
Signaling Pathways Involved in Myocardial Ischemia–Reperfusion Injury and Cardioprotection: A Systematic Review of Transcriptomic Studies in Sus scrofa

 ,
,
Abstract
:
1. Introduction
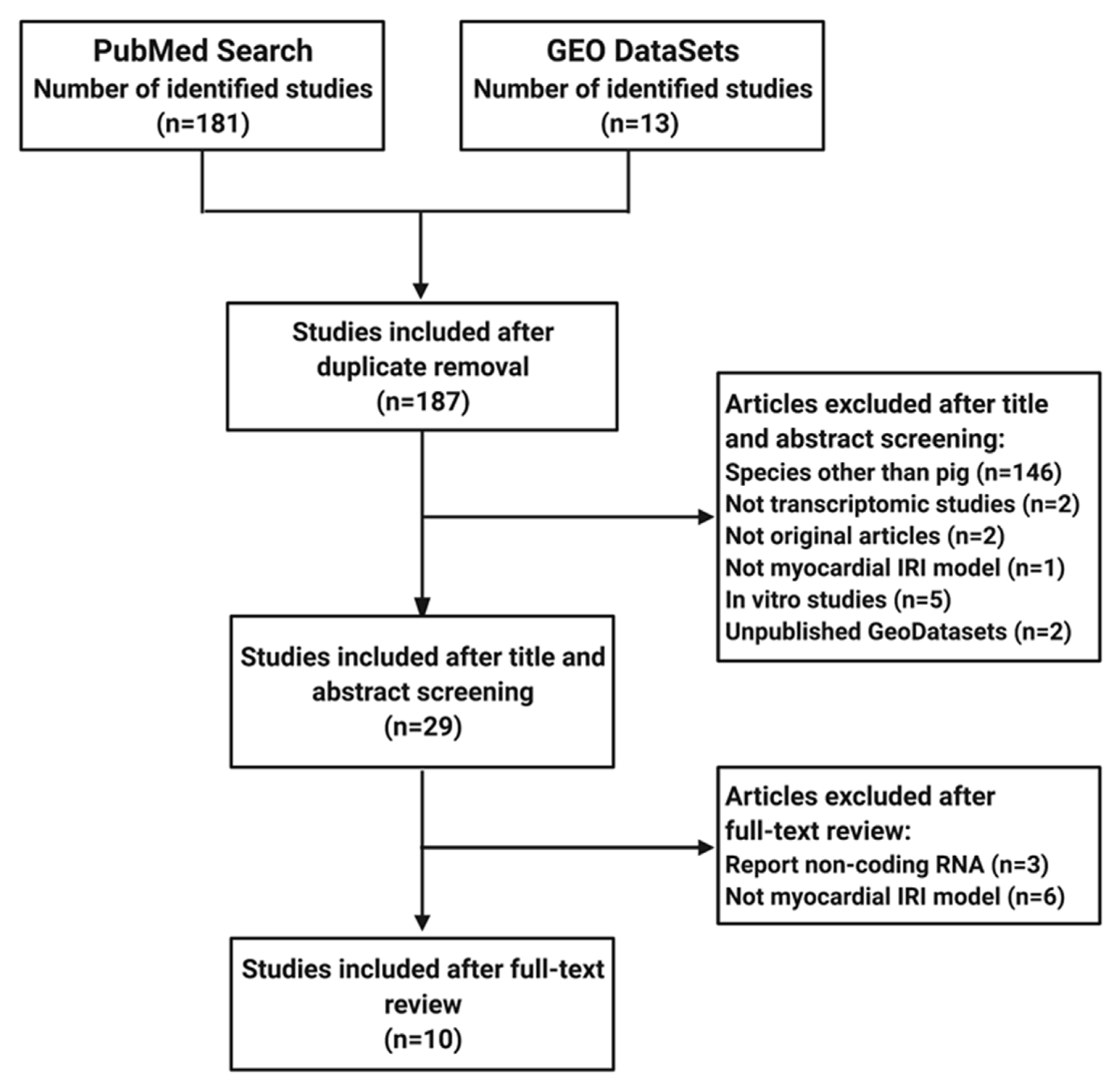
2. Materials and Methods
2.1. Elegibility Criteria
2.2. Search Strategy
2.3. Data Collection
2.4. Enrichment Analysis
2.5. Venn Diagrams
2.6. Reactome Pathway Analysis
3. Results
3.1. Time Course of Transcriptional Events in Myocardial IRI
3.1.1. Signaling Pathways Upregulated in the IC and the BZ
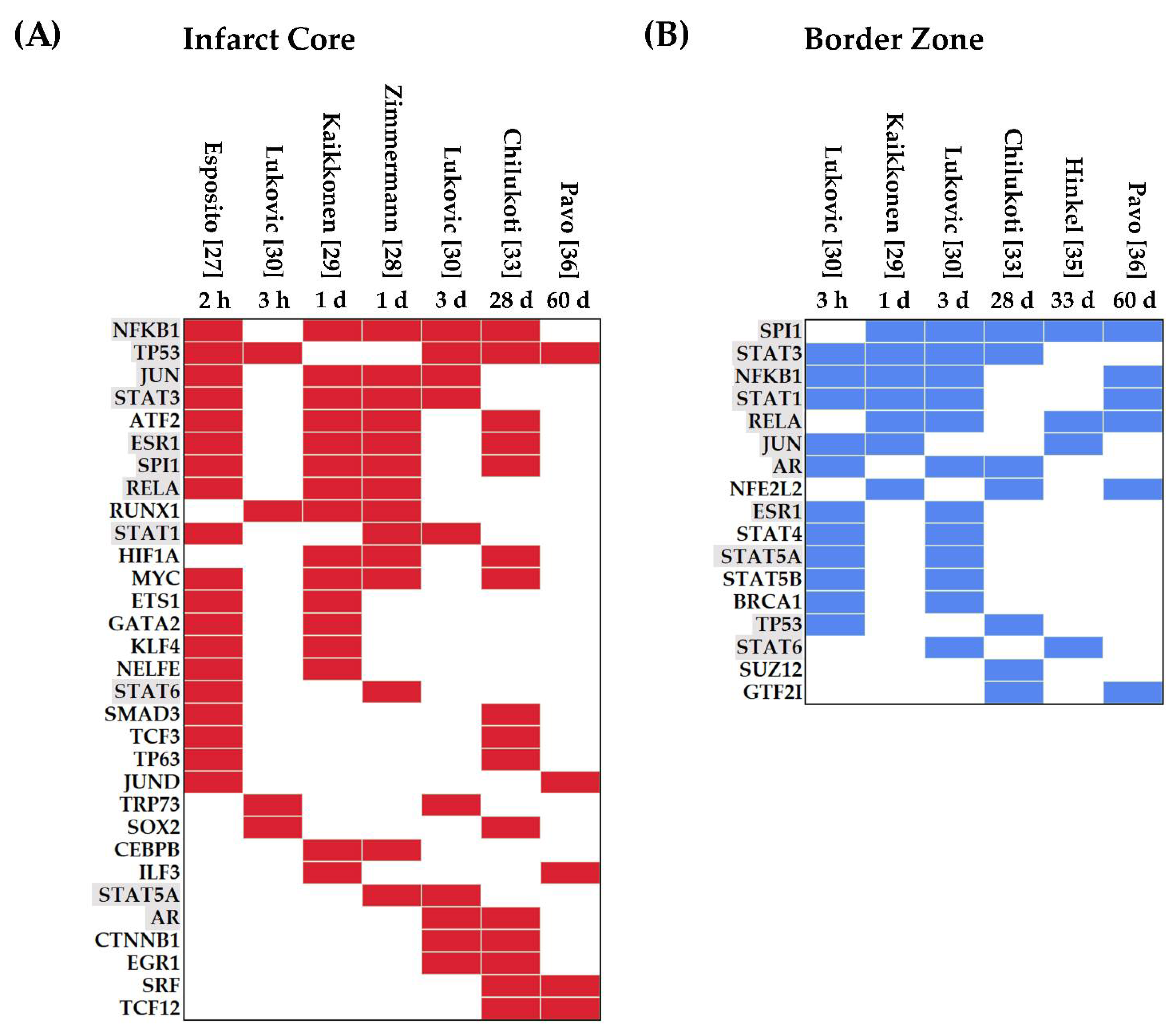
3.1.2. Transcription Factors Upregulated in Myocardial IRI
3.1.3. Molecular Functions and Biological Processes in Myocardial IRI
3.2. Gene Expression Profiles Induced by Cardioprotective Strategies
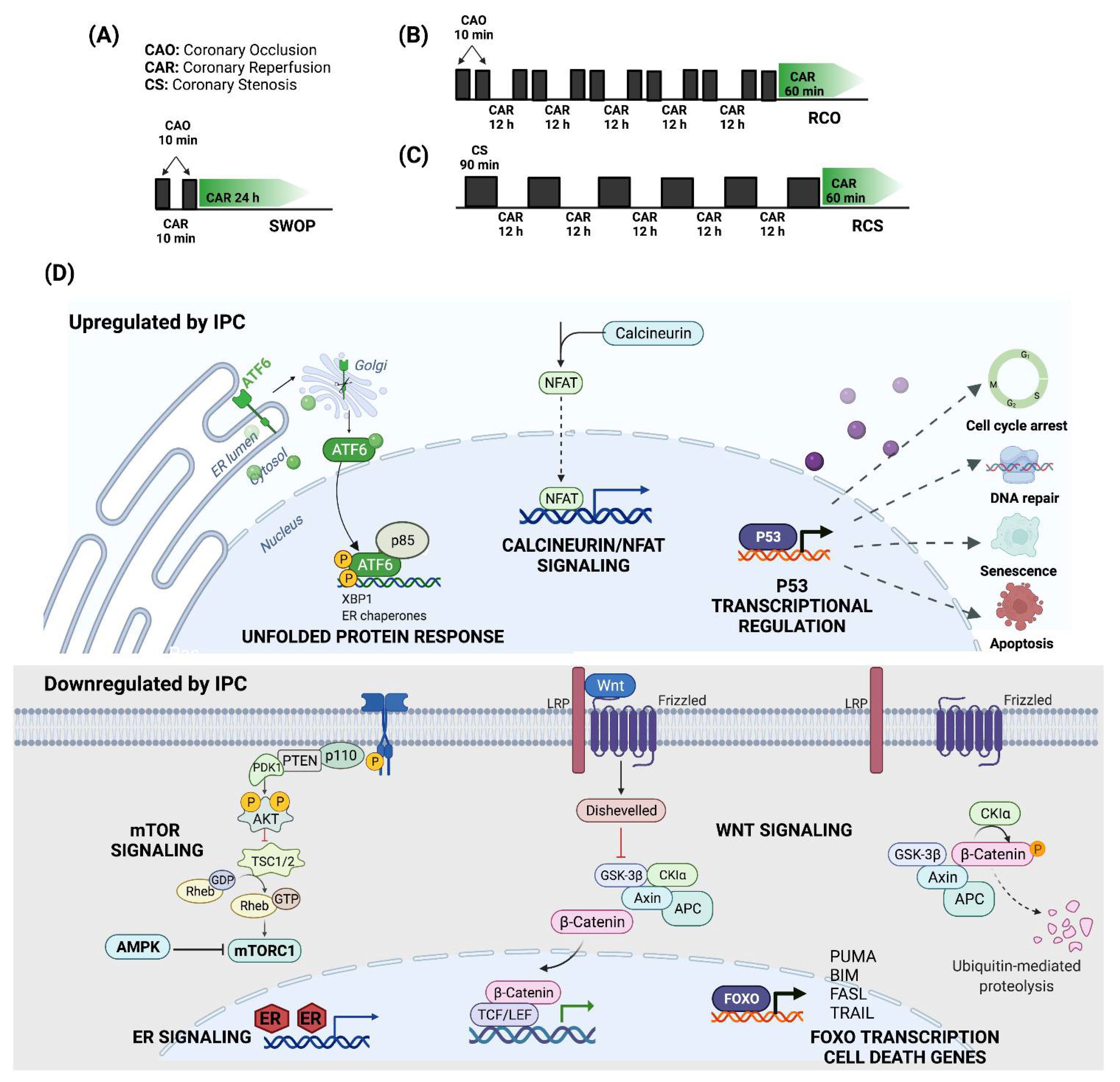
3.2.1. Ischemic Preconditioning
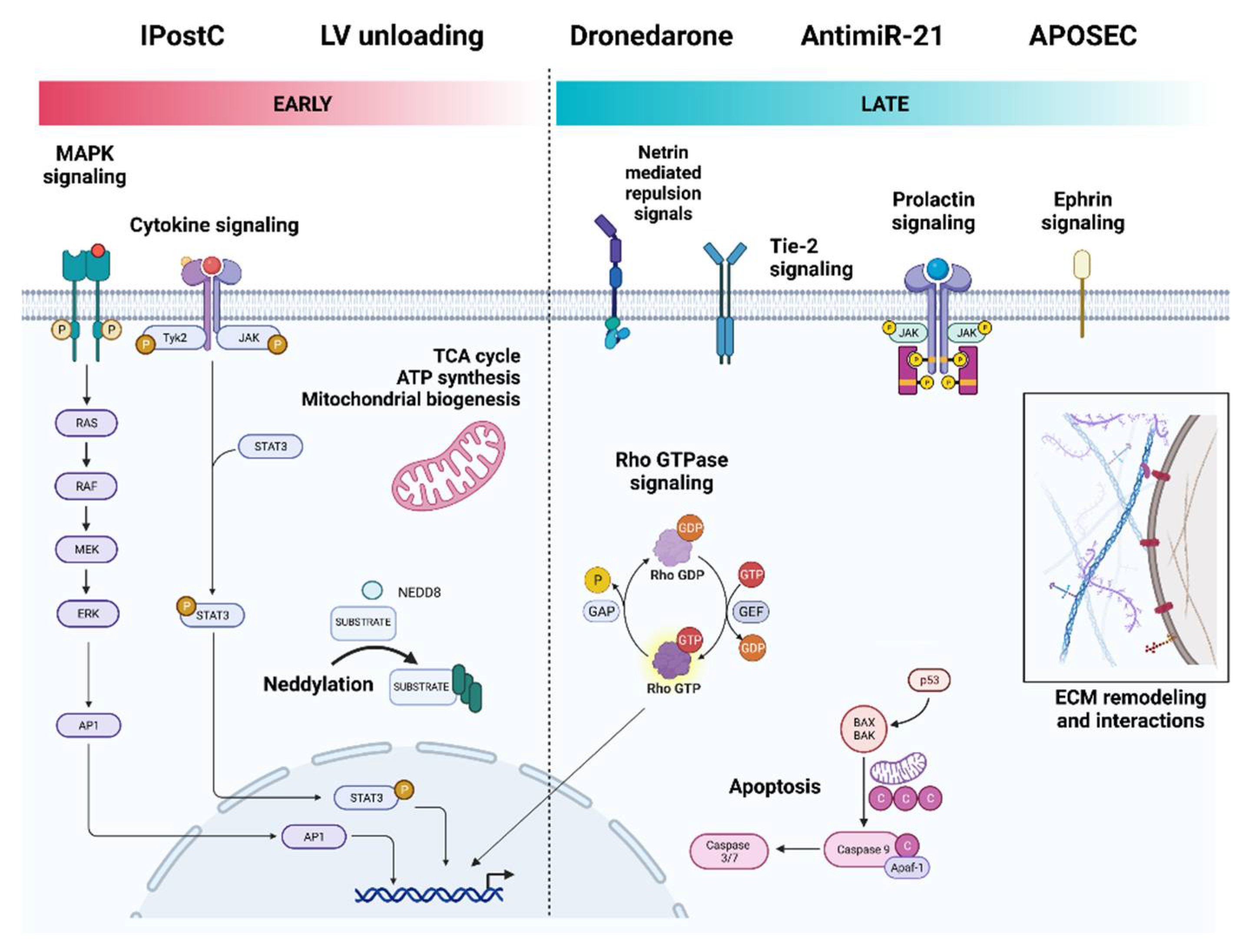
3.2.2. Ischemic Postconditioning
3.2.3. Primary LV Unloading
3.2.4. Pharmacological Cardioprotection
3.2.5. AntagomiRs
3.2.6. Regenerative Therapies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anderson, J.L.; Morrow, D.A. Acute Myocardial Infarction. N. Engl. J. Med. 2017, 376, 2053–2064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Virani, S.S.; Alonso, A.; Aparicio, H.J.; Benjamin, E.J.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Cheng, S.; Delling, F.N.; et al. Heart Disease and Stroke Statistics-2021 Update: A Report From the American Heart Association. Circulation 2021, 143, e254–e743. [Google Scholar] [CrossRef] [PubMed]
- Jennings, R.B.; Murry, C.E.; Steenbergen, C., Jr.; Reimer, K.A. Development of cell injury in sustained acute ischemia. Circulation 1990, 82, II2-12. [Google Scholar] [PubMed]
- Reimer, K.A.; Lowe, J.E.; Rasmussen, M.M.; Jennings, R.B. The wavefront phenomenon of ischemic cell death. 1. Myocardial infarct size vs duration of coronary occlusion in dogs. Circulation 1977, 56, 786–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hausenloy, D.J.; Yellon, D.M. Myocardial ischemia-reperfusion injury: A neglected therapeutic target. J. Clin. Investig. 2013, 123, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Frangogiannis, N.G. The inflammatory response in myocardial injury, repair, and remodelling. Nat. Rev. Cardiol. 2014, 11, 255–265. [Google Scholar] [CrossRef] [Green Version]
- Fröhlich, G.M.; Meier, P.; White, S.K.; Yellon, D.M.; Hausenloy, D.J. Myocardial reperfusion injury: Looking beyond primary PCI. Eur. Heart J. 2013, 34, 1714–1722. [Google Scholar] [CrossRef] [Green Version]
- Vyas, M.V.; Gros, R.; Hackam, D.G. Translation of Cardiovascular Animal Models to Human Randomized Trials. Am. J. Cardiol. 2020, 137, 141. [Google Scholar] [CrossRef]
- Sabroe, I.; Dockrell, D.H.; Vogel, S.N.; Renshaw, S.A.; Whyte, M.K.; Dower, S.K. Identifying and hurdling obstacles to translational research. Nat. Rev. Immunol. 2007, 7, 77–82. [Google Scholar] [CrossRef]
- Fernandez-Moure, J.S. Lost in Translation: The Gap in Scientific Advancements and Clinical Application. Front. Bioeng. Biotechnol. 2016, 4, 43. [Google Scholar] [CrossRef] [Green Version]
