Wine Contamination with Ochratoxins: A Review




1
Department of Microbiology III, Faculty of Biology, University Complutense of Madrid, Jose Antonio Novais 12, 28040 Madrid, Spain
2
Department of Genetics, Faculty of Biology, University Complutense of Madrid, Jose Antonio Novais 12, 28040 Madrid, Spain
*
Author to whom correspondence should be addressed.
Beverages 2018, 4(1), 6; https://doi.org/10.3390/beverages4010006
Submission received: 31 October 2017
/
Revised: 15 December 2017
/
Accepted: 29 December 2017
/
Published: 15 January 2018
(This article belongs to the Special Issue Mycotoxins in Beverages)
Abstract
:Ochratoxin A (OTA) is the main mycotoxin occurring in wine. This review article is focused on the distribution of this toxin and its producing-fungi in grape berries, as well as on the fate of OTA during winemaking procedures. Due to its toxic properties, OTA levels in wine are regulated in different countries; therefore, it is necessary to apply control and detoxification methods that are also discussed in this revision.
1. Introduction
Ochratoxin A (OTA) is a widespread mycotoxin that is produced by several Aspergillus and Penicillium species. OTA is the most toxic compound included in the ochratoxin group, and, structurally is a chlorinated isocoumarin compound (Figure 1) [1]. There are several metabolites related to OTA although they are considered less important. The most relevant ones are the dechloro analog of OTA (ochratoxin B, OTB), its ethyl ester (ochratoxin C, OTC), the isocoumaric derivative of OTA (ochratoxin α, OTα), and its dechloro analog (ochratoxin β, OTβ) [2] (Figure 1).
P. nordicum and P. verrucosum are the main Penicillium species that are capable of producing OTA and are often related to temperate regions [2]. OTA-producing Aspergillus species are included in sections Circumdati and Nigri. The new advances in phylogenetic analysis and secondary metabolite profile characterization have allowed for the reclassification and description of new species inside these sections. Nowadays, the main OTA-producing species in Aspergillus section Circumdati are A. steynii, A. westerdijkiae and A. ochraceus [1], whereas A. carbonarius, A. niger and A. welwitschiae are the most important producers in section Nigri [3].
OTA has been found in a variety of products worldwide, such as cereals and derivatives, grape and grape products, coffee, nuts, spices, and cocoa [4]. This toxin presents nephrotoxic, neurotoxic, teratogenic, and immunotoxic properties towards both animals and humans and has been classified as a possible human carcinogen by the International Agency for Research on Cancer [5].
Since the first report on the occurrence of OTA in wine [6], many reports have studied the causes of this contamination, as well as how to prevent its presence in this product [7]. Since 2006, the maximum OTA levels in wine are strictly regulated by the European Commission and several countries have also laid down limits for OTA in wine following EU directives [8].
Contamination by ochratoxigenic species seems to occur in the vineyard and many factors are reported to affect OTA levels in wine, including both biotic and abiotic factors. OTA concentration is also strongly modified during winemaking procedures [9]. This paper is a revision of the most relevant studies about this issue and compiles the results obtained related to ochratoxin presence in wine.
2. Ochratoxigenic Species on Grapes
It is widely known that OTA presence in wine is closely related to fungal contamination in the vineyards. Many studies have been performed to unravel the most important ochratoxigenic species occurring in grapes. Several reports evidenced the contribution of Aspergillus section Nigri species to OTA contamination of grapes and their occurrence on the surface of healthy berries [10]. However, their prevalence differs depending on the geographical region where the grapes are cultivated [11]. In general, A. carbonarius is considered as the most important ochratoxigenic species in grapes due to its common occurrence and its high production ability (almost all of the strains are able to produce the toxin at different levels). The species included in Aspergillus niger aggregate (mainly A. niger, A. welwitschiae, and A. tubingensis) are also frequently found in grapes worldwide, although few strains are able to produce the toxin and usually reach low production levels; therefore, they are considered to slightly contribute to OTA contamination of grapes. Some studies have reported grape contamination by other ochratoxigenic species that are included in Aspergillus section Nigri such as A. japonicus and A. aculeatus, as well as by OTA-producing species of Aspergillus section Circumdati, such as A. steynii, A. westerdijkiae, and A. ochraceus. Penicillium species have been also reported to occur in grapes but their contribution to OTA contamination is not well established yet [11].
The contribution of each species to OTA content of grapes and wine is hardly influenced by geographical origin. The surveys performed in Mediterranean countries in Southern Europe including Spain [12,13,14], Portugal [15,16], France [17,18], Italy [19,20], and Greece [21] seem to indicate that A. carbonarius is the most important OTA-producing species in grapes in this region although A. niger aggregate species are the most frequently isolated.
This pattern was also found in other Mediterranean countries in Northern Africa, indicating that climate might be influencing OTA-producing species distribution. Lasram et al. [22] indicated that Aspergillus section Nigri isolates were the only potential OTA-producing fungi from grapes in Tunez being A. niger aggregate species the most abundant, followed by A. carbonarius. However, the highest risk of OTA contamination was also attributed to A. carbonarius with a 97% of producing strains, whereas only 3% of A. niger aggregate isolates were able to produce OTA. In this country, similar results were obtained by Fredj et al. [23] who reported a high incidence of A. niger aggregate isolates. In this study, A. carbonarius also represented an important risk for OTA contamination with more than 80% of producing strains (compared to only 5% of producers in A. niger aggregate). A survey carried out in Lebanon also reported that A. carbonarius was the most important OTA producer in wine grapes reaching 100% of producing isolates but A. niger aggregate species were far more frequent [24].
The situation is similar in South American countries, as well as in Australia, where the main species isolated from wine grapes belong to A. niger aggregate [25,26]. However, A. niger aggregate species from Brazilian and Argentinean grape berries present a high percentage of OTA producers with levels up to 68% of producing isolates [27,28,29].
As mentioned before, many reports indicate that A. niger aggregate isolates are commonly found in the surface of grape berries; however, little information is available regarding which species are the most frequently occurring and their distribution seems to be related to geographical patterns. Recently, Pantelides et al. [30] performed an extensive analysis of OTA-producing species in Cyprus vineyards and found that A. tubingensis was the predominant species (92% of isolates), followed by A. niger (5%) and A. welwitschiae (2%). However, Medina et al. [31] reported that the most frequent species of A. niger aggregate in Spanish grapes was A. niger (75% of isolates) and only 25% of the isolates were classified as A. tubingensis. This paper was published before A. welwitschiae description and it might be possible that some A. niger isolates needed to be reclassified. The situation seems to be quite different out of Europe. Garmendia and Vero [32] described A. welwitschiae as the main species of A. niger aggregate in Uruguayan grapes, whereas no A. tubingensis isolates were found. In a recently published work, Qi et al. [33] also described A. welwitschiae as the most frequently isolated species included in A. niger aggregate in Canadian grapes and they found that only A. welwitschiae and A. niger were able to synthesize OTA.
A. japonicus is also occasionally isolated from grapes, but its production ability in vitro has not been demonstrated yet [12,15,17,24]. A. uvarum is also an uniseriate species that is included in Aspergillus section Nigri that has been reported to occur in grapes in Mediterranean countries but any isolate has been described as OTA producer [34]. The presence of uniseriate species seems to be more related to the first growth stages in the vineyard and their occurrence is drastically reduced at veraison and harvest periods [19].
Aspergillus section Nigri isolates are adapted to survive in warm climates and opportunistically infect grapes when berries are damaged by biotic and abiotic factors. Soil is the primary source of inoculum and spores can easily move from soil to berry surface [26,35]. Moreover, black spores of Aspergillus section Nigri species are quite protected from sun and UV light, which provide them a competitive advantage to resist in warm climates, mainly in the case of A. carbonarius [36]. Several works have studied ecophysiological factors affecting fungal growth and OTA production by Aspergillus section Nigri strains in grape-based medium [37,38,39,40]. A. niger and A. carbonarius present an optimum temperature for growth around 30–35 °C although maximum OTA production is reached at lower temperatures (15–20 °C) closer to that found during night when OTA accumulation might occur. High water activity levels favor both growth and OTA production; therefore, rainfall close to harvest might potentiate toxin accumulation on grapes.
OTA-producing species of Aspergillus section Circumdati have been also isolated from grapes worldwide. Their contamination levels are usually very low although in some cases they show high ochratoxigenic ability [12,15,27,41].
The source of OTA contamination in grapes in colder temperate regions is not well established yet. Black Aspergilli have not been isolated from grape berries although in a few cases ochratoxigenic Penicillia have been described. This may indicate that Penicillium species might be responsible for OTA presence in grapes and wines in these regions [11].
3. Factors Affecting Ochratoxin A Presence on Grapes, Musts and Wines
Many studies reveal that OTA contamination of wine occur worldwide. Zimmerli and Dick [6] first reported the presence of OTA in grape juices and wines. Since then, the presence of this toxin in wines has been described in many countries and different types of wine. The levels of contamination seem to be related to several factors that are explained below. They were selected because they analyzed a significant numbers of samples and applied appropriate sampling and analytical methods. Table 1 collects the most significant and extensive analyses of OTA occurrence in wines worldwide. The incidence of the toxin in wines is quite extensive, but rarely OTA content exceeds the limits set by the European Union and some countries (usually 2 µg/L). Taking into account all of the reports indicated in Table 1, only 1.2% of the samples analyzed are above legal maximum levels.
As mentioned before, the source of OTA contamination of wine is the raw material, and, therefore, sanitary condition of grapes is a very important factor to avoid OTA presence in the final product [66]. OTA levels in wine also depend on several factors that are related to environmental and managing conditions in the vineyards as well as to winemaking procedures. These factors are presented and discussed below.
3.1. Location (Latitude)
Several studies on the occurrence of OTA in wines in Europe have been reported since the first report was published [6]. An extensive review collected data from several countries, indicating that OTA incidence was significantly higher in Southern Europe (72% of positive samples) than in Northern regions (50% of contaminated wines) [67]. The considerable climatic variations among geographic regions might be influencing growth and toxin production by ochratoxigenic species. This tendency is more obvious in red wines than in white ones [20]. European countries are divided into Wine Growing Zones depending on their climatic conditions. Zone A covers growing areas in Germany, Luxembourg, and the United Kingdom, whereas zone B includes southern Germany and the North of France. Zone C represents the most southern growing zones in Europe including Italy, Spain, and Greece [68]. Otteneder and Majerus [68] confirmed that wines produced in Northern regions (zones A and B) were considerably less contaminated than those produced in South Europe (zone C) with 12% and 95% of contaminated samples, respectively.
This situation could be extrapolated to Northern and Southern regions inside the same country. Pietri et al. [54] noticed a gradual increase of OTA contamination from North to South Italy that attributed to considerable climatic differences occurring in those latitudes. Something similar was found in Greece by Stefanaki et al. [50] who reported a concentration gradient from North to South probably due to high humidity and temperature in Southern regions of the country.
However, some reports disagreed with this north to south gradient and did not found higher contamination levels in Southern Europe countries [69]. Therefore, this factor might be interacting with some others and OTA levels could vary depending on the samples analyzed.
3.2. Biotic Factors
As mentioned before, ochratoxigenic Aspergillus species are able to survive in vineyard soils or the surface of berries, but they are only able to enter grape tissues through skin damages.
Several insects and pathogenic fungi are capable of producing mechanical damages to berries that are used by saprophytic ochratoxigenic species as a door to invade grape tissues [11]. Lobesia botrana is the major grape pest in vineyards of Southern European regions. Its larva is able to attack berries, damaging the skin and allowing for ochratoxigenic species to colonize and grow inside the grapes; therefore, OTA levels in damaged bunches have been found to be significantly higher [70]. L. botrana larvae usually trap fungal conidia in their cuticle ornamentations and facilitate fungal penetration since they effectively tunnel berries [71]. Insecticide treatments in the field, either chemical or biological, are able to reduce L. botrana infection, and, consequently, OTA levels in harvested grapes [72,73].
