
Photoautotrophic Production of Docosahexaenoic Acid- and Eicosapentaenoic Acid-Enriched Biomass by Co-Culturing Golden-Brown and Green Microalgae

Abstract
:1. Introduction
2. Materials and Methods
2.1. Microalgae Strains, Seed Culture Maintenance, and Media Composition
2.2. Pre-Cultures and Process Conditions
2.3. Flow Cytometry for Cell Ratio Measurements
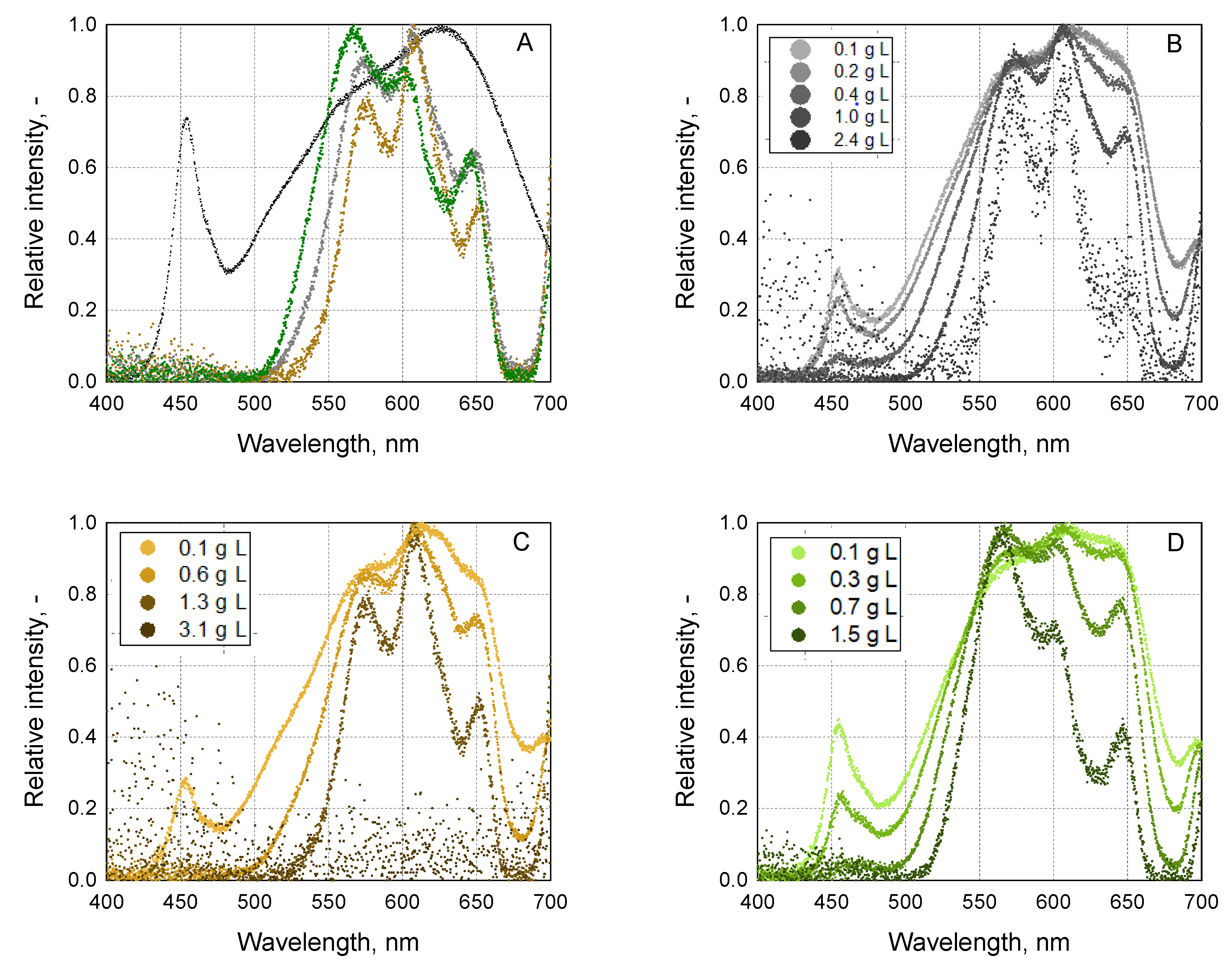
2.4. Spectra and Photon Flux Density Measurements
2.5. Determination of Urea and Nitrate
2.6. Determination of Optical Density, Cell Dry Weight, and Salinity
2.7. Lipid Transesterification and GC Analysis
3. Results and Discussion
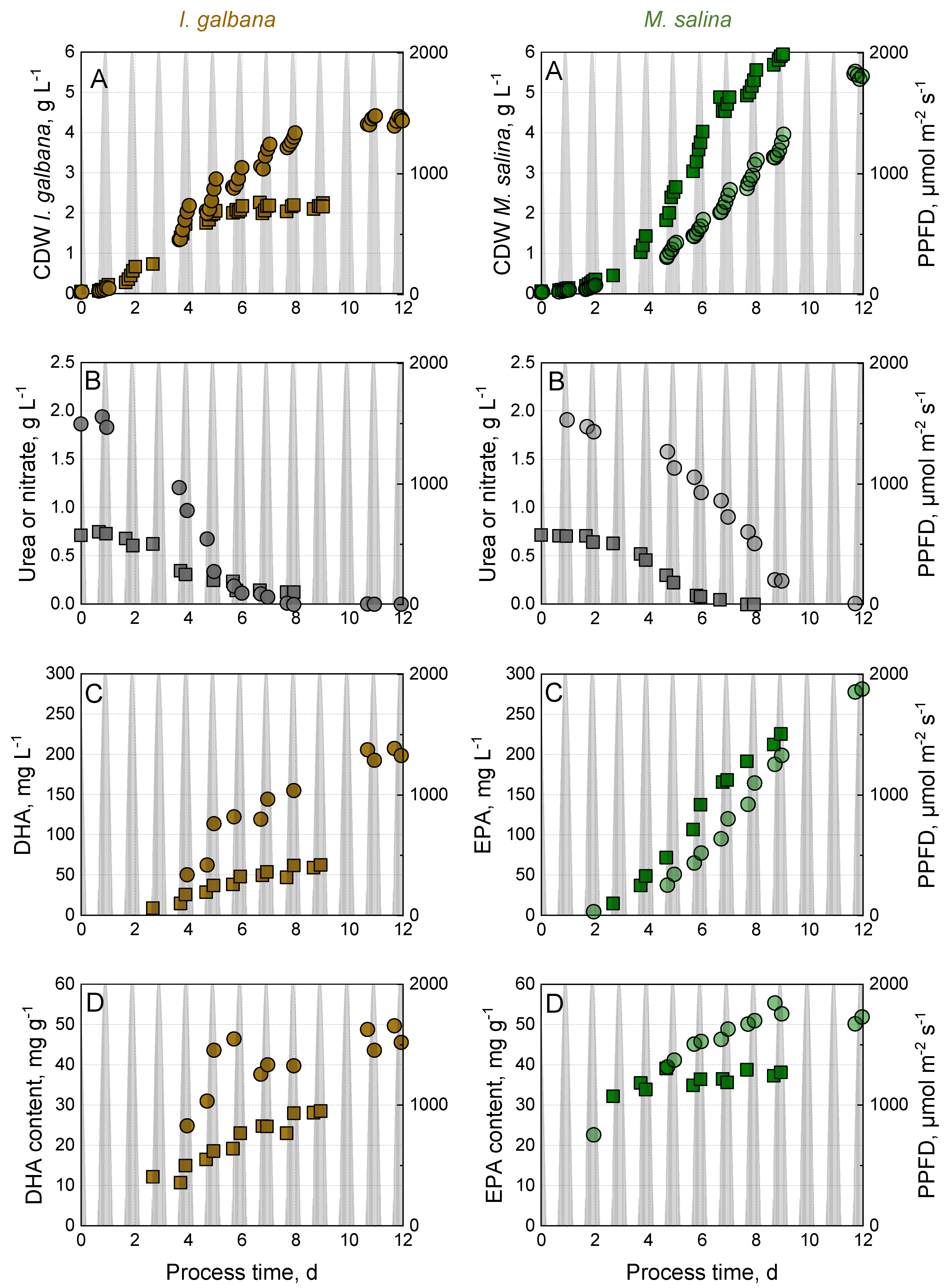
3.1. Effect of Nitrogen Source on Growth and Omega-3-Fatty Acid Production by I. galbana and M. salina in Monoculture
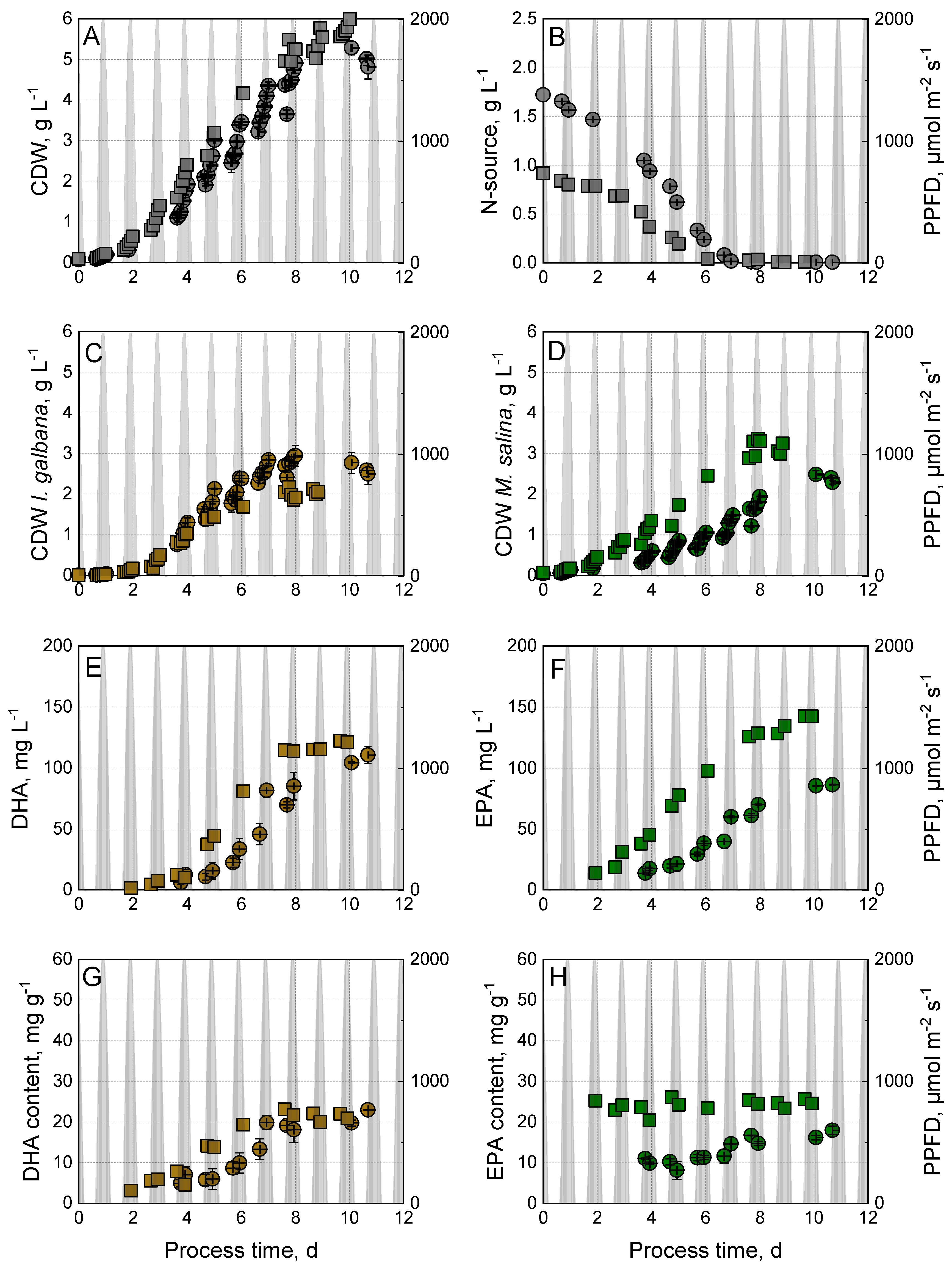
3.2. Effect of Nitrogen Source on Growth and Omega-3-Fatty Acid Production by I. galbana and M. salina in Co-Culture
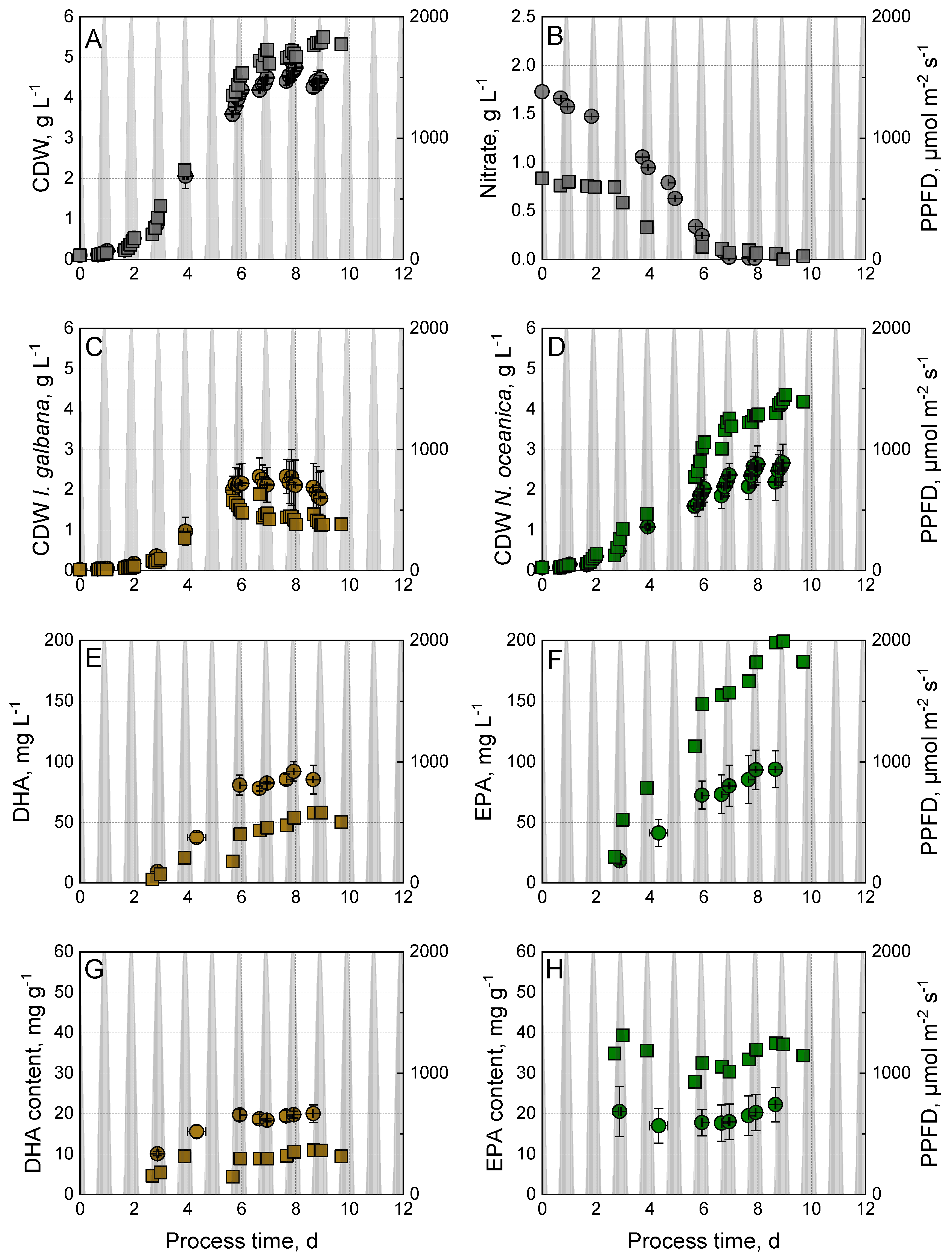
3.3. Phototrophic Co-Culture of I. galbana and N. oceanica Using Nitrate or Urea
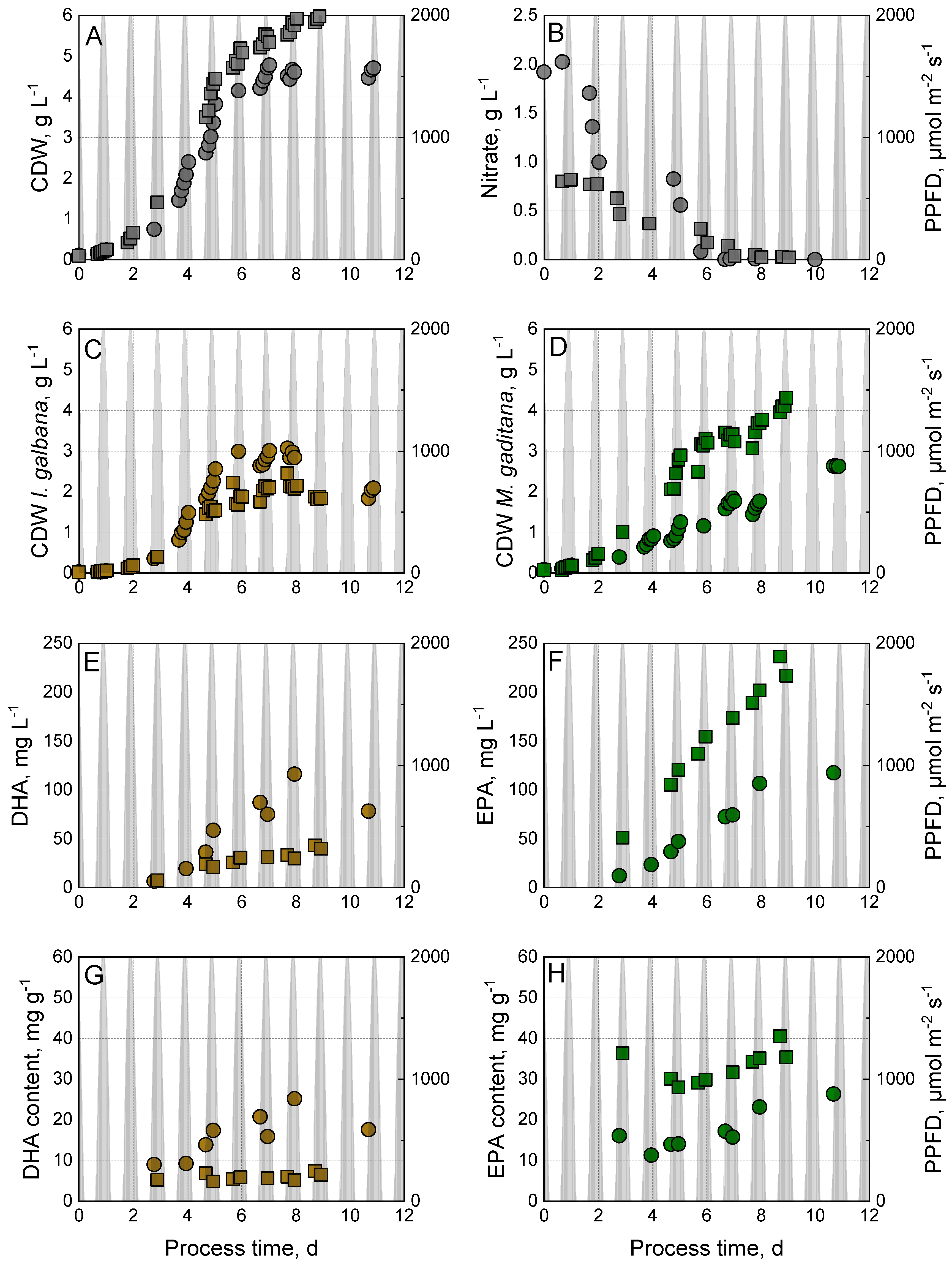
3.4. Phototrophic Co-Cultivation of I. galbana and M. gaditana Using Nitrate or Urea
3.5. Comparison of Phototrophic Co-Culture Processes with Golden-Brown Microalgae (DHA Producer) and Green Microalgae (EPA Producer)
3.6. Enhanced Light Utilization in Co-Cultures of Golden-Brown and Green Microalgae
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barta, D.G.; Coman, V.; Vodnar, D.C. Microalgae as sources of omega-3 polyunsaturated fatty acids: Biotechnological aspects. Algal Res. 2021, 58, 102410. [Google Scholar] [CrossRef]
- Khan, M.I.; Shin, J.H.; Kim, J.D. The promising future of microalgae: Current status, challenges, and optimization of a sustainable and renewable industry for biofuels, feed, and other products. Microb. Cell Factories 2018, 17, 36. [Google Scholar] [CrossRef] [PubMed]
- Tocher, D.R. Omega-3 long-chain polyunsaturated fatty acids and aquaculture in perspective. Aquaculture 2015, 449, 94–107. [Google Scholar] [CrossRef]
