
A Pilot Study on Bioaccumulation and Tissue Distribution of Mercury in Barn Swallow (Hirundo rustica)
1
Life Science and Technology School, Lingnan Normal University, Zhanjiang 524048, China
2
Maoming Branch, Guangdong Laboratory for Lingnan Modern Agriculture, College of Natural Resources and Environment, South China Agricultural University, Guangzhou 510642, China
3
School of Global Public Health, New York University, New York, NY 10044, USA
*
Author to whom correspondence should be addressed.
Toxics 2024, 12(3), 206; https://doi.org/10.3390/toxics12030206
Submission received: 14 December 2023
/
Revised: 1 March 2024
/
Accepted: 6 March 2024
/
Published: 8 March 2024
(This article belongs to the Special Issue Human Exposure to Emerging Contaminants Associated with E-waste Dismantling: Environmental Processes, Human Exposure and Environmental Management)
Abstract
:Although extensive research has been carried out on the occurrence of mercury (Hg) in biota, bioaccumulation and tissue distribution of Hg in songbirds have not been well characterized. In the present study, Hg was investigated in insects and barn swallows (Hirundo rustica) to explore the bioaccumulation characteristics of Hg. Hg in swallow feathers and tissues including muscle, liver, and bone was investigated to determine the tissue distribution of Hg. The concentrations of Hg were 1.39 ± 1.01 μg/g, 0.33 ± 0.09 μg/g, 0.47 ± 0.10 μg/g, and 0.23 ± 0.09 μg/g in feather, muscle, liver, and bone samples, respectively. The trophic magnification factor of Hg in swallows and insects was higher than 1. However, the Hg concentrations in swallow feathers were not significantly correlated with stable isotope values of carbon or nitrogen, which implies the complex food sources and exposure processes of Hg for swallows. Feathers had significantly higher concentrations of Hg than liver, muscle, and bone samples (p < 0.01 for all comparisons). Feather, muscle, bone, and other organs had fractions of 64.4 ± 11.9%, 6.07 ± 2.06%, 20.0 ± 8.19%, and 9.56 ± 2.96% in total body burden of Hg in swallows. Hg in feathers contributed more than half of Hg in the whole body for most swallow individuals. Swallows may efficiently eliminate Hg by molting, and the excretion flux of Hg and other contaminants via molting deserves more investigation.

1. Introduction
Mercury (Hg) is a widespread and persistent environmental contaminant, which poses significant risks to wildlife and also humans. Mercury acts as a neurotoxin and potentially brings significant risks to the health of wildlife at sufficient concentrations [1,2]. Hg exposure will exert various adverse effects, including low reproduction output, suppressed immune function, altered breeding activity, and reduced cognitive performance [1,2,3,4]. In the aquatic anaerobic environment, inorganic Hg can be converted to methyl mercury (MeHg) under microbial processes. MeHg is more bioavailable and bioaccumulative than inorganic Hg in organisms and is prone to transfer through food chains into organisms at high trophic levels [5,6].
Birds have long been recognized as biosentinels of environmental pollutants, especially persistent, bioaccumulative, and toxic chemicals like Hg [6]. Seabirds and raptors have been frequently investigated for Hg pollution because they occupy high trophic levels in local ecosystems and can accumulate considerable concentrations of Hg [7,8]. Songbirds are less studied for Hg but are recently becoming popular environmental indicators of Hg exposure and risk [9,10,11]. Songbirds are abundant and widely distributed in different regions. They also feed on a range of dietary items increasing their exposure to anthropogenic pollution [6]. Though songbirds inhabit terrestrial environments, it has been proved that Hg in wetlands can accumulate in aquatic invertebrates and be transferred to terrestrial predators (including songbirds) via the consumption of aquatic emergent insects [12,13,14]. Songbirds usually have a wide range of movements in various landscapes and can also ingest nutrients and contaminants from terrestrial and aquatic prey. Although it is well known that Hg biomagnifies in food chains, diverse food source complicates the biomagnification factors of Hg [12,15]. Dietary sources and habitat-specific foraging led to highly variable concentrations of MeHg, and the ranges of MeHg levels differed up to 283-fold in a songbird species from the same sampling site [15]. Insectivores with higher fractions of aquatic prey sources may have higher MeHg concentrations than those with less aquatic prey sources and similar trophic positions. Hence, it is critical to trace various food sources and trophic positions when evaluating Hg concentrations in riparian wildlife.
Internal tissues of birds, such as blood, eggs, muscle, and liver, are generally considered the most reliable indicators of Hg exposure in birds [1]. However, analysis of the internal tissues requires lethal methods, which is always contradictory to ethical regulations and conservation goals. Feathers have been considered useful non-lethal alternatives for monitoring environmental pollutants [16,17,18]. In addition, feathers are more chemically and physically stable. Thus, feathers are suitable biomonitoring tools that are non-invasive and easy to preserve for a long time. Although feathers have been commonly used for monitoring Hg in avian populations, the reliability of feathers as a sampling matrix has not been thoroughly assessed for many songbird species [10]. In some cases, such as nonmigratory species, body feathers show a strong correlation with internal tissues. A previous study in songbirds reported that feather growth reduced the concentrations of Hg in blood and Hg levels in blood rose after the completion of feather growth [17]. Moreover, a significant relationship has been observed between blood and feather samples in the songbird species, Carolina Wrens [9]. Although molt was probably the main mercury excretion pathway in birds to accelerate the rate of depuration of Hg from blood or other internal tissues, research suggested that Hg depuration from organs in some songbirds was not consistently affected by the rate of molt [2,19]. For example, in the contrast analysis of accelerated molt and gradual molt finches, the difference in Hg levels in internal tissues (including brain, muscle, liver, and kidney) was not significant [2]. These studies reported inconsistent relationships between tissue and feather Hg concentrations [9,20,21], indicating that feathers do not accurately represent recent Hg exposure in some bird species. In addition, the tissue distribution of Hg in feathers and different internal tissues including muscle, liver, and bone was scarcely reported in the literature [22] and still needs to be testified for songbird species.
In order to better understand the food chain transfer and tissue distribution of Hg in swallows, the present study aimed to (1) examine the concentrations of Hg in different tissues of swallows, including muscle, liver, bone, and feather; (2) explore the potential trophic transfer of Hg; (3) elucidate the tissue distribution of Hg in swallows, which can indicate if feather can serve as an indicator of Hg pollution for different internal tissues.