3.3. Weather
Different reports reflected important changes in OTA levels on grapes and wine in the same region between several years of harvest probably due to meteorogical circumstances [7]. For example, López de Cerain et al. [75] reported a significant variation in OTA incidence between two years in Spanish wines produced in a viticultural experimental station. They found an 85% of contaminated wines one year and only 15% the following one. The authors attributed it to the different quality of the grapes because of bad weather conditions that were reported during the first year, with low temperatures and heavy rains during the summer. These conditions favored the attack of pathogenic fungi, such as Oidium and Botrytis, and allowed for the entrance of ochratoxigenic fungi to grape tissues.
The effect of different meteorological parameters on OTA content of wines is controversial and it is not clearly established yet. Rotaru et al. [59] found a clear increase in positive samples depending on cultivation year, but they could not relate that difference to any weather parameter. Zhang et al. [47] found a negative correlation between OTA content and the average temperature from June to August, as well as a positive relationship between rainfall during grape veraison and OTA concentration in wines. Bellí et al. [13] did not study OTA contamination on wine but presence of black Aspergilli on grapes demonstrating that fungal occurrence was positively related to the average temperature although rainfall did not seem to have an influence in fungal infection. However, Lucchetta et al. [41] in a five-year long study performed in Italy described a negative relationship between ochratoxigenic species occurrence and relative humidity. The authors reported far more contaminated grapes in very dry years.
An extensive study performed in more than 150 vineyards in France established a correlation between OTA content in wine and meteorological characteristics in the vineyards provided that previous biotic damages had occurred in grape bunches [76]. These authors described that temperatures over 21 °C during the susceptible berry period (20 days before harvest) might result in critical levels of OTA concentration in wine.
Recent studies have been focused on how climate change would affect mycotoxin occurrence in foodstuffs. Climate change scenarios predicted higher temperatures and drier conditions that may favor A. tubingensis and A. niger development over A. carbonarius since they are better adapted to that new conditions [77].
3.4. Period of Harvest
It is well known that contamination by ochratoxigenic Aspergillus species is quite low in immature green grapes, since they are a hostile environment for spore germination [35]. From veraison to harvest, berries are most susceptible to infection by OTA-producing fungi due to their softer skin and higher sugar content, which favor Aspergillus spp. colonization and development [26]. Consequently, if harvest of mature berries is delayed, a high risk of OTA contamination will be found [78].
3.5. Agricultural and Harvesting Practices
Good agricultural and harvesting practices are a key point to minimize OTA occurrence in grapes and wine [79,80]. Even if environmental conditions are favoring fungal growth, the application of appropriate vineyard management can significantly reduce the final concentration of OTA in grapes [71]. The measures to reduce the incidence of Aspergillus spp. in mature berries include the minimization of damage to grapes during harvest and the removal of damaged or discolored berries, selection of grape varieties resistant to rain damage, or the application of control methods to reduce pests and other fungal diseases [35]. Minimizing Aspergillus spp. propagules in soil by careful management of cultivation, irrigation and pruning are also crucial tools to reduce OTA contamination of grapes [35].
The type of crop management applied in the vineyard does not seem to influence OTA contamination of grapes. Chiodini et al. [81] determined OTA content in organically and conventionally produced wines from different origins. Statistical analysis revealed that OTA levels in organic wines were not significantly different from that found in conventionally produced ones. The same results were found by Labrinea et al. [52] in conventional and organic wines from Greece.
3.6. Type of Wine
Table 1 gathers the most significant reports regarding the occurrence of OTA in wine worldwide. Regardless of geographical region, there is a clear tendency of red wines to be more contaminated than white varieties. Moreover, OTA usually occurs in sweet wines. Therefore, it seems that winemaking procedure is one of the most critical factors in the contamination of OTA in wines [6,7].
During white wine production, grapes are crushed and pressed, clear juice is separated from the skins and seeds, and then they rapidly undergo fermentation. However, red wines are left to macerate with the lees for several days allowing for higher amounts of OTA to be transferred to the juice [65,82]. Moreover, this maceration is usually performed at elevated temperature and aerobic conditions; therefore, no fermentation is occurring, which strongly favor fungal growth and OTA production [68].
Sweet wines are frequently contaminated and they often reach high levels of OTA that sometimes exceed the legal regulations [6,49,50,52,58]. This fact could also be related to winemaking techniques, although oenological practices vary widely among the different types of sweet wine [71]. For example, some sweet wines from Spain require a sunning process after harvesting when grapes lose water, and, consequently, increase their sugar content. These sun-dried grapes are exposed to an open environment for long periods (up to 15 days) when Aspergillus spp. become predominant over other fungi due to their high tolerance to sunlight and low water activity [36]. Some dessert wines are fortified by adding exogenous alcohol to the must or wine in order to stop fermentation [71]. During fermentation, OTA levels are considerably reduced (see Section 4); therefore, since this step is reduced or eliminated in fortified wines, OTA contamination is expected to be higher [83].
4. Fate of Ochratoxin A during Winemaking
A detailed protocol of winemaking process in white and red wines is shown in Figure 2, for a better understanding of this section. The complete winemaking procedure is known to reduce OTA content in wine by up to 80% [73]. The predicted increase or decrease in OTA content produced in each step is also indicated in Figure 2. As mentioned before, OTA contamination is due to ochratoxigenic fungal species that can grow and produce the toxin in grape tissues. The first step of winemaking requires grape crushing; the crushed grapes are transferred to the tanks where maceration takes place supposing the extraction of several compounds from the pomaces, including OTA [66]. Although OTA is also naturally present in the pulp, different studies report that berry skins are the most contaminated tissue of grapes; therefore, after crushing, OTA goes from the skin to must significantly increasing toxin content [82,84]. It is estimated that maceration causes an increase in OTA content around 20% [85]. From that step, oenological practices are quite different in white and red wines, which drastically modify OTA content.
In white vinification, pomace is removed before fermentation starts; therefore, pressing happens in absence of alcohol allowing for OTA to bind more effectively to grape solids [86]. However, in red wines, maceration occurs simultaneously to the first steps of alcoholic fermentation, which strongly affects OTA content. Lasram et al. [82] and Dachery et al. [87] observed an extremely high rise in OTA concentration one day after fermentation started (concentration around three and six times higher than OTA levels in grape juice, respectively).
Some winemaking procedures, usually for white wine production, need a flash pasteurization of must or the addition of sulfite at the beginning of the process that prevent fungal growth and OTA production [35].
Fermentation processes, both alcoholic and malolactic, significantly reduce OTA levels from must to wine. Csutorás et al. [88] studied OTA concentration during a 90-day long fermentation in macro-scale experiments using musts spiked with different amounts of OTA under similar conditions to real ripening processes. In all kind of wines (white, rose, and red), OTA content significantly decreased during the fermentation process regardless of initial toxin concentration.
OTA synthesis does not happen during fermentation since high ethanol concentration and generally anaerobic conditions inhibit fungal growth [7,85]. Moreover, it is well known that OTA removal from must during fermentation occurs mainly by adsorption to binding solids, such as grape skins and yeast cell walls [89]. In order to evaluate the adsorption ability of grape skins, Esti et al. [90] monitored OTA concentration in micro-fermentation experiments in presence or absence of grape skins. The results of these trials indicated that OTA removal was significantly higher (5–12% more reduction) in the presence of grape skins confirming the ability of these solids to adsorb OTA from musts.
Alcoholic fermentation is mainly performed by selected yeast species. OTA reduction during this period might be also attributed to adsorption to yeast cell walls. Caridi et al. [91] studied the ability of 20 Saccharomyces cerevisiae strains previously selected for winemaking to remove OTA during vinification. Their results showed that all of the tested strains noticeably reduced the initial OTA content, but there were significant differences in the behavior of each strain to remove the toxin in naturally contaminated musts. Cecchini et al. [92] observed a significant OTA reduction during alcoholic fermentation independently if it was produced by Saccharomyces or non-Saccharomyces species, although the percentage of OTA removal in wine varied widely (from 46% to 70%) depending on the yeast strain used. The strains of S. cerevisiae, Saccharomyces bayanus and Schizosaccharomyces pombe were the most efficient to remove OTA [92]. Meca et al. [93] studied the interaction between OTA and 16 S. cerevisiae strains during alcoholic fermentation of Moscato wine. The authors found that, although all of the strains were able to reduce OTA concentration to some extent, their detoxification ability varied widely (from 32% to 50%), probably due to the different composition of cell wall observed in these strains. Therefore, yeast selection during alcoholic fermentation offers an interesting option to maximize OTA removal from wines.
In the production of red wine, pomace pressing occurs after primary alcoholic fermentation. As the case of white wine, this pressing may reduce OTA levels since the toxin is adsorbed by the solids [86]; however, Gambuti et al. [78] observed that intensive pressing of the pomaces drastically increased OTA concentration in the wine, up to four times higher than conventionally produced ones.
Red wine production often requires a malolactic process that supposes a biological deacidification of wines. This fermentation is performed by lactic acid bacteria (LAB) and is also related to OTA decontamination [66]. Oenococcus oeni is the main LAB species that is involved in malolactic fermentation and some strains are reported to be able to remove OTA from extracts. In most cases, this removal appears to occur by a cell-binding process since toxin reduction is independent of bacterial viability and no degradation products are found [94,95]. However, some strains of Pediococcus parvulus isolated from Douro wines have been reported to degrade OTA to its less toxic derivative OTα. The synthesis of the enzyme responsible for biodegradation is inducible by OTA presence and the ability to synthesize it is strain specific [96]. Up to our knowledge, this is the only biochemical transformation of OTA that has been reported to occur during winemaking.
Many wine production processes require a long storage period in tanks before fining. Lasram et al. [82] demonstrated that OTA decreased around 50% after five months of storage. The authors attributed this OTA reduction to precipitation of solid compounds present in wine, such as pomace (in red wine) and debris of yeast and bacteria that could efficiently bound the toxin.
After fermentation, a clarification (fining) process is usually required to remove suspended solids that make wine cloudy. The clarification is widely known to contribute to OTA decrease due to adsorption to the sediment (lees) and could be natural sedimentation or performed by centrifugation or by the use of some adjuvants [66]. OTA bond to lees is quite stable and the toxin is not released even after centrifugation to recover additional wine [97]. García-Moruno et al. [98] demonstrated the ability of lees to bind OTA by adding lees from other vinification process to wine samples contaminated by the toxin. Only after 90 min of lees-wine contact, a significant reduction of OTA was detected and the maximum effect was observed after seven days of contact. Leong et al. [97] found significant variations in OTA content before and after clarification although the use of pectolytic enzymes did not seem to affect OTA removal.
Several fining agents used for wine clarification also contribute to detoxification by removing OTA. Some examples of fining adsorbents that have been reported to be highly efficient to remove OTA from contaminated wines include activated carbon [78,99,100], bentonite [97,101,102], gelatin [102,103], egg albumin [101,103], potassium caseinate [99], and naturals polymers, such as chitin and chitosan [102,104,105]. Despite their efficiency to detoxify, almost all of them modify organoleptic properties of wine even when they are used at the recommended dosage range. The concentration of the fining agent applied should be carefully controlled to be sure they do not affect flavor, color, and bouquet of the final product by removing polyphenols [106]. Changes in total polyphenol index lead to negative effects on wine quality. The loss of anthocyans supposes a reduction of color intensity, whereas the decrease of other sensory odorants affects wine aroma [107].
Nowadays, the EU allows the use of activated carbon for oenological use for treatment of white wines and musts, which might also reduce OTA concentration [108]. The International Code of Oenological Practices suggests the treatment using chitosan and chitin-glucan to reduce OTA in wine provided the doses do not exceed 500 g/hL [109,110].
