Microbiological Profile and Bioactive Properties of Insect Powders Used in Food and Feed Formulations

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Raw Materials and Microbiological Analyses
2.2. V3-V4 Amplification and Illumina Data Analysis
2.3. Phenotypic and Genotypic Characterization of LAB
2.4. Safety Aspects of Dominant Insect Powder LAB
2.5. Proximate Composition
2.6. Enzymatic hydrolysis
2.7. Sodium Dodecyl Sulphate–Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.8. DPPH Radical Scavenging Activity
2.9. Statistical Analyses
3. Results and Discussion
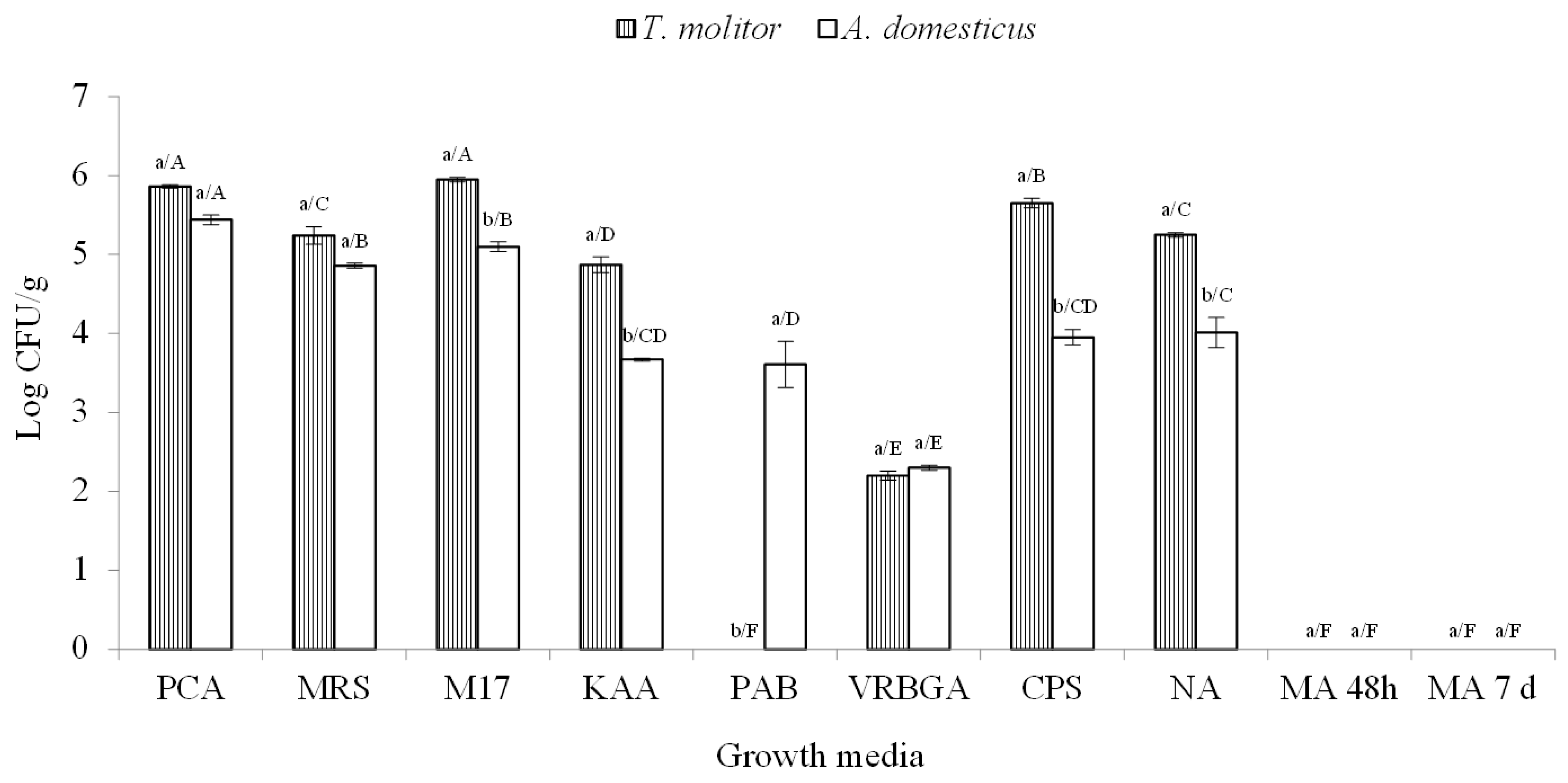
3.1. Microbial Loads of Insect Flours
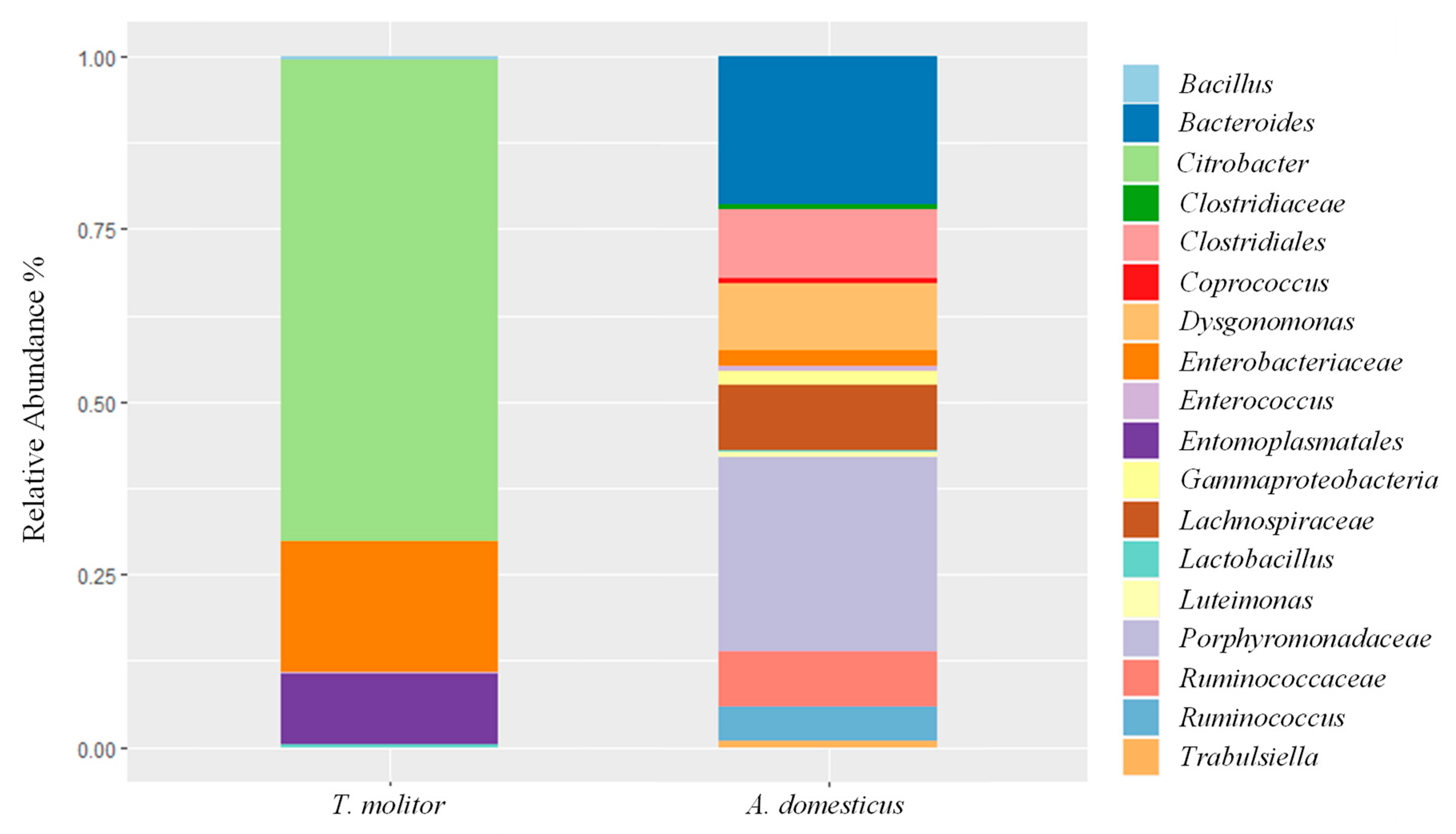
3.2. Culture-Independent Microbiological Analysis
3.3. Characterization of LAB
3.4. Proximate Composition, Energy and Fatty Acid Profile of Insect Powders
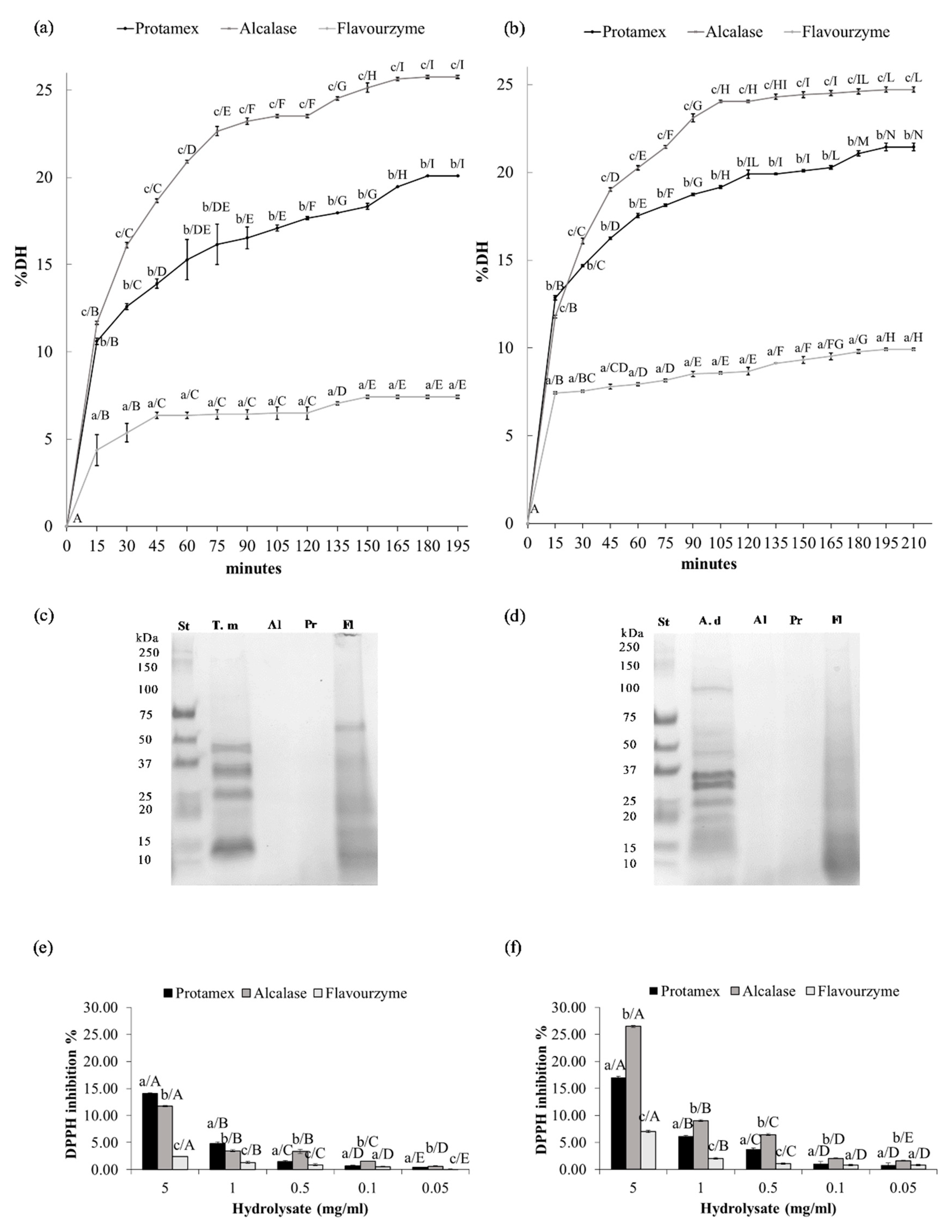
3.5. Enzymatic Hydrolysis
3.6. SDS PAGE
3.7. DPPH Radical Scavenging Activity
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Belluco, S.; Losasso, C.; Ricci, A.; Maggioletti, M.; Alonzi, C.; Paoletti, M.G. Edible insects: A food security solution or a food safety concern? Anim. Front. 2015, 5, 25–30. [Google Scholar]
- van Huis, A. Potential of Insects as Food and Feed in Assuring Food Security. Annu. Rev. Entomol. 2013, 58, 563–583. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Elorduy, J.; Moreno, J.M.P.; Prado, E.E.; Perez, M.A.; Otero, J.L.; de Guevara, O.L. Nutritional Value of Edible Insects from the State of Oaxaca, Mexico. J. Food Compos. Anal. 1997, 10, 142–157. [Google Scholar] [CrossRef]
- Chapagain, A.K.; Hoekstra, A.Y. Virtual Water Flows between Nations in Relation to Trade in Livestock and Livestock Products Value of Water; UNESCO-IHE: DA Delft, The Netherlands, 2003. [Google Scholar]
- Oonincx, D.G.A.B.; van Itterbeeck, J.; Heetkamp, M.J.W.; van den Brand, H.; van Loon, J.J.A.; van Huis, A. An Exploration on Greenhouse Gas and Ammonia Production by Insect Species Suitable for Animal or Human Consumption. PLoS ONE 2010, 5, e14445. [Google Scholar] [CrossRef] [PubMed]
- Woolf, E.; Zhu, Y.; Emory, K.; Zhao, J.; Liu, C. Willingness to consume insect-containing foods: A survey in the United States. LWT-Food Sci. Technol. 2019, 102, 100–105. [Google Scholar] [CrossRef]
- Orsi, L.; Voege, L.L.; Stranieri, S. Eating edible insects as sustainable food? Exploring the determinants of consumer acceptance in Germany. Food Res. Int. 2019, 125, 108573. [Google Scholar] [CrossRef]