2. Materials and Methods
2.1. Sample Collection
Body feather samples were collected from dead barn swallows (Hirundo rustica, n = 22, including 4 adults and 18 juveniles) discovered in a village close to Zhanjiang Bay, South China (20°53′ E, 110°10′ N), in 2021. Mineral exploitation, shipyard activities, and agricultural activities caused pollution in that region. The occurrence of Hg has been reported in previous studies [23,24]. The study area is a typical agricultural area of the subtropical plain in South China dominated by vegetable fields, sugar cane fields, and paddy fields including C4 and C3 plants. The main crop is rice, which can be harvested two or three times every year. The livestock reared in villages includes buffalos and chickens.
Most dead birds (n = 15) were completely dry when found on the road or in grassland, so only feather samples were collected for analysis. The birds were local breeding populations and were sampled from April to August, which was the breeding season for the swallows in the studied site. The swallows were found dead on the road or in grassland, but we were not sure if the swallows were roadkill or if they were dead because of the harsh summer conditions [25]. The bodies of seven birds were fresh and dissected for analysis of Hg in tissues. The muscle, liver, and sternum samples were acquired from seven swallows with fresh body conditions. Body feathers were rinsed with ultrapure water and freeze-dried. All samples were wrapped in aluminum foil and kept in freezers at −20 °C before further analysis. The tissues of individual swallows were analyzed. Insect samples were collected by sweep nets during the day and light traps at night. Locust and dragonfly samples were collected during the day, while cricket and beetle samples were collected during both day and night time. Because of the small body size of insects, insect individuals belonging to cricket, locust, dragonfly, and beetle were combined as composites to fulfill the analysis mass of stable isotopes and Hg concentrations. A total of 44 crickets, 48 locusts, 27 dragonflies, and 30 beetles were captured. In order to determine the detection limit of Hg and stable isotope, the insect individuals were mixed as four cricket, four locust, three dragonfly, and five beetle composite samples. The subsamples from each composite sample were used for Hg.
2.2. Sample Extraction and Analysis
All samples were first freeze-dried, and approximately 0.05 g of the dry sample was accurately weighed. The dry samples were ground to powder finer than 0.5 mm. Then, the samples were dipped in 4 mL HNO3 (65%) overnight. Then, samples were added with 3 mL H2O2 (30%) and digested at 120 °C for 4 h. The digestive fluid was cooled to ambient temperature, transferred to a volumetric flask, and diluted to 10 mL for instrumental analysis. Hg was quantified by atomic fluorescence spectrometry (AFS-8510, Beijing Haiguang, Beijing, China). All data in this study were expressed in μg/g dry weight (dw).
2.3. Isotope Analysis and Calculations
The measurements of δ13C and δ15N were the same as in previous studies [12]. Approximately 0.5 mg of dry sample was placed in a tin capsule and analyzed by a Flash EA 112 series elemental analyzer coupled with a Finnigan MAT ConFlo III isotope ratio mass spectrometer. Stable isotope abundance was calculated as follows:
where X represents 13C or 15N, and Rsample/Rstandard is the 13C/12C or 15N/14N ratio. The precisions were ±0.2‰ (2 SD) for δ13C and ±0.5‰ (2 SD) for δ15N.
δX = (Rsample/Rstandard − 1) × 1000
2.4. Trophic Magnification Factor (TMF)
The trophic level (TL) was determined by the following equation:
where δ15Nsample is the δ15N of a swallow or an insect sample, and δ15Nprimary consumer is the δ15N value of the beetle in the present study. The trophic discrimination factor (TDF) is set as 3.4.
TL = (δ15Nsample − δ 15Nprimary consumer)/TDF + 2
TMF was calculated as follows:
where CHg is the Hg concentration of the barn swallow feather or the whole insect, a is the y-intercept (constant), and b is the slope. Statistical significance of Equation (3) is defined at p < 0.05.
lg CHg= a + b × TL
TMF = 10b
2.5. QA/QC and Statistical Analysis
Quality control was carried out by analyzing blanks and standard reference materials. Two blank samples were included with each batch, and the concentrations of chemicals in blank samples were deducted from those in experimental samples. Recoveries of Hg in standard reference materials (GBW10018 (GSB-9) standard substance for chicken [26]) ranged from 85% to 110% with relative standard deviations (SDs) less than 15%. The limit of quantification (LOQ) of Hg was three times the standard deviation of Hg detected in blanks and was 0.001 μg/g dry weight (dw) in the present study.
All statistical analyses were performed by SPSS 22.0 (SPSS Inc., Chicago, IL, USA). Concentrations of Hg were log-transformed to obtain normal distribution. T-test was employed to test for the significance of differences between Hg levels in different groups of samples. Pearson correlation analysis was used to test the correlations between Hg levels in different groups of samples. Significance was set as p < 0.05.
3. Results
3.1. Hg in Swallow Feather
Hg was detected in all swallow samples with concentrations of 1.39 ± 1.01 μg/g in feathers. Hg concentrations in swallow feathers were lower than Hg in feathers of seabirds, such as penguins (1.88 μg/g, [27]) and petrels (19.7 μg/g, [28]). The results of swallows were also lower than the concentrations of Hg (1.95 ± 1.37 μg/g) in feathers of raptors from the Marin Headlands of California, USA [7]. It is reasonable to observe lower concentrations of Hg in insectivorous birds than in seabirds and raptors because of the high trophic positions of seabirds and raptors. Songbirds (such as Yuhina gularis, Pycnonotus xanthorrhous, and Suthora nipalensis) from Southwestern China [11] had comparable concentrations of Hg (1.66 ± 1.03 μg/g) with swallows in this study. In the present study, Hg concentrations in swallow individuals were highly variable, ranging from 0.30 to 4.34 μg/g. Though the swallows stayed in the breeding seasons from April to August in the studied site, it is possible that some swallows have different wintering sites. Thus, the exposure residue during the wintering season can bring uncertainty in Hg bioaccumulation. The swallows have different foraging strategies and therefore different exposure values of Hg. As there were only 22 samples, we did not explore the temporal trend in the five months by statistical analysis method. The concentrations of Hg in feathers from fresh (n = 7) and dry (n = 15) swallow bodies were compared by t-test, and no significant differences were observed in Hg concentrations in the two groups (p = 0.21). The feather samples are more stable than other bird tissues under ambient conditions. The results indicate that the body condition of swallows does not have a significant effect on Hg concentrations in feather samples. The age of swallows also makes it difficult to predict the contamination values. In this study, the feather samples were from juvenile and adult swallows. However, it is difficult to confirm their specific age and molting time. The effect of age on Hg concentrations in feathers requires further study in the future. And more notably, Ackerman et al. [1] suggested that if Hg concentrations in bird blood were greater than 0.2 μg/g, it can be considered to have risk. According to the following formula [1],
where CBlood Hg and CBird Feather Hg are concentrations of Hg in bird blood (μg/g wet weight) and bird feather (μg/g dry weight), respectively.
ln CBlood Hg= 0.673 × ln CBird Feather Hg − 1:673
The threshold of 0.2 μg/g Hg in blood was converted to 1.09 μg/g Hg concentrations in bird feathers. It is worth noting that eleven out of twenty-two swallows had concentrations of Hg in feathers higher than the threshold of 1.09 μg/g Hg in feathers, which could pose risk deleterious effects to birds [1]. Most studies focused on the exposure of Hg in aquatic and riparian food webs with high Hg basal and have noted the high exposure risk of Hg in these organisms. The result of the present study suggests that insectivorous birds may also suffer from elevated Hg exposure risks, even though insectivorous birds like swallows inhabit low basal Hg input terrestrial ecosystems. Terrestrial organisms like insectivorous birds may take up a certain amount of Hg via aquatic prey.