- Hooijmans, C.R.; de Vries, R.B.M.; Ritskes-Hoitinga, M.; Rovers, M.M.; Leeflang, M.M.; IntHout, J.; Wever, K.E.; Hooft, L.; de Beer, H.; Kuijpers, T.; et al. Facilitating healthcare decisions by assessing the certainty in the evidence from preclinical animal studies. PLoS ONE 2018, 13, e0187271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hacker, T.A. Animal Models and Cardiac Extracellular Matrix Research. Adv. Exp. Med. Biol. 2018, 1098, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Perleberg, C.; Kind, A.; Schnieke, A. Genetically engineered pigs as models for human disease. Dis. Model. Mech. 2018, 11, dmm030783. [Google Scholar] [CrossRef] [Green Version]
- Gabriel, G.C.; Devine, W.; Redel, B.K.; Whitworth, K.M.; Samuel, M.; Spate, L.D.; Cecil, R.F.; Prather, R.S.; Wu, Y.; Wells, K.D.; et al. Cardiovascular Development and Congenital Heart Disease Modeling in the Pig. J. Am. Heart Assoc. 2021, 10, e021631. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, S.; Hammoudi, N.; Watanabe, S.; Bikou, O.; Yamada, K.; Aguero, J.; Nomoto, K.; Kariya, T.; Fish, K.; Hajjar, R.J.; et al. Echocardiographic Left Ventricular Mass Estimation: Two-Dimensional Area-Length Method is Superior to M-Mode Linear Method in Swine Models of Cardiac Diseases. J. Cardiovasc. Transl. Res. 2020, 13, 648–658. [Google Scholar] [CrossRef] [PubMed]
- Wernersson, R.; Schierup, M.H.; Jørgensen, F.G.; Gorodkin, J.; Panitz, F.; Staerfeldt, H.H.; Christensen, O.F.; Mailund, T.; Hornshøj, H.; Klein, A.; et al. Pigs in sequence space: A 0.66X coverage pig genome survey based on shotgun sequencing. BMC Genom. 2005, 6, 70. [Google Scholar] [CrossRef] [Green Version]
- Fang, X.; Mou, Y.; Huang, Z.; Li, Y.; Han, L.; Zhang, Y.; Feng, Y.; Chen, Y.; Jiang, X.; Zhao, W.; et al. The sequence and analysis of a Chinese pig genome. Gigascience 2012, 1, 16. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.; Wu, J.; Zhou, W.; Liu, X.; Liu, Y.; Zhang, J.; Jia, S.; Li, J.; Wang, H. Identification and analysis of key genes associated with acute myocardial infarction by integrated bioinformatics methods. Medicine 2021, 100, e25553. [Google Scholar] [CrossRef]
- Kabir, M.H.; Patrick, R.; Ho, J.W.K.; O’Connor, M.D. Identification of active signaling pathways by integrating gene expression and protein interaction data. BMC Syst. Biol. 2018, 12, 120. [Google Scholar] [CrossRef]
- Movahed, M.; Brockie, S.; Hong, J.; Fehlings, M.G. Transcriptomic Hallmarks of Ischemia-Reperfusion Injury. Cells 2021, 10, 1838. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. Ann. Intern. Med. 2009, 151, 264–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. NCBI GEO: Archive for functional genomics data sets--update. Nucleic Acids Res. 2013, 41, D991–D995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [Green Version]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. g:Profiler: A web server for functional enrichment analysis and conversions of gene lists (2019 update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef] [Green Version]
- Gillespie, M.; Jassal, B.; Stephan, R.; Milacic, M.; Rothfels, K.; Senff-Ribeiro, A.; Griss, J.; Sevilla, C.; Matthews, L.; Gong, C.; et al. The reactome pathway knowledgebase 2022. Nucleic Acids Res. 2022, 50, D687–D692. [Google Scholar] [CrossRef]