- Tan, H.S.G.; Fischer, A.R.H.; Tinchan, P.; Stieger, M.; Steenbekkers, L.P.A.; van Trijp, H.C.M. Insects as food: Exploring cultural exposure and individual experience as determinants of acceptance. Food Qual. Prefer. 2015, 42, 78–89. [Google Scholar] [CrossRef]
- Nonaka, K. Feasting on insects. Entomol. Res. 2009, 39, 304–312. [Google Scholar] [CrossRef]
- Durst, P.B.; Johnson, D.V.; Leslie, R.N.; Shono, K. Forest Insects as Food: Humans Bite back Proceedings of a Workshop on Asia-Pacific Resources and Their Potential for Development; Food and Agriculture of the United Nations: Rome, Italy, 2010; ISBN 9789251064887. [Google Scholar]
- Klunder, H.C.; Wolkers-Rooijackers, J.; Korpela, J.M.; Nout, M.J.R. Microbiological aspects of processing and storage of edible insects. Food Control 2012, 26, 628–631. [Google Scholar] [CrossRef]
- Aguilar-Miranda, E.D.; López, M.G.; Escamilla-Santana, C.; Barba de la Rosa, A.P. Characteristics of Maize Flour Tortilla Supplemented with Ground Tenebrio molitor Larvae. J. Agric. Food Chem. 2001, 50, 192–195. [Google Scholar] [CrossRef]
- Kim, H.-W.; Setyabrata, D.; Lee, Y.J.; Jones, O.G.; Kim, Y.H.B. Pre-treated mealworm larvae and silkworm pupae as a novel protein ingredient in emulsion sausages. Innov. Food Sci. Emerg. Technol. 2016, 38, 116–123. [Google Scholar] [CrossRef]
- Bußler, S.; Rumpold, B.A.; Jander, E.; Rawel, H.M.; Schlüter, O.K. Recovery and techno-functionality of flours and proteins from two edible insect species: Meal worm (Tenebrio molitor) and black soldier fly (Hermetia illucens) larvae. Heliyon 2016, 2, e00218. [Google Scholar] [CrossRef] [PubMed]
- Megido, R.C.; Desmedt, S.; Blecker, C.; Béra, F.; Haubruge, É.; Alabi, T.; Francis, F. Microbiological load of edible insects found in Belgium. Insects 2017, 8, 1–8. [Google Scholar]
- Garofalo, C.; Osimani, A.; Milanović, V.; Taccari, M.; Cardinali, F.; Aquilanti, L.; Riolo, P.; Ruschioni, S.; Isidoro, N.; Clementi, F. The microbiota of marketed processed edible insects as revealed by high-throughput sequencing. Food Microbiol. 2017, 62, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Osimani, A.; Milanović, V.; Cardinali, F.; Garofalo, C.; Clementi, F.; Pasquini, M.; Riolo, P.; Ruschioni, S.; Isidoro, N.; Loreto, N.; et al. The bacterial biota of laboratory-reared edible mealworms (Tenebrio molitor L.): From feed to frass. Int. J. Food Microbiol. 2018, 272, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Osimani, A.; Garofalo, C.; Milanovic´, V.; Milanovic´, M.; Taccari, M.; Cardinali, F.; Aquilanti, L.; Pasquini, M.; Mozzon, M.; Raffaelli, N.; et al. Insight into the proximate composition and microbial diversity of edible insects marketed in the European Union. Eur. Food Res. Technol. 2017, 243, 1157–1171. [Google Scholar] [CrossRef]
