
The Biological Role of the S-Layer Produced by Lactobacillus helveticus 34.9 in Cell Protection and Its Probiotic Properties

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. S-Layer Extraction
2.3. Co-Aggregation
2.4. Microbial Adhesion to Solvents (MATS)
2.5. Influence of S-Layer on Bacterial Resistance during Freezing and Freeze-Drying
2.6. Influence of the S-Layer on the Survival of the Producing Cells under Conditions Simulating the Passage through the Gastrointestinal Tract (GIT)
2.7. Scanning Electron Microscopy
2.8. HT-29 Cell Culture
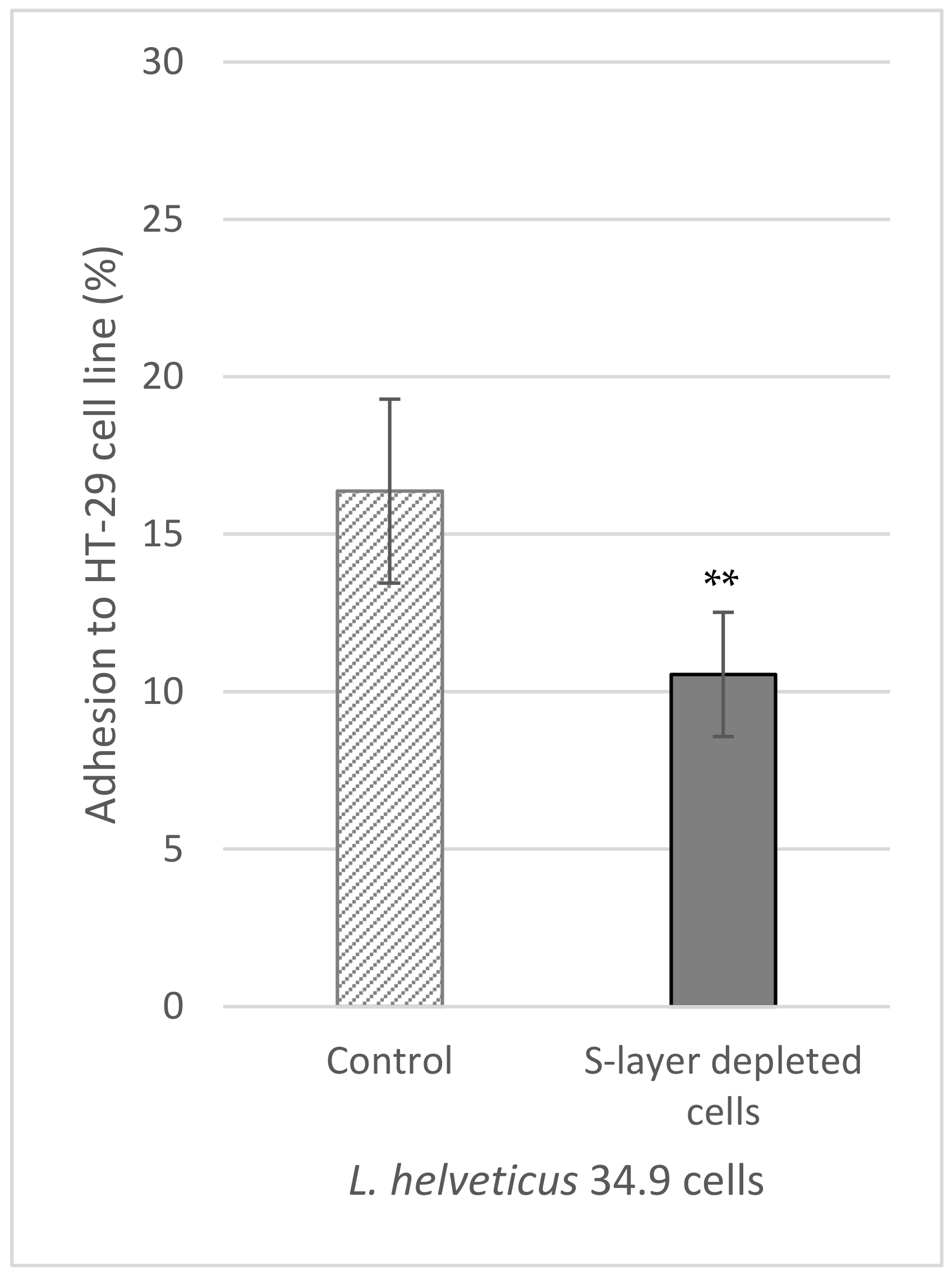
2.9. In Vitro Adhesion of L. helveticus 34.9 to HT-29 Cells
2.10. Statistical Analysis
3. Results
3.1. S-Layer Extraction
3.2. Co-Aggregation
3.3. Bacterial Adhesion to Solvents
3.4. Influence of the S-Layer on Bacterial Resistance during Freezing and Freeze-Drying
3.5. The Importance of the S-Layer for the GIT Survival
3.6. Scanning Electron Microscopy
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martin, R.; Langella, P. Emerging Health Concepts in the Probiotics Field: Streamlining the Definitions. Front. Microbiol. 2019, 10, 1047. [Google Scholar] [CrossRef] [PubMed]
- FAO/WHO. Probiotic in Foods: Health and Nutritional Properties and Guidelines for Evaluation; FAO Food and Nutrition; FAO: Rome, Italy, 2006; p. 85. [Google Scholar]
- Ohashi, Y.; Ushida, K. Health-beneficial effects of probiotics: Its mode of action. Anim. Sci. J. Nihon Chikusan Gakkaiho 2009, 80, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, J.; Yang, K.; Liu, M.; Zhang, J.; Xinyuan, W.; Fan, M. Screening for potential probiotic from spontaneously fermented non-dairy foods based on in vitro probiotic and safety properties. Ann. Microbiol. 2018, 68, 803–813. [Google Scholar] [CrossRef]
- Bodke, H.; Jogdand, S. Role of Probiotics in Human Health. Cureus 2022, 14, e31313. [Google Scholar] [CrossRef] [PubMed]
- Hynönen, U.; Palva, A. Lactobacillus surface layer proteins: Structure, function and applications. Appl. Microbiol. Biotechnol. 2013, 97, 5225–5243. [Google Scholar] [CrossRef] [PubMed]
- Palomino, M.M.; Allievi, M.C.; Gordillo, T.B.; Bockor, S.S.; Fina Martin, J.; Ruzal, S.M. Surface layer proteins in species of the family Lactobacillaceae. Microb. Biotechnol. 2023, 16, 1232–1249. [Google Scholar] [CrossRef] [PubMed]
- Mobarak Qamsari, E.; Kasra Kermanshahi, R.; Erfan, M.; Ghadam, P.; Sardari, S.; Eslami, N. Characteristics of surface layer proteins from two new and native strains of Lactobacillus brevis. Int. J. Biol. Macromol. 2017, 95, 1004–1010. [Google Scholar] [CrossRef] [PubMed]
- Grosu-Tudor, S.S.; Angelescu, I.R.; Brinzan, A.; Zamfir, M. Characterization of S-layer proteins produced by lactobacilli isolated from Romanian artisan fermented products. J. Appl. Microbiol. 2023, 134, lxac063. [Google Scholar] [CrossRef]
- Deepika, G.; Charalampopoulos, D. Chapter 4—Surface and Adhesion Properties of Lactobacilli. In Advances in Applied Microbiology; Academic Press: Cambridge, MA, USA, 2010; pp. 127–152. [Google Scholar]
