Effects of MP Polyethylene Microparticles on Microbiome and Inflammatory Response of Larval Zebrafish
by

Nicholas Kurchaba
, 
Bryan J. Cassone
,
Caleb Northam
,
Bernadette F. Ardelli
and
Christophe M. R. LeMoine
* Department of Biology, Brandon University, Brandon, MB R7A 6A9, Canada
*
Author to whom correspondence should be addressed.
Toxics 2020, 8(3), 55; https://doi.org/10.3390/toxics8030055
Submission received: 8 July 2020
/
Revised: 29 July 2020
/
Accepted: 3 August 2020
/
Published: 11 August 2020
(This article belongs to the Special Issue Prevalence, Fate and Effects of Plastic in Freshwater Environments)
Abstract
:Plastic polymers have quickly become one of the most abundant materials on Earth due to their low production cost and high versatility. Unfortunately, some of the discarded plastic can make its way into the environment and become fragmented into smaller microscopic particles, termed secondary microplastics (MP). In addition, primary MP, purposely manufactured microscopic plastic particles, can also make their way into our environment via various routes. Owing to their size and resilience, these MP can then be easily ingested by living organisms. The effect of MP particles on living organisms is suspected to have negative implications, especially during early development. In this study, we examined the effects of polyethylene MP ingestion for four and ten days of exposure starting at 5 days post-fertilization (dpf). In particular, we examined the effects of polyethylene MP exposure on resting metabolic rate, on gene expression of several inflammatory and oxidative stress linked genes, and on microbiome composition between treatments. Overall, we found no evidence of broad metabolic disturbances or inflammatory markers in MP-exposed fish for either period of time. However, there was a significant increase in the oxidative stress mediator L-FABP that occurred at 15 dpf. Furthermore, the microbiome was disrupted by MP exposure, with evidence of an increased abundance of Bacteroidetes in MP fish, a combination frequently found in intestinal pathologies. Thus, it appears that acute polyethylene MP exposure can increase oxidative stress and dysbiosis, which may render the animal more susceptible to diseases.

1. Introduction
Plastic materials are inexpensively produced and provide a high level of durability and flexibility, making them a key material for various applications. While an increasing number of plastics are re-used and recycled, many “single use” plastics are readily discarded and accumulate as waste worldwide [1]. Over 4900 metric tons of plastic waste have been discarded in landfills and the environment since 1950 [1,2]. Discarded plastics are derived from different polymers, such as polyethylene, polypropylene, and polystyrene, with each polymer providing unique and desirable qualities. Polyethylene is particularly desirable for packaging due to its resistance to degradation and low economic cost and thus is of major environmental importance [3].
Plastic pollution affects most ecosystems, including freshwater and oceanic systems. In these environments, plastic litter undergoes photo-oxidation and physical breakdown, generating smaller and chemically stable particles. When these particles are smaller than 5 mm in diameter, they are called secondary microplastics (MP) and may persist extensively in the environment and in organisms [4,5,6,7]. In contrast, primary MP are intentionally manufactured for various applications [6]. Regardless of their provenance, primary and secondary MP have been found in oceans for decades, though more recently they have been detected in freshwater, reaching some of the most remote lakes around the world [2,8,9]. In freshwater reserves, such as the Ottawa River and the Nantaizi Lake, MP can attain concentrations between 0.1 and 6162.5 p/L [10,11,12]. As MP can easily be ingested, their presence in aquatic environments poses a potential risk to many animal species, chronically exposing them to this pervasive pollutant. In aquatic animals, MP primarily accumulate both in the alimentary canal and respiratory structures in adult animals, though they appear to be limited to the intestine in aquatic larvae [13,14].
Once ingested, MP can negatively impact organismal health at different levels [15,16,17]. First, MP can disrupt intestinal transit and their accumulation may trigger intestinal villi swelling [18]. In addition, at the cellular level, MP can also cause physical injury to surrounding cells via abrasion-induced oxidative stress and accumulation of intercellular reactive oxygen species (ROS) [19]. Enhanced intestinal ROS production may also indicate an inflammatory response and participate in a feedback loop [20]. Regardless of the cause, excess ROS production jeopardizes tissue integrity and overall organismal health [21].
The inflammatory response is coordinated by multiple cascades, including pro- and anti-inflammatory mediators. These cascades can be initiated by multiple stressors, such as toxins, tissue damage, or microorganisms [22], and may trigger cytokines to flood surrounding tissues and recruit nearby immune cells to initiate inflammation. While inflammation allows for the removal of the foreign stimuli and promotion of active healing of the injured area, prolonged inflammation can also result in various pathologies, such as inflammatory bowel disease, leading to metabolic dysfunction and nutrient deficiency [23,24].
The intestinal accumulation of MP may also affect its microbiome, another crucial player in animal wellbeing. In many organisms, including fish, it is clear that the microbiome plays a crucial role in both health and diseases; thus, disturbances in the gut microbiome can be detrimental to overall animal health [25,26,27]. As in other organisms, the fish microbiome can be affected by MP, with presumed widespread effects on animal health [28,29,30,31,32,33]. Indeed, gut dysbiosis has been repeatedly linked to various metabolic disorders [34,35,36,37].
Finally, the combined effect of oxidative stress, inflammation, and gut dysbiosis could have an even more pronounced impact on fish during early development when energy usage and physiological stress is high [38,39,40]. In particular, increased energy expenditure could negatively impact vital processes, such as tissue maintenance, locomotion, and growth of the individual. MP can also indirectly affect energy budgets by promoting an immune response to purge and remove plastic particles via inflammation [41].
In order to investigate the potential negative effects of MP on a freshwater fish model organism, we exposed larval zebrafish to polyethylene MP (10–40 μm) at a concentration of 20 mg/L. The range selected simulates a mixed-size exposure while providing an adequate representation of the type of microplastics found in the environment, and provides consistency with previous laboratory investigations using supra-ecological concentrations [42,43,44,45]. The experiment examined the acute and prolonged effects of MP exposure throughout early development by assessing oxygen consumption rates (OCRs), directed gene expression analysis, and microbiome diversity to assess the various impacts of differential MP exposure. Our results show that MP exposure during early life has no impact on metabolic or immunological systems but appears to have localized effects within the gut by increasing oxidative stress and disturbing the gut microbiome.
2. Methods
2.1. Caretaking
Commercially obtained adult zebrafish (Danio rerio) were maintained in an aquatic housing system (Aquaneering, San Diego, CA, USA) with recirculating UV-treated de-chlorinated water maintained at 27 °C with a 14:10 h light dark cycle. Adult animals were fed twice daily with Adult Zebrafish Complete Diet (Zeigler, Gardners, PA, USA) and once with Artemia naupii. Adult fish were bred in tanks using standard procedures [46], and embryos were collected before being washed twice in 0.05% bleach solution and transferred to 90 × 15 mm petri plates filled with E3 buffer solution (5 mM NaCl, 0.17 mM KCl, 0.33 mM CaCl2, 0.33 mM MgSO4, 0.0006 mM methylene blue). The fish were then kept at 28 °C until day 5 post-fertilization (5 dpf), with E3 medium being replaced once every 24 h. At 5 dpf, the fish were transferred into 300 mL beakers and maintained in de-chlorinated control water or water containing MP. The MP fish were exposed to commercially obtained 10–45 µm polyethylene microspheres (Cospheric, Goleta, CA, USA) at a concentration of 20 mg/L. All fish (MP and control) were kept under these conditions for either 4 or 10 days, corresponding to 9 dpf and 15 dpf, respectively. Under similar experimental conditions, this treatment previously yielded consistent ingestion and accumulation of MP in all zebrafish larvae [13]. All treatments were maintained in a 27 °C water bath for the duration of the experiment. Daily, larvae were fed ad libitum Larval AP100 feed (Ziegler, Gardners, PA, USA), and approximately one third of the water, 100 mL of either MP or pristine freshwater, was replaced to maintain water quality and remove deceased larvae. As with prior work [13], mortality rates remained low (<2%) throughout the experiment. Bioassays were run in parallel using fish from 6 independent breeding events. All animal experimentations conform to the guidelines of the Canadian Council on Animal Care and were approved by the Brandon University Animal Care Committee on July 12 2016 under the research protocol 2016R03.
2.2. Respirometry
At 9 and 15 dpf, the oxygen consumption rates of zebrafish larvae were measured using a fibre-optic respirometry system (OXY-4 mini, PreSens, Regensburg, Germany). Both oxygen consumption and temperature values were recorded in parallel with the AutoResp software v2.1.2 (Loligo® Systems, Viborg, Denmark), and data were analyzed using RespR v1.1.0.0 [47]. The respirometry procedure was conducted in a temperature-controlled water bath (26.91 °C ± 0.02) for the duration of the experiment. Fish were placed into borosilicate chambers (2–3 mL) fitted with small optodes using aquarium-grade silicon (Marineland, Blacksburg, VA, USA). Magnetic stir bars allowed for adequate oxygen circulation and their consistent rotation was improved by the addition of rubber rings at the bottom of the tube. The system was calibrated with 100% O2 saturated E3 medium or dissolved sodium sulfite (0.02 g/mL solution). The fish were acclimated to the borosilicate chambers for 30 min and half of the total volume was replaced with fresh E3 prior to data collection. Each trial consisted of a 20 min sampling period featuring an early section (2 min), an intermediate section (8 min), and a late section (10 min). A sample size of 6 replicates (where each replicate included 5–10 fish depending on the size of the tube used) was used to determine OCRs between the treatments. We also measured background OCR (with no fish in chambers) and found these values to be negligible compared to larval OCRs. The intermediate sections were plotted using RespR to identify the most linear portion of the slope used for OCR calculation [47]. The OCRs were converted to nanomol/fish/hour to remain consistent with previous literature [13,48].