3.2. Influences on Hg Pollution
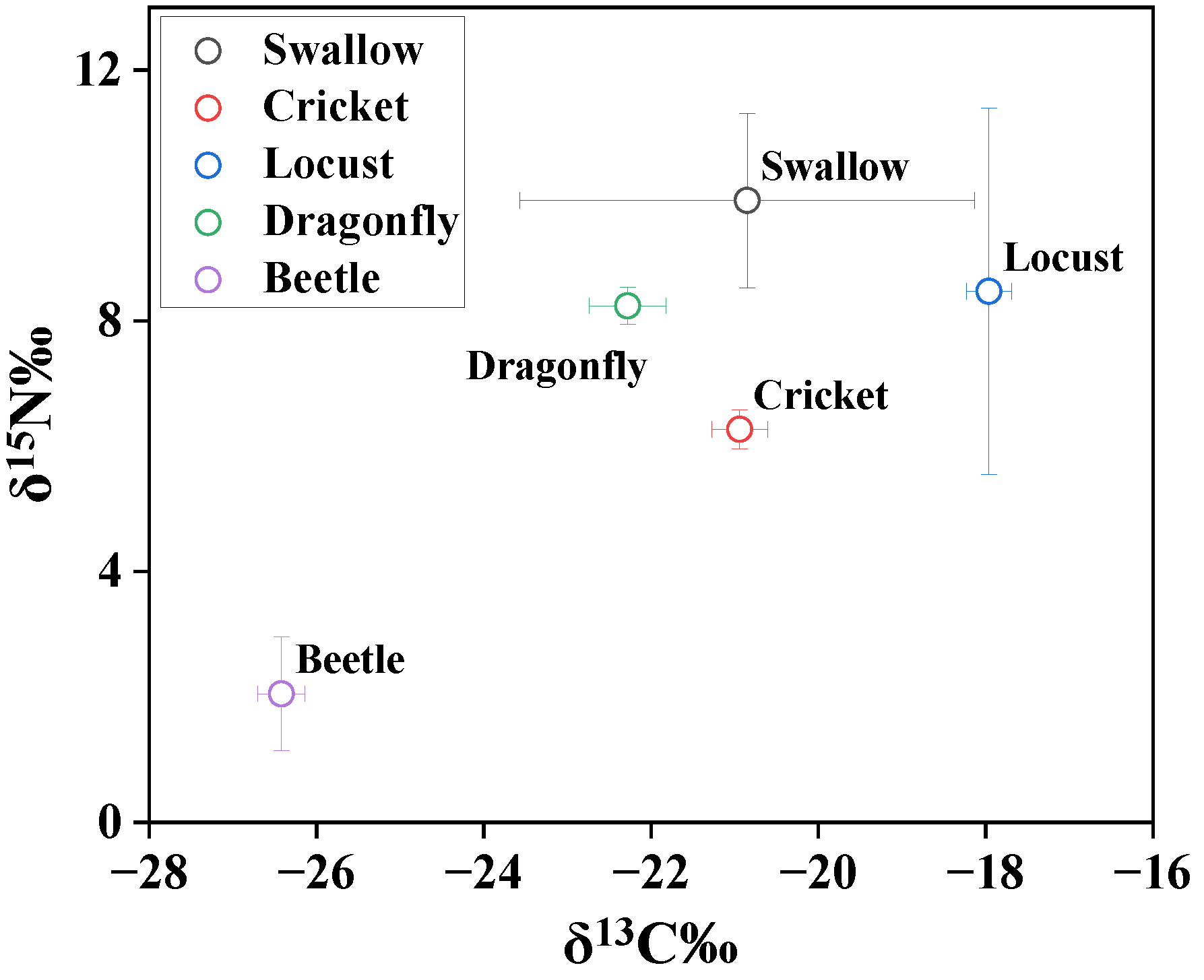
In order to explore the food chain transfer of Hg from insects to swallows, Hg and stable isotopes of carbon and nitrogen were measured in insect species and swallow feathers (Table 1 and Figure 1). The stable carbon and nitrogen isotope signature indicates the diet source and trophic position, respectively, because δ13C increases by 0–1‰ and δ15N increases by 3–5‰ per trophic level [29,30]. The δ13C and δ15N values were −20.8 ± 2.72‰ and 9.92 ± 1.39‰ in swallow feathers, respectively (Figure 1). The δ13C values ranged from −18.0 ± 0.27‰ to −26.4 ± 0.28‰, and the δ15N values ranged from 2.05 ± 0.91‰ to 8.47 ± 2.92‰ in the insect species. The insect species had a broad δ13C range, indicating different carbon sources for insect species. δ13C in C3 plants ranged from 30.6‰ to 22.2‰, while C4 plants usually have δ13C values ranging from 20‰ to 10‰ [31]. The locust and beetle species seem to feed on C4 and C3 plants, respectively, while dragonflies and crickets are more likely to feed on mixed sources of C3 and C4. It was difficult to trace the feed sources of swallows by stable isotopes of carbon and nitrogen, as swallows showed δ13C values in the range of those in insects and higher δ15N values than those in insects. The investigated insects were all potential feeding sources for swallows.
The mean concentrations of Hg were 0.03, 0.04, 0.04, and 0.15 μg/g in locusts, crickets, beetles, and dragonflies, respectively. Dragonflies have significantly higher Hg concentrations than other insect species (p < 0.05), which can be attributed to the aquatic habitat of dragonflies. Aquatic insects always exhibit greater exposure to Hg compared with terrestrial insects because of the microbial production of bioaccumulative MeHg in sediment [13]. Though the diet sources of swallows cannot be accurately estimated in this study, the insects and swallows were collected from the same village, which enabled discussions on the trophic transfer of Hg. Beetles had the lowest δ15N value (2.5 ± 0.91‰); therefore, it was deemed as the lowest TL (2.0 ± 0.27) herein. Among other species, crickets had the lowest TLs (3.24 ± 0.11), followed by dragonflies (3.82 ± 0.089), locusts (3.89 ± 0.086), and barn swallows (4.31 ± 0.41). Based on TL and δ15N values, the food web of insects–swallows was established in the present study. A significant positive correlation was found between trophic levels and log-transformed concentrations of Hg (p = 0.0013), as shown in Figure 2, which suggests a distinct transfer of Hg from lower to higher trophic levels.