- Martínez, M.G.; Prado Acosta, M.; Candurra, N.A.; Ruzal, S.M. S-layer proteins of Lactobacillus acidophilus inhibits JUNV infection. Biochem. Biophys. Res. Commun. 2012, 422, 590–595. [Google Scholar] [CrossRef]
- Sleytr, U.B.; Schuster, B.; Egelseer, E.-M.; Pum, D. S-layers: Principles and applications. FEMS Microbiol. Rev. 2014, 38, 823–864. [Google Scholar] [CrossRef]
- Gerbino, E.; Carasi, P.; Mobili, P.; Serradell, M.A.; Gomez-Zavaglia, A. Role of S-layer proteins in bacteria. World J. Microbiol. Biotechnol. 2015, 31, 1877–1887. [Google Scholar] [CrossRef] [PubMed]
- Alp, D.; Kuleaşan, H.; Korkut Altıntaş, A. The importance of the S-layer on the adhesion and aggregation ability of Lactic acid bacteria. Mol. Biol. Rep. 2020, 47, 3449–3457. [Google Scholar] [CrossRef] [PubMed]
- Angelescu, I.R.; Grosu-Tudor, S.S.; Cojoc, L.R.; Maria, G.M.; Chiritoiu, G.N.; Munteanu, C.V.A.; Zamfir, M. Isolation, characterization, and mode of action of a class III bacteriocin produced by Lactobacillus helveticus 34.9. World J. Microbiol. Biotechnol. 2022, 38, 220. [Google Scholar] [CrossRef] [PubMed]
- Lortal, S.; Van Heijenoort, J.; Gruber, K.; Sleytr, U.B. S-layer of Lactobacillus helveticus ATCC 12046: Isolation, chemical characterization and re-formation after extraction with lithium chloride. J. Gen. Microbiol. 1992, 138, 611–618. [Google Scholar] [CrossRef]
- Han, Q.; Kong, B.; Chen, Q.; Sun, F.; Zhang, H. In vitro comparison of probiotic properties of lactic acid bacteria isolated from Harbin dry sausages and selected probiotics. J. Funct. Foods 2017, 32, 391–400. [Google Scholar] [CrossRef]
- Zamfir, M.; Stefan, I.R.; Stancu, M.M.; Grosu-Tudor, S.S. Production, mode of action and sequencing of the corresponding gene of a bacteriocin produced by Lactococcus lactis 19.3. Int. J. Food Sci. Technol. 2016, 51, 2164–2170. [Google Scholar] [CrossRef]
- Angelescu, I.-R.; Zamfir, M.; Stancu, M.-M.; Grosu-Tudor, S.-S. Identification and probiotic properties of lactobacilli isolated from two different fermented beverages. Ann. Microbiol. 2019, 69, 1557–1565. [Google Scholar] [CrossRef]
- Zamfir, M.; Callewaert, R.; Cornea, P.C.; Savu, L.; Vatafu, I.; De Vuyst, L. Purification and characterization of a bacteriocin produced by Lactobacillus acidophilus IBB 801. J. Appl. Microbiol. 1999, 87, 923–931. [Google Scholar] [CrossRef]
- Grosu-Tudor, S.S.; Brown, L.; Hebert, E.M.; Brezeanu, A.; Brinzan, A.; Fadda, S.; Mozzi, F.; Zamfir, M. S-layer production by Lactobacillus acidophilus IBB 801 under environmental stress conditions. Appl. Microbiol. Biotechnol. 2016, 100, 4573–4583. [Google Scholar] [CrossRef]
- Hernández-Alcántara, A.; Wacher, C.; Llamas-Arriba, M.; López, P.; Pérez-Chabela, M. Probiotic properties and stress response of thermotolerant lactic acid bacteria isolated from cooked meat products. LWT 2018, 91, 249–257. [Google Scholar] [CrossRef]