2.3. RNA Extraction, Reverse Transcription (RT), and qPCR
Following 4 or 10 days of exposure (9 dpf or 15 dpf), the larvae were anesthetized in 100 mg/L tricaine methanesulfonate (MS–222, Sigma-Aldrich, Darmstadt, Germany) and quick-frozen in liquid nitrogen before being stored at −80 °C until further manipulation. RNA was isolated from a total of 36 samples, comprising 9 replicates for control and MP treatments at both 9 and 15 dpf, using an RNeasy Mini extraction kit in accordance to the manufacturer’s protocol (RNeasy, Qiagen, Aarhus, Denmark). A final volume of 20 µL was obtained for each sample, and the quantity and quality of the RNA was determined by the use of a NP80 NanoPhotometer® (Implen, Munich, Germany). Optical density ratios of 260/280 nm above 1.9 and concentrations above 500 ng/µL were targeted in order to provide a total of 2 μg of RNA for reverse transcription using the GoScript™ Reverse Transcription System (Promega, Madison, WI, USA). The transcribed cDNA was then diluted (1:10) prior to qPCR, as described before [49].
2.4. qPCR
Nine genes were selected in order to examine the inflammatory response of larval zebrafish. Primer sets were obtained from previous publications or custom designed with the use of Primer3 software (Table 1, [50,51,52,53,54,55,56,57]). Primer specificity was confirmed by examining the top e-value in BLASTn (NCBI) and single product amplification confirmed by melt analysis of all reactions.
Gene expression analysis was carried by qPCR amplification using the QuantiNova™ SYBR Green PCR kit (Qiagen, Aarhus, Denmark), with a final volume of 15 µL per reaction (7.5 µL 2X SYBR Green PCR Master Mix, 0.7 µM forward and reverse primer, respectively, and 1 µL cDNA). Standard curves were created for each of the primer sets and the efficiency of each primer set was computed using Quantinova Rotor-Gene Q Series Software (Qiagen). Melting curve analysis was also performed to ensure single product amplification in each reaction. Only primer sets with suitable efficiency values (1.00 +/− 0.20) and single amplicon were used for further testing. The cycle threshold (Ct) values were automatically calculated using Rotor-Gene software using the corresponding standard curve. Relative gene expression levels were calculated using the ΔΔCt method [58], using the geometric mean of two housekeeping genes (eF1α and Rpl-13a) as a reference [49,58].
2.5. Respirometry and Gene Expression Statistical Analysis
Respirometry data and gene expression data were analysed in SigmaPlot software (SigmaPlot 14.0, Systat Software, San Jose, CA, USA). A two-way ANOVA was used to compare both the respirometry and gene expression data. All tests were performed at a significance level of p = 0.05, after passing equal variance and normality tests.
2.6. Microbiome Analysis
2.6.1. Sampling, DNA Extraction, Sequencing, and Pre-Processing
At both 9 and 15 dpf, 6 replicate samples of each treatment containing 20 individual zebrafish larvae each were anesthetized in 100 mg/L tricaine methanesulfonate, thoroughly washed with 100% ethanol, flash frozen, and stored at −80 °C until further analyses. For each sample, genomic DNA was isolated using the One-4-All Genomic DNA Miniprep Kit (BioBasic, Markham, ON, Canada).
Samples were then sent to Génome Québec Innovation Centre (McGill University, Montreal, QC, Canada) for quality control, PCR validation and reaction, amplicon barcoding, and normalization. The 16S primers used were 515F: GTGCCAGCMGCCGCGGTAA and 806R: GGACTACHVGGGTWTCTAAT [59]. A BioAnalyzer 2100 (Agilent Technologies, Santa Clara, CA, USA) was used to check the quality of amplicon libraries that were subsequently sequenced on the MiSeq platform in a paired-end 300 bp fashion. Raw sequence reads were deposited in the NCBI short sequence read archive (SRA) under the accession number PRJNA643286. The amplicons were adapter and quality trimmed in CLC Genomics Workbench v11.0.1 (Qiagen, Aarhus, Denmark; Quality Limit = 0.00316, Minimum Length = 15, default parameters herein). The reads were then processed in CLC v11.0.1 for downstream analyses, as described previously [60].
2.6.2. CLC Analysis
We used the CLC Microbial Genomics Module 3.5 (Qiagen) pipeline for 16S analysis. First, operational taxonomic units (OTUs) were clustered at 97% similarity against the reference SILVA 132 database [61]. Chimeric sequences as well as OTUs detected in less than 0.05% of the total reads were then filtered out.
Bacterial communities were compared across ages and between control and MP fish. The OTU table was rarefied before OTU alpha diversity analysis was carried out using Shannon and Simpson’s alpha diversity indices as well as total OTU in CLC. Differences between all groups were assessed with a non-parametric Kruskall–Wallis test followed by Mann–Whitney U pairwise tests (α = 0.05).
Furthermore, beta diversity was evaluated with weighted and unweighted UNIFRAC as well as the Bray–Curtis index. Principal coordinates analyses (PCoA) of the distance plots were visualized, and multivariate analyses (PERMANOVA) were used to compare treatment differences. Differential abundance was computed using a binomial general linear model framework, and a likelihood test was used to detect differences across groups, followed by a Wald test for pairwise comparisons (FDR < 0.05).
3. Results
3.1. Respirometry
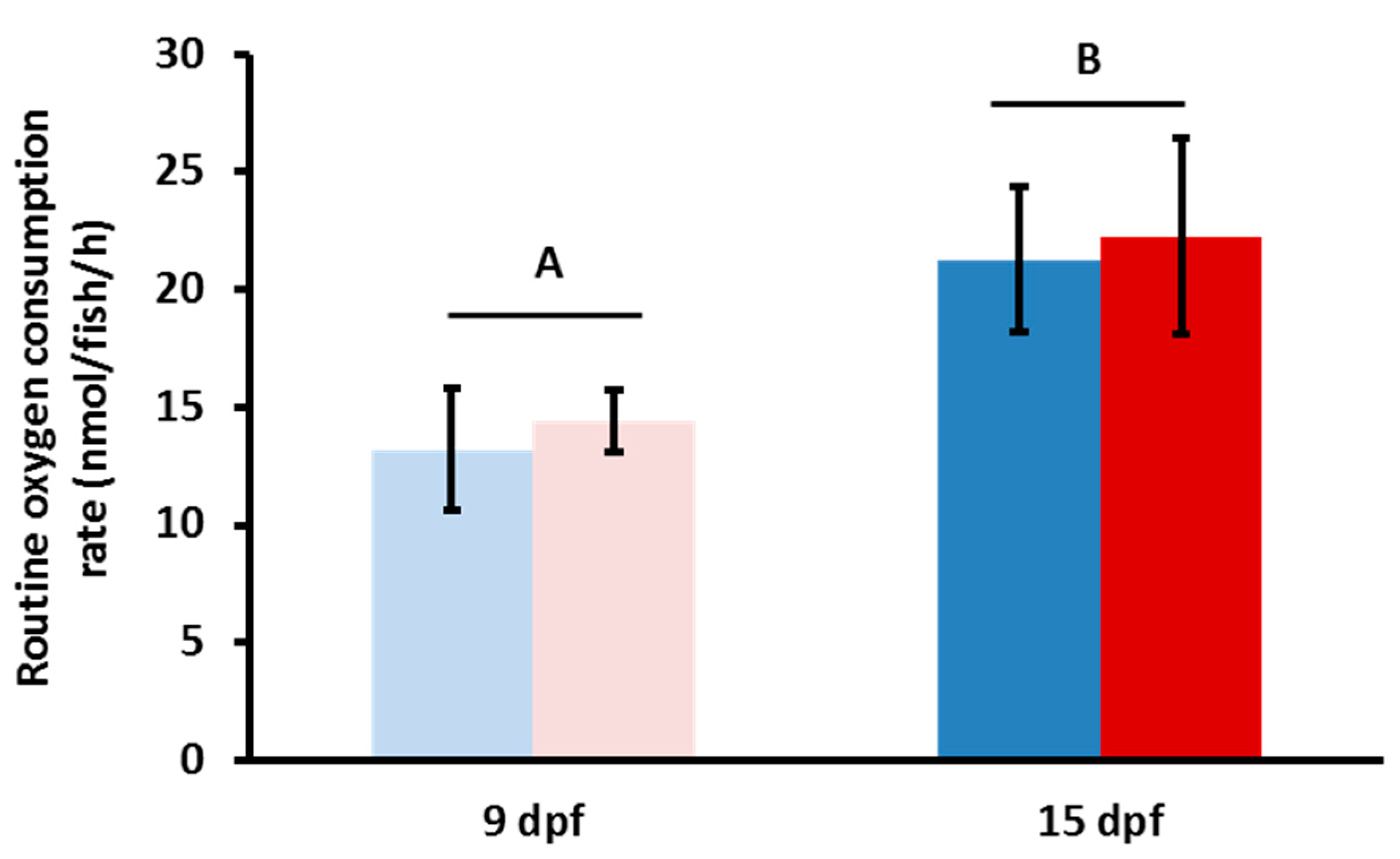
We explored the effects of MP exposure on routine metabolic rate using closed-system respirometry (Figure 1). Mean oxygen consumption rates (OCRs) between fish raised in clear water (control) and MP-exposed fish at both developmental time points were not statistically significant (Figure 1). However, we were able to identify a difference in OCRs when comparing the two time points, 9 dpf and 15 dpf (p = 0.015). To test whether the developmental time point of the zebrafish was more important than the overall MP duration, we also examined a 10–15 dpf exposure period to MP; however, no significant difference was found when compared to the 5–15 dpf treatment (data not shown). Ultimately, no statistically significant difference in mean OCRs was identified in any of the treatments in regard to MP exposure (Figure 1).
