Environmentally Relevant Concentrations of Bisphenol A Interact with Doxorubicin Transcriptional Effects in Human Cell Lines



Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Cultures and Reagents
2.2. Drugs and Treatments
2.3. RNA Isolation and Real-Time PCR
2.4. Statistical Analysis
3. Results
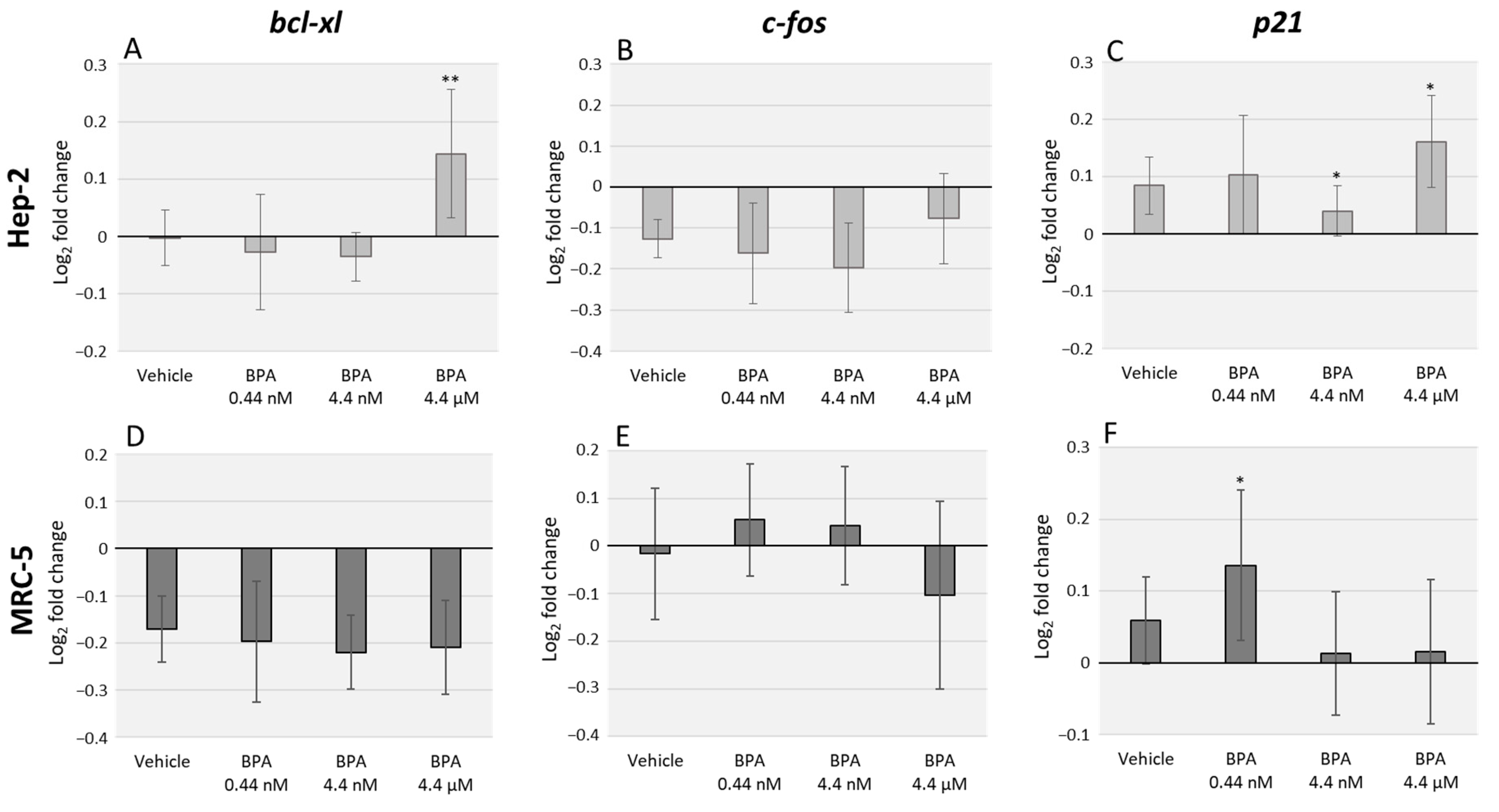
3.1. BPA Alters bcl-xl and p21 Transcription Levels but Does Not Significantly Affect c-fos Expression
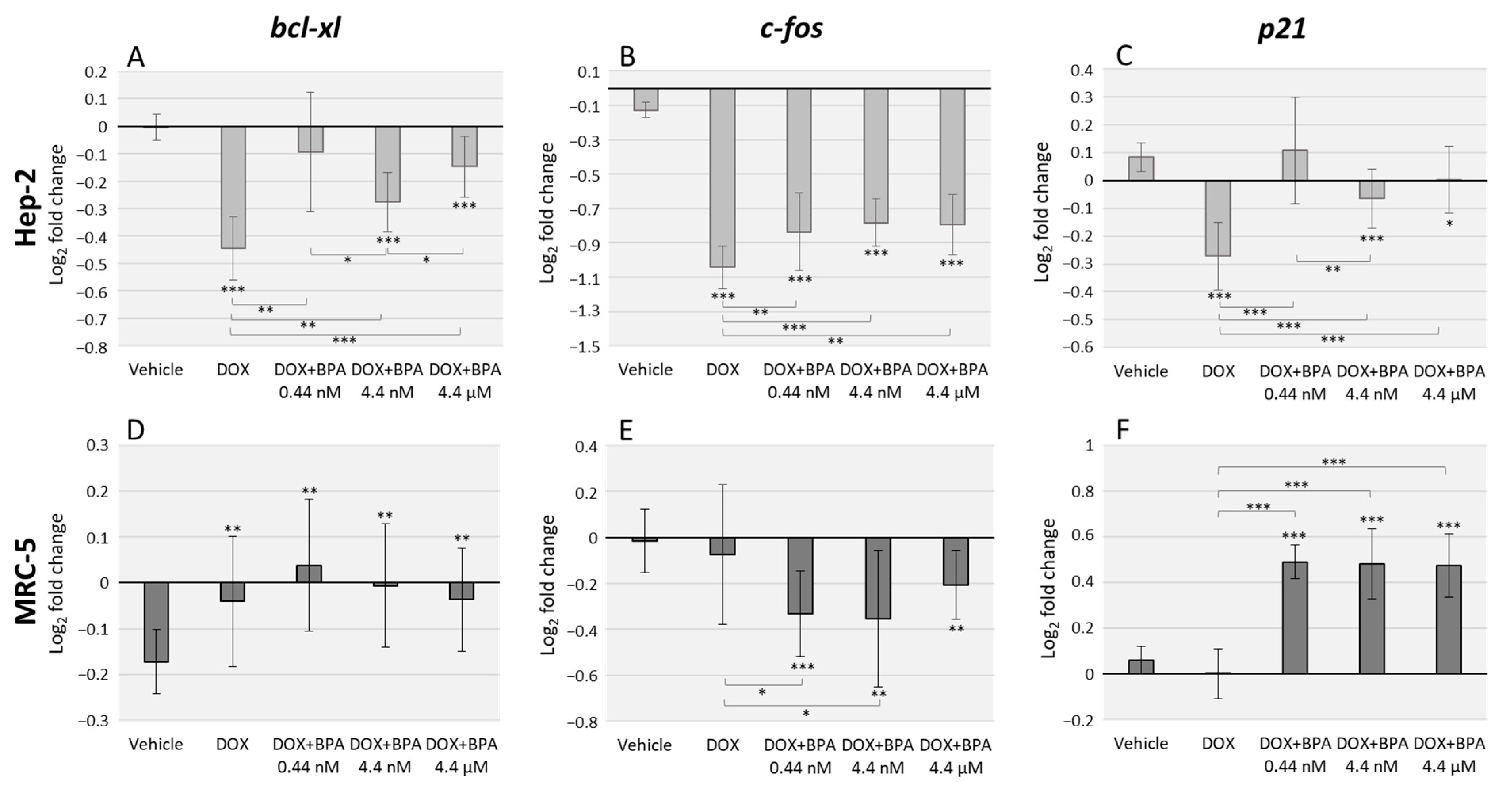
3.2. BPA Interacts with DOX Transcriptional Effects in Hep-2 and MRC-5 Cell Lines with Cell Type Specificity
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Report on the Burden of Endemic Health Care-Associated Infection Worldwide; Clean Care is Safer Care; WHO: Geneva, Switzerland, 2011. [Google Scholar]
- EFSA. Scientific Report of the Endocrine Active Substances Task Force. EFSA J. 2010, 8, 1932. [Google Scholar] [CrossRef]
- Podein, R.J.; Hernke, M.T.; Fortney, L.W.; Rakel, D.P. Sustainability, Synthetic Chemicals, and Human Exposure. Explor. J. Sci. Heal. 2010, 6, 186–188. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Kannan, K.; Tan, H.; Zheng, Z.; Feng, Y.L.; Wu, Y.; Widelka, M. Bisphenol Analogues Other Than BPA: Environmental Occurrence, Human Exposure, and Toxicity-A Review. Environ. Sci. Technol. 2016, 50, 5438–5453. [Google Scholar] [CrossRef] [PubMed]
- Schöpel, M.; Shkura, O.; Seidel, J.; Kock, K.; Zhong, X.; Löffek, S.; Helfrich, I.; Bachmann, H.S.; Scherkenbeck, J.; Herrmann, C.; et al. Allosteric Activation of GDP-Bound Ras Isoforms by Bisphenol Derivative Plasticisers. Int. J. Mol. Sci. 2018, 19, 1133. [Google Scholar] [CrossRef] [PubMed]