The TMF of Hg in the present food web was calculated. The TMF (3.16) was greater than 1, indicating significant trophic magnification of Hg in the present food web. The TMF herein was slightly higher than the TMF of MeHg in a riparian food web (2.39), including insectivorous birds from another site in South China [12], and was similar to TMFs of Hg (2.64–3.02) in two marine food webs including seabirds from North China and Norway [8,32]. Cao et al. [33] reported the TMF value (1.69) of Hg from phytoplankton–invertebrate–fish food webs in Laizhou Bay, which was lower than the TMF of Hg in the present study, whereas a higher TMF value (5.7) of Hg from the primary producer–invertebrate–fish food webs in Poyang Lake was reported by Zhang et al. [34]. Variations in TMF values among different ecosystems might be due to factors such as food web structure, taxa groups, energy requirements of organisms, and temporal and spatial changes in sources of Hg for organisms [35]. Furthermore, due to the differences in the binding ability of Hg among tissues, different tissue types of organisms used in the analysis might also affect the TMF estimation. Previous research has reported that available binding sites in tissues for MeHg decreased with an increase in lipid contents [36]. Therefore, tissues containing a higher lipid content might be expected to have lower Hg concentrations [36]. However, only the feathers of swallows in the present study were used to estimate the TMF because of the limited swallow tissue samples. Further work is needed to evaluate the impact of the different tissue types on the TMF estimation in the future.
Correlations between stable isotope values and concentrations of Hg were tested for swallows, and no obvious relationship was observed between concentrations of Hg and δ15N or δ13C (Figure 3). It is demonstrated that Hg in bird feathers is mainly composed of MeHg, which is derived from trophic transfer in food chains [37]. However, concentrations of Hg in swallow feathers were not correlated with δ15N values in the present study, which indicates no obvious trophic transfer of Hg. The result could be explained by the different baseline values of δ15N. The diet sources are complex for the studied swallows based on the δ13C results. The different baseline values of δ15N in different insects and swallows affected the correlations between Hg concentrations and δ15N values. The basis of δ15N can also be affected by landscape types and agricultural activities in the studied region [38,39]. Wild creatures may receive organic and inorganic agricultural inputs of nitrogen from a broad area.
The high burden of Hg in certain terrestrial predators such as spiders [40], songbirds [11], and bats [41] has been frequently reported, which is mainly attributed to the contribution of aquatic food sources containing Hg. In adjacent food webs from South China, concentrations of Hg were significantly and positively correlated with δ13C rather than δ15N in terrestrial insects and birds [12]. Aquatic insects have been identified as a potential route for dietary metal exposure for terrestrial insectivores [11,12]. Higher values of δ13C indicate more proportions of aquatic prey in the total diet for songbirds, leading to high exposure levels of Hg. In contrast, no obvious relationship has been observed between the concentrations of Hg and δ13C values in swallows in the present study. Aside from the complex carbon sources for swallows, the concentrations and bioavailability of Hg are also suggested as important factors in the bioaccumulation of Hg. For instance, beetles have indigestible shells for birds and may have different bioavailability of Hg compared to other insects. In addition, the present study only collected three orders of insects, whereas swallows are known to forage on various species of insects. More ecology and contamination information on more insect species can help to elucidate the pollution risks for swallows.
3.3. Tissue Distribution of Hg in Swallows
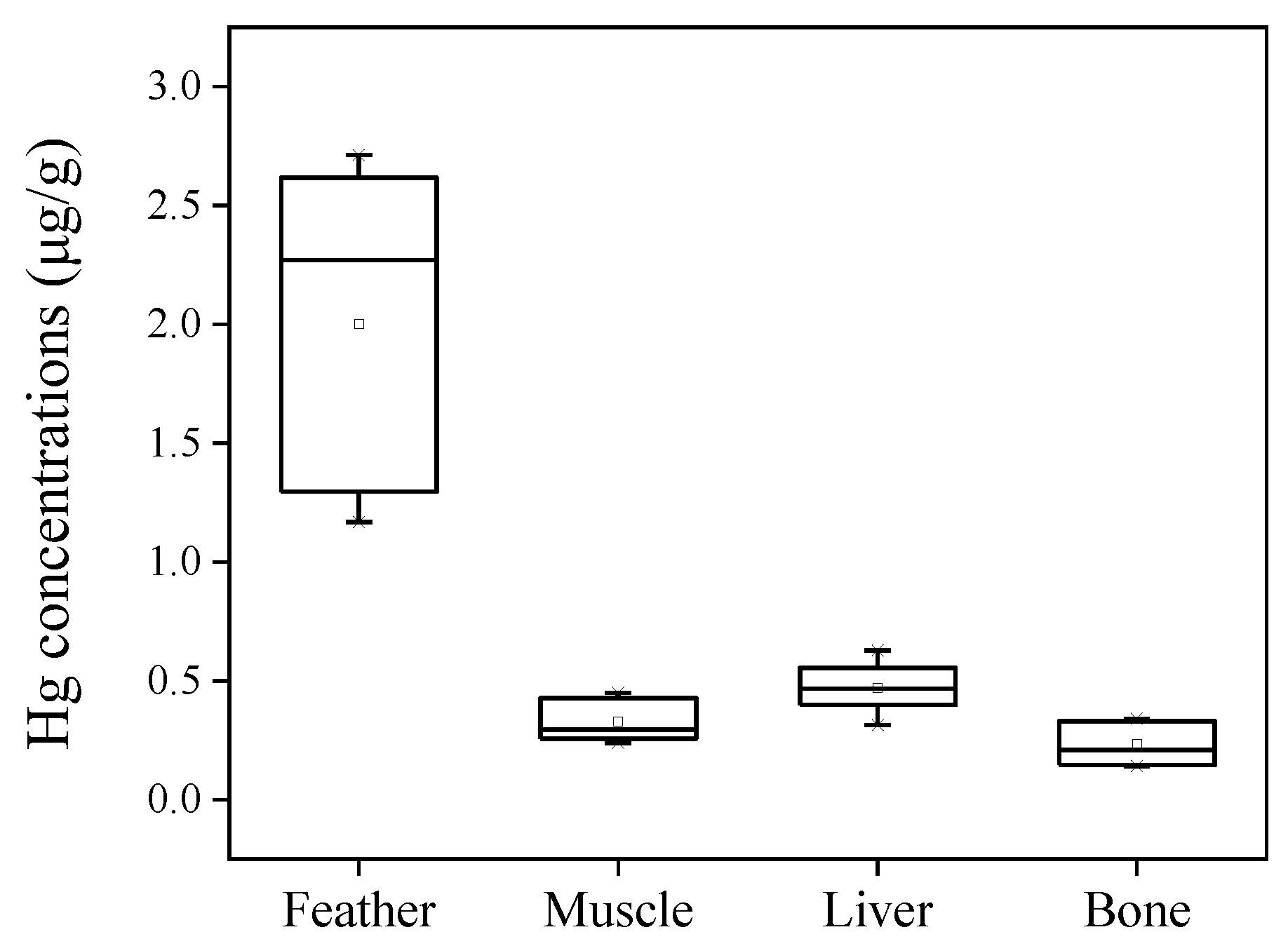