- Esposito, M.L.; Zhang, Y.; Qiao, X.; Reyelt, L.; Paruchuri, V.; Schnitzler, G.R.; Morine, K.J.; Annamalai, S.K.; Bogins, C.; Natov, P.S.; et al. Left Ventricular Unloading Before Reperfusion Promotes Functional Recovery After Acute Myocardial Infarction. J. Am. Coll. Cardiol. 2018, 72, 501–514. [Google Scholar] [CrossRef]
- Zimmermann, M.; Beer, L.; Ullrich, R.; Lukovic, D.; Simader, E.; Traxler, D.; Wagner, T.; Nemec, L.; Altenburger, L.; Zuckermann, A.; et al. Analysis of region specific gene expression patterns in the heart and systemic responses after experimental myocardial ischemia. Oncotarget 2017, 8, 60809–60825. [Google Scholar] [CrossRef] [Green Version]
- Kaikkonen, M.U.; Halonen, P.; Liu, O.H.; Turunen, T.A.; Pajula, J.; Moreau, P.; Selvarajan, I.; Tuomainen, T.; Aavik, E.; Tavi, P.; et al. Genome-Wide Dynamics of Nascent Noncoding RNA Transcription in Porcine Heart After Myocardial Infarction. Circ. Cardiovasc. Genet. 2017, 10, e001702. [Google Scholar] [CrossRef] [Green Version]
- Lukovic, D.; Gugerell, A.; Zlabinger, K.; Winkler, J.; Pavo, N.; Baranyai, T.; Giricz, Z.; Varga, Z.V.; Riesenhuber, M.; Spannbauer, A.; et al. Transcriptional Alterations by Ischaemic Postconditioning in a Pig Infarction Model: Impact on Microvascular Protection. Int. J. Mol. Sci. 2019, 20, 344. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.T.; Depre, C.; Yan, L.; Park, J.Y.; Tian, B.; Jain, K.; Chen, L.; Zhang, Y.; Kudej, R.K.; Zhao, X.; et al. Repetitive ischemia by coronary stenosis induces a novel window of ischemic preconditioning. Circulation 2008, 118, 1961–1969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Depre, C.; Park, J.Y.; Shen, Y.T.; Zhao, X.; Qiu, H.; Yan, L.; Tian, B.; Vatner, S.F.; Vatner, D.E. Molecular mechanisms mediating preconditioning following chronic ischemia differ from those in classical second window. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H752–H762. [Google Scholar] [CrossRef] [PubMed]
- Chilukoti, R.K.; Lendeckel, J.; Darm, K.; Bukowska, A.; Goette, A.; Sühling, M.; Utpatel, K.; Peters, B.; Homuth, G.; Völker, U.; et al. Integration of “omics” techniques: Dronedarone affects cardiac remodeling in the infarction border zone. Exp. Biol. Med. 2018, 243, 895–910. [Google Scholar] [CrossRef] [PubMed]
- Agnew, E.J.; Velayutham, N.; Matos Ortiz, G.; Alfieri, C.M.; Hortells, L.; Moore, V.; Riggs, K.W.; Baker, R.S.; Gibson, A.M.; Ponny, S.R.; et al. Scar Formation with Decreased Cardiac Function Following Ischemia/Reperfusion Injury in 1 Month Old Swine. J. Cardiovasc. Dev. Dis. 2019, 7, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinkel, R.; Ramanujam, D.; Kaczmarek, V.; Howe, A.; Klett, K.; Beck, C.; Dueck, A.; Thum, T.; Laugwitz, K.L.; Maegdefessel, L.; et al. AntimiR-21 Prevents Myocardial Dysfunction in a Pig Model of Ischemia/Reperfusion Injury. J. Am. Coll. Cardiol. 2020, 75, 1788–1800. [Google Scholar] [CrossRef] [PubMed]
- Pavo, N.; Zimmermann, M.; Pils, D.; Mildner, M.; Petrási, Z.; Petneházy, Ö.; Fuzik, J.; Jakab, A.; Gabriel, C.; Sipos, W.; et al. Long-acting beneficial effect of percutaneously intramyocardially delivered secretome of apoptotic peripheral blood cells on porcine chronic ischemic left ventricular dysfunction. Biomaterials 2014, 35, 3541–3550. [Google Scholar] [CrossRef] [Green Version]