- Schöpel, M.; Jockers, K.F.G.; Düppe, P.M.; Autzen, J.; Potheraveedu, V.N.; Ince, S.; Yip, K.T.; Heumann, R.; Herrmann, C.; Scherkenbeck, J.; et al. Bisphenol A binds to Ras proteins and competes with guanine nucleotide exchange: Implications for GTPase-selective antagonists. J. Med. Chem. 2013, 56, 9664–9672. [Google Scholar] [CrossRef]
- National Institute of Environmental Health Sciences. Endocrine Disruptors; NIEHS: Durham, NC, USA.
- Gore, A.C.; Chappell, V.A.; Fenton, S.E.; Flaws, J.A.; Nadal, A.; Prins, G.S.; Toppari, J.; Zoeller, R.T. EDC-2: The Endocrine Society’s Second Scientific Statement on Endocrine-Disrupting Chemicals. Endocr. Rev. 2015, 36, E1–E150. [Google Scholar] [CrossRef] [PubMed]
- Rubin, B.S. Bisphenol A: An endocrine disruptor with widespread exposure and multiple effects. J. Steroid Biochem. Mol. Biol. 2011, 127, 27–34. [Google Scholar] [CrossRef]
- Routledge, E.J.; White, R.; Parker, M.G.; Sumpter, J.P. Differential effects of xenoestrogens on coactivator recruitment by estrogen receptor (ER) α and ERβ. J. Biol. Chem. 2000, 275, 35986–35993. [Google Scholar] [CrossRef]
- Welshons, W.V.; Nagel, S.C.; Vom Saal, F.S. Large effects from small exposures. III. Endocrine mechanisms mediating effects of bisphenol A at levels of human exposure. Endocrinology 2006. [Google Scholar] [CrossRef]
- Wetherill, Y.B.; Akingbemi, B.T.; Kanno, J.; McLachlan, J.A.; Nadal, A.; Sonnenschein, C.; Watson, C.S.; Zoeller, R.T.; Belcher, S.M. In Vitro molecular mechanisms of bisphenol A action. Reprod. Toxicol. 2007, 24, 178–198. [Google Scholar] [CrossRef]