Before bottling, a filtration is often performed in white wines to finish clarification process. Gambuti et al. [78] reported an 80% of OTA decrease after microfiltration through 0.45 µm membranes, probably due to toxin binding by the retained wine macromolecules above the filter.
Bottled ageing for at least 12 months seems to have an effect on the decrease of OTA content of wines, and this reduction increases over time if correct storage conditions are maintained [66,100]. This effect is lower in white wines probably due to a more complex matrix occurring in red wines that presents a greater potential for binding OTA [97].
Taking into account all the presented data, it can be concluded that OTA is mainly removed from musts through adsorption to solids, such as pomaces, grape skins, or the microorganisms that are involved in alcoholic or malolactic fermentation, which are discarded at the end of winemaking.
5. Worldwide Regulations of Ochratoxin A in Wine and Grape Juice
The FAO/WHO Joint Expert Committee on Food Additives (JECFA) has revised several times the maximum tolerable weekly intake value for OTA. Levels of OTA contamination of foodstuffs and potential intakes are considered by the Committee. After cereals, wine and grape juice are the most important sources of OTA in human diet. The contribution from wine is estimated to be 21% of the overall exposure (8.9 ng/kg bw per week) based on a daily consumption of 240 g, whereas the contribution from grape juice constitutes the 7% of the exposure (3.1 ng/kg bw per week) based on a daily consumption of 69 g [74,111].
Maximum levels for OTA in wine have been set by Commission Regulation (CE) No. 1881/2006, whereas the sampling methods have been regulated by Commission Regulation (CE) No. 401/2006 [8,112]. These EU standards were adopted based on the opinion of the Scientific Panel on Contaminants in Food of the European Food Safety Authority [113]. According to this regulation, the maximum tolerable levels of OTA are established at 2 µg/L in wine (including sparkling wine, aromatized wine, aromatized wine-based drinks, and aromatized wine-product cocktails), as well as in grape juice and derivatives that are intended for direct human consumption. This regulation is applied by all European Union Countries and other countries in the European region, including Albania, Macedonia, Norway, Ukraine, Turkey, Andorra, Azerbaijan, Iceland, Luxemburg, Bosnia-Herzegovina, Moldavia, Serbia, Georgia, Montenegro, Switzerland, Armenia, and Byelorussia [113].
Apart from those limits established in European Countries, few regulations are found worldwide regarding OTA in wine or grape juices [114]. Currently, maximum permitted levels of 10 µg/L and 2 µg/L have been established for OTA in wines in Brazil and Ecuador, respectively. Chile and Colombia have adopted regulations for OTA in grape juice and maximum limits of 5 and 2 µg/L have been set, respectively. In Asia, Indonesia regulates OTA levels in grape juice with maximum allowed levels of 2 µg/L, and, recently, China also established this maximum level of OTA in wine and grape juice [115].
6. Control and Detoxification Methods to Reduce OTA Content in Wine
To prevent OTA presence in wine and grape juice, it is essential to avoid grape contamination by toxigenic fungal species during pre and postharvest. The establishment of a correct Hazard Analysis and Critical Control Points (HACCP) system to implement good agricultural and manufactural practices might drastically reduce mycotoxins in the final product [65,79].
As discussed before, ochratoxigenic species infect grapes before harvest; therefore, controlling fungal proliferation in the field is a good option to minimize OTA contamination. Up to date, several active ingredients of fungicides frequently used in vineyards have demonstrated antifungal properties in vitro towards Aspergillus section Nigri species isolated from grapes, such as cyprodinil or sulfur [116]. On the contrary, some authorized products for their use in vineyards have been found to promote fungal growth and/or OTA production i.e., carbendazim [117] or copper [116]. Therefore, care must be taken before the application of chemical products and field trials are imperative. Tjamos et al. [118] evidenced the effectiveness of Switch (active ingredients cyprodinil and fludioxonil) in reducing the level of Aspergillus section Nigri populations on wine-producing grape vineyards and indicated that the closer the harvest the more efficient the application. Bellí et al. [119] compared the effectiveness of Switch and Chorus (active ingredient cyprodinil) to control black Aspergilli in the vineyards. The authors observed a much higher efficiency of Switch and suggested a possible synergistic effect of cyprodinil and fludioxonil against A. carbonarius in the field.
In the last few years, a strict regulation for the use of chemical compounds has been established since negative effect towards environment and human health of these compounds has been reported. Therefore, there is a need to investigate sustainable methods that effectively control fungal growth, and, subsequently, OTA production in grapes. Some natural fungicides have been tested in vitro against black Aspergilli isolated from grapes. Perillaldehyde and natamycin showed antifungal activity in vitro and reduced OTA production by black Aspergilli, but their effectiveness has not been tested on grapes yet [120,121]. Favilla et al. [122] tested the effectiveness of fusopyrone on artificially contaminated grapes in highly conductive conditions to mold development and observed an almost complete inhibition of growth and a drastic reduction on OTA content of grapes.
Biological control using antagonistic microorganism is also considered an ecofriendly alternative method to substitute or complement chemical treatments on grapes [123]. There are many reports that show the efficiency of saprophytic yeast to control ochratoxigenic Aspergillus spp. in artificially inoculated grapes. The potential biocontrol agents described up to now include strains of Aureobasidium pullulans, Candida incommunis, Candida intermedia, Candida friedrichii, Candida sake, Cryptococcus magnus, Issatchenkia terricola, Issatchenkia orientalis, Kluyveromyces thermotolerans, Lanchacea thermotolerans, and Metschnikowia pulcherrima [124,125,126,127].
Aureobasidium pullulans Y-1 isolated from the grapevine phyllosphere is effective in controlling A. carbonarius infection of berries in the field, and, therefore, OTA contamination in the must [128]. The application of this yeast in vineyards is as effective as the fungicide Switch to reduce fungal growth [128]. Ponsone et al. [129] reported the effective control of black Aspergilli and OTA contamination of wine grapes by two strains of the epiphytic yeast L. thermotolerans under both greenhouse and field trials. The authors indicated that this effect was significantly higher when mechanical damage was applied, suggesting that the yeasts rapidly colonized the wounds preventing Aspergillus spp. infection. Despite the promising results of biopesticides in the vineyards, their effectiveness is very depending on environmental factors and their protection decreases when temperature and humidity are optimum for fungal development [130]. Therefore, it is necessary to found biocontrol agents, which can efficiently compete at environmental conditions more conductive to infection by Aspergillus spp.
Taking into account that ochratoxigenic species enter grape tissues through skin lesions, a carefully performed harvest, and transport to minimize grape damage, as well as cool storage of harvested grapes to prevent fungal growth might be crucial to reduce OTA contamination [78]. OTA presence might also be reduced up to 98% by the rejection of moldy grapes or bunches before entering the winemaking process [106]. Minimizing the delay between harvest and crushing is also a good measure to prevent toxin contamination of unprocessed grapes [74].
Solfrizzo et al. [131] proposed a novel decontamination procedure using grape pomaces to efficiently reduce remaining OTA after fermentation without affecting oenological parameters. The authors proposed the repassage of contaminated wines over no contaminated grape pomaces from the same variety. The pomaces showed high affinity for OTA even after being used four times. The effect was found after only 24 h contact and OTA content in wines was found to decrease by 55–65%.
Many yeast have been reported as detoxifying agents due to OTA adsorption to their cell walls or enzymatic degradation. Adsorption is a passive phenomenon, which supposes OTA link to yeast cell wall components, such as α-d-glucans and mannoproteins, and does not depend on cell viability [123]. Therefore, detoxification based on microbial adsorption might be a clear advantage because it can be performed by autoclaved yeast cells that cannot neither consume substrate components nor release secondary metabolites, which would lead to an alteration of organoleptic properties of the product [132]. Several inactivated yeast or their cell wall preparations have shown their ability to adsorb OTA in grape juice or wine, including strains of S. cerevisiae, S. bayanus, C. intermedia, C. fiedrichii, and L. thermotolerans [126,133,134]. Despite the good results obtained using biological decontamination, the addition of yeasts to wine needs to be controlled and a careful selection of suitable yeast should be performed. Although a high OTA adsorption ability of yeast cell walls and inactivated cells of S. cerevisiae W13 and BM45, wine treatment with these agents caused color loss of wines [134].
On the other hand, a major constraint in microbiological detoxification is the removal of decontaminating agents after treatment. Farbo et al. [132] proposed alginate beads encapsulation of Candida intermedia dead cells to be applied in grape juice detoxification. The maximum reduction (80%) was found after 48 h of incubation and the adsorption efficiency was similar when cells were free or immobilized.
7. Analytical Methods for OTA Determination in Wine
Simple, reliable, and sensitive methods to monitor ochratoxin levels are indispensable to guarantee food safety and to demonstrate that wines accomplish legal regulations [9]. The European Commission established some requirements for toxin analysis methods regarding their recovery and reproducibility [112] and recommended a protocol for OTA determination in wines by inmunoaffinity columns (IAC) clean-up, followed by HPLC coupled with fluorescent detection [56,135].
Mycotoxin analysis usually consist of three steps (1) extraction from the matrix, (2) purification from the extract (clean-up) and concentration if necessary, and (3) detection and quantification by analytical instruments [9]. Up to now, many suitable protocols are available to determine OTA in wine that differ in some of these steps, usually in extraction or clean up procedures. The main protocols that are described up to now together with their features are indicated in Table 2.
In the last few years, novel technologies have been proposed for rapid analysis of OTA in food products to replace the expensive IAC clean-up. As can be seen in Table 2, different clean-up methods have been developed based on solid-phase extractions with peptides, molecular imprinted polymers, or aptamers that are able to bind OTA as efficiently as IAC. On the other hand, chromatography based methods for detection and quantification of the toxin require a highly equipped laboratory and qualified staff, and, therefore, the current research is focused on the development of new detection methods being immunosensors and fluorescence polarization immunoassays, which have yielded the best results (Table 2).
Different, quite inexpensive, fast, and sensitive innovative methods to determine OTA have been also developed in the last few years. Barthelmebs et al. [147] developed a direct competitive enzyme-linked aptamer assay (ELAA) successfully applied for the detection of very low amounts of OTA in wine samples (detection limit 1 µg/L). The authors propose this method to perform a useful screening for routine use in OTA determination in wine since it is rapid (125 min) and overcome the disadvantages of the unstable and expensive ELISA-based methods. Yin et al. [148] have recently designed a novel aptamer-based colorimetric bioassay to determine OTA in fortified white wine samples. Free aptamers are absorbed onto gold nanoparticles surfaces, but when aptamers recognize OTA, gold particles aggregate. This aggregation supposes a change from red to blue color that is easily observed by naked eye or measured by a spectrophotometer. These new biosensors need to be improved to get better detection limits, but they are promising tools for rapid detection of OTA in wine samples.
Rapid tests for OTA screening in wineries are of value to evaluate OTA content in situ and check if samples fulfil legislation requirements. To eliminate matrix interferences, it is necessary to perform a rapid clean up protocol that could be performed in a quite simply equipped laboratory. Different tests are available that indicate if OTA presence is under or above legal limits established for wine in the UE (2 µg/L). These rapid methods are based on competitive enzyme immunoassays on nitrocellulose membranes. Anti-OTA immunoglobulins are linked to an enzyme that allows for a chromogenic detection when toxin is above the corresponding limits. Several tests are available from different manufacturers i.e., Ochracard (R-Biopharm), OCHRATOXIN-A (wine) rapid test (BioSystems) or Ochratoxin A in wine flow-through rapid test (EuroProxima).
8. Other Mycotoxins in Wine
Although OTA is by far the most frequently found mycotoxin in wine, an increasing number of systematic studies have reported contamination by some of its derivatives. However, only a few studies have been focused on the importance of OTA metabolites regarding their toxicity and occurrence [2]. Some surveys that were performed in Mediterranean countries show that OTA appears simultaneously with OTB in all the wine samples and OTC is also an important analog due to its high occurrence at high levels [149,150]. OTC is hydrolyzed to OTA in red wine, and, therefore, OTA levels may be underestimated and it would be necessary to include multitoxin detection of OTA and its analogs in routine analysis of wine [149]. Other analogs, such as metil-OTA, metil-OTB, and etil-OTB have been also frequently found in Mediterranean wine [69]. The possible synergic or additive effects posed by these toxins need to be evaluated to determine the potential risk for human and animal health.