3.2. Gene Expression Analysis
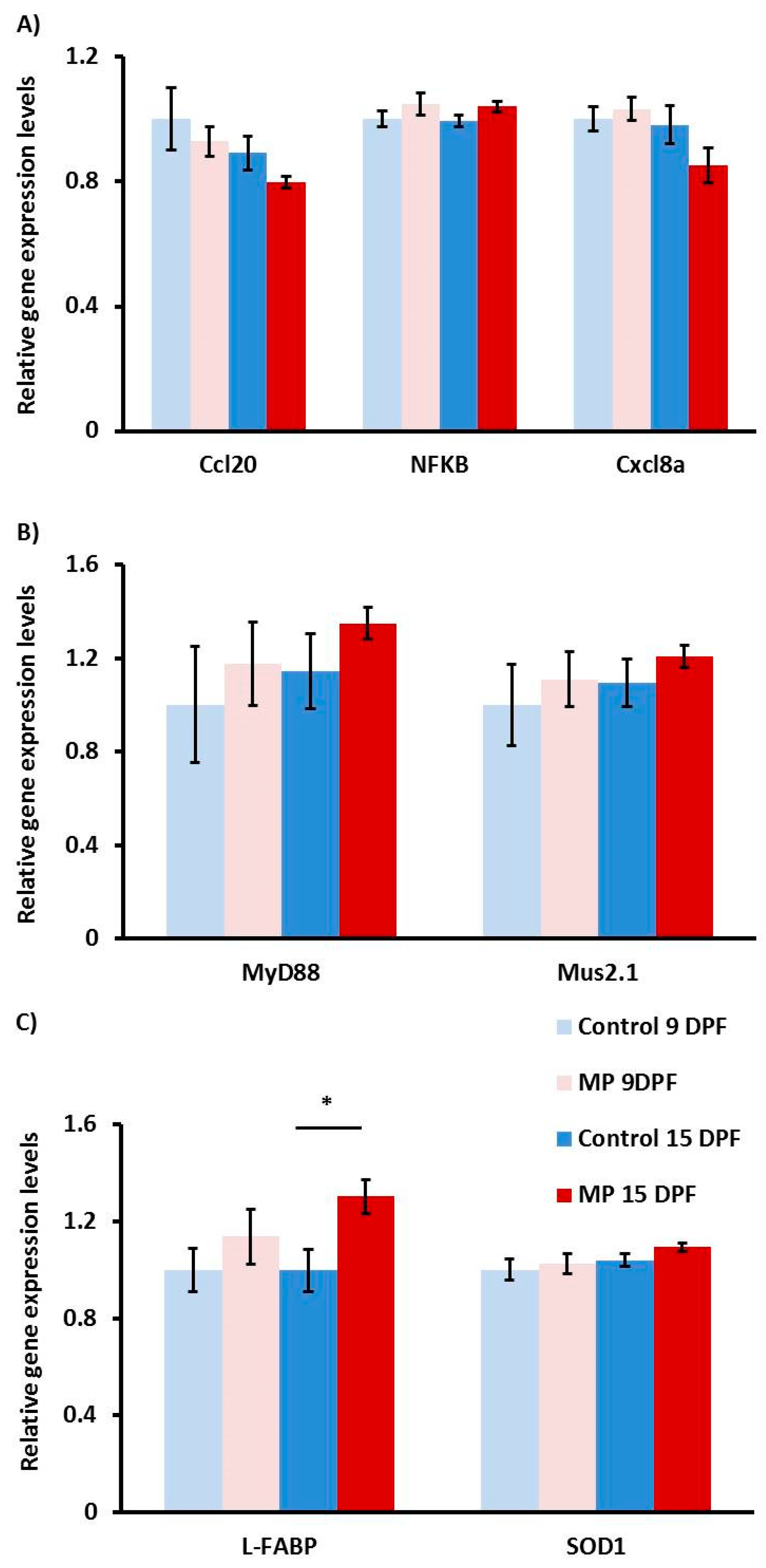
In order to assess the effects of MP exposure on various cellular response pathways, we used qPCR and evaluated patterns of gene expression of nine gene targets involved with the inflammatory response, the gut microbiome, or oxidative stress (Figure 2). In terms of MP inflammatory response, none of the target genes (ccl20, cxcl8a, and NF-κβ) appeared to be affected across treatments. Although ccl20 and cxcl8a both show a marginal reduction in gene expression in 15 dpf MP fish, this effect was not statistically significant (Figure 2A). Similarly, genes associated with the gut microbiome (MyD88 and Mus2.1) exhibited relatively high variability resulting in no appreciable difference between treatments (Figure 2B). Finally, genes that are typically upregulated in periods of oxidative stress, L-FABP and SOD1, were also examined. In this case, we detected a significant increase in L-FABP expression in MP fish at day 15 (p = 0.022), but a marginal and insignificant increase in SOD1 in the same fish (Figure 2C).
3.3. Structure of Gut Bacteria Communities
We aimed to characterize the effect of MP exposure on the microbiome of zebrafish larvae for different exposure duration. Fish were exposed to MP or control conditions and sampled at 4 and 10 days, and their DNA was extracted (n = 6 for each treatment and time point) and was subjected to high throughput 16S sequencing. We obtained a total of 3,535,330 reads, of which 42% were retained as OTU reads after quality trimming and chimeric read removal. CLC analysis assigned these reads to 1239 OTUs at >97% similarity, and after selecting OTUs present in over 0.05% (n = 105) of the reads, these were identified from 7 phyla, 9 classes, 24 orders, 33 families, and 62 genera (see Supplementary File 1).
3.4. Core Microbial Composition
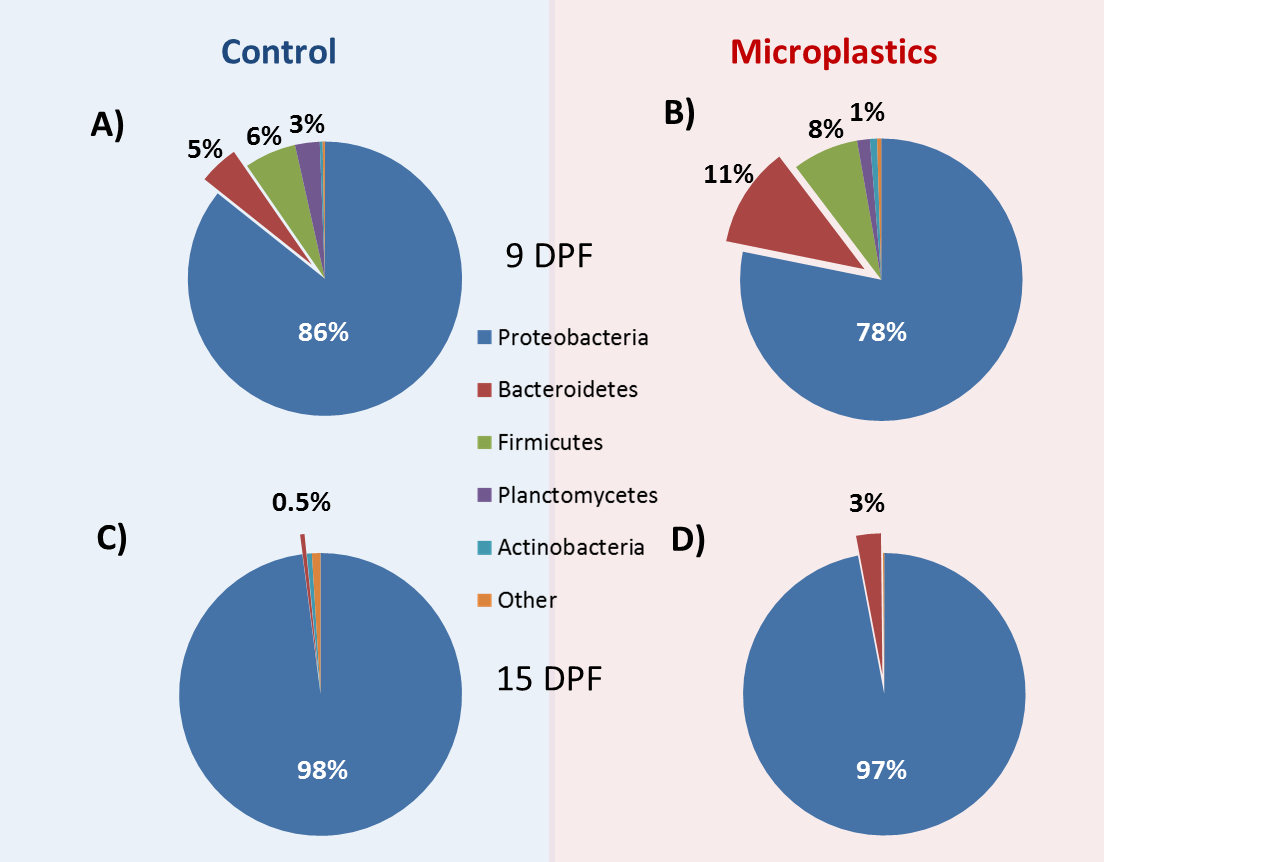
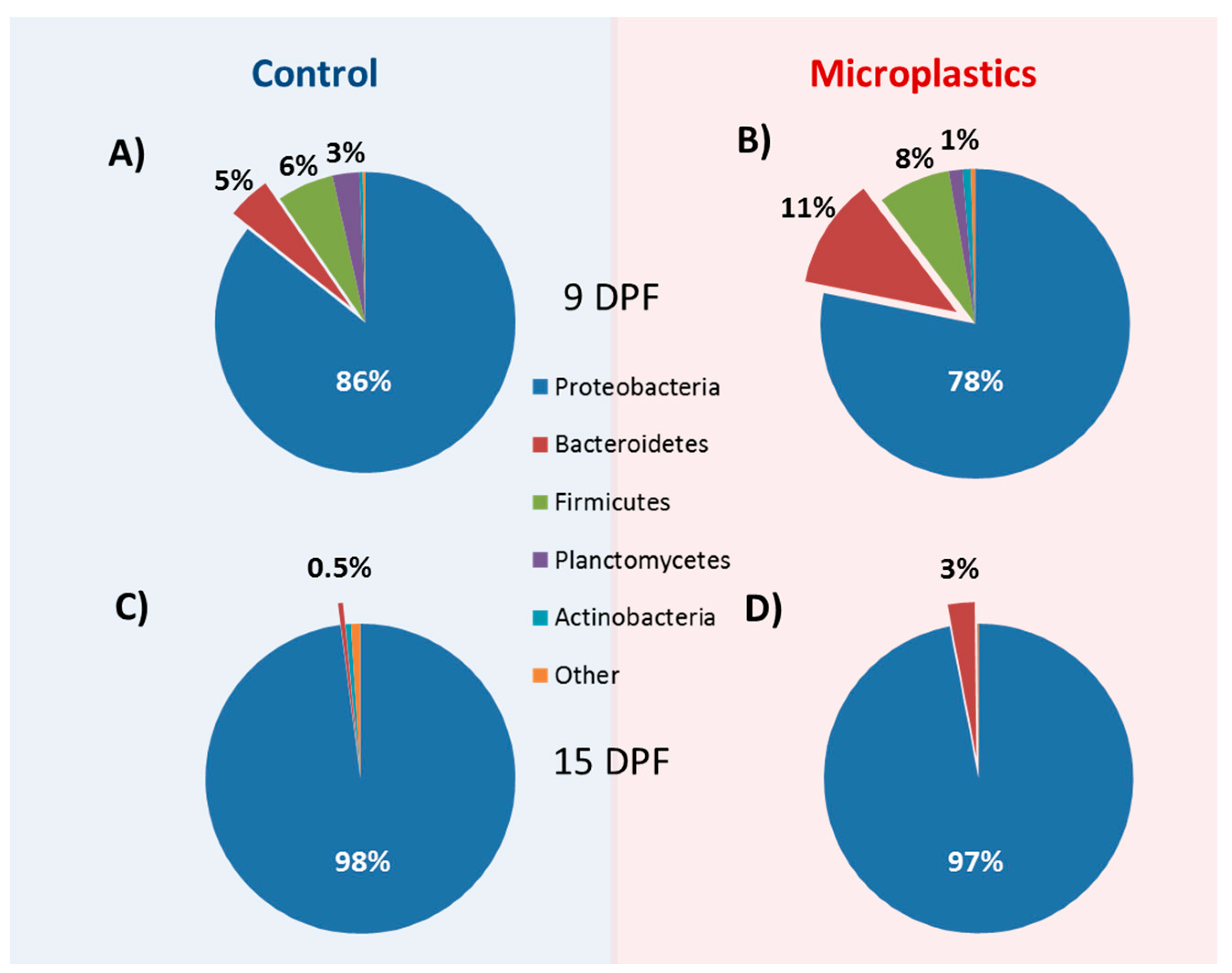
In this study, we describe the core microbiome as taxa that are present in all 24 of the samples sequenced, regardless of treatment and time point. At the phylum level, Proteobacteria had a consistent dominating presence as the most abundant phylum (90%) across all samples and treatments. Indeed, the next most abundant ubiquitous phyla, Bacteroidetes and Cyanobacteria, only each averaged 4% of the total reads in all samples (Figure 3). At the genus level, Aeromonas, Pseudomonas, and Vibrio were overall the most abundant and were among the 17 genera identified as the core microbiome common (Supplementary File 1). Thus, while there were some marked differences in microbial composition across samples and treatments (see below), there was a relatively strong presence of two major phyla and several genera in the larval zebrafish microbiome.
3.5. Microbial Community Changes through Ontogeny
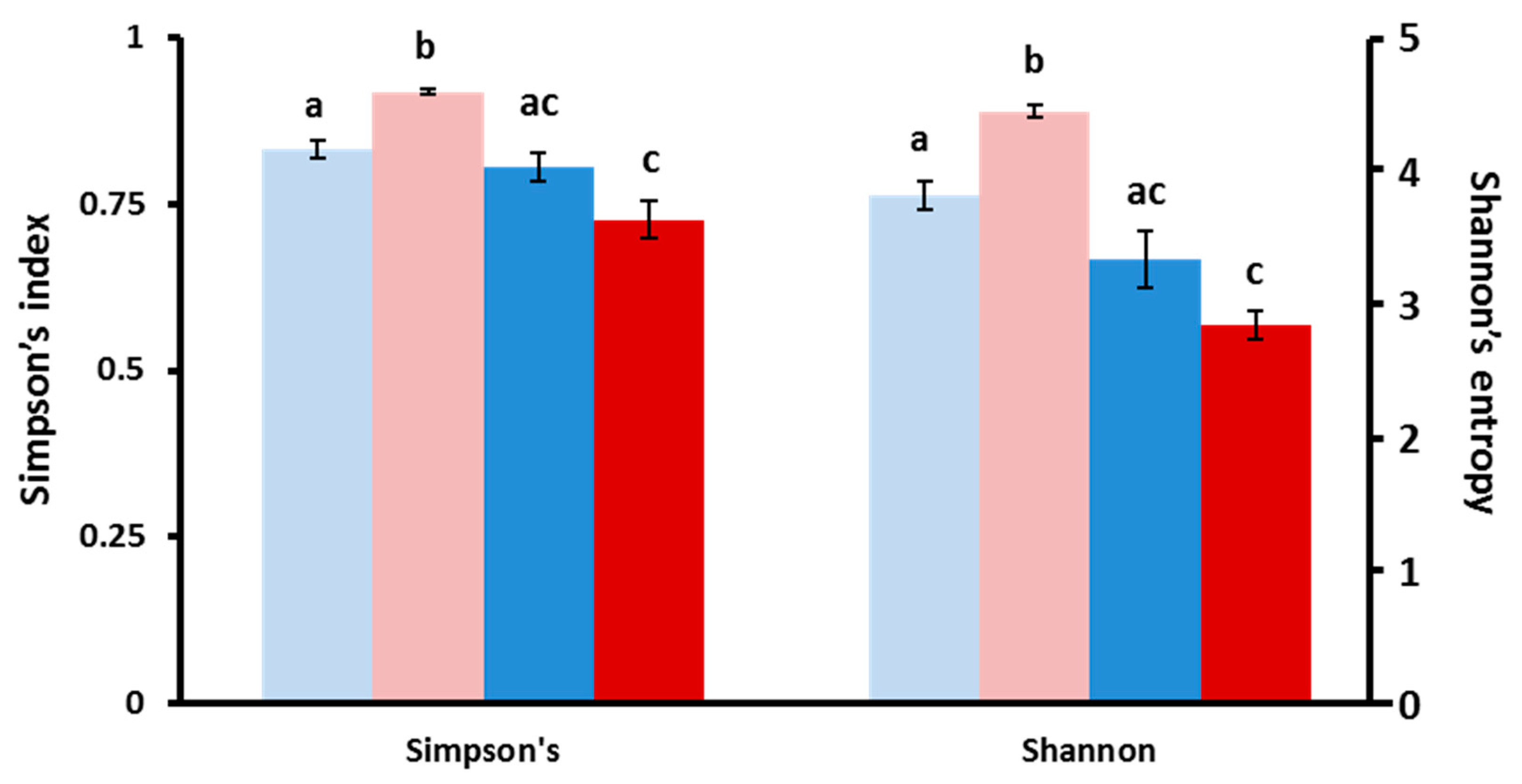