- Grosu-Tudor, S.S.; Ștefan, I.R.; Zamfir, M. Growth/survival of some functional lactic acid bacteria under different stress conditions. AgroLife Sci. J. 2016, 5, 71–78. [Google Scholar]
- Beldarrain-Iznaga, T.; Villalobos-Carvajal, R.; Sevillano-Armesto, E.; Leiva-Vega, J. Functional properties of Lactobacillus casei C24 improved by microencapsulation using multilayer double emulsion. Food Res. Int. 2021, 141, 110136. [Google Scholar] [CrossRef] [PubMed]
- Kos, B.; Šušković, J.; Vuković, S.; Šimpraga, M.; Frece, J.; Matošić, S. Adhesion and aggregation ability of probiotic strain Lactobacillus acidophilus M92. J. App. Microbiol. 2003, 94, 981–987. [Google Scholar] [CrossRef]
- Abdulla, A.A.; Abed, T.A.; Saeed, A.M. Adhesion, autoaggregation and hydrophobicity of six Lactobacillus strains. Br. Microbiol. Res. J. 2014, 4, 381–391. [Google Scholar] [CrossRef]
- Uroić, K.; Novak, J.; Hynönen, U.; Pietilä, T.E.; Leboš Pavunc, A.; Kant, R.; Kos, B.; Palva, A.M.; Suskovic, J. The role of S-layer in adhesive and immunomodulating properties of probiotic starter culture Lactobacillus brevis D6 isolated from artisanal smoked fresh cheese. LWT Food Sci. Technol. 2016, 69, 623–632. [Google Scholar] [CrossRef]
- Banić, M.; Uroić, K.; Pavunc, A.; Novak, J.; Zorić, K.; Durgo, K.; Petković, H.; Jamnik, P.; Kazazić, S.; Kazazić, S.; et al. Characterization of S-layer proteins of potential probiotic starter culture Lactobacillus brevis SF9B isolated from sauerkraut. LWT 2018, 93, 257–267. [Google Scholar] [CrossRef]
- Gharbi, Y.; Fhoula, I.; Ruas-Madiedo, P.; Afef, N.; Boudabous, A.; Gueimonde, M.; Ouzari, H.-I. In-vitro characterization of potentially probiotic Lactobacillus strains isolated from human microbiota: Interaction with pathogenic bacteria and the enteric cell line HT29. Ann. Microbiol. 2019, 69, 61–72. [Google Scholar] [CrossRef]
- Hojjati, M.; Behabahani, B.A.; Falah, F. Aggregation, adherence, anti-adhesion and antagonistic activity properties relating to surface charge of probiotic Lactobacillus brevis gp104 against Staphylococcus aureus. Microb. Pathog. 2020, 147, 104420. [Google Scholar] [CrossRef]
- Vanitha, P.R.; Somashekaraiah, R.; Divyashree, S.; Pan, I.; Sreenivasa, M.Y. Antifungal activity of probiotic strain Lactiplantibacillus plantarum MYSN7 against Trichophyton tonsurans. Front. Microbiol. 2023, 14, 1192449. [Google Scholar] [CrossRef]
- Yadav, R.; Puniya, A.K.; Shukla, P. Probiotic Properties of Lactobacillus plantarum RYPR1 from an Indigenous Fermented Beverage Raabadi. Front. Microbiol. 2016, 7, 1683. [Google Scholar] [CrossRef]
- Bouchard, D.S.; Seridan, B.; Saraoui, T.; Rault, L.; Germon, P.; Gonzalez-Moreno, C.; Nader-Macias, F.M.; Baud, D.; François, P.; Chuat, V.; et al. Lactic Acid Bacteria Isolated from Bovine Mammary Microbiota: Potential Allies against Bovine Mastitis. PLoS ONE 2015, 10, e0144831. [Google Scholar] [CrossRef] [PubMed]