- Shanle, E.; Xu, W. Endocrine disrupting chemicals targeting estrogen receptor signaling: Identification and mechanisms of action. Chem. Res. Toxicol. 2010, 24, 6–19. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Son, T.G.; Kim, S.J.; Kim, H.S.; Kim, T.S.; Han, S.Y.; Lee, J. Suppressive effects of bisphenol a on the proliferation of neural progenitor cells. J. Toxicol. Environ. Heal. Part A 2007, 70, 1288–1295. [Google Scholar] [CrossRef] [PubMed]
- Dolinoy, D.C.; Huang, D.; Jirtle, R.L. Maternal nutrient supplementation counteracts bisphenol A-induced DNA hypomethylation in early development. Proc. Natl. Acad. Sci. USA 2007, 104, 13056–13061. [Google Scholar] [CrossRef] [PubMed]
- Soto, A.M.; Sonnenschein, C. Environmental causes of cancer: Endocrine disruptors as carcinogens. Nat. Rev. Endocrinol. 2010, 6, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Norgard, R.J.; Stanger, B.Z. Cellular Plasticity in Cancer. Cancer Discov. 2019. [Google Scholar] [CrossRef] [PubMed]
- LaPensee, E.W.; Tuttle, T.R.; Fox, S.R.; Ben-Jonathan, N. Bisphenol A at low nanomolar doses confers chemoresistance in estrogen receptor-α-positive and -negative breast cancer cells. Environ. Health Perspect. 2009, 117, 175–180. [Google Scholar] [CrossRef] [PubMed]
- LaPensee, E.; LaPensee, C.; Fox, S.; Schwemberger, S.; Afton, S.; Ben-Jonathan, N. Bisphenol A and estradiol are equipotent in antagonizing cisplatin-induced cytotoxicity in breast cancer cells. Cancer Lett. 2010, 290, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Delgado, M.; Ribeiro-Varandas, E. Bisphenol A at the reference level counteracts doxorubicin transcriptional effects on cancer related genes in HT29 cells. Toxicol. Vitr. 2015, 29, 2009–2014. [Google Scholar] [CrossRef]