- Chua, E.T.; Schenk, P.M. A biorefinery for Nannochloropsis: Induction, harvesting, and extraction of EPA-rich oil and high-value protein. Bioresour. Technol. 2017, 244, 1416–1424. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, H.A.; Newton, R.; Auchterlonie, N.A.; Müller, D.B. Systems approach to quantify the global omega-3 fatty acid cycle. Nat. Food 2020, 1, 59–62. [Google Scholar] [CrossRef]
- Aranceta, J.; Perez-Rodrigo, C. Recommended dietary reference intakes, nutritional goals and dietary guidelines for fat and fatty acids: A systematic review. Br. J. Nutr. 2012, 107 (Suppl. S2), S8–S22. [Google Scholar] [CrossRef] [PubMed]
- Stark, K.D.; Van Elswyk, M.E.; Higgins, M.R.; Weatherford, C.A.; Salem, N., Jr. Global survey of the omega-3 fatty acids, docosahexaenoic acid and eicosapentaenoic acid in the blood stream of healthy adults. Prog. Lipid Res. 2016, 63, 132–152. [Google Scholar] [CrossRef] [PubMed]
- Tocher, D.R.; Betancor, M.B.; Sprague, M.; Olsen, R.E.; Napier, J.A. Omega-3 Long-Chain Polyunsaturated Fatty Acids, EPA and DHA: Bridging the Gap between Supply and Demand. Nutrients 2019, 11, 89. [Google Scholar] [CrossRef]
- Oliver, L.; Dietrich, T.; Marañón, I.; Villarán, M.C.; Barrio, R.J. Producing Omega-3 Polyunsaturated Fatty Acids: A Review of Sustainable Sources and Future Trends for the EPA and DHA Market. Resources 2020, 9, 148. [Google Scholar] [CrossRef]
- Draaisma, R.B.; Wijffels, R.H.; Slegers, P.M.; Brentner, L.B.; Roy, A.; Barbosa, M.J. Food commodities from microalgae. Curr. Opin. Biotechnol. 2013, 24, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Polishchuk, A.; Valev, D.; Tarvainen, M.; Mishra, S.; Kinnunen, V.; Antal, T.; Yang, B.; Rintala, J.; Tyystjärvi, E. Cultivation of Nannochloropsis for eicosapentaenoic acid production in wastewaters of pulp and paper industry. Bioresour. Technol. 2015, 193, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Borowitzka, M.A. Culturing microalgae in outdoor ponds. In Algal Culturing Techniques; Andersen, R.A., Ed.; Elsevier/Academic Press: Burlington, VT, USA, 2005; pp. 205–218. [Google Scholar]
- Aussant, J.; Guihéneuf, F.; Stengel, D.B. Impact of temperature on fatty acid composition and nutritional value in eight species of microalgae. Appl. Microbiol. Biotechnol. 2018, 102, 5279–5297. [Google Scholar] [CrossRef] [PubMed]