- Sandes, S.; Alvim, L.; Silva, B.; Acurcio, L.; Santos, C.; Campos, M.; Santos, C.; Nicoli, J.; Neumann, E.; Nunes, Á. Selection of new lactic acid bacteria strains bearing probiotic features from mucosal microbiota of healthy calves: Looking for immunobiotics through in vitro and in vivo approaches for immunoprophylaxis applications. Microbiol. Res. 2017, 200, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wasko, A.; Polak-Berecka, M.; Paduch, R.; Jozwiak, K. The effect of moonlighting proteins on the adhesion and aggregation ability of Lactobacillus helveticus. Anaerobe 2014, 30, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Johnson-Henry, K.C.; Hagen, K.E.; Gordonpour, M.; Tompkins, T.A.; Sherman, P.M. Surface-layer protein extracts from Lactobacillus helveticus inhibit enterohaemorrhagic Escherichia coli O157:H7 adhesion to epithelial cells. Cell. Microbiol. 2007, 9, 356–367. [Google Scholar] [CrossRef] [PubMed]
- Boonaert, C.J.P.; Rouxhet, P.G. Surface of Lactic Acid Bacteria: Relationships between Chemical Composition and Physicochemical Properties. Appl. Environ. Microbiol. 2000, 66, 2548–2554. [Google Scholar] [CrossRef] [PubMed]
- Rong, J.; Zheng, H.; Liu, M.; Hu, X.; Wang, T.; Zhang, X.; Jin, F.; Wang, L. Probiotic and anti-inflammatory attributes of an isolate Lactobacillus helveticus NS8 from Mongolian fermented koumiss. BMC Microbiol. 2015, 15, 196. [Google Scholar] [CrossRef] [PubMed]
- Samet, K.; Icen, T.I. Screening of Potential Probiotic Properties of Lactobacillus and Lactococcus strains. Res. J. Biotechnol. 2022, 17, 119–126. [Google Scholar]
- Schar-Zammaretti, P.; Dillmann, M.L.; D’Amico, N.; Affolter, M.; Ubbink, J. Influence of fermentation medium composition on physicochemical surface properties of Lactobacillus acidophilus. Appl. Environ. Microbiol. 2005, 71, 8165–8173. [Google Scholar] [CrossRef]
- Van der Mei, H.C.; van de Belt-Gritter, B.; Pouwels, P.H.; Martinez, B.; Busscher, H.J. Cell surface hydrophobicity is conveyed by S-layer proteins—A study in recombinant lactobacilli. Colloids Surf. B Biointerfaces 2003, 28, 127–134. [Google Scholar] [CrossRef]
- Xu, H.; Jeong, H.S.; Lee, H.-Y.; Ahn, J. Assessment of cell surface properties and adhesion potential of selected probiotic strains. Lett. Appl. Microbiol. 2009, 49, 434–442. [Google Scholar] [CrossRef]
- Corcoran, B.M.; Ross, R.P.; Fitzgerald, G.F.; Dockery, P.; Stanton, C. Enhanced survival of GroESL-overproducing Lactobacillus paracasei NFBC 338 under stressful conditions induced by drying. Appl. Environ. Microbiol. 2006, 72, 5104–5107. [Google Scholar] [CrossRef] [PubMed]
- Davoodabadi, A.; Soltan Dallal, M.M.; Rahimi Foroushani, A.; Douraghi, M.; Sharifi Yazdi, M.K.; Amin Harati, F. Antibacterial activity of Lactobacillus spp. isolated from the feces of healthy infants against enteropathogenic bacteria. Anaerobe 2015, 34, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Jabbari, V.; Rezaei Mokarram, R.; Khiabani, M.; Askari, F.; Ahmadi, E.; Hassanzadeh, A.; Aghazadeh, S.B.; Asgharzadeh, M.; Kafilet, H.S. Molecular Identification of Lactobacillus acidophilus as a probiotic potential from traditional doogh samples and evaluation of their antimicrobial activity against some pathogenic bacteria. Biomed. Res. 2017, 28, 1458–1463. [Google Scholar]