9. Conclusions
OTA is a widespread mycotoxin frequently found in wine worldwide. The source of OTA in this product is ochratoxigenic Aspergillus species that grow and produce the toxin in grapes. Although OTA levels drastically decrease during winemaking, control, and detoxification methods are necessary to avoid OTA presence in the final product. Climate change scenarios predict a modification in fungal species distribution, as well as in ochratoxin occurrence in wine. Therefore, more studies on the occurrence of OTA derivatives are necessary to unravel their importance in wine worldwide.
Acknowledgments
This work was supported by the Spanish Ministry of Science and Innovation (AGL 2014-53928-C2-2-R).
Author Contributions
All authors contributed equally to this work.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gil-Serna, J.; Vázquez, C.; Sardiñas, N.; González-Jaén, M.T.; Patiño, B. Revision of ochratoxin A production capacity by the main species of Aspergillus section Circumdati. Aspergillus steynii revealed as the main risk of OTA contamination. Food Control 2011, 22, 343–345. [Google Scholar] [CrossRef]
- El Khoury, A.; Atoui, A. Ochratoxin A: General overview and actual molecular status. Toxins 2010, 2, 461–493. [Google Scholar] [CrossRef] [PubMed]
- Palumbo, J.D.; O’Keeffe, T.L. Detection and discrimination of four Aspergillus section Nigri species by PCR. Lett. Appl. Microbiol. 2015, 60, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Gil-Serna, J.; Vázquez, C.; González-Jaén, M.T.; Patiño, B. Mycotoxins: Toxicology. In Encyclopedia of Food Microbiology, 2nd ed.; Batt, C., Tortorello, M.L., Eds.; Elsevier Ltd. Academic Press: Amsterdam, The Netherlands, 2014; pp. 887–892. ISBN 978-0-12-384733-1. [Google Scholar]
- International Agency for Research on Cancer (IARC). Some naturally occurring substances: Food items and constituents, heterocyclic aromatic amines and mycotoxins. IARC Monogr. Eval. Carcinog. Risks Hum. 1993, 56, 245–395. [Google Scholar]
- Zimmerli, B.; Dick, R. Ochratoxin A in table wine and grape juice: Occurrence and risk assessment. Food Addit. Contam. 1996, 13, 655–668. [Google Scholar] [CrossRef] [PubMed]
- Blesa, J.; Soriano, J.M.; Moltó, J.C.; Mañes, J. Factors affecting the presence of ochratoxin A in wines. Crit. Rev. Food Sci. 2006, 46, 473–478. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Commission Regulation EC No 1881/2006 setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Union 2006, 364, 5–24. [Google Scholar]
- Anli, R.E.; Bayram, M. Ochratoxin A in wines. Food Rev. Int. 2009, 25, 214–232. [Google Scholar] [CrossRef]
- Cabañes, F.J.; Accensi, F.; Bragulat, M.R.; Abarca, M.L.; Castellá, G.; Minguez, S.; Pons, A. What is the source of ochratoxin A in wine? Int. J. Food Microbiol. 2002, 79, 213–215. [Google Scholar] [CrossRef]
- Varga, J.; Kozakiewicz, Z. Ochratoxin A in grapes and grape derived products. Trends Food Sci. Technol. 2006, 17, 72–81. [Google Scholar] [CrossRef]
- Bau, M.; Bragulat, M.R.; Abarca, M.L.; Minguez, S.; Cabañes, F.J. Ochratoxigenic species from Spanish wine grapes. Int. J. Food Microbiol. 2005, 98, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Bellí, N.; Bau, M.; Marín, S.; Abarca, M.L.; Ramos, A.J.; Bragulat, R. Mycobiota and ochratoxin A producing fungi from Spanish wine grapes. Int. J. Food Microbiol. 2006, 111, S40–S45. [Google Scholar] [CrossRef] [PubMed]
- García-Cela, E.; Crespo-Sempere, A.; Gil-Serna, J.; Porqueres, A.; Marín, S. Fungal diversity, incidence and mycotoxin contamination in grapes from two agro-climatic Spanish regions with emphasis on Aspergillus species. J. Sci. Food Agric. 2014, 95, 1716–1729. [Google Scholar] [CrossRef] [PubMed]
- Serra, R.; Abrunhosa, L.; Kozakiewicz, Z.; Venancio, A. Black Aspergillus species as ochratoxin A producers in Portuguese wine grapes. Int. J. Food Microbiol. 2003, 88, 63–68. [Google Scholar] [CrossRef]
- Serra, R.; Mendoça, C.; Venancio, A. Ochratoxin A occurrence and formation in Portuguese wine grapes at various stages of maturation. Int. J. Food Microbiol. 2006, 111, S35–S39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bejaoui, H.; Mathieu, F.; Taillandier, P.; Lebrihi, A. Black Aspergilli and ochratoxin A production in French vineyards. Int. J. Food Microbiol. 2006, 111, S46–S52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dachoupakan, C.; Ratomahenina, R.; Martinez, V.; Guiraud, J.P.; Baccou, J.C.; Schorr-Galindo, S. Study of the phenotypic and genotypic biodiversity of potentially ochratoxigenic black aspergilli isolated from grapes. Int. J. Food Microbiol. 2009, 132, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Battilani, P.; Pietri, A.; Bertuzzi, T.; Languasco, L.; Giorni, P.; Kozakiewicz, Z. Occurrence of ochratoxin A—Producing fungi in grapes grown in Italy. J. Food Prot. 2003, 66, 633–636. [Google Scholar] [CrossRef] [PubMed]
- Battilani, P.; Giorni, P.; Bertuzzi, T.; Formenti, S.; Pietri, A. Black Aspergilli and ochratoxin A in grapes in Italy. Int. J. Food Microbiol. 2006, 111, S53–S60. [Google Scholar] [CrossRef] [PubMed]
- Tjamos, S.E.; Antoniou, P.P.; Tjamos, E.C. Aspergillus spp., distribution, population composition and ochratoxin A production in wine producing vineyards in Greece. Int. J. Food Microbiol. 2006, 111, S61–S66. [Google Scholar] [CrossRef] [PubMed]
- Lasram, S.; Oueslati, S.; Mliki, A.; Ghorbel, A.; Silar, P.; Chebil, S. Ochratoxin A and ochratoxigenic black Aspergillus species in Tunisian grapes cultivated in different geographic areas. Food Control 2012, 25, 75–80. [Google Scholar] [CrossRef]
- Fredj, S.M.; Chebil, S.; Mliki, A. Isolation and characterization of ochratoxin A and aflatoxin B1 producing fungi infecting grapevines cultivated in Tunisia. Afr. J. Microbiol. Res. 2009, 3, 523–527. [Google Scholar]
- El Khoury, A.; Rizk, T.; Lteif, R.; Azouri, H.; Delia, M.L.; Lebrihi, A. Fungal contamination and aflatoxin B1 and ochratoxin A in Lebanese wine-grapes and musts. Food Chem. Toxicol. 2008, 46, 2244–2250. [Google Scholar] [CrossRef] [PubMed]
- Chulze, S.N.; Magnoli, C.E.; Dalcero, A.M. Occurrence of ochratoxin A in wine and ochratoxigenic mycoflora in grapes and dried vine fruits in South America. Int. J. Food Microbiol. 2006, 111, S5–S9. [Google Scholar] [CrossRef] [PubMed]
- Leong, S.L.; Hocking, A.D.; Pitt, J.I.; Kazi, B.A.; Emmett, R.W.; Scott, E.S. Australian research on ochratoxigenic fungi and ochratoxin A. Int. J. Food Microbiol. 2006, 111, S10–S17. [Google Scholar] [CrossRef] [PubMed]
- Da Rocha, C.A.; Palacios, V.; Combina, M.; Fraga, M.E.; De Oliveira, A.; Magnoli, C.E.; Dalcero, A.M. Potential ochratoxin A producers from wine grapes in Argentina and Brazil. Food Addit. Contam. 2002, 19, 408–414. [Google Scholar] [CrossRef]
- Magnoli, C.; Violante, M.; Combina, M.; Palacio, G.; Dalcero, A. Mycoflora and ochratoxin-producing strains of Aspergillus section Nigri in wine grapes in Argentina. Lett. Appl. Microbiol. 2003, 37, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Chiotta, M.L.; Ponsone, M.L.; Combina, M.; Torres, A.M.; Chulze, S.N. Aspergillus section Nigri species isolated from different wine-grape growing regions in Argentina. Int. J. Food Microbiol. 2009, 136, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Pantelides, I.S.; Aristeidou, E.; Lazari, M.; Tsolakidou, M.D.; Tsaltas, D.; Christofidou, M.; Kafouris, D.; Christou, E.; Ioannou, N. Biodiversity and ochratoxin A profile of Aspergillus section Nigri populations isolated from wine grapes in Cyprus vineyards. Food Microbiol. 2017, 67, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Medina, A.; Mateo, R.; López-Ocaña, L.; Valle-Algarra, F.M.; Jiménez, M. Study of Spanish grape mycobiota and ochratoxin A production by isolates of Aspergillus tubingensis and other members of Aspergillus section Nigri. Appl. Environ. Microbiol. 2006, 71, 4696–4702. [Google Scholar] [CrossRef] [PubMed]
- Garmendia, G.; Vero, S. Occurrence and biodiversity of Aspergillus section Nigri on ‘Tannat’ grapes in Uruguay. Int. J. Food Microbiol. 2016, 216, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Qi, T.F.; Renaud, J.B.; McDowell, T.; Seifert, K.A.; Yeung, K.K.C.; Sumarah, M.W. Diversity of Mycotoxin-Producing Black Aspergilli in Canadian Vineyards. J. Agric. Food Chem. 2016, 64, 1583–1589. [Google Scholar] [CrossRef] [PubMed]
- Perrone, G.; Varga, J.; Susca, A.; Frisvad, J.C.; Stea, G.; Kocsube, S.; Kozakiewicz, Z.; Samson, R.A. Aspergillus uvarum sp. nov., an uniseriate black Aspergillus species isolated from grapes in Europe. Int. J. Syst. Evol. Microbiol. 2007, 58, 1032–1039. [Google Scholar] [CrossRef] [PubMed]
- Hocking, A.D.; Leong, S.L.; Kazi, B.A.; Emmett, R.W.; Scott, E.S. Fungi and mycotoxins in vineyards and grape products. Int. J. Food Microbiol. 2007, 119, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Valero, A.; Begum, M.; Leong, S.L.; Hocking, A.D.; Ramos, A.J.; Sanchis, V.; Marín, S. Effect of germicidal UVC light on fungi isolated from grapes and raisins. Lett. Appl. Microbiol. 2007, 45, 238–243. [Google Scholar] [CrossRef] [PubMed]