- Osimani, A.; Milanović, V.; Cardinali, F.; Roncolini, A.; Garofalo, C.; Clementi, F.; Pasquini, M.; Mozzon, M.; Foligni, R.; Raffaelli, N.; et al. Bread enriched with cricket powder (Acheta domesticus): A technological, microbiological and nutritional evaluation. Innov. Food Sci. Emerg. Technol. 2018, 48, 150–163. [Google Scholar] [CrossRef]
- Hall, F.G.; Jones, O.G.; O’Haire, M.E.; Liceaga, A.M. Functional properties of tropical banded cricket (Gryllodes sigillatus) protein hydrolysates. Food Chem. 2017, 224, 414–422. [Google Scholar] [CrossRef]
- Hall, F.; Johnson, P.E.; Liceaga, A. Effect of enzymatic hydrolysis on bioactive properties and allergenicity of cricket (Gryllodes sigillatus) protein. Food Chem. 2018, 262, 39–47. [Google Scholar] [CrossRef]
- Li-Chan, E.C. Bioactive peptides and protein hydrolysates: Research trends and challenges for application as nutraceuticals and functional food ingredients. Curr. Opin. Food Sci. 2015, 1, 28–37. [Google Scholar] [CrossRef]
- Tang, Y.; Debnath, T.; Choi, E.-J.; Kim, Y.W.; Ryu, J.P.; Jang, S.; Chung, S.U.; Choi, Y.-J.; Kim, E.-K. Changes in the amino acid profiles and free radical scavenging activities of Tenebrio molitor larvae following enzymatic hydrolysis. PLoS ONE 2018, 13, e0196218. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, A.; Bunge, J. Estimating the Number of Species in a Stochastic Abundance Model. Biometrics 2002, 58, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shen, D.; Fang, Z.; Jie, Z.; Qiu, X.; Zhang, C.; Chen, Y.; Ji, L. Human Gut Microbiota Changes Reveal the Progression of Glucose Intolerance. PLoS ONE 2013, 8, e71108. [Google Scholar] [CrossRef] [PubMed]
- Gregersen, T. Rapid method for distinction of gram-negative from gram-positive bacteria. Eur. J. Appl. Microbiol. Biotechnol. 1978, 5, 123–127. [Google Scholar] [CrossRef]
- Alfonzo, A.; Ventimiglia, G.; Corona, O.; Di Gerlando, R.; Gaglio, R.; Francesca, N.; Moschetti, G.; Settanni, L. Diversity and technological potential of lactic acid bacteria of wheat flours. Food Microbiol. 2013, 36, 343–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaglio, R.; Francesca, N.; Di Gerlando, R.; Mahony, J.; De Martino, S.; Stucchi, C.; Moschetti, G.; Settanni, L. Enteric bacteria of food ice and their survival in alcoholic beverages and soft drinks. Food Microbiol. 2017, 67, 17–22. [Google Scholar] [CrossRef] [Green Version]