- Prabhu, S.D.; Frangogiannis, N.G. The Biological Basis for Cardiac Repair After Myocardial Infarction: From Inflammation to Fibrosis. Circ. Res. 2016, 119, 91–112. [Google Scholar] [CrossRef]
- Weidemüller, P.; Kholmatov, M.; Petsalaki, E.; Zaugg, J.B. Transcription factors: Bridge between cell signaling and gene regulation. Proteomics 2021, 21, e2000034. [Google Scholar] [CrossRef]
- Bhoumik, A.; Takahashi, S.; Breitweiser, W.; Shiloh, Y.; Jones, N.; Ronai, Z. ATM-dependent phosphorylation of ATF2 is required for the DNA damage response. Mol. Cell 2005, 18, 577–587. [Google Scholar] [CrossRef] [Green Version]
- Bellissimo, D.C.; Chen, C.H.; Zhu, Q.; Bagga, S.; Lee, C.T.; He, B.; Wertheim, G.B.; Jordan, M.; Tan, K.; Worthen, G.S.; et al. Runx1 negatively regulates inflammatory cytokine production by neutrophils in response to Toll-like receptor signaling. Blood Adv. 2020, 4, 1145–1158. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, B.; Liu, X.; Deng, Y.; Zhu, Y.; Zhu, F.; Liang, Y.; Li, H. Sp1 Targeted PARP1 Inhibition Protects Cardiomyocytes From Myocardial Ischemia-Reperfusion Injury via Downregulation of Autophagy. Front. Cell Dev. Biol. 2021, 9, 621906. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, Y.; Aihara, K.; Sato, T.; Akaike, M.; Yoshizumi, M.; Suzaki, Y.; Izawa, Y.; Fujimura, M.; Hashizume, S.; Kato, M.; et al. Androgen receptor gene knockout male mice exhibit impaired cardiac growth and exacerbation of angiotensin II-induced cardiac fibrosis. J. Biol. Chem. 2005, 280, 29661–29666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Y.; Ho, B.X.; Pang, J.K.S.; Pek, N.M.Q.; Hor, J.H.; Ng, S.Y.; Soh, B.S. Wnt/β-catenin-mediated signaling re-activates proliferation of matured cardiomyocytes. Stem. Cell Res. Ther. 2018, 9, 338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zins, K.; Pomyje, J.; Hofer, E.; Abraham, D.; Lucas, T.; Aharinejad, S. Egr-1 upregulates Siva-1 expression and induces cardiac fibroblast apoptosis. Int. J. Mol. Sci. 2014, 15, 1538–1553. [Google Scholar] [CrossRef] [Green Version]
- Barron, M.R.; Belaguli, N.S.; Zhang, S.X.; Trinh, M.; Iyer, D.; Merlo, X.; Lough, J.W.; Parmacek, M.S.; Bruneau, B.G.; Schwartz, R.J. Serum response factor, an enriched cardiac mesoderm obligatory factor, is a downstream gene target for Tbx genes. J. Biol. Chem. 2005, 280, 11816–11828. [Google Scholar] [CrossRef] [Green Version]
- Tandon, P.; Miteva, Y.V.; Kuchenbrod, L.M.; Cristea, I.M.; Conlon, F.L. Tcf21 regulates the specification and maturation of proepicardial cells. Development 2013, 140, 2409–2421. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.C.; Nguyen, T.; Pickett, C.B. Regulation of the antioxidant response element by protein kinase C-mediated phosphorylation of NF-E2-related factor 2. Proc. Natl. Acad. Sci. USA 2000, 97, 12475–12480. [Google Scholar] [CrossRef] [Green Version]
- Eggler, A.L.; Small, E.; Hannink, M.; Mesecar, A.D. Cul3-mediated Nrf2 ubiquitination and antioxidant response element (ARE) activation are dependent on the partial molar volume at position 151 of Keap1. Biochem. J. 2009, 422, 171–180. [Google Scholar] [CrossRef] [Green Version]