- Gu, W.; Kavanagh, J.M.; McClure, D.D. Photoautotrophic production of eicosapentaenoic acid. Crit. Rev. Biotechnol. 2021, 41, 731–748. [Google Scholar] [CrossRef] [PubMed]
- Guihéneuf, F.; Stengel, D.B. Interactive effects of light and temperature on pigments and n-3 LC-PUFA-enriched oil accumulation in batch-cultivated Pavlova lutheri using high-bicarbonate supply. Algal Res. 2017, 23, 113–125. [Google Scholar] [CrossRef]
- Nogueira, N.; Nascimento, F.; Cunha, C.; Cordeiro, N. Nannochloropsis gaditana grown outdoors in annular photobioreactors: Operation strategies. Algal Res. 2020, 48, 101913. [Google Scholar] [CrossRef]
- Sukenik, A.; Zmora, O.; Carmeli, Y. Biochemical quality of marine unicellular algae with special emphasis on lipid composition. II. Nannochloropsis sp. Aquaculture 1993, 117, 313–326. [Google Scholar] [CrossRef]
- Wikfors, G.H.; Patterson, G.W. Differences in strains of Isochrysis of importance to mariculture. Aquaculture 1994, 123, 127–135. [Google Scholar] [CrossRef]
- Sun, Z.; Wang, X.; Liu, J. Screening of Isochrysis strains for simultaneous production of docosahexaenoic acid and fucoxanthin. Algal Res. 2019, 41, 101545. [Google Scholar] [CrossRef]
- Gao, F.; Woolschot, S.; Cabanelas, I.T.D.; Wijffels, R.H.; Barbosa, M.J. Light spectra as triggers for sorting improved strains of Tisochrysis lutea. Bioresour. Technol. 2021, 321, 124434. [Google Scholar] [CrossRef]
- Kim, S.M.; Kang, S.-W.; Kwon, O.N.; Chung, D.; Pan, C.-H. Fucoxanthin as a major carotenoid in Isochrysis aff. galbana: Characterization of extraction for commercial application. J. Korean Soc. Appl. Biol. Chem. 2012, 55, 477–483. [Google Scholar] [CrossRef]
- Hulatt, C.J.; Wijffels, R.H.; Bolla, S.; Kiron, V. Production of Fatty Acids and Protein by Nannochloropsis in Flat-Plate Photobioreactors. PLoS ONE 2017, 12, e0170440. [Google Scholar] [CrossRef] [PubMed]
- Gu, W.; Kavanagh, J.M.; McClure, D.D. Towards a sustainable supply of omega-3 fatty acids: Screening microalgae for scalable production of eicosapentaenoic acid (EPA). Algal Res. 2022, 61, 102564. [Google Scholar] [CrossRef]
- Schadler, T.; Caballero Cerbon, D.; de Oliveira, L.; Garbe, D.; Bruck, T.; Weuster-Botz, D. Production of lipids with Microchloropsis salina in open thin-layer cascade photobioreactors. Bioresour. Technol. 2019, 289, 121682. [Google Scholar] [CrossRef] [PubMed]