- Jackson, C.R.; Fedorka-Cray, P.J.; Barrett, J.B. Use of a genus- and species-specific multiplex PCR for identification of enterococci. J. Clin. Microbiol. 2004, 42, 3558–3565. [Google Scholar] [CrossRef]
- Clinical Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing; Twenty-seventh Informational Supplement (M100); CLSI: Wayne, PA, USA, 2017. [Google Scholar]
- Lopes, M.D.F.S.; Simões, A.P.; Tenreiro, R.; Marques, J.J.F.; Crespo, M.T.B. Activity and expression of a virulence factor, gelatinase, in dairy enterococci. Int. J. Food Microbiol. 2006, 112, 208–214. [Google Scholar] [CrossRef]
- AOAC Official methods of analysis. J. Pharm. Sci. 1990, 1, 1230.
- AOAC Association of Official Analytical Chemists Official Method, 981.10 Crude protein in meat block digestion method. J. AOAC Int. 1992, 65, 1339.
- Díaz-Rojas, E.I.; Argüelles-Monal, W.M.; Higuera-Ciapara, I.; Hernández, J.; Lizardi-Mendoza, J.; Goycoolea, F.M. Determination of chitin and protein contents during the isolation of chitin from shrimp waste. Macromol. Biosci. 2006, 6, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Lepage, G.; Roy, C.C. Improved recovery of fatty acid through direct transesterification without prior extraction or purification. J. Lipid Res. 1984, 25, 1391–1396. [Google Scholar] [PubMed]
- Messina, C.M.; Renda, G.; La Barbera, L.; Santulli, A. By-products of farmed European sea bass (Dicentrarchus labrax L.) as a potential source of n-3 PUFA. Biologia 2013, 68, 288–293. [Google Scholar] [CrossRef]
- Messina, C.M.; Renda, G.; Randazzo, M.; Laudicella, V.A.; Gharbi, S.; Pizzo, F.; Morghese, M.; Santulli, A. Extraction of bioactive compounds from shrimp waste. Bull. Inst. Natl. Sci. Tech. Mer de Salammbô 2015, 42, 27–29. [Google Scholar]
- Dumay, J.; Donnay-Moreno, C.; Barnathan, G.; Jaouen, P.; Bergé, J.P. Improvement of lipid and phospholipid recoveries from sardine (Sardina pilchardus) viscera using industrial proteases. Process Biochem. 2006, 41, 2327–2332. [Google Scholar] [CrossRef]
- de Holanda, H.D.; Netto, F.M. Recovery of Components from Shrimp (Xiphopenaeus kroyeri) Processing Waste by Enzymatic Hydrolysis. J. Food Sci. 2006, 71, C298–C303. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–267. [Google Scholar]
- Sharma, O.P.; Bhat, T.K. DPPH antioxidant assay revisited. Food Chem. 2009, 113, 1202–1205. [Google Scholar] [CrossRef]