- Frangogiannis, N.G. The role of the chemokines in myocardial ischemia and reperfusion. Curr. Vasc. Pharmacol. 2004, 2, 163–174. [Google Scholar] [CrossRef]
- Murry, C.E.; Jennings, R.B.; Reimer, K.A. Preconditioning with ischemia: A delay of lethal cell injury in ischemic myocardium. Circulation 1986, 74, 1124–1136. [Google Scholar] [CrossRef] [Green Version]
- Laskey, W.K.; Yoon, S.; Calzada, N.; Ricciardi, M.J. Concordant improvements in coronary flow reserve and ST-segment resolution during percutaneous coronary intervention for acute myocardial infarction: A benefit of postconditioning. Catheter. Cardiovasc. Interv. 2008, 72, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Lou, B.; Cui, Y.; Gao, H.; Chen, M. Meta-analysis of the effects of ischemic postconditioning on structural pathology in ST-segment elevation acute myocardial infarction. Oncotarget 2018, 9, 8089–8099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burkhoff, D.; Sayer, G.; Doshi, D.; Uriel, N. Hemodynamics of Mechanical Circulatory Support. J. Am. Coll. Cardiol. 2015, 66, 2663–2674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapur, N.K.; Qiao, X.; Paruchuri, V.; Morine, K.J.; Syed, W.; Dow, S.; Shah, N.; Pandian, N.; Karas, R.H. Mechanical Pre-Conditioning with Acute Circulatory Support Before Reperfusion Limits Infarct Size in Acute Myocardial Infarction. JACC Heart Fail. 2015, 3, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Bromage, D.I.; Davidson, S.M.; Yellon, D.M. Stromal derived factor 1α: A chemokine that delivers a two-pronged defence of the myocardium. Pharmacol. Ther. 2014, 143, 305–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, S.P.; Bolli, R. The ubiquitous role of nitric oxide in cardioprotection. J. Mol. Cell Cardiol. 2006, 40, 16–23. [Google Scholar] [CrossRef]
- Lasley, R.D. Adenosine Receptor-Mediated Cardioprotection-Current Limitations and Future Directions. Front. Pharmacol. 2018, 9, 310. [Google Scholar] [CrossRef]
- Pagliaro, P.; Penna, C. Redox signalling and cardioprotection: Translatability and mechanism. Br. J. Pharmacol. 2015, 172, 1974–1995. [Google Scholar] [CrossRef] [Green Version]
- Connolly, S.J.; Camm, A.J.; Halperin, J.L.; Joyner, C.; Alings, M.; Amerena, J.; Atar, D.; Avezum, Á.; Blomström, P.; Borggrefe, M.; et al. Dronedarone in high-risk permanent atrial fibrillation. N. Engl. J. Med. 2011, 365, 2268–2276. [Google Scholar] [CrossRef] [Green Version]
- Skyschally, A.; Heusch, G. Reduction of myocardial infarct size by dronedarone in pigs—A pleiotropic action? Cardiovasc. Drugs Ther. 2011, 25, 197–201. [Google Scholar] [CrossRef] [Green Version]
- Engelhorn, T.; Schwarz, M.A.; Heusch, G.; Doerfler, A.; Schulz, R. Reduction of cerebral infarct size by dronedarone. Cardiovasc. Drugs Ther. 2011, 25, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, S.; Cheng, Y.; Yang, J.; Li, J.; Liu, X.; Wang, X.; Wang, D.; Krall, T.J.; Delphin, E.S.; Zhang, C. MicroRNA expression signature and the role of microRNA-21 in the early phase of acute myocardial infarction. J. Biol. Chem. 2009, 284, 29514–29525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, P.D.; de Bakker, D.E.M.; Bakkers, J. Cardiac regenerative capacity: An evolutionary afterthought? Cell Mol. Life Sci. 2021, 78, 5107–5122. [Google Scholar] [CrossRef] [PubMed]