- Rodolfi, L.; Chini Zittelli, G.; Bassi, N.; Padovani, G.; Biondi, N.; Bonini, G.; Tredici, M.R. Microalgae for oil: Strain selection, induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactor. Biotechnol. Bioeng. 2009, 102, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Russo, G.L.; Langellotti, A.L.; Martín-García, B.; Verardo, V.; Romano, R.; Sacchi, R.; Masi, P. New Biotechnological Production of EPA by Pythium irregulare Using Alternative Sustainable Media Obtained from Food Industry By-Products and Waste. Sustainability 2023, 15, 1147. [Google Scholar] [CrossRef]
- Lee Chang, K.J.; Nichols, C.M.; Blackburn, S.I.; Dunstan, G.A.; Koutoulis, A.; Nichols, P.D. Comparison of Thraustochytrids Aurantiochytrium sp., Schizochytrium sp., Thraustochytrium sp., and Ulkenia sp. for Production of Biodiesel, Long-Chain Omega-3 Oils, and Exopolysaccharide. Mar. Biotechnol. 2014, 16, 396–411. [Google Scholar] [CrossRef] [PubMed]
- Borowitzka, M.A. Commercial production of microalgae: Ponds, tanks, tubes and fermenters. J. Biotechnol. 1999, 70, 313–321. [Google Scholar] [CrossRef]
- Remize, M.; Brunel, Y.; Silva, J.L.; Berthon, J.Y.; Filaire, E. Microalgae n-3 PUFAs Production and Use in Food and Feed Industries. Mar. Drugs 2021, 19, 113. [Google Scholar] [CrossRef] [PubMed]
- Thurn, A.-L.; Stock, A.; Gerwald, S.; Weuster-Botz, D. Simultaneous photoautotrophic production of DHA and EPA by Tisochrysis lutea and Microchloropsis salina in co-culture. Bioresour. Bioprocess. 2022, 9, 130. [Google Scholar] [CrossRef]
- Harwood, J.L. Algae: Critical Sources of Very Long-Chain Polyunsaturated Fatty Acids. Biomolecules 2019, 9, 708. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-Y.; Nagarajan, D.; Cheah, W.Y. Eicosapentaenoic acid production from Nannochloropsis oceanica CY2 using deep sea water in outdoor plastic-bag type photobioreactors. Bioresour. Technol. 2018, 253, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Juneja, A.; Ceballos, R.M.; Murthy, G.S. Effects of Environmental Factors and Nutrient Availability on the Biochemical Composition of Algae for Biofuels Production: A Review. Energies 2013, 6, 4607–4638. [Google Scholar] [CrossRef]