In order to assess the differences in microbiota across treatments, we assessed the relative diversity of the samples. First, we normalized for sequencing depth and rarified all samples to 37,895 reads to examine changes in community structure. Overall, as the fish aged, we saw a relative decrease in community diversity (Figure 4). Although the general pattern applies to both MP and control treatments, it was only statistically significant for all three indicators used in MP fish (observed OTUs, Shannon and Simpson indices), while only the total observed OTU index was significantly decreased in older control fish. However, when we delve into the specifics, we see the number of ubiquitous phyla decreasing from five to only two, Bacteriodetes and Proteobacteria, during the 9 to 15 dpf transition (Supplementary File 1). Incidentally, the abundance of these two phyla increased with the age of the fish, while others, such as Firmicutes and Planctomycetes, were reduced in older fish. These trends were further confirmed as several genera of the Burkholderiaceae and Comamonadaceae family, for example, markedly decreased in older fish (Supplementary File 1). Overall, there was strong evidence of an ontogenic shift in microbial fauna in the developing fish.
3.6. MP Exposure Disturbs the Microbiome
While the age of the fish appears to have an influence on the microbiota composition within each treatment group, we were mostly interested in the differences within and between treatments. We compared beta diversity differences among our samples using permutational multivariate analysis (PERMANOVA). Overall, microbial communities were different across treatments/exposures (PERMANOVA: F = 6.16, p = 3 × 10−5) and particularly according to the age of the fish (PERMANOVA: p = 3 × 10−5). However, there was no overall significant impact of MP treatment compared to control when combining both time points (PERMANOVA: F = 1.11, p = 0.342). When examined within age groups, only 9 dpf fish showed a significant difference in bacterial communities in MP-treated fish compared to controls (p = 0.013). However, at both time points, there was an increased abundance of Bacteroidetes in MP fish, as well as changes in the abundance of several genera (Supplementary File 1). This overall suggests that, while larval zebrafish undergo broad ontogenic microbial changes over the exposure period, there is an evident effect of MP on the microbiome of larval fish, particularly during early development.
4. Discussion
Plastic pollution is quickly becoming one of the most concerning forms of anthropogenic pollution, with numerous studies indicating increased levels of MP in our water systems [62,63,64]. MP can accumulate inside aquatic species, rapidly leading to bioaccumulation and homeostatic imbalances [65,66]. For example, wild fish are likely encountering MP chronically and thus serve as excellent models for the study of MP toxicity in a living system [67]. A number of studies have demonstrated that MP pollution is indeed harmful, even under acute exposures, with polystyrene MP causing gut dysbiosis, inflammation, oxidative stress, and impaired swimming performance in adult zebrafish [41,68]. However, exposure to polyethylene MP has shown conflicting results in adult zebrafish, with some suggesting that polyethylene MP pose little to no harm [69], while others describe signs of oxidative stress and inflammation [16]. Here, we present additional evidence that there is a limited impact of acute MP exposure on developing larval zebrafish. Indeed, ten days of exposure to supra-ecological concentrations of MP does not seem to affect the overall metabolism of the fish and has a limited impact on inflammatory molecular responses, but it seems to trigger an elevated ROS response in the larvae and a concurrent dysbiosis of the larval microbiome.