- Bellí, N.; Marín, S.; Sanchis, V.; Ramos, A.J. Influence of water activity and temperature on growth of isolates of Aspergillus section Nigri obtained from grapes. Int. J. Food Microbiol. 2004, 96, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.; Parra, R.; Aldred, D.; Magan, N. Water and temperature relations of growth and ochratoxin A production by Aspergillus carbonarius strains from grapes in Europe and Israel. J. Appl. Microbiol. 2004, 97, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Leong, S.L.; Hocking, A.D.; Scott, E.S. Effect of temperature and water activity on growth and ochratoxin A production by Australian Aspergillus carbonarius and A. niger isolates on a simulated grape juice medium. Int. J. Food Microbiol. 2006, 110, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Marín, S.; Bellí, N.; Lasram, S.; Chebil, S.; Ramos, A.J.; Ghorbel, A.; Sanchis, V. Kinetics of ochratoxin A production and accumulation by Aspergillus carbonarius on synthetic grape medium at different temperature levels. J. Food Sci. 2006, 71, M196–M200. [Google Scholar] [CrossRef]
- Lucchetta, G.; Bazzo, I.; Dal Cortivo, G.; Stringher, L.; Bellotto, D.; Borgo, M.; Angelini, E. Occurrence of black Aspergilli and ochratoxin A on grapes in Italy. Toxins 2010, 2, 840–855. [Google Scholar] [CrossRef] [PubMed]
- Ponsone, M.L.; Chiotta, M.L.; Combina, M.; Torres, A.; Knass, P.; Dalcero, A.; Chulze, S.N. Natural occurrence of ochratoxin A in musts, wines and grape vine fruits from grapes harvested in Argentina. Toxins 2010, 2, 1984–1996. [Google Scholar] [CrossRef] [PubMed]
- Hocking, A.D.; Varelis, P.; Pitt, J.I.; Cameron, S.F.; Leong, S.L. Occurrence of ochratoxin A in Australian wine. Aust. J. Grape Wine Res. 2003, 9, 72–78. [Google Scholar] [CrossRef]
- Ng, W.; Mankotia, M.; Pantazopoulos, P.; Neil, R.J.; Scott, P.M. Ochratoxin A in wine and grape juice sold in Canada. Food Addit. Contam. 2004, 21, 971–981. [Google Scholar] [CrossRef] [PubMed]
- Vega, M.; Ríos, G.; Baer, D.; Mardones, C.; Tessini, C.; Herlitz, E.; Saelzer, R.; Ruiz, M.A. Ochratoxin A occurrence in wines produced in Chile. Food Control 2012, 28, 147–150. [Google Scholar] [CrossRef]
- Wu, J.; Tan, Y.; Wang, Y.; Xu, R. Occurrence of ochratoxin A in wine and beer samples from China. Food Addit. Contam. B 2011, 4, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Chen, L.; Li, J.; Zhu, B.; Ma, L. Occurrence of ochratoxin A in Chinese wines: Influence of local meteorological parameters. Eur. Food Res. Technol. 2013, 236, 277–283. [Google Scholar] [CrossRef]
- Mikulikova, R.; Belakova, S.; Benesoba, K.; Svoboda, Z. Study of ochratoxin A content in South Moravian and foreign wines by the UPLC method with fluorescence detection. Food Chem. 2012, 133, 55–59. [Google Scholar] [CrossRef]
- Burdaspal, P.; Legarda, T. Occurrence of ochratoxin A in sweet wines produced in Spain and other countries. Food Addit. Contam. 2007, 24, 976–986. [Google Scholar] [CrossRef] [PubMed]
- Stefanaki, I.; Foufa, E.; Tsatsou-Dritsa, A.; Dais, P. Ochratoxin A concentrations in Greek domestic wines and dried vine fruits. Food Addit. Contam. 2003, 20, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Soufleros, E.H.; Tricard, C.; Bouloumpasi, E.C. Occurrence of ochratoxin A in Greek wines. J. Sci. Food Agric. 2003, 83, 173–179. [Google Scholar] [CrossRef]
- Labrinea, E.P.; Natskoulis, P.I.; Spiropoulos, A.E.; Magan, N.; Tassou, C.C. A survey of ochratoxin A occurrence in Greek wines. Food Addit. Contam. B 2011, 4, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Varga, J.; Kiss, R.; Mátrai, T.; Mátrai, T.; Téren, J. Detection of ochratoxin A in Hungarian wines and beers. Acta Aliment. 2005, 34, 381–392. [Google Scholar] [CrossRef]
- Pietri, A.; Bertuzzi, T.; Pallaroni, L.; Piva, G. Occurrence of ochratoxin A in Italian wines. Food Addit. Contam. 2001, 18, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Spadaro, D.; Lorè, A.; Garibaldi, A.; Gullino, M.L. Occurrence of ochratoxin A before bottling in DOC and DOCG wines produced in Piedmont (Northern Italy). Food Control 2010, 21, 1294–1297. [Google Scholar] [CrossRef]
- Visconti, A.; Pascale, M.; Centonze, G. Determination of ochratoxin A in wine by means of immunoaffinity column clean-up and high-performance liquid chromatography. J. Chromatogr. A 1999, 864, 89–101. [Google Scholar] [CrossRef]
- Brera, C.; Debegnach, F.; Minardi, V.; Prantera, E.; Pannunzi, E.; Faleo, S.; De Santis, B.; Miraglia, M. Ochratoxin A contamination in Italian wine samples and evaluation of the exposure in the Italian population. J. Agric. Food Chem. 2008, 56, 10611–10618. [Google Scholar] [CrossRef] [PubMed]
- Di Stefano, V.; Pitonzo, R.; Avellone, G.; Di Fiore, A.; Monte, L.; Ogorka, A.Z. Determination of aflatoxins and ochratoxins in Sicilian Sweet Wines by High-Performance Liquid Chromatography with Fluorometric Detection and Immunoaffinity Cleanup. Food Anal. Method 2015, 8, 569–577. [Google Scholar] [CrossRef]
- Rotaru, S.; Israel-Roming, F.; Campeanu, G.; Deciu, G. Correlation of ochratoxin A level in wine with vine environment. Rom. Biotechnol. Lett. 2011, 16, 126–130. [Google Scholar]
- Bellí, N.; Marín, S.; Duaigües, A.; Ramos, A.J.; Sanchis, V. Ochratoxin A in wines, musts and grape juices from Spain. J. Sci. Food Agric. 2004, 84, 591–594. [Google Scholar] [CrossRef]
- Quintela, S.; Villarán, M.C.; López de Armentia, I.; Elejalde, E. Occurrence of ochratoxin A in Rioja Alavesa wines. Food Chem. 2011, 126, 302–305. [Google Scholar] [CrossRef]
- Murillo-Arbizu, M.T.; Amézqueta, S.; González-Peñas, E.; López de Cerain, A. Occurrence of Ochratoxin A in Southern Spanish Generous Wines under the Denomination of Origin “Jerez-Xérès-Sherry and Manzanilla Sanlúcar de Barrameda”. Toxins 2010, 2, 1054–1064. [Google Scholar] [CrossRef] [PubMed]
- Shepard, G.S.; Fabiani, A.; Stockenstro, S.; Mschicileli, N.; Sewram, V. Quantitation of ochratoxin A in South African wines. J. Agric. Food Chem. 2003, 51, 1102–1106. [Google Scholar] [CrossRef] [PubMed]
- Stander, M.A.; Steyn, P.S. Survey of ochratoxin A in South African wines. S. Afr. J. Enol. Vitic. 2002, 23, 9–13. [Google Scholar] [CrossRef]
- Var, I.; Kabak, B. Occurrence of ochratoxin A in Turkish wines. Microchem. J. 2007, 86, 241–247. [Google Scholar] [CrossRef]
- Grazioli, B.; Fumi, M.D.; Silva, A. The role of processing on ochratoxin A content in Italian must and wine: A study on naturally contaminated grapes. Int. J. Food Microbiol. 2006, 111, S93–S96. [Google Scholar] [CrossRef] [PubMed]
- Miraglia, M.; Brera, C. Task 3.2.7. Assessment of Dietary Intake of Ochratoxin A by the Population of EU Member States. 2002. Available online: https://ec.europa.eu/food/sites/food/files/safety/docs/cs_contaminants_catalogue_ochratoxin_task_3-2-7_en.pdf (accessed on 25 October 2017).
- Otteneder, H.; Majerus, P. Occurrence of ochratoxin A (OTA) in wines: Influence of the type of wine and its geographical origin. Food Addit. Contam. 2000, 17, 793–798. [Google Scholar] [CrossRef] [PubMed]
- Remiro, R.; Irigoyen, A.; González-Peñas, E.; Lizarraga, E.; López de Cerain, A. Levels of ochratoxin in Mediterranean red wines. Food Control 2013, 32, 63–68. [Google Scholar] [CrossRef]
- Cozzi, G.; Pascale, M.; Perrone, G.; Visconti, A.; Logrieco, A. Effect of Lobesia botrana damages on black aspergilli rot and ochratoxin A content in grapes. Int. J. Food Microbiol. 2006, 111, S88–S92. [Google Scholar] [CrossRef] [PubMed]
- Covarelli, L.; Beccari, G.; Marini, A.; Tosi, L. A review on the occurrence and control of ochratoxigenic fungal species and ochratoxin A in dehydrated grapes, non-fortified dessert wines and dried vine fruit in the Mediterranean area. Food Control 2012, 26, 347–356. [Google Scholar] [CrossRef]
- Cozzi, G.; Haidukowski, M.; Perrone, G.; Visconti, A.; Logrieco, A. Influence of Lobesia botrana field control on black aspergilli rot and ochratoxin A contamination in grapes. J. Food Prot. 2009, 72, 894–897. [Google Scholar] [CrossRef] [PubMed]
- Cozzi, G.; Somma, S.; Haidukowski, M.; Logrieco, A. Ochratoxin A management in vineyards by Lobesia botrana biocontrol. Toxins 2013, 5, 49–59. [Google Scholar] [CrossRef] [PubMed]
- The Joint FAO/WHO Expert Committee on Food Additives (JECFA). Safety Evaluation of Certain Food Additives and Contaminants. Ochratoxin A. WHO Food Additives Series 2008, No. 59. Available online: http://apps.who.int/iris/bitstream/ 10665/43823/1/9789241660594_eng.pdf (accessed on 25 October 2017).
- López de Cerain, A.; González-Peñas, E.; Jiménez, A.M.; Bello, J. Contribution to the study of ochratoxin A in Spanish wines. Food Addit. Contam. 2002, 19, 1058–1064. [Google Scholar] [CrossRef] [PubMed]
- Clouvel, P.; Bonvarlet, L.; Martínez, A.; Lagouarde, P.; Dieng, I.; Martin, P. Wine contamination by ochratoxin A in relation to vine environment. Int. J. Food Microbiol. 2008, 123, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Paterson, M.; Venancio, A.; Lima, N.; Guilloux-Bénatier, M.; Rousseaux, S. Predominant mycotoxins, mycotoxigenic fungi and climate change related to wine. Food Res. Int. 2017, in press. [Google Scholar] [CrossRef]
- Gambuti, A.; Strollo, D.; Genovese, A.; Ugliano, M.; Ritieni, A.; Moio, L. Influence of enological practices on ochratoxin A concentration in wine. Am. J. Enol. Vitic. 2005, 56, 155–162. [Google Scholar]
- Food and Agriculture Organization (FAO). Manual on the Application of the HACCP System in Mycotoxin Prevention and Control; FAO Food and Nutrition Paper; Food and Agriculture Organization: Roma, Italy, 2001; Volume 73, pp. 1–124. Available online: http://www.fao.org/3/a-y1390e.pdf (accessed on 25 October 2017).
- International Organization of Vine and Wine (OIV). Resolution VITI/OENO 1/2005. Code of Sound Vitivinicultural Practices in Order to Minimise Levels of Ochratoxin A in Vine-Based Products. 2005. Available online: http://www.oiv.int/public/medias/2771/viti-oeno-1-2005-en.pdf (accessed on 25 October 2017).