- Manuguerra, S.; Caccamo, L.; Mancuso, M.; Arena, R.; Rappazzo, A.C.; Genovese, L.; Santulli, A.; Messina, C.M.; Maricchiolo, G. The antioxidant power of horseradish, Armoracia rusticana, underlies antimicrobial and antiradical effects, exerted in vitro. Nat. Prod. Res. 2018, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Osimani, A.; Milanović, V.; Cardinali, F.; Garofalo, C.; Clementi, F.; Ruschioni, S.; Riolo, P.; Isidoro, N.; Loreto, N.; Galarini, R.; et al. Distribution of transferable antibiotic resistance genes in laboratory-reared edible mealworms (Tenebrio molitor L.). Front. Microbiol. 2018, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Motcha Anthony Reetha, B.; Mohan, M. Diversity of commensal bacteria from mid-gut of pink stem borer (Sesamia inferens [Walker])-Lepidoptera insect populations of India. J. Asia. Pac. Entomol. 2018, 21, 937–943. [Google Scholar] [CrossRef]
- Sakamoto, M. The Family Porphyromonadaceae. In The Prokaryotes; Springer: Berlin/Heidelberg, Germany, 2014; pp. 811–824. [Google Scholar]
- Gaglio, R.; Cruciata, M.; Di Gerlando, R.; Scatassa, M.L.; Cardamone, C.; Mancuso, I.; Sardina, M.T.; Moschetti, G.; Portolano, B.; Settanni, L. Microbial activation of wooden vats used for traditional cheese production and evolution of neoformed biofilms. Appl. Environ. Microbiol. 2016, 82, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Gaglio, R.; Francesca, N.; Maniaci, G.; Corona, O.; Alfonzo, A.; Giosuè, C.; Di Noto, A.; Cardamone, C.; Sardina, M.T.; Portolano, B.; et al. Valorization of indigenous dairy cattle breed through salami production. Meat Sci. 2016, 114, 58–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfonzo, A.; Gaglio, R.; Miceli, A.; Francesca, N.; Di Gerlando, R.; Moschetti, G.; Settanni, L. Shelf life evaluation of fresh-cut red chicory subjected to different minimal processes. Food Microbiol. 2018, 73, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Gaglio, R.; Couto, N.; Marques, C.; de Fatima Silva Lopes, M.; Moschetti, G.; Pomba, C.; Settanni, L. Evaluation of antimicrobial resistance and virulence of enterococci from equipment surfaces, raw materials, and traditional cheeses. Int. J. Food Microbiol. 2016, 236, 107–114. [Google Scholar] [CrossRef] [Green Version]
- Foulquié Moreno, M.R.; Sarantinopoulos, P.; Tsakalidou, E.; De Vuyst, L. The role and application of enterococci in food and health. Int. J. Food Microbiol. 2006, 106, 1–24. [Google Scholar] [CrossRef]
- Rumpold, B.A.; Schlüter, O.K. Nutritional composition and safety aspects of edible insects. Mol. Nutr. Food Res. 2013, 57, 802–823. [Google Scholar] [CrossRef]
- Zielińska, E.; Baraniak, B.; Karaś, M.; Rybczyńska, K.; Jakubczyk, A. Selected species of edible insects as a source of nutrient composition. Food Res. Int. 2015, 77, 460–466. [Google Scholar] [CrossRef]