In order to assess the effect of MP ingestion at the whole organism level, we monitored larval oxygen consumption in a closed respirometry system. Indeed, changes in oxygen consumption could provide an indication of changes in the metabolic status of the animal, including changes in immunological activity and physiological stress [70,71]. Overall, larval oxygen consumption rates were in line with previous studies, with an evident increase in oxygen consumption as the larvae ages [48]. However, we were unable to detect any differences in oxygen consumption between age-matched control and MP-exposed fish, suggesting no global effects on the larval metabolic function. Increased OCRs may indicate the presence of physiological stressors such as widespread inflammation or decreased nutrient absorption [72]. Although similar oxygen consumption rates could indicate a lack of response, we cannot rule out a localized immune response to MP in the fish which may not have broad metabolic effects [13,44]. To further investigate this possibility, we examined the expression of several immune response genes following distinct durations of MP exposure.
Early on in larval zebrafish development, the immune system relies entirely on the innate response for protection [73]. Though efficient, this system requires activation via several pattern recognition receptors that recognize specific exogenous molecules and trigger the release of inflammatory cytokines [22]. We hypothesized that these receptors could be activated by polyethylene particles and thus decided to examine the expression of two genes involved in the broad spectrum MyD88-dependent pathway. MyD88 is largely responsible for the signal transmission of toll-like receptors associated with inflammation, ultimately upregulating cytokine NF-Kβ [74,75,76]. We were unable to find any significant difference in MyD88 or NF-Kβ expression between either of the time points and treatments, suggesting an absence of generalized inflammation via this pathway. However, it is possible that, given the broad tissue distribution of these markers, our approach measuring whole body gene expression levels may mask small localized changes in intestinal tissues. Considering this caveat, we investigated two cytokines (ccl20 and cxcl8a) closely associated with the gut region to provide a more targeted assessment. Both cytokines have been associated with localized gut inflammation in zebrafish in various contexts, which may identify them as key contributors to gut inflammation [77,78]. However, once again, we did not detect any differences in gene expression, further suggesting that intestinal inflammation does not occur in larval zebrafish exposed to MP.
Despite the apparent lack of inflammation, we suspected that MP in the gut could lead to increased oxidative stress resulting from physical abrasion of enterocytes. Therefore, we examined the transcript accumulation of two intestinal antioxidants found in the intestine of larval zebrafish: liver fatty-acid binding protein (L-FABP) and superoxide dismutase 1 (SOD1). L-FABP mitigates hepatic and intestinal ROS production by binding to fatty acid metabolites and heme ligands [79]. Similarly, SOD1 limits ROS using zinc and copper ions, producing non-reactive hydrogen and oxygen atoms [80]. We observed an increase in L-FABP expression during the MP 9–15 dpf time period; however, there was no observable difference in SOD1 expression, possibly due to the wide expression of SOD1 in contrast to a more tissue-specific expression of L-FABP. Together, as previously suggested, our results suggest that MP exposure promotes increased ROS in the zebrafish gut [16,28,81].
Microbiome
The gut of the zebrafish is quickly colonized by microorganisms during the early phases of larval development following the completion of the intestinal tract, resulting in a microbial community known as the “microbiome”. The microbiome is primarily colonized by bacteria that originate from the chorion of the larvae and the surrounding environment during the initial feeding event [82]. Consequently, variability in environmental conditions has made it complex to assess the composition of a core intestinal microbiome in zebrafish, particularly when considering the rapid development of these fish and the variability among fish facilities and strains [83,84]. With these aspects in mind, we ensured that zebrafish larvae were treated equally during our two periods of MP exposure (5–9 dpf and 5–15 dpf) to identify potential time-sensitive responses to polyethylene MP.
As the fish aged, the diversity of the microbiome was significantly reduced. Furthermore, trends of dominant genera changed from 9 dpf to 15 dpf in both the absence and presence of MP. In all fish, dominant genera such as Variovorax increased with age, while others such as Rheinheimera disappeared. When comparing the endpoints of both treatments, we were able to identify a decrease in Aeromonas sp. and an increase in Vibrio sp., a change which may be indicative of natural microbiome development [85]. Increased microbiome diversity is expected to occur at approximately 10 dpf as the zebrafish larvae transition from larvae to juveniles, meaning that this period of development is naturally highly dynamic, even in the absence of MP or other contaminants [84]. We were able to detect two distinct microbial community profiles between half of our samples in each of the MP treatments, which suggests that our time points represent this transitional state and may have increased the susceptibility to dysbiosis via MP exposure.
Larval microbiota was overall more affected by MP at 9 dpf than 15 dpf, indicating that dysbiosis in response to this pollutant may be time- or life stage-sensitive. In contrast to older phases of development, larval zebrafish seem to experience greater gut microbial shifts due to environmental factors [84,86,87]. However, at the phylum level, MP induced an increase in Bacteroidetes at both life-stages and a decrease in Actinobacteria at 15 dpf, a combination that has been found in intestinal inflammation pathologies [88]. Furthermore, some members of the sub-genera Bacteroids have also had their presence associated with various human metabolic diseases such as obesity, autism, type II diabetes, and colorectal cancer [34,35,36,89]. Therefore, although MP exposure only transiently affects bacterial communities, these changes could negatively impact the health of the fish during these transitional life stages.
Interestingly, 15 dpf MP larvae demonstrate concurrently a higher abundance of Bacteriodetes and an increase in ROS, as evidenced by an increase in L-FABP expression compared to control fish at 15 dpf. While we can only speculate regarding the interaction between ROS production and increased abundance of this bacterial phylum, it is possible that increased oxidative stress in MP fish promotes the growth of these bacteria. Indeed, while cells of many micro and macro-organisms are typically affected by ROS [68,90,91], some microbial species, including members of the Bacteriodetes, have effective defense mechanisms against damaging oxidative stress, which may give them a selective advantage over other micro-organisms [92,93,94]. Alternatively, it is possible that the proliferation of these bacteria may be the underlying cause of oxidative stress. Regardless of the proximate causal agent, we confirm that MP exposure is detrimental to larval zebrafish.
5. Conclusions
MP exposure is an environmental concern, as virtually every single ecosystem is plagued with these stable microscopic particles. Here, we confirm that MP exposure negatively affects a freshwater organism as it can result in microbial dysbiosis and increased oxidative stress in larval zebrafish. These effects were asynchronous, with pronounced dysbiosis after only four days of exposure and a subsequent increase in ROS after longer exposure. Furthermore, we were unable to identify any determinants of metabolic distress or inflammation across three independent time points, which suggests that, during early development, MP toxicity may be localized (guts) and does not occur at the whole organism level. Our time course also suggests that the zebrafish gut is most susceptible to MP-induced dysbiosis following the transition from the larval to juvenile state and that, by promoting changes in the microbial communities, such as the proliferation of potentially pathogenic bacteria, MP has the potential to negatively affect fish health at a particularly vulnerable phase of development. Overall, our results corroborate the accumulating evidence that aquatic organisms are negatively affected by MP exposure, though the consequences at the tissue and whole organismal level remain complex and should certainly remain an active area of research.
Supplementary Materials
The following are available online at https://www.mdpi.com/2305-6304/8/3/55/s1, file 1: Differential microbial abundance after 4 and 10 days of MP exposure in larval zebrafish.
Author Contributions
Conceptualization, C.M.R.L.; Formal analysis, N.K. and C.M.R.L.; Funding acquisition, B.J.C. and C.M.R.L.; Investigation, N.K. and C.N.; Methodology, N.K. and B.F.A.; Supervision, B.J.C., B.F.A. and C.M.R.L.; Writing—Original draft, N.K. and C.M.R.L.; Writing—Review & editing, N.K., B.J.C. and B.F.A. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by individual NSERC funding to C.M.R.L., B.A. and B.J.C., as well as collaborative CFI funding to C.M.R.L. and B.J.C.