- Johnson, B.; Selle, K.; O’Flaherty, S.; Goh, Y.J.; Klaenhammer, T. Identification of extracellular surface-layer associated proteins in Lactobacillus acidophilus NCFM. Microbiology 2013, 159, 2269–2282. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, L.; Margolles, A.; Sanchez, B. Bile resistance mechanisms in Lactobacillus and Bifidobacterium. Front. Microbiol. 2013, 4, 396. [Google Scholar] [CrossRef] [PubMed]
- Duary, R.K.; Batish, V.K.; Grover, S. Relative gene expression of bile salt hydrolase and surface proteins in two putative indigenous Lactobacillus plantarum strains under in vitro gut conditions. Mol. Biol. Rep. 2012, 39, 2541–2552. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Wei, H.; Yuan, J.; Li, Q.; Li, Y.; Li, N.; Li, J. Identification of a Surface Protein from Lactobacillus reuteri JCM1081 That Adheres to Porcine Gastric Mucin and Human Enterocyte-Like HT-29 Cells. Curr. Microbiol. 2008, 57, 33–38. [Google Scholar] [CrossRef]
- Jakava-Viljanen, M.; Palva, A. Isolation of surface (S) layer protein carrying Lactobacillus species from porcine intestine and faeces and characterization of their adhesion properties to different host tissues. Vet. Microbiol. 2007, 124, 264–273. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Survival (%) | ||||
|---|---|---|---|---|
| Storage at −20 °C | ||||
| 3 Days | 7 Days | 14 Days | Freeze-Drying | |
| Control cells | 99.8 ± 1.12 | 99.2 ± 0.27 | 99.0 ± 2.37 | 68.6 ± 0.04 |
| S-layer depleted cells | 99.4 ± 1.28 | 98.2 ± 1.35 | 97.9 ± 3.76 | 41.8 ± 1.75 |
| Bacterial Survival (%) | ||
|---|---|---|
| pH 3 + Pepsin | pH 8 + Pancreatin + Bile Salts | |
| Control cells | 70.3 ± 0.08 | 66.2 ± 0.29 |
| S-layer depleted cells | 55.6 ± 1.07 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Angelescu, I.-R.; Zamfir, M.; Ionetic, E.-C.; Grosu-Tudor, S.-S. The Biological Role of the S-Layer Produced by Lactobacillus helveticus 34.9 in Cell Protection and Its Probiotic Properties. Fermentation 2024, 10, 150. https://doi.org/10.3390/fermentation10030150
Angelescu I-R, Zamfir M, Ionetic E-C, Grosu-Tudor S-S. The Biological Role of the S-Layer Produced by Lactobacillus helveticus 34.9 in Cell Protection and Its Probiotic Properties. Fermentation. 2024; 10(3):150. https://doi.org/10.3390/fermentation10030150
Chicago/Turabian StyleAngelescu, Iulia-Roxana, Medana Zamfir, Emanuela-Cătălina Ionetic, and Silvia-Simona Grosu-Tudor. 2024. "The Biological Role of the S-Layer Produced by Lactobacillus helveticus 34.9 in Cell Protection and Its Probiotic Properties" Fermentation 10, no. 3: 150. https://doi.org/10.3390/fermentation10030150