Most studies have monitored pollutants in the feathers and internal tissues (including muscle, liver, and blood) of birds [6]. Feather was proposed as a non-invasive monitoring sample for birds. Some studies have tried to assess the relationships between Hg concentrations in feathers and internal doses in blood [9,16,17]. In the present study, the muscle, liver, and bone samples were available for seven swallows. The tissue distribution of Hg was then discussed. Hg was detected in all swallow tissues. The concentrations of Hg were 0.33 ± 0.09 μg/g, 0.47 ±0.10 μg/g, and 0.23 ± 0.09 μg/g in the muscle, liver, and bone samples, respectively (Figure 4). Feathers had significantly higher concentrations of Hg than internal tissues including muscle, liver, and bone (p < 0.01), whereas bones had the lowest Hg concentrations among all tissues (p < 0.05). Correlations between the concentrations of Hg in feathers and other tissues were assessed by Pearson correlation analysis, and no significant correlations were observed between feathers and muscles, livers, or bones (p > 0.05). Previous studies have reported that Hg concentrations were the highest in feathers and nails, followed by the liver and then the muscle, among tissues [21,42,43]. For instance, mean Hg concentrations in the thrush family (Turdidae) approximated a 13:3.57:1 ratio for feather, liver, and muscle tissues, respectively, as reported by Low et al. [44]. This trend was consistent with the findings in the present study.
The tissue burden of Hg (g) in feathers, muscles, bones, and other organs was roughly estimated as concentrations of Hg multiplied by the weight of different tissues. The liver concentration of Hg was used to calculate the burden of Hg in all viscera organs. Feather, muscle, bone, and other organs had fractions of 64.4 ± 11.9%, 6.07 ± 2.06%, 20.0 ± 8.19%, and 9.56 ± 2.96% in the total body burden of Hg in swallows (Figure 5). Hg in feathers contributed more than half of Hg in all tissues for most swallow individuals, indicating that swallows can efficiently eliminate Hg via molting. Previous studies verified that variations in Hg concentrations existed between inner tissues and feathers, and the order of magnitude of higher Hg concentrations in feathers compared to inner tissues led to an overestimation of Hg burden in birds [44]. In comparison with other tissues, feathers deposited the largest proportion (i.e., the highest concentrations) of Hg originating from diet exposures. The results indicate that feathers may not be a suitable indicator of Hg contamination in internal tissues. However, Hg in feathers is still worth attention because feather molting seems to be an important excretion pathway of Hg for swallowing. As the chick gains mass, the pollution of Hg will dilute in the body burden. Thus, the newly growing feathers allocate Hg, and Hg concentrations in blood and other internal organs will decrease. The excretion flux of Hg and other contaminants via molting warrants more studies in comparison with contaminants in the eggs and feces of birds.
Overall, the present study performed a pilot investigation on the bioaccumulation and tissue distribution of Hg in swallows from a tropical region. There are some limitations in the present study. The swallow populations seem to have complex diet sources, which is indicated by the relatively wide range of δ13C and δ15N values. The manuscript collected limited swallow and insect samples, and the insect composites were analyzed. A more comprehensive survey on swallow diet composition and diet pollution is necessary to elucidate the exposure, bioaccumulation, and risk of Hg and more contaminants in swallows. It should be noted that the survival and reproductive success of swallows in the studied region were relatively lower than in other temperate regions according to our recent observations [25]. The harsh summer conditions in the studied tropical region such as high temperatures, high humidity, rainstorms, and typhoons were suspected to pose negative effects on swallows. However, the toxicological risks of Hg and other contaminants for the local bird populations are not well understood. More research is required to fill the knowledge gap on swallows, which can serve as a useful bioindicator of environmental and ecological conditions.
4. Conclusions
The present study measured Hg in swallow tissues and insect species. However, the Hg concentrations were highly variable in swallow feathers and were not significantly correlated with δ13C or δ15N values, which may be attributed to the pollution levels and bioavailability of Hg in complex diet sources of swallows. Feathers had the highest concentrations of Hg, while bones had the lowest concentrations of Hg among the three tissue types of swallow samples. The estimation of the body burden of Hg shows that feathers are the primary deposit for Hg, with fractions of 64.4 ± 11.9% in the total body burden of Hg in swallows. These results indicate that swallows can efficiently eliminate Hg via molting, which should be of interest in the evaluation of exposure and risk of Hg for swallows and other bird species, and the excretion flux of Hg via molting warrants further study.