- Carotenuto, F.; Teodori, L.; Maccari, A.M.; Delbono, L.; Orlando, G.; Di Nardo, P. Turning regenerative technologies into treatment to repair myocardial injuries. J. Cell. Mol. Med. 2020, 24, 2704–2716. [Google Scholar] [CrossRef] [Green Version]
- Lichtenauer, M.; Mildner, M.; Hoetzenecker, K.; Zimmermann, M.; Podesser, B.K.; Sipos, W.; Berényi, E.; Dworschak, M.; Tschachler, E.; Gyöngyösi, M.; et al. Secretome of apoptotic peripheral blood cells (APOSEC) confers cytoprotection to cardiomyocytes and inhibits tissue remodelling after acute myocardial infarction: A preclinical study. Basic Res. Cardiol. 2011, 106, 1283–1297. [Google Scholar] [CrossRef] [Green Version]
- Dixon, J.A.; Gorman, R.C.; Stroud, R.E.; Bouges, S.; Hirotsugu, H.; Gorman, J.H., 3rd; Martens, T.P.; Itescu, S.; Schuster, M.D.; Plappert, T.; et al. Mesenchymal cell transplantation and myocardial remodeling after myocardial infarction. Circulation 2009, 120, S220–S229. [Google Scholar] [CrossRef] [Green Version]
- Renner, S.; Blutke, A.; Clauss, S.; Deeg, C.A.; Kemter, E.; Merkus, D.; Wanke, R.; Wolf, E. Porcine models for studying complications and organ crosstalk in diabetes mellitus. Cell Tissue Res. 2020, 380, 341–378. [Google Scholar] [CrossRef]
- Ramachandran, P.; Iredale, J.P. Liver fibrosis: A bidirectional model of fibrogenesis and resolution. Qjm 2012, 105, 813–817. [Google Scholar] [CrossRef] [Green Version]
- Travers, J.G.; Kamal, F.A.; Robbins, J.; Yutzey, K.E.; Blaxall, B.C. Cardiac Fibrosis: The Fibroblast Awakens. Circ. Res. 2016, 118, 1021–1040. [Google Scholar] [CrossRef] [Green Version]
- Mahtta, D.; Sudhakar, D.; Koneru, S.; Silva, G.V.; Alam, M.; Virani, S.S.; Jneid, H. Targeting Inflammation After Myocardial Infarction. Curr. Cardiol. Rep. 2020, 22, 110. [Google Scholar] [CrossRef] [PubMed]
- Baines, C.P.; Zhang, J.; Wang, G.W.; Zheng, Y.T.; Xiu, J.X.; Cardwell, E.M.; Bolli, R.; Ping, P. Mitochondrial PKCepsilon and MAPK form signaling modules in the murine heart: Enhanced mitochondrial PKCepsilon-MAPK interactions and differential MAPK activation in PKCepsilon-induced cardioprotection. Circ. Res. 2002, 90, 390–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gross, G.J.; Peart, J.N. KATP channels and myocardial preconditioning: An update. Am. J. Physiol. Heart Circ. Physiol. 2003, 285, H921–H930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baines, C.P.; Liu, G.S.; Birincioglu, M.; Critz, S.D.; Cohen, M.V.; Downey, J.M. Ischemic preconditioning depends on interaction between mitochondrial KATP channels and actin cytoskeleton. Am. J. Physiol. 1999, 276, H1361–H1368. [Google Scholar] [CrossRef]
- Yehualashet, A.S.; Belachew, T.F.; Kifle, Z.D.; Abebe, A.M. Targeting Cardiac Metabolic Pathways: A Role in Ischemic Management. Vasc. Health Risk Manag. 2020, 16, 353–365. [Google Scholar] [CrossRef]
- Li, J.; Ma, W.; Li, H.; Hou, N.; Wang, X.; Kim, I.M.; Li, F.; Su, H. NEDD8 Ultimate Buster 1 Long (NUB1L) Protein Suppresses Atypical Neddylation and Promotes the Proteasomal Degradation of Misfolded Proteins. J. Biol. Chem. 2015, 290, 23850–23862. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zou, J.; Littlejohn, R.; Liu, J.; Su, H. Neddylation, an Emerging Mechanism Regulating Cardiac Development and Function. Front. Physiol. 2020, 11, 612927. [Google Scholar] [CrossRef]