Acknowledgments
We thank O.E. Elebute for guidance in the microbiome analysis.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Geyer, R.; Jambeck, J.R.; Law, L.K. Production, Use, and Fate of All Plastics Ever Made. Sci. Adv. 2017, 3, e1700782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnes, D.K.A.; Galgani, F.; Thompson, C.R.; Barlaz, M. Accumulation and Fragmentation of Plastic Debris in Global Environments. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2009, 364, 1985–1998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, P.K.; Hakkarainen, M.; Varma, I.K.; Albertsson, A.C. Degradable Polyethylene: Fantasy or Reality. Environ. Sci. Technol. 2011, 45, 4217–4227. [Google Scholar] [CrossRef] [PubMed]
- Andrady, A.L. Microplastics in the Marine Environment. Mar. Pollut. Bull. 2011, 62, 1596–1605. [Google Scholar] [CrossRef] [PubMed]
- Cheshire, A.; Ellik, A.; Barbière, J.; Cohen, Y.; Evans, S.; Jarayabhand, S.; Jeftic, L.; Jung, R.T.; Kinsey, S.; Kusui, E.T.; et al. UNEP/IOC Guidelines on Survey and Monitoring of Marine Litter; Regional Seas Reports and Studies No. 186 IOC Technical Series No. 83. UNEP Regional Seas Reports and Studies; Intergovernmental Oceanographic Commission: Paris, France, 2009. [Google Scholar]
- Frias, J.P.; Nash, R. Microplastics: Finding a consensus on the definition. Mar. Pollut. Bull. 2019, 138, 145–147. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, R.V.; Gutow, L.; Thompson, R.C.; Thiel, M. Microplastics in the Marine Environment: A Review of the Methods Used for Identification and Quantification. Environ. Sci. Technol. 2012, 46, 3060–3075. [Google Scholar] [CrossRef]
- Obbard, R.W.; Sadri, S.; Wong, Q.Y.; Khitun, A.A.; Baker, I.; Thompson, R.C. Global Warming Releases Microplastic Legacy Frozen in Arctic Sea Ice. Earths Future 2014, 2, 315–320. [Google Scholar] [CrossRef]
- Sun, J.; Dai, X.; Wang, Q.; Loosdrecht, V.M.C.M.; Ni, B.J. Microplastics in Wastewater Treatment Plants: Detection, Occurrence and Removal. Water Res. 2019, 152, 21–37. [Google Scholar] [CrossRef]
- Li, J.; Liu, H.; Chen, J.P. Microplastics in Freshwater Systems: A Review on Occurrence, Environmental Effects, and Methods for Microplastics Detection. Water Res. 2018, 137, 362–374. [Google Scholar] [CrossRef]
- Shahul, H.F.; Bhatti, M.S.; Anuar, N.; Anuar, N.; Mohan, P.; Periathamby, A. Worldwide Distribution and Abundance of Microplastic: How Dire Is the Situation? Waste Manag. Res. 2018, 36, 873–897. [Google Scholar] [CrossRef]
- Vermaire, C.J.; Pomeroy, C.; Herczegh, M.S.; Haggart, O.; Murphy, M. Microplastic Abundance and Distribution in the Open Water and Sediment of the Ottawa River, Canada, and Its Tributaries. FACETS. 2017, 2, 301–314. [Google Scholar] [CrossRef] [Green Version]
- LeMoine, C.M.R.; Kelleher, B.M.; Lagarde, R.; Northam, C.; Elebute, O.O.; Cassone, B.J. Transcriptional Effects of Polyethylene Microplastics Ingestion in Developing Zebrafish (Danio rerio). Environ. Pollut. 2018, 243 Pt A, 591–600. [Google Scholar] [CrossRef]
- Wright, S.L.; Thompson, R.C.; Galloway, T.S. The Physical Impacts of Microplastics on Marine Organisms: A Review. Environ. Pollut. 2013, 178, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Chambers, T.K.; Chen, Z.C.; Crawford, P.A.; Fu, X.; Burgess, S.C.; Lai, L.; Leone, T.C.; Kelly, D.P.; Finck, B.N. Liver-Specific PGC-1beta Deficiency Leads to Impaired Mitochondrial Function and Lipogenic Response to Fasting-Refeeding. PLoS ONE 2012, 7, e52645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Zhang, Y.; Deng, Y.; Jiang, W.; Zhao, Y.; Geng, J.; Ding, L.; Ren, H. Uptake and Accumulation of Polystyrene Microplastics in Zebrafish (Danio rerio) and Toxic Effects in Liver. Environ. Sci. Technol. 2016, 50, 4054–4060. [Google Scholar] [CrossRef]
- Pitt, J.A.; Kozal, J.S.; Jayasundara, N.; Massarsky, A.; Trevisan, R.; Geitner, N.; Wiesner, M.; Levin, E.D.; Di Giulio, R.T. Uptake, Tissue Distribution, and Toxicity of Polystyrene Nanoparticles in Developing Zebrafish (Danio rerio). Aquat. Toxicol. 2018, 194, 185–194. [Google Scholar] [CrossRef]
- Pedà, C.; Leteria, C.; Fossi, C.M.; Gai, F.; Andaloro, F.; Genovese, L.; Perdichizzi, A.; Romeo, T.; Maricchiolo, G. Intestinal Alterations in European Sea Bass Dicentrarchus labrax (Linnaeus, 1758) Exposed to Microplastics: Preliminary Results. Environ. Pollut. 2016, 212, 251–256. [Google Scholar] [CrossRef]
- Jeong, C.B.; Won, E.J.; Kang, H.-M.; Lee, M.C.; Hwang, D.S.; Hwang, U.K.; Zhou, B.; Souissi, S.; Lee, S.J.; Lee, J.S. Microplastic Size-Dependent Toxicity, Oxidative Stress Induction, and p-JNK and p-P38 Activation in the Monogonont Rotifer (Brachionus koreanus). Environ. Sci. Technol. 2016, 50, 8849–8857. [Google Scholar] [CrossRef]
- Blaser, H.; Dostert, C.; Mak, T.W.; Brenner, D. TNF and ROS crosstalk in inflammation. Trends Cell Biol. 2016, 26, 249–261. [Google Scholar] [CrossRef]
- Duan, X.D.; Feng, L.; Jiang, W.D.; Wu, P.; Liu, Y.; Kuang, S.Y.; Tang, T.; Tang, W.N.; Zhang, Y.A.; Zhou, X.Q. Dietary soybean β-conglycinin suppresses growth performance and inconsistently triggers apoptosis in the intestine of juvenile grass carp (Ctenopharyngodon idella) in association with ROS-mediated MAPK signalling. Aquac. Nutr. 2019, 25, 770–782. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, C.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory Responses and Inflammation-Associated Diseases in Organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaser, A.; Tilg, H. “Metabolic Aspects” in Inflammatory Bowel Diseases. Curr. Drug Deliv. 2012, 9, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Weisshof, R.; Chermesh, I. Micronutrient Deficiencies in Inflammatory Bowel Disease. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 576–581. [Google Scholar] [CrossRef] [PubMed]
- Slyepchenko, A.; Maes, M.; Machado-Vieira, R.; Anderson, G.; Solmi, M.; Sanz, Y.; Berk, M.; Köhler, C.A.; Carvalho, A.F. Intestinal dysbiosis, gut hyperpermeability and bacterial translocation: Missing links between depression, obesity and type 2 diabetes. Curr. Pharm Des. 2016, 22, 6087–6106. [Google Scholar] [CrossRef] [PubMed]
- Lynch, S.V.; Pedersen, O. The human intestinal microbiome in health and disease. N. Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef] [Green Version]
- Kim, A.; Kim, N.; Roh, J.H.; Chun, W.-K.; Ho, D.T.; Lee, Y.; Kim, D.-H. Administration of antibiotics can cause dysbiosis in fish gut. Aquaculture 2019, 512, 734330. [Google Scholar] [CrossRef]
- Qiao, R.; Cheng, S.; Lu, Y.; Zhang, Y.; Ren, H.; Lemos, B. Microplastics Induce Intestinal Inflammation, Oxidative Stress, and Disorders of Metabolome and Microbiome in Zebrafish. Sci. Total Environ. 2019, 662, 246–253. [Google Scholar] [CrossRef]
- Wan, Z.; Wang, C.; Zhou, J.; Shen, M.; Wang, X.; Fu, Z.; Jin, Y. Effects of polystyrene microplastics on the composition of the microbiome and metabolism in larval zebrafish. Chemosphere 2019, 217, 646–658. [Google Scholar] [CrossRef]
- Fackelmann, G.; Sommer, S. Microplastics and the gut microbiome: How chronically exposed species may suffer from gut dysbiosis. Mar. Pollut. Bull. 2019, 143, 193–203. [Google Scholar] [CrossRef]
- Lu, L.; Luo, T.; Zhao, Y.; Cai, C.; Fu, Z.; Jin, Y. Interaction between microplastics and microorganism as well as gut microbiota: A consideration on environmental animal and human health. Sci. Total Environ. 2019, 667, 94–100. [Google Scholar] [CrossRef]
- Luo, T.; Wang, C.; Pan, Z.; Jin, C.; Fu, Z.; Jin, Y. Maternal polystyrene microplastic exposure during gestation and lactation altered metabolic homeostasis in the dams and their F1 and F2 offspring. Environ. Sci. Technol. 2019, 53, 10978–10992. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Chen, Q.-L.; An, X.-L.; Yang, X.-R.; Christie, P.; Ke, X.; Wu, L.H.; Zhu, Y.G. Exposure of soil collembolans to microplastics perturbs their gut microbiota and alters their isotopic composition. Soil Biol. Biochem. 2018, 116, 302–310. [Google Scholar] [CrossRef]
- Sobhani, I.; Tap, J.; Roudot-Thoraval, F.; Roperch, J.P.; Letulle, S.; Langella, P.; Corthier, G.; Nhieu, J.T.V.; Furet, J.P. Microbial Dysbiosis in Colorectal Cancer (CRC) Patients. PLoS ONE 2011, 6, e16393. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.W.; Sanderson, J.D.; Churcher, C.; Parkes, G.C.; Hudspith, B.N.; Rayment, N.; Brostoff, J.; Parkhill, J.; Gordon, D.; Petrovska, L. High-Throughput Clone Library Analysis of the Mucosa-Associated Microbiota Reveals Dysbiosis and Differences between Inflamed and Non-Inflamed Regions of the Intestine in Inflammatory Bowel Disease. BMC Microbiol. 2011, 11, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Ma, C.; Han, L.; Nawaz, M.; Gao, F.; Zhang, X.; Yu, P.; Zhao, C.; Li, L.; Zhou, A.; et al. Molecular Characterisation of the Faecal Microbiota in Patients with Type II Diabetes. Curr. Microbiol. 2010, 61, 69–78. [Google Scholar] [CrossRef]