- Finke, M.D. Complete nutrient composition of commercially raised invertebrates used as food for insectivores. Zoo Biol. 2002, 21, 269–285. [Google Scholar] [CrossRef]
- Sánchez-Muros, M.J.; Barroso, F.G.; Manzano-Agugliaro, F. Insect meal as renewable source of food for animal feeding: A review. J. Clean. Prod. 2014, 65, 16–27. [Google Scholar] [CrossRef]
- Yang, L.-F.; Siriamornpun, S.; Li, D. Polyunsaturated fatty acid content of edible insects in Thailand. J. Food Lipids 2006, 13, 277–285. [Google Scholar] [CrossRef]
- Baek, H.H.; Cadwallader, K.R. Enzymatic Hydrolysis of Crayfish Processing By-products. J. Food Sci. 1995, 60, 929–935. [Google Scholar] [CrossRef]
- Mizani, M.; Aminlari, M.; Khodabandeh, M. An Effective Method for Producing a Nutritive Protein Extract Powder from Shrimp-head Waste. Food Sci. Technol. Int. 2005, 11, 49–54. [Google Scholar] [CrossRef]
- Quaglia, G.B.; Orban, E. Influence of the degree of hydrolysis on the solubility of the protein hydrolysates from sardine (Sardina pilchardus). J. Sci. Food Agric. 1987, 38, 271–276. [Google Scholar] [CrossRef]
- Rebeca, B.D.; Peña-Vera, M.T.; Díaz-Castañeda, M. Production of Fish Protein Hydrolysates with Bacterial Proteases; Yield and Nutritional Value. J. Food Sci. 1991, 56, 309–314. [Google Scholar] [CrossRef]
- Shahidi, F.; Han, X.-Q.; Synowiecki, J. Production and characteristics of protein hydrolysates from capelin (Mallotus villosus). Food Chem. 1995, 53, 285–293. [Google Scholar] [CrossRef]
- Yang, R.; Zhao, X.; Kuang, Z.; Ye, M.; Luo, G.; Xiao, G.; Liao, S.; Li, L.; Xiong, Z. Optimization of antioxidant peptide production in the hydrolysis of silkworm (Bombyx mori L.) pupa protein using response surface methodology. J. Food Agric. Environ. 2013, 11, 952–956. [Google Scholar]
- Abtahi, S.; Aminlari, M. Effect of Sodium Sulfite, Sodium Bisulfite, Cysteine, and pH on Protein Solubility and Sodium Dodecyl Sulfate−Polyacrylamide Gel Electrophoresis of Soybean Milk Base. J. Agric. Food Chem. 1997, 45, 4768–4772. [Google Scholar] [CrossRef]
- Kristinsson, H.G.; Rasco, B.A. Fish Protein Hydrolysates: Production, Biochemical, and Functional Properties. Crit. Rev. Food Sci. Nutr. 2000, 40, 43–81. [Google Scholar] [CrossRef] [PubMed]
- Picot, L.; Ravallec, R.; Fouchereau-Péron, M.; Vandanjon, L.; Jaouen, P.; Chaplain-Derouiniot, M.; Guérard, F.; Chabeaud, A.; LeGal, Y.; Alvarez, O.M.; et al. Impact of ultrafiltration and nanofiltration of an industrial fish protein hydrolysate on its bioactive properties. J. Sci. Food Agric. 2010, 90, 1819–1826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raghavan, S.; Kristinsson, H.G. Antioxidative Efficacy of Alkali-Treated Tilapia Protein Hydrolysates: A Comparative Study of Five Enzymes. J. Agric. Food Chem. 2008, 56, 1434–1441. [Google Scholar] [CrossRef] [PubMed]