- Zhang, J.; Cui, J.; Zhao, F.; Yang, L.; Xu, X.; Shi, Y.; Wei, B. Cardioprotective effect of MLN4924 on ameliorating autophagic flux impairment in myocardial ischemia-reperfusion injury by Sirt1. Redox. Biol. 2021, 46, 102114. [Google Scholar] [CrossRef]
- Dries, J.L.; Kent, S.D.; Virag, J.A. Intramyocardial administration of chimeric ephrinA1-Fc promotes tissue salvage following myocardial infarction in mice. J. Physiol. 2011, 589, 1725–1740. [Google Scholar] [CrossRef]
- O’Neal, W.T.; Griffin, W.F.; Dries-Devlin, J.L.; Kent, S.D.; Chen, J.; Willis, M.S.; Virag, J.A. Ephrin-Eph signaling as a potential therapeutic target for the treatment of myocardial infarction. Med. Hypotheses 2013, 80, 738–744. [Google Scholar] [CrossRef] [Green Version]
- Suri, C.; Jones, P.F.; Patan, S.; Bartunkova, S.; Maisonpierre, P.C.; Davis, S.; Sato, T.N.; Yancopoulos, G.D. Requisite role of angiopoietin-1, a ligand for the TIE2 receptor, during embryonic angiogenesis. Cell 1996, 87, 1171–1180. [Google Scholar] [CrossRef] [Green Version]
- Dumont, D.J.; Gradwohl, G.; Fong, G.H.; Puri, M.C.; Gertsenstein, M.; Auerbach, A.; Breitman, M.L. Dominant-negative and targeted null mutations in the endothelial receptor tyrosine kinase, tek, reveal a critical role in vasculogenesis of the embryo. Genes Dev. 1994, 8, 1897–1909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.W.; Won, J.Y.; Lee, H.Y.; Lee, H.J.; Youn, S.W.; Lee, J.Y.; Cho, C.H.; Cho, H.J.; Oh, S.; Chae, I.H.; et al. Angiopoietin-1 protects heart against ischemia/reperfusion injury through VE-cadherin dephosphorylation and myocardiac integrin-β1/ERK/caspase-9 phosphorylation cascade. Mol. Med. 2011, 17, 1095–1106. [Google Scholar] [CrossRef]
- Guthrie, S. Axon guidance: Netrin receptors are revealed. Curr. Biol. 1997, 7, R6–R9. [Google Scholar] [CrossRef] [Green Version]
- Layne, K.; Ferro, A.; Passacquale, G. Netrin-1 as a novel therapeutic target in cardiovascular disease: To activate or inhibit? Cardiovasc. Res. 2015, 107, 410–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Cai, H. Induction of cardioprotection by small netrin-1-derived peptides. Am. J. Physiol. Cell Physiol. 2015, 309, C100–C106. [Google Scholar] [CrossRef] [PubMed]
- Kilian, L.S.; Voran, J.; Frank, D.; Rangrez, A.Y. RhoA: A dubious molecule in cardiac pathophysiology. J. Biomed. Sci. 2021, 28, 33. [Google Scholar] [CrossRef]
- Sah, V.P.; Minamisawa, S.; Tam, S.P.; Wu, T.H.; Dorn, G.W., 2nd; Ross, J., Jr.; Chien, K.R.; Brown, J.H. Cardiac-specific overexpression of RhoA results in sinus and atrioventricular nodal dysfunction and contractile failure. J. Clin. Investig. 1999, 103, 1627–1634. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.H.; Del Re, D.P.; Sussman, M.A. The Rac and Rho hall of fame: A decade of hypertrophic signaling hits. Circ. Res. 2006, 98, 730–742. [Google Scholar] [CrossRef] [Green Version]