Author Contributions
L.T.: Conceptualization, methodology, sampling, data analysis, and original draft preparation; Y.Z. and R.Y.: Sample preparation, instrumental analysis, data analysis, and manuscript review and editing; X.Z.: data analysis, and manuscript review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This study was financially supported by the National Natural Science Foundation of China (No. 42177205), Guangzhou Science and Technology Plan Project (202201010413), and Research Fund of Maoming Branch, Guangdong Laboratory for Lingnan Modern Agriculture (No. 2022ZD001).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ackerman, J.T.; Eagles-Smith, C.A.; Herzog, M.P.; Hartman, C.A.; Peterson, S.H.; Evers, D.C.; Jackson, A.K.; Elliott, J.E.; Vander Pol, S.S.; Bryan, C.E. Avian mercury exposure and toxicological risk across western North America: A synthesis. Sci. Total Environ. 2016, 15, 749–769. [Google Scholar] [CrossRef]
- Whitney, M.; Cristol, D. Rapid depuration of mercury in songbirds accelerated by feather molt. Environ. Toxicol. Chem. 2017, 36, 3120–3126. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Perez, C.R.; Branfireun, B.A.; Guglielmo, C.G. Dietary exposure to methylmercury affects flight endurance in a migratory songbird. Environ. Pollut. 2018, 234, 894–901. [Google Scholar] [CrossRef]
- Seewagen, C.L. The threat of global mercury pollution to bird migration: Potential mechanisms and current evidence. Ecotoxicology 2018, 29, 1254–1267. [Google Scholar] [CrossRef] [PubMed]
- Alberts, J.M.; Sullivan, S.M.; Kautza, A. Riparian swallows as integrators of landscape change in a multiuse river system: Implications for aquatic-to-terrestrial transfers of contaminants. Sci. Total Environ. 2013, 463–464, 42–50. [Google Scholar] [CrossRef]
- Chételat, J.; Ackerman, J.T.; Eagles-Smith, C.A.; Hebert, C.E. Methylmercury exposure in wildlife: A review of the ecological and physiological processes affecting contaminant concentrations and their interpretation. Sci. Total Environ. 2020, 711, 135117. [Google Scholar] [CrossRef] [PubMed]
- Bourbour, R.P.; Martinico, B.L.; Ackerman, J.T.; Herzog, M.P.; Hull, A.C.; Fish, A.M.; Hull, J.M. Feather mercury concentrations in North American raptors sampled at migration monitoring stations. Ecotoxicology 2019, 28, 379–391. [Google Scholar] [CrossRef]
- Jaeger, I.; Hop, H.; Gabrielsen, G.W. Biomagnification of mercury in selected species from an Arctic marine food web in Svalbard. Sci. Total Environ. 2009, 407, 4744–4751. [Google Scholar] [CrossRef]
- Jackson, A.K.; Evers, D.C.; Etterson, M.A.; Condon, A.M.; Folsom, S.B.; Detweiler, J.; Schmerfeld, J.; Cristol, D.A. Mercury exposure affects the reproductive success of a free-living terrestrial songbird, the Carolina Wren (Thryothorus ludovicianus). Auk 2011, 128, 759–769. [Google Scholar] [CrossRef]
- Kardynal, K.J.; Jardine, T.D.; Genier, C.S.V.; Bumelis, K.H.; Mitchell, G.W.; Evans, M.; Hobson, K.A. Mercury exposure to swallows breeding in Canada inferred from feathers grown on breeding and non-breeding grounds. Ecotoxicology 2020, 29, 876–891. [Google Scholar] [CrossRef]
- Li, C.; Xu, Z.; Luo, K.; Chen, Z.; Xu, X.; Xu, C.; Qiu, G. Biomagnification and trophic transfer of total mercury and methylmercury in a sub-tropical montane forest food web, southwest China. Chemosphere 2021, 277, 130371. [Google Scholar] [CrossRef]
- Wu, X.; Chen, L.; Li, X.; Cao, X.; Zheng, X.; Li, R.; Zhang, J.; Luo, X.; Mai, B. Trophic transfer of methylmercury and brominated flame retardants in adjacent riparian and aquatic food webs: 13C indicates biotransport of contaminants through food webs. Environ. Pollut. 2022, 306, 119433. [Google Scholar] [CrossRef] [PubMed]
- Haro, R.J.; Bailey, S.W.; Northwick, R.M.; Rolfhus, K.R.; Sandheinrich, M.B.; Wiener, J.G. Burrowing dragonfly larvae as biosentinels of methylmercury in freshwater food webs. Environ. Sci. Technol. 2013, 47, 8148–8156. [Google Scholar] [CrossRef]
- Twining, C.W.; Razavo, N.R.; Brenna, J.T.; Dzielski, S.A.; Gonzalez, S.T.; Lawrence, P.; Cleckner, L.B.; Flecker, A.S. Emergent freshwater insects serve as subsidies of methylmercury and beneficial fatty acids for riparian predators across an agricultural gradient. Environ. Sci. Technol. 2021, 55, 5868–5877. [Google Scholar] [CrossRef]
- Ackerman, J.T.; Hartman, C.A.; Herzog, M.P. Mercury contamination in resident and migrant songbirds and potential effects on body condition. Environ. Pollut. 2019, 246, 797–810. [Google Scholar] [CrossRef] [PubMed]
- García-Fernández, A.J.; Espín, S.; Martínez-López, E. Feathers as a Biomonitoring Tool of Polyhalogenated Compounds: A Review. Environ. Sci. Technol. 2013, 47, 3028–3043. [Google Scholar] [CrossRef] [PubMed]
- Peterson, S.H.; Ackerman, J.T.; Toney, M.; Herzog, M.P. Mercury Concentrations Vary within and among Individual Bird Feathers: A Critical Evaluation and Guidelines for Feather Use in Mercury Monitoring Programs. Environ. Toxicol. Chem. 2019, 38, 1164–1187. [Google Scholar] [CrossRef]
- Jaspers, V.L.; Covaci, A.; Herzke, D.; Eulaers, I.; Eens, M. Bird feathers as a biomonitor for environmental pollutants: Prospects and pitfalls. Trends Analyt. Chem. 2019, 118, 223–226. [Google Scholar] [CrossRef]
- Costa, R.A.; Eeva, T.; Eira, C.; Vaqueiro, J.; Medina, P.; Vingada, J.V. Great tits breeding performance and mercury contamination from the paper and pulp industry in the west coast of Portugal. Chem. Ecol. 2014, 30, 206–215. [Google Scholar] [CrossRef]
- Mikoni, N.A.; Poppenga, R.; Ackerman, J.T.; Foley, J.; Hazlehurst, J.; Purdin, G.; Aston, L.; Hargrave, S.; Jelks, K.; Tell, L.A. Trace elements in feather and tissue samples from Anna’s hummingbirds. Ecol. Indic. 2017, 80, 96–105. [Google Scholar] [CrossRef]
- Kenow, K.P.; Meyer, M.W.; Hines, R.K.; Karasov, W.H. Distribution and accumulation of mercury in tissues of captive-reared common loon (Gavia immer) chicks. Environ. Toxicol. 2007, 26, 1047–1055. [Google Scholar] [CrossRef] [PubMed]
- Deng, H.; Zhang, Z.; Chang, C.; Wang, Y. Trace metal concentration in Great Tit (Parus major) and Greenfinch (Carduelis sinica) at the Western Mountains of Beijing, China. Environ. Pollut. 2007, 148, 620–626. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Bao, K.; Yan, Y.; Neupane, B.; Gao, C. Spatial distribution of potentially harmful trace elements and ecological risk assessment in Zhanjiang mangrove wetland, South China. Mar. Pollut. Bull. 2022, 182, 114033. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wang, W.X.; Zhang, L. Arsenic speciation and spatial and interspecies differences of metal concentrations in mollusks and crustaceans from a South China estuary. Ecotoxicology 2013, 22, 671–682. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Liu, Y.; Zhou, Z.; Zhou, H.; Lu, S.; Zhang, Z. Reproductive Success of a Tropical Barn Swallow Hirundo rustica Population Is Lower Than That in Temperate Regions. Animals 2023, 13, 62. [Google Scholar] [CrossRef] [PubMed]
- National Sharing Platform for Reference Materials, National Institute of Metrology, China. Available online: https://www.ncrm.org.cn/Web/Home/EnglishIndex (accessed on 5 March 2024).