- Wehr, J.D. Algae: Anatomy, Biochemistry, and Biotechnology by Barsanti, L. & Gualtieri, P. J. Phycol. 2006, 43, 412–414. [Google Scholar] [CrossRef]
- Converti, A.; Scapazzoni, S.; Lodi, A.; Carvalho, J.C.M. Ammonium and urea removal by Spirulina platensis. J. Ind. Microbiol. Biotechnol. 2006, 33, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Campos, H.; Boeing, W.J.; Dungan, B.N.; Schaub, T. Cultivating the marine microalga Nannochloropsis salina under various nitrogen sources: Effect on biovolume yields, lipid content and composition, and invasive organisms. Biomass Bioenergy 2014, 66, 301–307. [Google Scholar] [CrossRef]
- Ren, M.; Ogden, K. Cultivation of Nannochloropsis gaditana on mixtures of nitrogen sources. Environ. Prog. Sustain. Energy 2014, 33, 551–555. [Google Scholar] [CrossRef]
- Schädler, T.; Neumann-Cip, A.-C.; Wieland, K.; Glöckler, D.; Haisch, C.; Brück, T.; Weuster-Botz, D. High-Density Microalgae Cultivation in Open Thin-Layer Cascade Photobioreactors with Water Recycling. Appl. Sci. 2020, 10, 3883. [Google Scholar] [CrossRef]
- Havel, J.; Franco-Lara, E.; Weuster-Botz, D. A parallel bubble column system for the cultivation of phototrophic microorganisms. Biotechnol. Lett. 2008, 30, 1197–1200. [Google Scholar] [CrossRef] [PubMed]
- Pfaffinger, C.E.; Severin, T.S.; Apel, A.C.; Göbel, J.; Sauter, J.; Weuster-Botz, D. Light-dependent growth kinetics enable scale-up of well-mixed phototrophic bioprocesses in different types of photobioreactors. J. Biotechnol. 2019, 297, 41–48. [Google Scholar] [CrossRef]
- Ge, F.; Song, K.; Yang, Z.; Li, J.; Yan, F.; Zhang, M.; Huang, B.; Fu, G.; Zheng, H. Enhancing Docosahexaenoic Acid Production of Isochrysis galbana from Starch-Rich Food Processing Byproducts. Fermentation 2023, 9, 158. [Google Scholar] [CrossRef]
- Zheng, H.; Ge, F.; Song, K.; Yang, Z.; Li, J.; Yan, F.; Wu, X.; Zhang, Q.; Liu, Y.; Ruan, R. Docosahexaenoic acid production of the marine microalga Isochrysis galbana cultivated on renewable substrates from food processing waste under CO2 enrichment. Sci. Total Environ. 2022, 848, 157654. [Google Scholar] [CrossRef]