- Reese, A.T.; Cho, E.H.; Kiltzman, B.; Nichols, S.P.; Wisniewski, N.A.; Villa, M.M.; Durand, H.K.; Jiang, S.; Midani, F.S.; Nimmagadda, S.N.; et al. Antibiotic-Induced Changes in the Microbiota Disrupt Redox Dynamics in the Gut. eLife 2018, 7, e35987. [Google Scholar] [CrossRef]
- Gil, D.; Alfonso, I.S.; Pérez, R.L.; Muriel, J.; Monclús, R. Harsh Conditions during Early Development Influence Telomere Length in an Altricial Passerine: Links with Oxidative Stress and Corticosteroids. J. Evol. Biol. 2019, 32, 111–125. [Google Scholar] [CrossRef] [Green Version]
- Jiang, N.M.; Cowan, M.; Moonah, S.N.; Petri, W.A. The Impact of Systemic Inflammation on Neurodevelopment. Trends Mol. Med. 2018, 24, 794–804. [Google Scholar] [CrossRef]
- Mead, K.S.; Denny, M.W. The Effects of Hydrodynamic Shear Stress on Fertilization and Early Development of the Purple Sea Urchin Strongylocentrotus purpuratus. Biol. Bull. 1995, 188, 46–56. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.; Xia, J.; Pan, Z.; Yang, J.; Wang, W.; Fu, Z. Polystyrene Microplastics Induce Microbiota Dysbiosis and Inflammation in the Gut of Adult Zebrafish. Environ. Pollut. 2018, 235, 322–329. [Google Scholar] [CrossRef]
- Song, Y.K.; Hong, H.S.; Jang, M.; Kang, J.-H.; Kwon, O.Y.; Han, G.M.; Shim, W.J. Large Accumulation of Micro-Sized Synthetic Polymer Particles in the Sea Surface Microlayer. Environ. Sci. Technol. 2014, 48, 9014–9021. [Google Scholar] [CrossRef] [PubMed]
- Watts, A.J.R.; Lewis, C.; Goodhead, R.M.; Beckett, S.J.; Moger, J.; Tyler, C.R.; Galloway, T.S. Uptake and retention of microplastics by the shore crab Carcinus maenas. Environ. Sci. Technol. 2014, 48, 8823e8830. [Google Scholar] [CrossRef] [PubMed]
- Cole, M.; Lindeque, P.; Fileman, E.; Halsband, C.; Galloway, T.S. The Impact of Polystyrene Microplastics on Feeding, Function and Fecundity in the Marine Copepod Calanus Helgolandicus. Environ. Sci. Technol. 2015, 49, 1130–1137. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.F.; Syberg, K.; Shashoua, Y. Influence of polyethylene microplastic beads on the uptake and localization of silver in zebrafish (Danio rerio). Environ. Pollut. 2015, 206, 73e79. [Google Scholar] [CrossRef] [PubMed]
- Westerfield, M. The Zebrafish Book; University of Oregon Press: Eugene, OR, USA, 1995. [Google Scholar]
- Harianto, J.; Carey, N.; Byrne, M. respR—An R package for the manipulation and analysis of respirometry data. Methods Ecol. Evol. 2019, 10, 912–920. [Google Scholar] [CrossRef] [Green Version]
- Rombough, P.; Drader, H. Hemoglobin Enhances Oxygen Uptake in Larval Zebrafish (Danio rerio) but Only under Conditions of Extreme Hypoxia. J. Exp. Biol. 2009, 212 Pt 6, 778–784. [Google Scholar] [CrossRef] [Green Version]
- Northam, C.; LeMoine, C.M.R. Metabolic Regulation by the PGC-1α and PGC-1β Coactivators in Larval Zebrafish (Danio rerio). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2019, 234, 60–67. [Google Scholar] [CrossRef]
- Untergasser, A.S.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3--new capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [Green Version]
- McCurley, A.T.; Callard, G.V. Characterization of housekeeping genes in zebrafish: Male-female differences and effects of tissue type, developmental stage and chemical treatment. BMC Mol. Biol. 2008, 9, 102. [Google Scholar] [CrossRef] [Green Version]
- Ibarra, M.S.; Etichetti, C.B.; Benedetto, C.D.; Rosano, G.L.; Margarit, E.; Sal, G.D.; Mione, M.; Girardini, J. Dynamic Regulation of Pin1 Expression and Function during Zebrafish Development. PLoS ONE 2017, 12, e0175939. [Google Scholar] [CrossRef]
- Vaart, V.D.M.; Soest, V.J.J.; Spaink, H.P.; Meijer, A.H. Functional analysis of a zebrafish myd88 mutant identifies key transcriptional components of the innate immune system. Dis. Model Mech. 2013, 6, 841–854. [Google Scholar] [CrossRef] [Green Version]
- Rotman, J.; Gils, V.W.; Butler, D.; Spaink, H.P.; Meijer, A.H. Rapid screening of innate immune gene expression in zebrafish using reverse transcription-multiplex ligation-dependent probe amplification. BMC Res. Notes 2011, 4, 196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, Y.; Zhang, Y.; Han, L.; He, Q.; Hou, H.; Han, J.; Wang, X.; Li, C.; Cen, J.; Liu, K. Oxidative stress-mediated developmental toxicity induced by isoniazide in zebrafish embryos and larvae. J. Appl. Toxicol. 2017, 37, 842–852. [Google Scholar] [CrossRef] [PubMed]
- Ryu, B.; Kim, C.Y.; Oh, H.; Kim, U.; Kim, J.; Jung, C.R.; Lee, B.H.; Lee, S.; Chang, S.N.; Lee, J.M.; et al. Development of an alternative zebrafish model for drug-induced intestinal toxicity. J. Appl. Toxicol. 2018, 38, 259–273. [Google Scholar] [CrossRef] [PubMed]
- Brugman, S.; Witte, M.; Scholman, R.C.; Klein, M.R.; Boes, M.; Nieuwenhuis, E.E. T Lymphocyte–Dependent and–Independent Regulation of Cxcl8 Expression in Zebrafish Intestines. J. Immunol. 2014, 192, 484–491. [Google Scholar] [CrossRef] [Green Version]
- Livak, J.K.; Schmittgen, D.T. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2-ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef] [Green Version]
- Cassone, B.J.; Grove, H.C.; Elebute, O.; Villanueva, S.M.; LeMoine, C.M. Role of the intestinal microbiome in low-density polyethylene degradation by caterpillar larvae of the greater wax moth, Galleria mellonella. Proc. Biol. Sci. 2020, 287, 20200112. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef]
- Gallo, F.; Fossi, C.; Weber, R.; Santillo, D.; Sousa, J.; Ingram, I.; Nadal, A.; Romano, D. Marine Litter Plastics and Microplastics and Their Toxic Chemicals Components: The Need for Urgent Preventive Measures. Environ. Sci. Eur. 2018, 30. [Google Scholar] [CrossRef]
- Galloway, S.T.; Lewis, C.N. Marine Microplastics Spell Big Problems for Future Generations. Proc. Natl. Acad. Sci. USA 2016, 113, 2331–2333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, S.; Chatterjee, S. Microplastic Pollution, a Threat to Marine Ecosystem and Human Health: A Short Review. Environ. Sci. Pollut. Res. Int. 2017, 24, 21530–21547. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Gundlach, M.; Shouye, Y.; Jiang, J.; Velki, M.; Yin, D.; Hollert, H. Quantitative Investigation of the Mechanisms of Microplastics and Nanoplastics toward Zebrafish Larvae Locomotor Activity. Sci. Total. Environ. 2017, 584–585, 1022–1031. [Google Scholar] [CrossRef] [PubMed]
- Mazurais, D.; Ernande, B.; Quazuguel, P.; Severe, A.; Huelvan, C.; Madec, L.; Mouchel, O.; Soudant, P.; Robbens, J.; Huvet, A.; et al. Evaluation of the Impact of Polyethylene Microbeads Ingestion in European Sea Bass (Dicentrarchus labrax) Larvae. Mar. Environ. Res. 2015, 112 Pt A, 78–85. [Google Scholar] [CrossRef] [Green Version]
- Hill, A.J.; Teraoka, H.; Heideman, W.; Peterson, R.E. Zebrafish as a Model Vertebrate for Investigating Chemical Toxicity. Toxicol. Sci. 2005, 86, 6–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiang, L.; Cheng, J. Exposure to Microplastics Decreases Swimming Competence in Larval Zebrafish (Danio rerio). Ecotoxicol. Environ. Saf. 2019, 176, 226–233. [Google Scholar] [CrossRef]
- Reniere, M.L. Reduce, Induce, Thrive: Bacterial Redox Sensing during Pathogenesis. J. Bacteriol. 2018, 200, e00128-18. [Google Scholar] [CrossRef] [Green Version]
- Bonneaud, C.; Wilson, R.S.; Seebacher, F. Immune-Challenged Fish Up-Regulate Their Metabolic Scope to Support Locomotion. PLoS ONE 2016, 11, e0166028. [Google Scholar] [CrossRef] [Green Version]
- Yuan, M.; Yan, C.; Ying, Y.H.; Lu, W. Behavioral and Metabolic Phenotype Indicate Personality in Zebrafish (Danio rerio). Front. Physiol. 2018, 9, 653. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.S.; Cho, H.-J.; Kim, E.; Huh, Y.H.; Kim, H.-J.; Kim, B.; Kang, T.; Lee, J.-S.; Jeong, J. Bioaccumulation of Polystyrene Nanoplastics and Their Effect on the Toxicity of Au Ions in Zebrafish Embryos. Nanoscale 2019, 11, 3200–3207. [Google Scholar] [CrossRef]
- Novoa, B.; Figueras, A. Zebrafish: Model for the Study of Inflammation and the Innate Immune Response to Infectious Diseases. Adv. Exp. Med. Biol. 2012, 946, 253–275. [Google Scholar] [CrossRef] [PubMed]