- Swift, L.P.; Rephaeli, A.; Nudelman, A.; Phillips, D.R.; Cutts, S.M. Doxorubicin-DNA adducts induce a non-topoisomerase II-mediated form of cell death. Cancer Res. 2006, 66, 4863–4871. [Google Scholar] [CrossRef]
- National Cancer Institute. Doxorubicin; National Cancer Institute: Bethesda, MD, USA, 2007.
- Rudel, R.A.; Gray, J.M.; Engel, C.L.; Rawsthorne, T.W.; Dodson, R.E.; Ackerman, J.M.; Rizzo, J.; Nudelman, J.L.; Brody, J.G. Food packaging and bisphenol A and bis(2-ethyhexyl) phthalate exposure: Findings from a dietary intervention. Environ. Health Perspect. 2011, 119, 914–920. [Google Scholar] [CrossRef]
- Ribeiro, E.; Ladeira, C.; Viegas, S. Occupational exposure to Bisphenol A (BPA): A reality that still needs to be unveiled. Toxics 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, E.; Ladeira, C.; Viegas, S. EDCs mixtures: A stealthy hazard for human health? Toxics 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Ramos, C.; Ladeira, C.; Dias, A.; Faria, I.; Cristovam, E. Cytotoxic and genotoxic effects of environmental relevant concentrations of bisphenol A and interactions with doxorubicin. Mutat. Res. Gen. Tox. Environ. Mutagen. 2019, 838, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Warfel, N.; El-Deiry, W. p21WAF1 and tumourigenesis: 20 years after. Curr. Opin. Oncol. 2013, 25, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Durchdewald, M.; Angel, P.; Hess, J. The transcription factor Fos: A Janus-type regulator in health and disease. Histol. Histopathol. 2009, 24, 1451–1461. [Google Scholar] [CrossRef] [PubMed]
- Bisphenol, A. Integrated Risk Information System; USEPA (U.S. Environmental Protection Agency): Washington, DC, USA, 1988.