- Liu, J.; Sommerfeld, M.; Hu, Q. Screening and characterization of Isochrysis strains and optimization of culture conditions for docosahexaenoic acid production. Appl. Microbiol. Biotechnol. 2013, 97, 4785–4798. [Google Scholar] [CrossRef]
- Fidalgo, J.P.; Cid, A.; Torres, E.; Sukenik, A.; Herrero, C. Effects of nitrogen source and growth phase on proximate biochemical composition, lipid classes and fatty acid profile of the marine microalga Isochrysis galbana. Aquaculture 1998, 166, 105–116. [Google Scholar] [CrossRef]
- Sassano, C.E.N.; Carvalho, J.C.M.; Gioielli, L.A.; Sato, S.; Torre, P.; Converti, A. Kinetics and bioenergetics of Spirulina platensis cultivation by fed-batch addition of urea as nitrogen source. Appl. Biochem. Biotechnol. 2004, 112, 143–150. [Google Scholar] [CrossRef]
- Roopnarain, A.; Sym, S.; Gray, V.M. Effect of nitrogenous resource on growth, biochemical composition and ultrastructure of Isochrysis galbana (Isochrysidales, Haptophyta). Phycol. Res. 2015, 63, 43–50. [Google Scholar] [CrossRef]
- Abeliovich, A.; Azov, Y. Toxicity of ammonia to algae in sewage oxidation ponds. Appl. Environ. Microbiol 1976, 31, 801–806. [Google Scholar] [CrossRef]
- Belkin, S.; Boussiba, S. High internal pH conveys ammonia resistance in Spirulina platensis. Bioresour. Technol. 1991, 38, 167–169. [Google Scholar] [CrossRef]
- Chen, S.Y.; Pan, L.I.; Hong, M.I.; Lee, A.N. The effects of temperature on the growth of and ammonia uptake by marine microalgae. Bot. Stud. 2012, 53, 125–133. [Google Scholar]
- Abeliovich, A. Water Pollution and Bioremediation by Microalgae: Water Purification: Algae in Wastewater Oxidation Ponds. In Handbook of Microalgal Culture; Wiley: Hoboken, NJ, USA, 2003; pp. 430–438. [Google Scholar] [CrossRef]
- Meng, Y.; Jiang, J.; Wang, H.; Cao, X.; Xue, S.; Yang, Q.; Wang, W. The characteristics of TAG and EPA accumulation in Nannochloropsis oceanica IMET1 under different nitrogen supply regimes. Bioresour. Technol. 2015, 179, 483–489. [Google Scholar] [CrossRef]
- Gao, F.; Teles, I.; Wijffels, R.H.; Barbosa, M.J. Process optimization of fucoxanthin production with Tisochrysis lutea. Bioresour. Technol. 2020, 315, 123894. [Google Scholar] [CrossRef]