- Thiansilakul, Y.; Benjakul, S.; Shahidi, F. Compositions, functional properties and antioxidative activity of protein hydrolysates prepared from round scad (Decapterus maruadsi). Food Chem. 2007, 103, 1385–1394. [Google Scholar] [CrossRef]
- Ahn, C.-B.; Kim, J.-G.; Je, J.-Y. Purification and antioxidant properties of octapeptide from salmon byproduct protein hydrolysate by gastrointestinal digestion. Food Chem. 2014, 147, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Taheri, A.; Sabeena Farvin, K.H.; Jacobsen, C.; Baron, C.P. Antioxidant activities and functional properties of protein and peptide fractions isolated from salted herring brine. Food Chem. 2014, 142, 318–326. [Google Scholar] [CrossRef]
- Allen, L.; De Benoist, B.; Dary, O.; Hurrell, R. Guidelines on Food Fortification with Micronutrients; WHO: Geneva, Switzerland, 2006; ISBN 9241594012. [Google Scholar]
- Starčević, K.; Gavrilović, A.; Gottstein, Ž.; Mašek, T. Influence of substitution of sunflower oil by different oils on the growth, survival rate and fatty acid composition of Jamaican field cricket (Gryllus assimilis). Anim. Feed Sci. Technol. 2017, 228, 66–71. [Google Scholar] [CrossRef]
- Wang, D.; Zhai, S.W.; Zhang, C.X.; Zhang, Q.; Chen, H. Nutrition value of the Chinese grasshopper Acrida cinerea (Thunberg) for broilers. Anim. Feed Sci. Technol. 2007, 135, 66–74. [Google Scholar] [CrossRef]
- Mlcek, J.; Borkovcova, M.; Rop, O.; Bednarova, M. Biologically active substances of edible insects and their use in agriculture, veterinary and human medicine—A review. J. Cent. Eur. Agric. 2014, 15, 225–237. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proximate Composition | Tenebrio molitor (T. molitor) | Acheta domesticus (A. domesticus) |
|---|---|---|
| Moisture | 3.12 ± 0.39 | 4.72 ± 0.15 |
| Ash | 3.32 ± 0.04 | 4.49 ± 0.04 |
| Lipid | 26.17 ± 0.21 | 21.66 ± 0.13 |
| Protein | 52.95 ± 0.33 | 63.62 ± 0.5 |
| Chitin | 14.42 ± 0.33 | 5.50 ± 0.5 |
| Energy | 1868.59 ± 2.26 | 1882.88 ± 3.65 |
| Fatty acids | ||
| Myristic acid (14:0) | 2.35 ± 0.07 | 1.66 ± 0.07 |
| Palmitic acid (16:0) | 17.96 ± 0.14 | 25.56 ± 0.44 |
| Palmitoleic acid (16:1 n-7) | 1.67 ± 0.01 | 0.81 ± 0.02 |
| 9,12-Hexadecadienoic acid (16:2 n-4) | 0.02 ± 0.01 | 0.08 ± 0.00 |
| 6,9,12-Hexadecatrienoic acid (16:3 n-4) | 0.15 ± 0.00 | 0.11 ± 0.00 |