- Carravieri, A.; Cherel, Y.; Jaeger, A.; Churlud, C.; Bustamante, P. Penguins as bioindicators of mercury contamination in the southern Indian Ocean: Geographical and temporal trends. Environ. Pollut. 2016, 213, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Carravieri, A.; Cherel, Y.; Blevin, P.; Brault-Favrou, M.; Chastel, O.; Bustamante, P. Mercury exposure in a large subantarctic avian community. Environ. Pollut. 2014, 190, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Post, D.M. Using stable isotopes to estimate trophic position: Models, methods, and assumptions. Ecology 2002, 83, 703–718. [Google Scholar] [CrossRef]
- Nielsen, J.M.; Clare, E.L.; Hayden, B.; Brett, M.T.; Kratina, P. Diet tracing in ecology: Method comparison and selection. Methods Ecol. Evol. 2017, 9, 278–291. [Google Scholar] [CrossRef]
- Traugott, M.; Kamenova, S.; Ruess, L.; Seeber, J.; Plantegenest, M. Empirically Characterising Trophic Networks: What Emerging DNA-Based Methods, Stable Isotope and Fatty Acid Analyses Can Offer. Adv. Ecol. Res. 2013, 49, 177–224. [Google Scholar]
- Tong, Y.; Zhang, W.; Hu, X.; Ou, L.; Hu, D.; Yang, T.; Wei, W.; Wang, X. Model description of trophodynamic behavior of methylmercury in a marine aquatic system. Environ. Pollut. 2012, 166, 89–97. [Google Scholar] [CrossRef]
- Cao, L.; Liu, J.; Dou, S.; Huang, W. Biomagnification of methylmercury in a marine food web in Laizhou Bay (North China) and associated potential risks to public health. Mar. Pollut. Bull. 2020, 150, 110762. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, W.; Zheng, N.; Cao, Y.; Xiao, H.; Zhu, R.; Guan, H.; Xiao, H. Methylmercury biomagnification in aquatic food webs of Poyang Lake, China: Insights from amino acid signatures. J. Hazard. Mater. 2021, 404, 123700. [Google Scholar] [CrossRef] [PubMed]
- Lavoie, R.A.; Jardine, T.D.; Chumchal, M.M.; Kidd, K.A.; Campbell, L.M. Biomagnification of mercury in aquatic food webs: A worldwide meta-analysis. Environ. Sci. Technol. 2013, 47, 13385–13394. [Google Scholar] [CrossRef]
- Lavoie, R.A.; Hebert, C.E.; Rail, J.; Braune, B.M.; Yumvihoze, E.; Hill, L.G.; Lean, D.R.S. Trophic structure and mercury distribution in a Gulf of St. Lawrence (Canada) food web using stable isotope analysis. Sci. Total Environ. 2010, 408, 5529–5539. [Google Scholar] [CrossRef] [PubMed]
- Luo, K.; Xu, Z.; Wang, X.; Quan, R.; Lu, Z.; Bi, W.; Zhao, H.; Qiu, G. Terrestrial methylmercury bioaccumulation in a pine forest food chain revealed by live nest videography observations and nitrogen isotopes. Environ. Pollut. 2020, 263, 114530. [Google Scholar] [CrossRef]
- Heaton, T.H. Isotopic studies of nitrogen pollution in the hydrosphere and atmosphere: A review. Chem. Geol. Isot. Geosci. Sect. 1986, 59, 87–102. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Aber, J.D.; Howarth, R.W.; Likens, G.E.; Matson, P.A.; Schindler, D.W.; Schlesinger, W.H.; Tilman, D.G. Human alteration of the global nitrogen cycle: Sources and consequences. Ecol. Appl. 1997, 7, 737–750. [Google Scholar] [CrossRef]
- Speir, S.L.; Chumchal, M.M.; Drenner, R.W.; Cocke, W.G.; Lewis, M.E.; Whitt, H.J. Methyl mercury and stable isotopes of nitrogen reveal that a terrestrial spider has a diet of emergent aquatic insects. Environ. Toxicol. Chem. 2014, 33, 2506–2509. [Google Scholar] [CrossRef]
- Becker, D.J.; Chumchal, M.M.; Broders, H.G.; Korstian, J.M.; Clare, E.L.; Rainwater, T.R.; Platt, S.G.; Simmons, N.B.; Fenton, M.B. Mercury bioaccumulation in bats reflects dietary connectivity to aquatic food webs. Environ. Pollut. 2018, 233, 1076–1085. [Google Scholar] [CrossRef]
- Caldwell, C.A.; Arnold, M.A.; Gould, W.R. Mercury distribution in blood, tissues, and feathers of double-crested cormorant nestlings from arid-lands reservoirs in south central New Mexico. Arch. Environ. Contam. Toxicol. 1999, 36, 456–461. [Google Scholar] [CrossRef]
- Szumilo, E.; Szubska, M.; Meissner, W.; Beldowska, M.; Falkowska, L. Mercury in immature and adults Herring Gulls (Larus argentatus) wintering on the Gulf of Gdansk area. Oceanol. Hydrobiol. Stud. 2013, 42, 260–267. [Google Scholar] [CrossRef]
- Low, K.E.; Ramsden, D.K.; Jackson, A.K.; Emery, C.; Robinson, W.D.; Randolph, J.; Eagles-Smith, C.A. Songbird feathers as indicators of mercury exposure: High variability and low predictive power suggest limitations. Ecotoxicology 2020, 29, 1281–1292. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Stable isotope values in organisms.

Figure 2.
Relationships between log-transformed concentrations of Hg and trophic levels in samples. The black line and grey area mean the regression line of samples and the 95% confidence interval of regression line, respectively.
Figure 2.
Relationships between log-transformed concentrations of Hg and trophic levels in samples. The black line and grey area mean the regression line of samples and the 95% confidence interval of regression line, respectively.

Figure 3.