- Pereira, H.; Sá, M.; Maia, I.; Rodrigues, A.; Teles, I.; Wijffels, R.H.; Navalho, J.; Barbosa, M. Fucoxanthin production from Tisochrysis lutea and Phaeodactylum tricornutum at industrial scale. Algal Res. 2021, 56, 102322. [Google Scholar] [CrossRef]
- Bendif, E.M.; Probert, I.; Schroeder, D.C.; de Vargas, C. On the description of Tisochrysis lutea gen. nov. sp. nov. and Isochrysis nuda sp. nov. in the Isochrysidales, and the transfer of Dicrateria to the Prymnesiales (Haptophyta). J. Appl. Phycol. 2013, 25, 1763–1776. [Google Scholar] [CrossRef]
- Wang, W.; Yu, L.-J.; Xu, C.; Tomizaki, T.; Zhao, S.; Umena, Y.; Chen, X.; Qin, X.; Xin, Y.; Suga, M.; et al. Structural basis for blue-green light harvesting and energy dissipation in diatoms. Science 2019, 363, eaav0365. [Google Scholar] [CrossRef] [PubMed]
- Premvardhan, L.; Bordes, L.; Beer, A.; Büchel, C.; Robert, B. Carotenoid Structures and Environments in Trimeric and Oligomeric Fucoxanthin Chlorophyll a/c2 Proteins from Resonance Raman Spectroscopy. J. Phys. Chem. B 2009, 113, 12565–12574. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, J. Analysis of light absorption and photosynthetic activity by Isochrysis galbana under different light qualities. Aquac. Res. 2020, 51, 2893–2902. [Google Scholar] [CrossRef]
- Keeling, P.J. The number, speed, and impact of plastid endosymbioses in eukaryotic evolution. Annu. Rev. Plant Biol. 2013, 64, 583–607. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Co-Cultivations | Nitrogen Source | Inoculation Ratio |
|---|---|---|
| I. galbana and M. salina | Nitrate | 1:3 |
| Urea | 1:3 | |
| I. galbana and N. oceanica | Nitrate | 1:3 |
| Urea | 1:3 | |
| I. galbana and M. gaditana | Nitrate | 1:3 |
| Urea | 1:3 |
| t = 8 d | I. galbana | M. salina | Co-Culture | Co-Culture |
|---|---|---|---|---|
| CDW, g L−1 | 4.0 | 3.2 | 4.8 ± 0.2 | +33 ± 4% |
| DHA, mg L−1 | 77.5 | - | 85.5 ± 11 | +10 ± 13% |
| DHA, mgDHA gCDW−1 | 20 | - | 18 ± 3 | - |
| EPA, mg L−1 | - | 82.5 | 70.2 ± 1 | −15 ± 1% |
| EPA, mgEPA gCDW−1 | - | 20.5 | 15 ± 0 | −27 ± 0% |
| t = 8 d | I. galbana | M. salina | Co-Culture | Co-Culture |
|---|---|---|---|---|
| CDW, g L−1 | 2.2 | 5.3 | 5.0 | +33% |
| DHA, mg L−1 | 31 | - | 114 | +3.7-fold |
| DHA, mgDHA gCDW−1 | 14 | - | 22 | +57% |