- Chiodini, A.M.; Scherpenisse, P.; Bergwerff, A.A. Ochratoxin A contents in wine: Comparison of organically and conventionally produced products. J. Agric. Food Chem. 2006, 54, 7399–7404. [Google Scholar] [CrossRef] [PubMed]
- Lasram, S.; Mani, A.; Zaied, C.; Chebil, S.; Abid, S.; Bacha, H.; Mliki, A.; Ghorbel, A. Evolution of ochratoxin A content during red and rose vinification. J. Sci. Food Agric. 2008, 88, 1696–1703. [Google Scholar] [CrossRef]
- Bellver, J.; Fernández-Franzón, M.; Ruiz, M.J.; Juan-García, A. Presence of ochratoxin A (OTA) mycotoxin in alcoholic drinks from Southern European countries: Wine and beer. J. Agric. Food Chem. 2014, 62, 7643–7651. [Google Scholar] [CrossRef] [PubMed]
- Battilani, P.; Pietri, A. Ochratoxin A in grapes and wine. Eur. J. Plant Pathol. 2002, 108, 639–643. [Google Scholar] [CrossRef]
- Battilani, P.; Magan, N.; Logrieco, A. European research on ochratoxin A in grapes and wine. Int. J. Food Microbiol. 2006, 11, S2–S4. [Google Scholar] [CrossRef] [PubMed]
- Leong, S.L.; Hocking, A.D.; Varelis, P.; Giannikopoulos, G.; Scott, E.S. Fate of ochratoxin A during vinification of Semillon and Shiraz grapes. J. Agric. Food Chem. 2006, 54, 6460–6464. [Google Scholar] [CrossRef] [PubMed]
- Dachery, B.; Fonseca, F.; Dal Magro, L.; Manfroi, V.; Welke, J.E. Exposure risk assessment to ochratoxin A through consumption of juice and wine considering the effect of steam extraction time and vinification stages. Food Chem. Toxicol. 2017, 109, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Csutorás, C.; Rácz, L.; Rácz, K.; Futo, P.; Forgó, P.; Kiss, A. Monitoring ochratoxin A during fermentation of different wines by applying high toxin concentrations. Microchem. J. 2013, 107, 182–184. [Google Scholar] [CrossRef]
- Fernandes, A.; Ratola, N.; Cerdeira, A.; Alves, A.; Venancio, A. Changes in ochratoxin A concentration during winemaking. Am. J. Enol. Vitic. 2007, 58, 92–96. [Google Scholar]
- Esti, M.; Benucci, I.; Liburdi, K.; Acciaro, G. Monitoring of ochratoxin A fate during alcoholic fermentation of wine-must. Food Control 2012, 27, 53–56. [Google Scholar] [CrossRef]
- Caridi, A.; Galvano, F.; Tafuri, A.; Ritieni, A. Ochratoxin A removal during winemaking. Enzyme Microb. Technol. 2006, 40, 122–126. [Google Scholar] [CrossRef]
- Cecchini, F.; Morassut, M.; García-Moruno, E.; Di Stefano, R. Influence of yeast strain on ochratoxin A content during fermentation of white and red must. Food Microbiol. 2006, 23, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Meca, G.; Blaiotta, G.; Ritieni, A. Reduction of ochratoxin A during the fermentation of Italian red wine Moscato. Food Control 2010, 21, 579–583. [Google Scholar] [CrossRef]
- Del Prete, V.; Rodríguez, H.; Carrascosa, A.V.; De las Rivas, B.; García-Moruno, E.; Muñoz, R. In vitro removal of ochratoxin A by wine lactic acid bacteria. J. Food Prot. 2007, 70, 2155–2160. [Google Scholar] [CrossRef] [PubMed]
- Mateo, E.M.; Medina, A.; Mateo, F.; Valle-Algarra, F.M.; Pardo, I.; Jiménez, M. Ochratoxin A removal in synthetic media by living and heat-inactivated cells of Oenococcus oeni isolated from wines. Food Control 2010, 21, 23–28. [Google Scholar] [CrossRef] [Green Version]
- Abrunhosa, L.; Ines, A.; Rodrigues, A.I.; Guimaraes, A.; Pereira, V.L.; Parpot, P.; Mendes-Faia, A.; Venancio, A. Biodegradation of ochratoxin A by Pediococcus parvulus isolated from Douro wines. Int. J. Food Microbiol. 2014, 188, 45–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leong, S.L.; Hocking, A.D.; Scott, E.S. The effect of juice clarification, static or rotary fermentation and fining on ochratoxin A in wine. Aust. J. Grape Wine Res. 2006, 12, 245–251. [Google Scholar] [CrossRef]
- García-Moruno, E.; Sanlorenzo, C.; Boccaccino, B.; Di Stefano, R. Treatment with yeast to reduce the concentration of ochratoxin A in red wine. Am. J. Enol. Vitic. 2005, 56, 73–76. [Google Scholar]
- Castellari, M.; Versari, A.; Fabiani, A.; Parpinello, G.P.; Galassi, S. Removal of ochratoxin A in red wines by means of adsorption treatments with commercial fining agents. J. Agric. Food Chem. 2001, 49, 3917–3921. [Google Scholar] [CrossRef] [PubMed]
- Var, I.; Kabak, B.; Erginkaya, Z. Reduction in ochratoxin A levels in white wine, following treatment with activated carbon and sodium bentonite. Food Control 2008, 19, 592–598. [Google Scholar] [CrossRef]
- Anli, R.E.; Vural, N.; Bayram, M. Removal of ochratoxin A (OTA) from naturally contaminated wines during the vinification process. J. Inst. Brew. 2011, 117, 456–461. [Google Scholar] [CrossRef]
- Quintela, S.; Villarán, M.C.; López de Armentia, I.; Elejalde, E. Ochratoxin A removal from red wine by several oenological fining agents: Bentonite, egg albumin, allergen-free adsorbents, chitin and chitosan. Food Addit. Contam. A 2012, 29, 1168–1174. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Niu, Y.; Ma, T.; Xu, P.; Huang, W.; Zhan, J. Determination, content analysis and removal efficiency of fining agents on ochratoxin A in Chinese wines. Food Control 2017, 73, 382–392. [Google Scholar] [CrossRef]
- Bornet, A.; Teissedre, P.L. Chitosan, chitin-glucan and chitin effects on minerals (iron, lead, cadmium) and organic (ochratoxin A) contaminants in wines. Eur. Food Res. Technol. 2008, 226, 681–689. [Google Scholar] [CrossRef]
- Kurtbay, H.M.; Bekçi, Z.; Merdivan, M.; Yurdakoç, K. Reduction of ochratoxin A levels in red wine by bentonite, modified bentonites and chitosan. J. Agric. Food Chem. 2008, 56, 2541–2545. [Google Scholar] [CrossRef] [PubMed]
- Quintela, S.; Villarán, M.C.; López de Armentia, I.; Elejalde, E. Ochratoxin A removal in wine: A review. Food Control 2013, 30, 439–445. [Google Scholar] [CrossRef]
- Varga, J.; Kocsubé, S.; Péteri, Z.; Vágvölgyi, C.; Tóth, B. Chemical, physical and biological approaches to prevent ochratoxin induced toxicoses in humans and animals. Toxins 2010, 2, 1718–1750. [Google Scholar] [CrossRef] [PubMed]
- European Council. Council Regulation EC No 1493/1999 on the common organization of the market in wine. Off. J. Eur. Communities 1999, 179, 1–84. [Google Scholar]
- International Organization of Vine and Wine (OIV). Resolution OIV/OENO 338A/2009. Wines-Treatment Using Chitosan. 2009. Available online: http://www.oiv.int/public/medias/3551/f-code-ii-3416.pdf (accessed on 25 October 2017).
- International Organization of Vine and Wine (OIV). Resolution OIV/OENO 338B/2009. Wines-Treatment Using Chitin Glucane. 2009. Available online: http://www.oiv.int/public/medias/3553/f-code-ii-3417.pdf (accessed on 25 October 2017).
- The Joint FAO/WHO Expert Committee on Food Additives (JECFA). Safety Evaluation of Certain Mycotoxins in Food. WHO Food Additives Series 2001, No. 47. Available online: http://www.fao.org/3/a-bc528e.pdf (accessed on 25 October 2017).
- European Commission. Commission Regulation EC No 401/2006 laying down the methods of sampling and analysis for the official control of the levels of mycotoxins in foodstuffs. Off. J. Eur. Union 2006, 70, 12–70. [Google Scholar]
- European Food Safety Authority (EFSA). Opinion of the Scientific Panel on Contaminants in the Food Chain on a request from the Commission related to ochratoxin A in food. EFSA J. 2006, 365, 1–56. [Google Scholar]
- VICAM. Global Mycotoxin Regulations. Available online: https://play.google.com/store/apps/details?id=com.vicam.Regulations (accessed on 25 October 2017).
- National Health and Family Planning Commission (NHFPC). Maximum Levels of Mycotoxins in Food (GB 2761-2017). 2017. Available online: http://www.nhfpc.gov.cn/sps/s7891/201704/b83ad058ff544ee39dea811264878981.shtml (accessed on 25 October 2017).