| Stearic acid (18:0) | 3.34 ± 0.05 | 12.47 ± 0.10 |
| Oleic acid (18:1 n-9) | 45.75 ± 0.38 | 22.59 ± 0.04 |
| Vaccenic acid (18:1 n-7) | 0.50 ± 0.66 | 0.93 ± 0.02 |
| Linoleic acid (18:2 n-6) | 25.73 ± 0.18 | 32.35 ± 0.42 |
| γ-linolenic acid (18:3 n-6) | n.d. | n.d. |
| 8,11,14-Octadecatrienoic acid (18:3 n-4) | 0.01 ± 0.02 | n.d. |
| α-Linolenic acid (18:3 n-3) | 2.30 ± 0.03 | 1.75 ± 0.01 |
| Stearidonic acid (18:4 n-3) | n.d. | 0.04 ± 0.04 |
| Eicosenoic acid (20:1 n-9) | 0.13 ± 0.04 | 0.21 ± 0.00 |
| Arachidonic acid (20:4 n-6) | n.d. | 0.14 ± 0.01 |
| Eicosatetraenoic acid (20:4, n-3) | n.d. | n.d. |
| Eicosapentaenoic acid (20:5, n-3) | 0.01 ± 0.01 | 0.48 ± 0.04 |
| Cetoleic acid (22:1 n-11) | 0.01 ± 0.02 | 0.11 ± 0.00 |
| Erucic acid (22:1 n-9) | 0.01 ± 0.02 | 0.01 ± 0.00 |
| Adrenic acid (22:4 n-6) | 0.03 ± 0.05 | n.d. |
| Osbond acid (22:5 n-6) | 0.00 ± 0.01 | n.d. |
| Docosapentaenoic acid (22:5 n-3) | n.d. | 0.10 ± 0.01 |
| Docosahexaenoic acid (22:6 n-3) | 0.02 ± 0.02 | 0.63 ± 0.02 |
| Nervonic acid (24:1 n-9) | 2.35 ± 0.07 | 0.00 ± 0.00 |
| Saturated | 23.65 ± 0.12 | 39.68 ± 0.41 |
| Monounsaturated | 48.06 ± 0.20 | 24.66 ± 0.00 |
| Polyunsaturated | 28.28 ± 0.31 | 35.66 ± 0.41 |
| Total n-3 | 2.33 ± 0.07 | 2.99 ± 0.02 |
| Total n-6 | 25.77 ± 0.24 | 32.49 ± 0.43 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Messina, C.M.; Gaglio, R.; Morghese, M.; Tolone, M.; Arena, R.; Moschetti, G.; Santulli, A.; Francesca, N.; Settanni, L. Microbiological Profile and Bioactive Properties of Insect Powders Used in Food and Feed Formulations. Foods 2019, 8, 400. https://doi.org/10.3390/foods8090400
Messina CM, Gaglio R, Morghese M, Tolone M, Arena R, Moschetti G, Santulli A, Francesca N, Settanni L. Microbiological Profile and Bioactive Properties of Insect Powders Used in Food and Feed Formulations. Foods. 2019; 8(9):400. https://doi.org/10.3390/foods8090400
Chicago/Turabian StyleMessina, Concetta Maria, Raimondo Gaglio, Maria Morghese, Marco Tolone, Rosaria Arena, Giancarlo Moschetti, Andrea Santulli, Nicola Francesca, and Luca Settanni. 2019. "Microbiological Profile and Bioactive Properties of Insect Powders Used in Food and Feed Formulations" Foods 8, no. 9: 400. https://doi.org/10.3390/foods8090400
APA StyleMessina, C. M., Gaglio, R., Morghese, M., Tolone, M., Arena, R., Moschetti, G., Santulli, A., Francesca, N., & Settanni, L. (2019). Microbiological Profile and Bioactive Properties of Insect Powders Used in Food and Feed Formulations. Foods, 8(9), 400. https://doi.org/10.3390/foods8090400