- Lauriol, J.; Keith, K.; Jaffré, F.; Couvillon, A.; Saci, A.; Goonasekera, S.A.; McCarthy, J.R.; Kessinger, C.W.; Wang, J.; Ke, Q.; et al. RhoA signaling in cardiomyocytes protects against stress-induced heart failure but facilitates cardiac fibrosis. Sci. Signal. 2014, 7, ra100. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, M.; Sadoshima, J. Mechanisms of physiological and pathological cardiac hypertrophy. Nat. Rev. Cardiol. 2018, 15, 387–407. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| References | Breed | Sex | Occlusion/Reperfusion | Area | Intervention | Platform | GEO Accession |
|---|---|---|---|---|---|---|---|
| Esposito ML et al., 2018 [27] * | Yorkshire | Male | 90 min/120 min | IC | LV unloading | Porcine 1.0 ST microarrays (Affymetrix) | GSE108644 |
| Zimmer-mann M et al., 2017 [28] * | Domestic | Female | 90 min/1 d | IC/BZ/RZ | - | Whole Porcine Genome Oligo Microarray (Agilent) | - |
| Kaikkonen MU et al., 2017 [29] * | Farm | Female | 90 min/1 d | IC/BZ | - | GRO-SeqIllumina HiSeq2000 | GSE81155 |
| Lukovic D et al., 2019 [30] * | Domestic | Female | 90 min/3 h 90 min/3 d | IC/RZ | IPostC | RNA-SeqIllumina HiSeq2500 | - |
| Shen YT et al., 2008 [31] | Unspecified | Unspecified | 60 min/4 d | SE | IPC | Porcine Genome Array (Affymetrix) | - |
| Depre C et al., 2010 [32] | Domestic | Female | 10 min Twice/1 d 10 min Twicex6/1 h | Heart | IPC | Porcine Genome Array (Affymetrix) | GSE21096 |
| Chilukoti RK et al., 2018 [33] * | German Landrace | Male, castrated | 90 min/28 d | IC/BZ | Dronedarone | GeneChip Porcine Genome Arrays (Affymetrix) | - |
| Agnew EJ et al., 2019 [34] | Yorkshire Landrace | Male/Female | 60 min/30 d | LV | Age | RNA-SeqIllumina NovaSeq6000 | GSE137293 |
| Hinkel R et al., 2020 [35] * | German Landrace | Male/Female | 60 min/33 d | BZ | LNA-antimiR-21 | RNA-SeqIllumina HiSeq2000 | - |
| Pavo N et al., 2014 [36] * | Domestic | Female | 90 min/60 d | IC/BZ | APOSEC | Sus Scrofa Oligo Microarray v2 (Agilent) | GSE47397 |
| IPC Model | Pathway Name | |
|---|---|---|
| Upregulated | Downregulated | |
| RCO |
|
|
| RCS |
|
|
| SWOP |
|
|
| RCP/RCS/SWOP |
|
|
| Cardioprotective Intervention | Pathway Name | |
|---|---|---|
| Upregulated | Downregulated | |
| IPostC (3 d) |
|
|
| LV unloading (2 h) |
|
|
| Dronedarone (28 d) |
|
|
| LNA-antimiR-21 (33 d) |
|
|
| APOSEC (60 d) |
|
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salazar-Gonzalez, H.; Gutierrez-Mercado, Y.K.; Munguia-Galaviz, F.J.; Echavarria, R. Signaling Pathways Involved in Myocardial Ischemia–Reperfusion Injury and Cardioprotection: A Systematic Review of Transcriptomic Studies in Sus scrofa. J. Cardiovasc. Dev. Dis. 2022, 9, 132. https://doi.org/10.3390/jcdd9050132
Salazar-Gonzalez H, Gutierrez-Mercado YK, Munguia-Galaviz FJ, Echavarria R. Signaling Pathways Involved in Myocardial Ischemia–Reperfusion Injury and Cardioprotection: A Systematic Review of Transcriptomic Studies in Sus scrofa. Journal of Cardiovascular Development and Disease. 2022; 9(5):132. https://doi.org/10.3390/jcdd9050132
Chicago/Turabian StyleSalazar-Gonzalez, Hector, Yanet Karina Gutierrez-Mercado, Francisco Javier Munguia-Galaviz, and Raquel Echavarria. 2022. "Signaling Pathways Involved in Myocardial Ischemia–Reperfusion Injury and Cardioprotection: A Systematic Review of Transcriptomic Studies in Sus scrofa" Journal of Cardiovascular Development and Disease 9, no. 5: 132. https://doi.org/10.3390/jcdd9050132