- Bellí, N.; Marín, S.; Sanchis, V.; Ramos, A.J. Impact of fungicides on Aspergillus carbonarius growth and ochratoxin A production on synthetic grape-like medium and on grapes. Food Addit. Contam. 2006, 23, 1021–1029. [Google Scholar] [CrossRef] [PubMed]
- Medina, A.; Mateo, R.; Valle-Algarra, F.; Mateo, E.M.; Jiménez, M. Effect of carbendazim and physicochemical factors on the growth and ochratoxin A production of Aspergillus carbonarius isolated from grapes. Int. J. Food Microbiol. 2007, 119, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Tjamos, S.E.; Antoniou, P.P.; Kazantzidou, A.; Antonopoulos, D.F.; Papageorgiou, I.; Tjamos, E.C. Aspergillus niger and Aspergillus carbonarius in Corinth raisin and wine-producing vineyards in Greece: Population composition, ochratoxin A production and chemical control. J. Phytopathol. 2004, 152, 250–255. [Google Scholar] [CrossRef]
- Bellí, N.; Marín, S.; Argilés, E.; Ramos, A.J.; Sanchis, V. Effect of chemical treatments on ochratoxigenic fungi and common mycobiota of grapes (Vitis vinifera). J. Food Prot. 2007, 70, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Medina, A.; Jiménez, M.; Mateo, R.; Magan, N. Efficacy of natamycin for control of growth and ochratoxin A production by Aspergillus carbonarius strains under different environmental conditions. J. Appl. Microbiol. 2007, 103, 2234–2239. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Wang, Y.; Zeng, H.; Li, Z.; Zhang, P.; Tessema, A.; Peng, X. Efficacy and possible mechanisms of perillaldehyde in control of Aspergillus niger causing grape decay. Int. J. Food Microbiol. 2015, 202, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Favilla, M.; Pascale, M.; Ricelli, A.; Evidente, A.; Amalfitano, C.; Altomare, C. Inhibition of species of the Aspergillus section Nigri and ochratoxin production in grapes by fusapyrone. Appl. Environ. Microbiol. 2008, 74, 2248–2253. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Apaliya, M.T.; Mahunu, G.K.; Chen, L.; Li, W. Control of ochratoxin A-producing fungi in grape berry by microbial antagonists: A review. Trends Food Sci. Technol. 2016, 51, 88–97. [Google Scholar] [CrossRef]
- Bleve, G.; Grieco, F.; Cozzi, G.; Logrieco, A.; Visconti, A. Isolation of epiphytic yeast with potential biocontrol of Aspergillus carbonarius and A. niger on grape. Int. J. Food Microbiol. 2006, 108, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Ponsone, M.L.; Chiotta, M.L.; Combina, M.; Dalcero, A.; Chulze, S. Biocontrol as a strategy to reduce the impact of ochratoxin A and Aspergillus section Nigri in grapes. Int. J. Food Microbiol. 2011, 151, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Fiori, S.; Urgeghe, P.P.; Hammami, W.; Razzu, S.; Jaoua, S.; Migueli, Q. Biocontrol activity of four non- and low-fermenting yeast strains against Aspergillus carbonarius and their ability to remove ochratoxin A from grape juice. Int. J. Food Microbiol. 2014, 189, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Pantelides, I.S.; Christou, O.; Tsolakidou, D.; Tsaltas, D.; Ioannou, N. Isolation, identification and in vitro screening of grapevine yeast for the control of black Aspergilli on grapes. Biol. Control 2015, 88, 46–53. [Google Scholar] [CrossRef]
- Dimakopoulou, M.; Tjamos, S.E.; Antoniou, P.P.; Pietri, A.; Battilani, P.; Avramidis, N.; Markakis, E.A.; Tjamos, E.C. Phyllosphere grapevine yeast Aureobasidium pullulans reduce Aspergillus carbonarius (sour rot) incidence in wine-producing vineyards in Greece. Biol. Control 2008, 46, 158–165. [Google Scholar] [CrossRef]
- Ponsone, M.L.; Nally, M.C.; Chiotta, M.L.; Combina, M.; Köhl, J.; Chulze, S.N. Evaluation of the effectiveness of potential biocontrol yeasts against black sour rot and ochratoxin A occurring under greenhouse and field grape production conditions. Biol. Control 2016, 103, 78–85. [Google Scholar] [CrossRef]
- De Curtis, F.; de Felice, D.V.; Ianiri, G.; De Cicco, V.; Castoria, R. Environmental factors affect the activity of biocontrol agents against ochratoxigenic Aspergillus carbonarius on wine grape. Int. J. Food Microbiol. 2012, 159, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Solfrizzo, M.; Avantaggiato, G.; Panzarini, G.; Visconti, A. Removal of ochratoxin A from contaminated red wines by repassage over grape pomaces. J. Agric. Food Chem. 2010, 58, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Farbo, M.G.; Urgeghe, P.P.; Fiori, S.; Marceddu, S.; Jaoua, S.; Migueli, Q. Adsorption of ochratoxin A from grape juice by yeast cells immobilised in calcium alginate beads. Int. J. Food Microbiol. 2015, 217, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Bejaoui, H.; Mathieu, F.; Taillandier, P.; Lebrihi, A. Ochratoxin A removal in synthetic and natural grape juices by selected oenological Saccharomyces strains. J. Appl. Microbiol. 2004, 97, 1038–1044. [Google Scholar] [CrossRef] [PubMed]
- Petruzzi, L.; Baiano, A.; De Gianni, A.; Sinigaglia, M.; Corbo, M.R.; Bevilacqua, A. Differential adsorption of ochratoxin A and anthocyanins by inactivated yeasts and yeast cell walls during simulation of wine aging. Toxins 2015, 7, 4350–4365. [Google Scholar] [CrossRef] [PubMed]
- European Committee for Standardization. CSN EN 14133—Foodstuffs—Determination of Ochratoxin A in Wine and Beer—HPLC with Immunoaffinity Column Clean-Up; Czech Office for Standards, Metrology and Testing: Praha, Czech Republic, 2009. [Google Scholar]
- Brera, C.; Grossi, S.; De Santis, B.; Miraglia, M. Automated HPLC method for the determination of ochratoxin A in wine samples. J. Liq. Chromatogr. Relat. Technol. 2003, 26, 119–133. [Google Scholar] [CrossRef]
- Giraudi, G.; Ferrero, V.E.V.; Anfossi, L.; Baggiani, C.; Giovannoli, C.; Tozzi, C. Solid-phase extraction of ochratoxin A from wine based on a binding hexapeptide prepared by combinatorial synthesis. J. Chromatogr. 2007, 1175, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Kong, W.; Zhou, S.; Yin, L.; Wan, L.; Yang, M. Molecularly imprinted polymer-based solid phase clean-up for analysis of ochratoxin A in beer, red wine, and grape juice. J. Sep. Sci. 2013, 36, 1291–1297. [Google Scholar] [CrossRef] [PubMed]
- Hernández, M.J.; García-Moreno, M.V.; Durán, E.; Guillén, D.; Barroso, G. Validation of two analytical methods for the determination of ochratoxin A by reversed-phased high-performance liquid chromatography coupled to fluorescence detection in must and sweet wines from Andalusia. Anal. Chim. Acta 2006, 566, 117–121. [Google Scholar] [CrossRef]
- Longobardi, F.; Iacovelli, V.; Catucci, L.; Panzarini, G.; Pascale, M.; Visconti, A.; Agostiano, A. Determination of ochratoxin A in wine by means of immunoaffinity and aminopropyl solid-phase column cleanup and fluorometric detection. J. Agric. Food Chem. 2013, 61, 1604–1608. [Google Scholar] [CrossRef] [PubMed]
- Dall’Asta, C.; Galaverna, G.; Dossena, A.; Marchelli, R. Reversed-phase liquid chromatographic method for the determination of ochratoxin A in wine. J. Chromatogr. A 2004, 1024, 275–279. [Google Scholar] [CrossRef] [PubMed]
- García-Fonseca, S.; Ballesteros-Gómez, A.; Rubio, S.; Pérez-Bendito, D. Coacervate extraction of ochratoxin A in wines prior to liquid chromatography/fluorescence determination. Anal. Chim. Acta 2008, 617, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Vidal, J.C.; Bonel, L.; Ezquerra, A.; Duato, P.; Castillo, J.R. An electrochemical immunosensor for ochratoxin A determination in wines based on a monoclonal antibody and paramagnetic microbeads. Anal. Bioanal. Chem. 2012, 403, 1585–1593. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, P.J.; Barros, N.; Camara, J.S. A survey of the occurrence of ochratoxin A in Madeira wines based on a modified QuEChERS extraction procedure combined with liquid chromatography-triple quadrupole tandem mass spectrometry. Food Res. Int. 2013, 54, 293–301. [Google Scholar] [CrossRef]
- Zezza, F.; Longobardi, F.; Pascale, M.; Eremin, S.A.; Visconti, A. Fluorescence polarization immunoassay for rapid screening of ochratoxin A in red wine. Anal. Bioanal. Chem. 2009, 395, 1317–1323. [Google Scholar] [CrossRef] [PubMed]
- Arroyo-Manzanares, N.; Gámiz-Gracia, L.; García-Campaña, A.M. Determination of ochratoxin A in wines by capillary liquid chromatography with laser induced fluorescence detection using dispersive liquid-liquid microextraction. Food Chem. 2012, 135, 368–372. [Google Scholar] [CrossRef] [PubMed]
- Barthelmebs, L.; Jonca, J.; Hayat, A.; Prieto-Simon, B.; Marty, J.L. Enzyme-Linked Aptamer Assays (ELAAs), based on a competition format for a rapid and sensitive detection of ochratoxin A in wine. Food Control 2011, 22, 737–743. [Google Scholar] [CrossRef]
- Yin, X.; Wang, S.; Liu, X.; He, C.; Tang, Y.; Li, Q.; Liu, Q.; Su, H.; Tan, T.; Dong, Y. Aptamer-based colorimetric biosensing of ochratoxin A in fortified white grape wine sample using unmodified gold nanoparticles. Anal. Sci. 2017, 33, 659–664. [Google Scholar] [CrossRef] [PubMed]
- Remiro, R.; González-Peñas, E.; Lizarraga, E.; López de Cerain, A. Quantification of ochratoxin A and five analogs in Navarra red wines. Food Control 2012, 27, 139–145. [Google Scholar] [CrossRef]
- Di Stefano, V.; Avellone, G.; Pitonzo, R.; Capocchiano, V.G.; Mazza, A.; Cicero, N.; Dugo, G. Natural co-occurrence of ochratoxin A, ochratoxin B and aflatoxins in Sicilian red wines. Food Addit. Contam. Part A 2015, 32, 1343–1351. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Chemical structure of the main natural occurring mycotoxins.
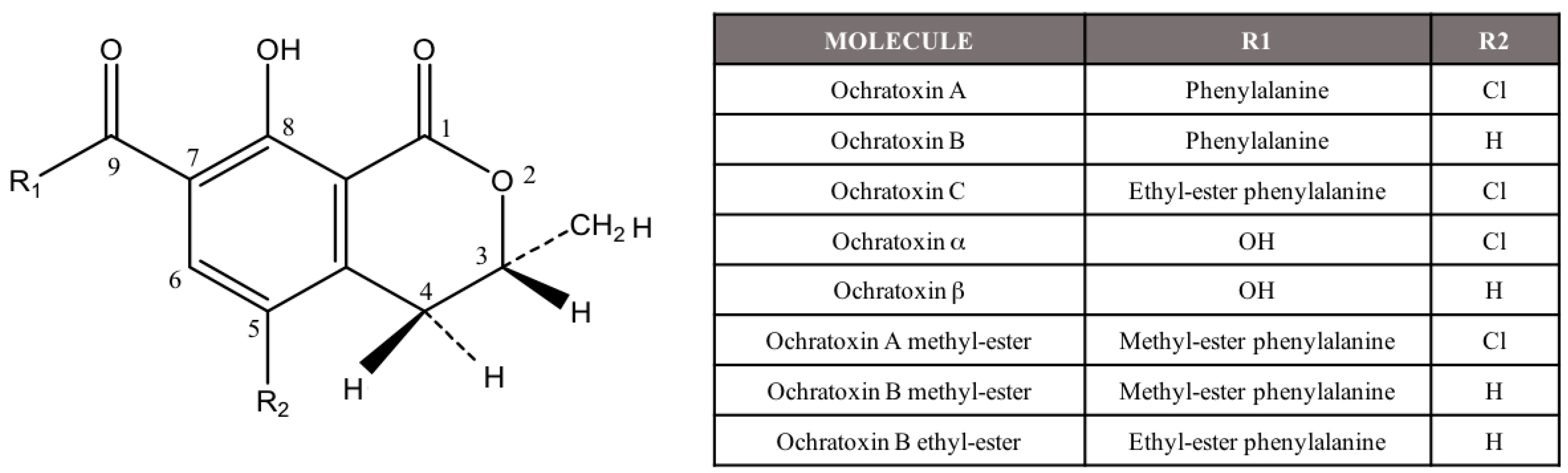
Figure 2.
Schematic representation of winemaking process. Steps for red (left) and white (right) wine production are indicated. The arrows are related to the increase or decrease in Ochratoxin A (OTA) content during each step.
Figure 2.
Schematic representation of winemaking process. Steps for red (left) and white (right) wine production are indicated. The arrows are related to the increase or decrease in Ochratoxin A (OTA) content during each step.