Relationships between Hg concentrations and stable isotope values in swallow feather samples. The black line and grey area mean the regression line of samples and the 95% confidence interval of regression line, respectively.
Figure 3.
Relationships between Hg concentrations and stable isotope values in swallow feather samples. The black line and grey area mean the regression line of samples and the 95% confidence interval of regression line, respectively.

Figure 4.
The ranges of Hg concentrations in paired swallow tissues. The boxes represent the range from the lower quartile to the upper quartile. The horizontal lines and the tiny squares in the boxes represent the median and average values, respectively.
Figure 4.
The ranges of Hg concentrations in paired swallow tissues. The boxes represent the range from the lower quartile to the upper quartile. The horizontal lines and the tiny squares in the boxes represent the median and average values, respectively.

Figure 5.
Proportions of Hg burden in swallow tissues. Each bar represents a swallow individual.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
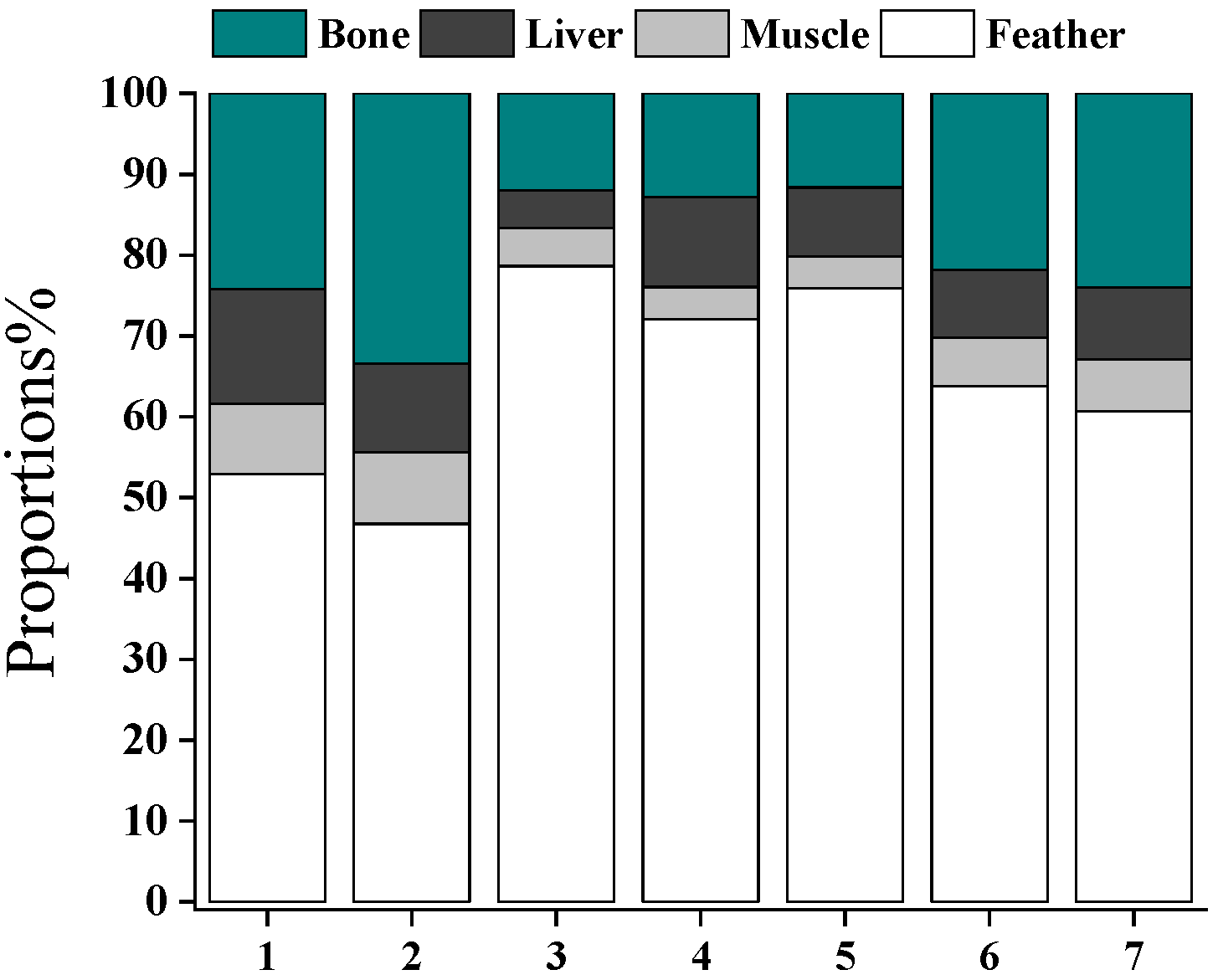{kind=link}
Table 1.
Sample information and Hg concentrations (μg/g dw).
| English Name | N | δ13C‰ | δ15N‰ | Hg |
|---|---|---|---|---|
| Barn Swallow (Hirundo rustica) | ||||
| Feather | 22 | −20.85 ± 2.72 | 9.92 ± 1.39 | 1.39 ± 1.01 |
| Muscle | 7 | 0.33 ± 0.09 | ||
| Liver | 7 | 0.47 ± 0.10 | ||
| Bone | 7 | 0.23 ± 0.09 | ||
| Insects | ||||
| Cricket | 4 | −20.94 ± 0.33 | 6.27 ± 0.31 | 0.04 ± 0.05 |
| Locust | 4 | −17.96 ± 0.27 | 8.47 ± 2.92 | 0.03 ± 0.04 |
| Dragonfly | 3 | −22.28 ± 0.46 | 8.24 ± 0.3 | 0.15 ± 0.00 |
| Beetle | 5 | −26.42 ± 0.28 | 2.05 ± 0.91 | 0.04 ± 0.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tian, L.; Zhu, Y.; Yu, R.; Zheng, X. A Pilot Study on Bioaccumulation and Tissue Distribution of Mercury in Barn Swallow (Hirundo rustica). Toxics 2024, 12, 206. https://doi.org/10.3390/toxics12030206
AMA Style
Tian L, Zhu Y, Yu R, Zheng X. A Pilot Study on Bioaccumulation and Tissue Distribution of Mercury in Barn Swallow (Hirundo rustica). Toxics. 2024; 12(3):206. https://doi.org/10.3390/toxics12030206
Chicago/Turabian StyleTian, Li, Yujing Zhu, Ruiming Yu, and Xiaobo Zheng. 2024. "A Pilot Study on Bioaccumulation and Tissue Distribution of Mercury in Barn Swallow (Hirundo rustica)" Toxics 12, no. 3: 206. https://doi.org/10.3390/toxics12030206
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.