- European Food Safety Authority. Bisphenol, A; European Food Safety Authority: Parma, Italy, 2017. [Google Scholar]
- Greene, R.F.; Collins, J.M.; Jenkins, J.F.; Speyer, J.L.; Myers, C.E. Plasma pharmacokinetics of adriamycin and adriamycinol: Implications for the design of in vitro experiments and treatment protocols. Cancer Res. 1983, 43, 3417–3421. [Google Scholar]
- Eom, Y.; Kim, M.A.; Park, S.S.; Goo, M.J.; Kwon, H.J.; Sohn, S.; Kim, W.-H.; Yoon, G.; Choi, K.S. Two distinct modes of cell death induced by doxorubicin: Apoptosis and cell death through mitotic catastrophe accompanied by senescence-like phenotype. Oncogene 2005, 4765–4777. [Google Scholar] [CrossRef]
- Park, S.S.; Kim, M.A.; Eom, Y.; Choi, K.S. Bcl-xL blocks high dose doxorubicin-induced apoptosis but not low dose doxorubicin-induced cell death through mitotic catastrophe. Biochem. Biophys. Res. Commun. 2007, 363, 1044–1049. [Google Scholar] [CrossRef]
- Poehlmann, A.; Reissig, K.; Just, A.; Walluscheck, D.; Hartig, R.; Schinlauer, A.; Lessel, W.; Guenther, T.; Silver, A.; Steinberg, P.; et al. Non-apoptotic function of caspases in a cellular model of hydrogen peroxide-associated colitis. J. Cell. Mol. Med. 2013, 17, 901–913. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Genes | Accession Number * | Forward Primer (5′→3′) | Reverse Primer (3′→5′) | Product Length (bp) |
|---|---|---|---|---|
| GAPDH | NM_002046 | GAGTCAACGGATTTGGTCGTA | GCAGAGATGATGACCCTTTTG | 245 |
| β-actin | NM_001101 | AGGCCAACCGCGAGAAG | ACAGCCTGGATAGCAACGTACA | 79 |
| bcl-xl | Z23115.1 | TTACCTGAATGACCACCTA | ATTTCCGACTGAAGAGTGA | 185 |
| c-fos | NM_005252 | AGGAGAATCCGAAGGGAAAG | CAAGGGAAGCCACAGACATC | 247 |
| p21 | NM_000389 | CTGGAGACTCTCAGGGTCGAA | CCAGGACTGCAGGCTTCCT | 123 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ribeiro, E.; Delgadinho, M.; Brito, M. Environmentally Relevant Concentrations of Bisphenol A Interact with Doxorubicin Transcriptional Effects in Human Cell Lines. Toxics 2019, 7, 43. https://doi.org/10.3390/toxics7030043
Ribeiro E, Delgadinho M, Brito M. Environmentally Relevant Concentrations of Bisphenol A Interact with Doxorubicin Transcriptional Effects in Human Cell Lines. Toxics. 2019; 7(3):43. https://doi.org/10.3390/toxics7030043
Chicago/Turabian StyleRibeiro, Edna, Mariana Delgadinho, and Miguel Brito. 2019. "Environmentally Relevant Concentrations of Bisphenol A Interact with Doxorubicin Transcriptional Effects in Human Cell Lines" Toxics 7, no. 3: 43. https://doi.org/10.3390/toxics7030043
APA StyleRibeiro, E., Delgadinho, M., & Brito, M. (2019). Environmentally Relevant Concentrations of Bisphenol A Interact with Doxorubicin Transcriptional Effects in Human Cell Lines. Toxics, 7(3), 43. https://doi.org/10.3390/toxics7030043