- Akira, S.; Hoshino, K. Myeloid Differentiation Factor 88–Dependent and –Independent Pathways in Toll-Like Receptor Signaling. J. Infect. Dis. 2003, 187 (Suppl. 2), S356–S363. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-ΚB Signaling in Inflammation. Signal Transduct. Target Ther. 2017, 17023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sar, A.M.; Stockhammer, O.W.; Laan, C.; Spaink, H.P.; Bitter, W.; Meijer, A.H. MyD88 Innate Immune Function in a Zebrafish Embryo Infection Model. Infect. Immun. 2006, 74, 2436–2441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brugman, S. The Zebrafish as a Model to Study Intestinal Inflammation. Dev. Comp. Immunol. 2016, 64, 82–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanyang, L.; Liu, X.; Chen, X.; Yujia, Q.; Jiarong, F.; Jun, S.; Zhihua, R. Application of Zebrafish Models in Inflammatory Bowel Disease. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.; Bonkovsky, H.L.; de Lemos, A.; Burczynski, F.J. Recent Insights into the Biological Functions of Liver Fatty Acid Binding Protein 1. J. Lipid Res. 2015, 56, 2238–2247. [Google Scholar] [CrossRef] [Green Version]
- Maragakis, N.J.; Rothstein, J.D. Motor Neuron Disease: Amyotrophic Lateral Sclerosis. Mol. Neurobiol. 2007, 307–319. [Google Scholar] [CrossRef]
- Jeong, C.B.; Kang, H.M.; Lee, M.C.; Kim, D.H.; Han, J.; Hwang, S.; Souissi, S.; Lee, S.J.; Shin, K.H.; Park, H.G.; et al. Adverse Effects of Microplastics and Oxidative Stress-Induced MAPK/Nrf2 Pathway-Mediated Defense Mechanisms in the Marine Copepod Paracyclopina nana. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Egerton, S.; Culloty, S.; Whooley, J.; Stanton, C.; Ross, P.R. The Gut Microbiota of Marine Fish. Front. Microbiol. 2018, 9, 873. [Google Scholar] [CrossRef]
- Roeselers, G.; Mittge, E.K.; Stephens, Z.W.; Parichy, D.M.; Cavanaugh, C.M.; Guillemin, K.; Rawls, J.F. Evidence for a Core Gut Microbiota in the Zebrafish. ISME J. 2011, 5, 1595–1608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephens, W.Z.; Burns, R.A.; Stagaman, K.; Wong, S.; Rawls, J.F.; Guillemin, K.; Bohannan, B.J. The Composition of the Zebrafish Intestinal Microbial Community Varies across Development. ISME J. 2016, 10, 644–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phelps, D.; Brinkman, N.E.; Keely, S.P.; Annken, E.M.; Catron, T.R.; Betancourt, D.; Wood, C.E.; Espenschied, S.T.; Rawls, J.F.; Tal, T. Microbial Colonization Is Required for Normal Neurobehavioral Development in Zebrafish. Sci. Rep. 2017, 7, 11244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, J.P.; Sapp, M.; Schratzberger, M.; Osborn, M.A. Interactions Between Microorganisms and Marine Microplastics: A Call for Research. Mar. Technol. Soc. J. 2011, 45, 12–20. [Google Scholar] [CrossRef]
- Lu, K.; Ruxia, Q.; Hao, A.; Zhang, Y. Influence of Microplastics on the Accumulation and Chronic Toxic Effects of Cadmium in Zebrafish (Danio rerio). Chemosphere 2018, 202, 514–520. [Google Scholar] [CrossRef]
- Prakash, S.; Rodes, L.; Coussa-Charley, M.; Tomaro-Duchnesneau, C. Gut Microbiota: Next Frontier in Understanding Human Health and Development of Biotherapeutics. Biologics 2011, 5, 71–86. [Google Scholar] [CrossRef] [Green Version]
- Vael, C.; Nelen, V.; Verhulst, L.S.; Goossens, H.; Desager, K.N. Early Intestinal Bacteroides Fragilis Colonisation and Development of Asthma. BMC Pulm. Med. 2008, 8, 19. [Google Scholar] [CrossRef] [Green Version]
- Cadet, J.; Wagner, R.J. DNA Base Damage by Reactive Oxygen Species, Oxidizing Agents, and UV Radiation. Cold Spring Harb. Perspect. Biol. 2013, 5, a012559. [Google Scholar] [CrossRef]
- Dumitrescu, L.; Popescu-Olaru, I.; Cozma, L.; Tulbă, D.; Eugen, M.H.; Ceafalan, L.C.; Gherghiceanu, M.; Popescu, B.O. Oxidative Stress and the Microbiota-Gut-Brain Axis. Oxid. Med. Cell Longev. 2018, 406594. [Google Scholar] [CrossRef] [Green Version]
- Johnson, L.A.; Hug, L.A. Distribution of reactive oxygen species defense mechanisms across domain bacteria. Free Radic. Biol. Med. 2019, 140, 93–102. [Google Scholar] [CrossRef]
- Machado, M.V.; Cortez-Pinto, H. Gut Microbiota and Nonalcoholic Fatty Liver Disease. Ann. Hepatol. 2012, 11, 440–449. [Google Scholar] [CrossRef]
- Suzuki, A.; Ito, M.; Hamaguchi, T.; Mori, H.; Takeda, Y.; Baba, R.; Watanabe, T.; Kurokawa, K.; Asakawa, S.; Hirayama, M.; et al. Quantification of Hydrogen Production by Intestinal Bacteria That Are Specifically Dysregulated in Parkinson’s Disease. PLoS ONE 2018, 13, e0208313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Oxygen consumption rates (OCRs) for microplastic (MP) exposed zebrafish. OCRs of MP fish after 5 days (pink) and 10 days (red) of exposure to MP and the respective age matched control fish (light blue/dark blue). No statistical significance was observed between the MP and control groups; however, there was a significant difference between time points (n = 6, p = 0.015).
Figure 1.
Oxygen consumption rates (OCRs) for microplastic (MP) exposed zebrafish. OCRs of MP fish after 5 days (pink) and 10 days (red) of exposure to MP and the respective age matched control fish (light blue/dark blue). No statistical significance was observed between the MP and control groups; however, there was a significant difference between time points (n = 6, p = 0.015).

Figure 2.
Gene expression in MP and control fish at 9 and 15 dpf. Gene expression analysis of inflammatory (A), microbial (B), and oxidative stress (C) genes in fish exposed to MP for 4 (pink) and 10 days (red) or aged matched control fish (light and dark blue). EF1α/RLP-13a were used as housekeeping genes. Statistically significant differences are indicated with an asterisk (n = 9, p < 0.05).
Figure 2.
Gene expression in MP and control fish at 9 and 15 dpf. Gene expression analysis of inflammatory (A), microbial (B), and oxidative stress (C) genes in fish exposed to MP for 4 (pink) and 10 days (red) or aged matched control fish (light and dark blue). EF1α/RLP-13a were used as housekeeping genes. Statistically significant differences are indicated with an asterisk (n = 9, p < 0.05).

Figure 3.
Relative phylum composition of control and MP-exposed zebrafish larvae. Zebrafish larvae were exposed to control conditions (A,C) and microplastics (B,D) for 4 days (A,B) and 10 days (C,D), respectively. Percentages represent the relative abundance of the most represented phyla.
Figure 3.
Relative phylum composition of control and MP-exposed zebrafish larvae. Zebrafish larvae were exposed to control conditions (A,C) and microplastics (B,D) for 4 days (A,B) and 10 days (C,D), respectively. Percentages represent the relative abundance of the most represented phyla.

Figure 4.
The effect of MP exposure on diversity indices in larval zebrafish. Simpson and Shannon OTU diversity indices obtained through CLC analysis for 9 dpf control (light blue) and microplastics (pink), as well as 15 dpf control (dark blue) and microplastics (red). Significant differences are indicated by different letters (p = 0.05).
Figure 4.
The effect of MP exposure on diversity indices in larval zebrafish. Simpson and Shannon OTU diversity indices obtained through CLC analysis for 9 dpf control (light blue) and microplastics (pink), as well as 15 dpf control (dark blue) and microplastics (red). Significant differences are indicated by different letters (p = 0.05).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
List of primers used in qPCR assays of gene expression.
| Gene | Forward Primer | Reverse Primer | Accession Number |
| ef1α51 | CTTCTCAGGCTGACTGTGC | CCGCTAGCATTACCCTCC | AY422992 |
| Rpl-13a52 | TCTGGAGGACTGTAAGAGGTATGC | AGACGCACAATCTTGAGAGCAG | NM_212784.1 |
| MyD8853 | GAGGCGATTCCAGTAACAGC | GAAAGCATCAAAGGTCTCAGGTG | NM_212814.2 |
| Ccl2054 | ATCAATCTGCGCTAATCCATCAC | TGGTGAACATGCTCATCGTCTT | NM_001113595.1 |
| L-FABP55 | ACGTGGCAGGTTTACGCTCAG | TTGGAGGTGATGGTGAAGTCG | BC164928.1 |
| SOD156 | GTCGTCTGGCTTGTGGAGTG | TGTCAGCGGGCTAGTGCTT | AY324390.1 |
| NF-κβ56 | CCAAATCCCAAAAGGTTAGAGATTT | CCTCTTAGGGCTGAGCGAATT | XM_005156814.2 |
| Cxcl8a57 | TGTTTTCCTGGCATTTCTGAC | TTTACAGTGTGGGCTTGGAGGG | XM_009306855.3 |
| Mus2.150 | TGGTGGACCAGTGTGAAAAA | GGTCCAAAACCCAGCTACAA | XM_021470771.1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kurchaba, N.; Cassone, B.J.; Northam, C.; Ardelli, B.F.; LeMoine, C.M.R. Effects of MP Polyethylene Microparticles on Microbiome and Inflammatory Response of Larval Zebrafish. Toxics 2020, 8, 55. https://doi.org/10.3390/toxics8030055
AMA Style
Kurchaba N, Cassone BJ, Northam C, Ardelli BF, LeMoine CMR. Effects of MP Polyethylene Microparticles on Microbiome and Inflammatory Response of Larval Zebrafish. Toxics. 2020; 8(3):55. https://doi.org/10.3390/toxics8030055
Chicago/Turabian StyleKurchaba, Nicholas, Bryan J. Cassone, Caleb Northam, Bernadette F. Ardelli, and Christophe M. R. LeMoine. 2020. "Effects of MP Polyethylene Microparticles on Microbiome and Inflammatory Response of Larval Zebrafish" Toxics 8, no. 3: 55. https://doi.org/10.3390/toxics8030055
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.