| EPA, mg L−1 | - | 96.0 | 129 | +34% |
| EPA, mgEPA gCDW−1 | - | 19.5 | 24 | +23% |
| t = 8 d | I. galbana | N. oceanica | Co-Culture | Co-Culture |
|---|---|---|---|---|
| CDW, g L−1 | 4.0 | 5.0 | 4.7 ± 0.2 | +18 ± 4% |
| DHA, mg L−1 | 77.5 | - | 92 ± 8 | +19 ± 9% |
| DHA, mgDHA gCDW−1 | 20 | - | 20 ± 1 | - |
| EPA, mg L−1 | - | 128 | 93 ± 16 | −27 ± 17% |
| EPA, mgEPA gCDW−1 | - | 25.5 | 20 ± 4 | −22 ± 20% |
| t = 8 d | I. galbana | M. gaditana | Co-Culture | Co-Culture |
|---|---|---|---|---|
| CDW, g L−1 | 4.0 | 4.6 | 4.6 | +15% |
| DHA, mg L−1 | 77.5 | - | 116 | +49% |
| DHA, mgDHA gCDW−1 | 20 | - | 25 | +25% |
| EPA, mg L−1 | - | 122 | 107 | −12% |
| EPA, mgEPA gCDW−1 | - | 25 | 23 | −8% |
| Nitrogen Source | CDW, g L−1 | CDW- Golden-Brown Microalgae, g L−1 | CDW- Green MICROALGAE, g L−1 | DHA, mgDHA gCDW−1 | EPA, mgEPA gCDW−1 | |
|---|---|---|---|---|---|---|
| I. galbana and M. salina | Nitrate | 4.8 ± 0.2 | 2.9 ± 0.2 | 1.8 ± 0 | 18 ± 3 | 15 ± 0 |
| Urea | 5.3 | 1.9 | 3.4 | 22 | 24 | |
| I. galbana and N. oceanica | Nitrate | 4.7 ± 0.2 | 2.1 ± 0.6 | 2.6 ± 0.5 | 20 ± 1 | 20 ± 4 |
| Urea | 5.1 | 1.3 | 3.8 | 11 | 36 | |
| I. galbana and M. gaditana | Nitrate | 4.6 | 2.8 | 1.8 | 25 | 23 |
| Urea | 5.8 | 2.1 | 3.7 | 5 | 35 | |
| T. lutea and M. salina [29] | Nitrate | 4.2 ± 0.3 | 2.1 ± 0.1 | 2.0 ± 0.3 | 26 ± 2 | 23 ± 4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thurn, A.-L.; Schobel, J.; Weuster-Botz, D. Photoautotrophic Production of Docosahexaenoic Acid- and Eicosapentaenoic Acid-Enriched Biomass by Co-Culturing Golden-Brown and Green Microalgae. Fermentation 2024, 10, 220. https://doi.org/10.3390/fermentation10040220
Thurn A-L, Schobel J, Weuster-Botz D. Photoautotrophic Production of Docosahexaenoic Acid- and Eicosapentaenoic Acid-Enriched Biomass by Co-Culturing Golden-Brown and Green Microalgae. Fermentation. 2024; 10(4):220. https://doi.org/10.3390/fermentation10040220
Chicago/Turabian StyleThurn, Anna-Lena, Josef Schobel, and Dirk Weuster-Botz. 2024. "Photoautotrophic Production of Docosahexaenoic Acid- and Eicosapentaenoic Acid-Enriched Biomass by Co-Culturing Golden-Brown and Green Microalgae" Fermentation 10, no. 4: 220. https://doi.org/10.3390/fermentation10040220