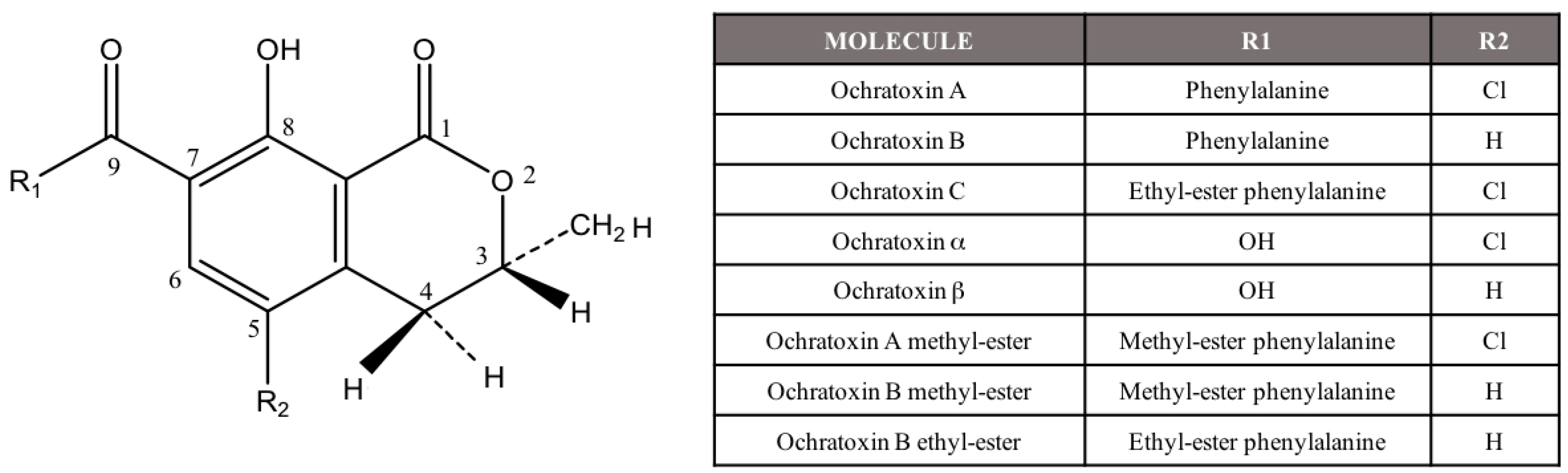{kind=link}
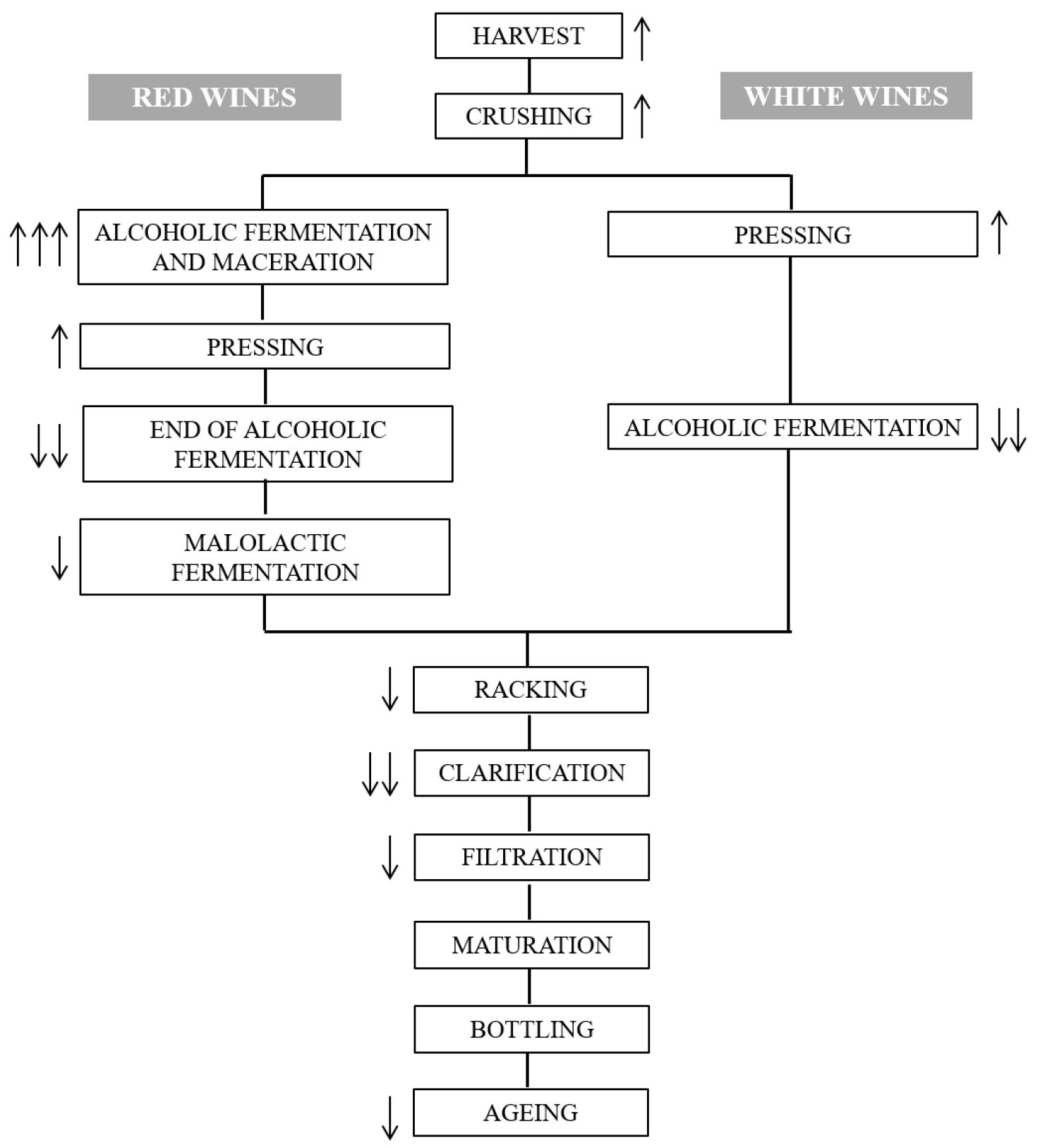{kind=link}
Table 1.
Main studies regarding the worldwide occurrence of OTA in red, white and dessert wines. The results are indicated as the number of positive samples/total samples (percentage of positive samples) of each type of wine. The number of samples exceeding the EU legal limits (2 µg/L) as well as the percentage represented by these samples (between parentheses) are also indicated. The analysis methods used and the highest concentration detected are included.
Table 1.
Main studies regarding the worldwide occurrence of OTA in red, white and dessert wines. The results are indicated as the number of positive samples/total samples (percentage of positive samples) of each type of wine. The number of samples exceeding the EU legal limits (2 µg/L) as well as the percentage represented by these samples (between parentheses) are also indicated. The analysis methods used and the highest concentration detected are included.
| COUNTRY | RED | WHITE | DESSERT | ANALYSIS METHOD | MAXIMUM OTA (µg/L) | SAMPLES ABOVE LIMITS | REFERENCE |
|---|---|---|---|---|---|---|---|
| Argentina | 4/47 (9%) | - | - | HPLC | 4.82 | 2 (4%) | [42] |
| Australia | 49/344 (14%) | 41/257 (16%) | 3/13 (23%) | HPLC | 0.62 | 0 (0%) | [43] |
| Canada | 10/43 (23%) | 5/36 (14%) | - | LC | 0.39 | 0 (0%) | [44] |
| Chile | 28/841 (3%) | 6/313 (2%) | - | HPLC | 0.35 | 0 (0%) | [45] |
| China | 18/63 (29%) | 4/42 (10%) | - | LC/MS | 1.18 | 0 (0%) | [46] |
| 44/77 (57%) | 1/34 (3%) | - | HPLC | 5.65 | 3 (3%) | [47] | |
| Czech Republic | 2/13 (15%) | 3/33 (9%) | - | UPLC | 0.07 | 0 (0%) | [48] |
| France | - | - | 47/49 (96%) | LC/MS | 1.22 | 0 (0%) | [49] |
| Greece | 71/104 (68%) | 63/118 (53%) | 15/18 (83%) | HPLC | 2.82 | n.a. | [50] |
| 9/14 (65%) | 7/13 (54%) | 6/7 (86%) | HPLC | 3.20 | 3 (9%) | [51] | |
| 45/64 (70%) | 31/49 (63%) | 22/27 (81%) | HPLC | 2.00 | 0 (0%) | [52] | |
| Hungary | 33/33 (100%) | 31/32 (97%) | - | HPLC | 0.53 | 0 (0%) | [53] |
| Italy | 86/92 (93%) | - | 9/15 (60%) | HPLC | 3.86 | n.a. | [54] |
| 695/1002 (69%) | 125/204 (61%) | - | HPLC | 2.63 | 29 (2%) | [55] | |
| 37/38 (97%) | 4/9 (57%) | - | HPLC | 7.63 | 6 (13%) | [56] | |
| 535/773 (69%) | 128/290 (44%) | 18/28 (64%) | HPLC | 7.50 | 22 (2%) | [57] | |
| - | - | 29/30 (97%) | HPLC | 1.56 | 0 (0%) | [58] | |
| Romania | 17/44 (39%) | 21/55 (38%) | - | HPLC | 1.89 | 0 (0%) | [59] |
| Spain | 24/130 (19%) | 4/50 (8%) | - | HPLC | 4.24 | 2 (1%) | [60] |
| 108/188 (57%) | 4/6 (67%) | - | HPLC | 0.18 | 0 (0%) | [61] | |
| - | - | 32/40 (80%) | LC | 0.64 | 0 (0%) | [62] | |
| - | - | 186/188 (99%) | LC/MS | 4.63 | 18 (1%) | [49] | |
| South Africa | 9/9 (100%) | 15/15 (100%) | - | HPLC | 0.39 | 0 (0%) | [63] |
| 19/65 (29%) | 20/42 (48%) | 11/15 (73%) | HPLC | 2.67 | n.a. | [64] | |
| Turkey | 44/51 (86%) | 29/34 (85%) | - | HPLC | 0.82 | 0 (0%) | [65] |
(-) is indicated when the paper did not analyze this type of wine. n.a. = data not available in the original paper.
Table 2.
Methods for determination of OTA in wine.
| Type of Wine | Extraction | Clean-up | Detection | Recovery | Reproducibility (%RSD) | Detection Limits | Special Remarks | Reference |
|---|---|---|---|---|---|---|---|---|
| White and red | PBS | IAC | HPLC-FD | 84–95% | <5% | 0.01 µg/L | Automated protocol | [136] |
| White, rosé, red, dessert | PEG8000 | Peptide SPE | HPLC | 75–113% | n.a | 0.10 µg/L | Efficiency similar to IAC | [137] |
| Red and must | - | MIP SPE | HPLC | 91–103% | <5% | 0.025 µg/L | MIP columns are reusable | [138] |
| White, rosé, red, dessert | PEG8000 + NaHCO3 | IAC | HPLC-MS | 88–104% | <10% | 0.01 µg/L | Reference method | [56] |
| Red and must | - | C18 SPE | HPLC-FD | 94–114% | <2% | 0.22–0.77 µg/L | Must do not require SPE | [139] |
| Red | PEG8000 + NaHCO3 | IAC + Aminopropyl SPE | HPLC-FD | 95–105% | <15% | 0.2 µg/L | Double clean up to reduce matrix interference | [140] |
| White, rosé, red, dessert | - | - | HPLC-FD | 93–96% | <5% | 0.05 µg/L | Without extraction and clean up | [141] |
| White, rosé, red | - | Coacervate of decanoic acid in THF | HPLC | 85–100% | <7% | 0.004–0.015 µg/L | - | [142] |
| Red | PVP + PEG3350 | Magnetic beads linked to anti-OTA antibodies | Electrochemical | 105–109% | <9% | 0.11 µg/L | Competitive electrochemical immunosensor | [143] |
| Dessert | QuEChERS | Dispersive SPE | HPLC-MS | 87–103% | <9% | 0.1 µg/L | - | [144] |
| Red | - | Aminopropyl SPE | FP immunoassay | 75–83% | <12% | 0.7 µg/L | Rapid procedure (less than 10 min) | [145] |
| White, rosé, red | - | DLLME | HPLC-LIF | 92–98% | <5% | 0.006 µg/L | Green analytical chemistry alternative | [146] |
DLLME: Dispersive liquid–liquid microextraction; IAC: Immunoaffinity column; FP: Fluorescence polarization; HPLC-FD: High performance liquid chromatography with fluorescence detection; HPLC-LIF: High performance liquid chromatography laser-induced fluorescence detection; MIP: Molecularly imprinted polymer; MS: Mass Spectometry; SPE: Solid Phase Extraction; THF: Tetrahydrofuran; (-) is indicated when this step is not necessary in the described method. n.a: Data not available in the original paper.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gil-Serna, J.; Vázquez, C.; González-Jaén, M.T.; Patiño, B. Wine Contamination with Ochratoxins: A Review. Beverages 2018, 4, 6. https://doi.org/10.3390/beverages4010006
AMA Style
Gil-Serna J, Vázquez C, González-Jaén MT, Patiño B. Wine Contamination with Ochratoxins: A Review. Beverages. 2018; 4(1):6. https://doi.org/10.3390/beverages4010006
Chicago/Turabian StyleGil-Serna, Jessica, Covadonga Vázquez, María Teresa González-Jaén, and Belén Patiño. 2018. "Wine Contamination with Ochratoxins: A Review" Beverages 4, no. 1: 6. https://doi.org/10.3390/beverages4010006
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.