

Phenolic Profile and the Antioxidant, Anti-Inflammatory, and Antimicrobial Properties of Açaí (Euterpe oleracea) Meal: A Prospective Study
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Açaí Byproduct
2.2. Chemicals and Microbial Strains
2.3. LC-ESI-QTOF-MS/MS Phenolic Profile
2.4. Total Phenolic Compounds, Reducing Power and Free Fadical Scavenging Capacity of Açaí Meal Extract
2.4.1. Total Phenolic Compounds (TPC) and Reducing Power
2.4.2. Free Radical Scavenging Capacity
2.5. In Vitro Cytotoxicity and Anti-Inflammatory Activity
2.5.1. Cell Culture
2.5.2. Cell Viability Assay
2.5.3. NF-κB Activation and TNF-α Quantification
2.6. Antimicrobial Activity of the Açaí Meal Extract
2.6.1. Strains and Growth Conditions
2.6.2. Minimum Inhibitory Concentration (MIC)
2.6.3. Minimum Bactericidal and Fungicidal Concentration (MBC/MFC)
2.7. Statistical Analysis
3. Results and Discussion
3.1. Comprehensive Chemical Characterization of the Extract from Açaí Meal
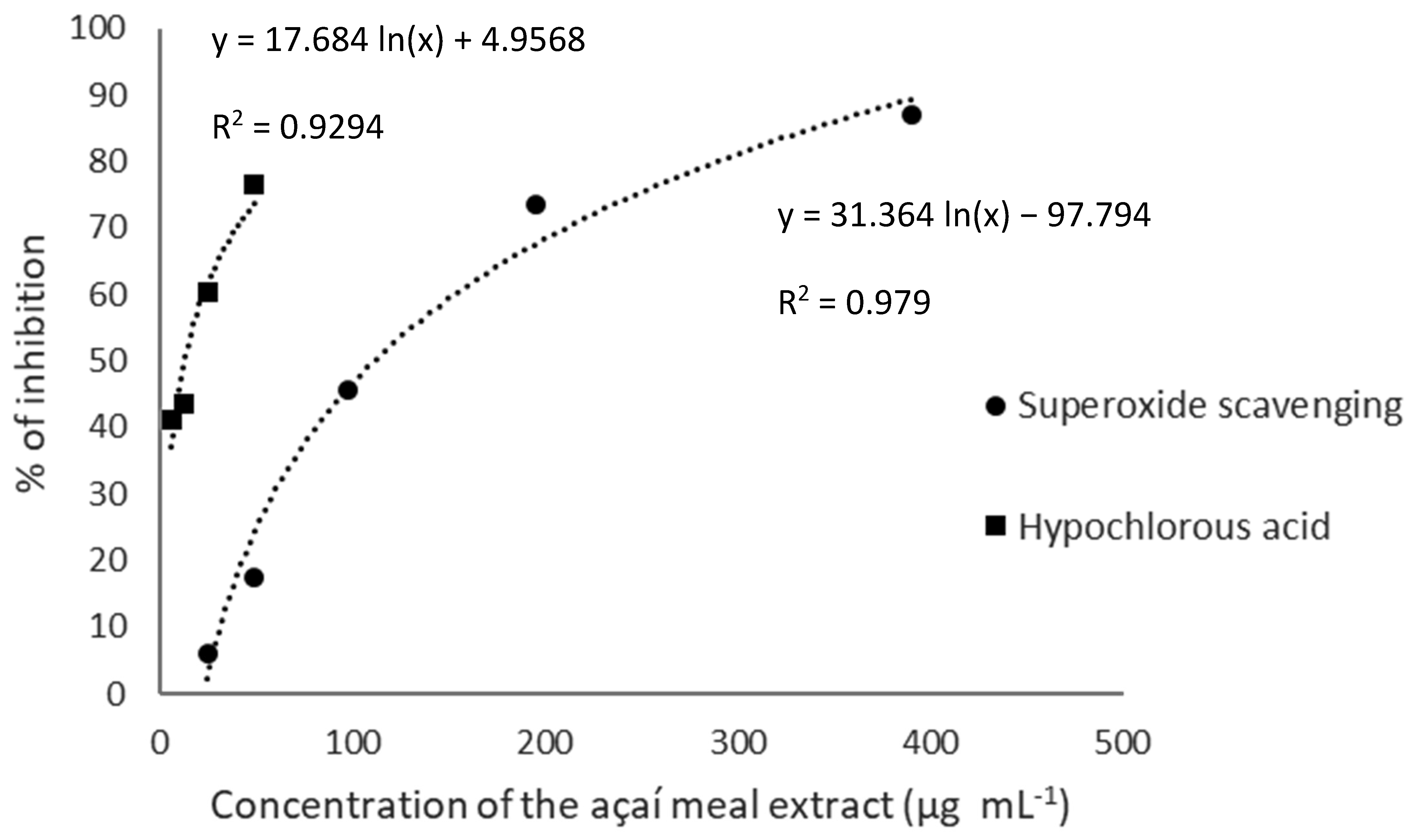
3.2. Antioxidant Capacity: TPC, Reducing Power and Free Radical Scavenging
3.3. In Vitro Cytotoxicity and Anti-Inflammatory Activity
3.4. Antimicrobial Activity
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leal, C.; Gouvinhas, I.; Santos, R.A.; Rosa, E.; Silva, A.M.; Saavedra, M.J.; Barros, A.I. Potential application of grape (Vitis vinifera L.) stem extracts in the cosmetic and pharmaceutical industries: Valorization of a by-product. Ind. Crop. Prod. 2020, 154, 112675. [Google Scholar] [CrossRef]
- Chang, S.K.; Alasalvar, C.; Shahidi, F. Superfruits: Phytochemicals, antioxidant efficacies, and health effects—A comprehensive review. Crit. Rev. Food Sci. Nutr. 2018, 59, 1580–1604. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, K.K.D.L.; Pereira, L.F.R.; Lamarão, C.V.; Lima, E.S.; da Veiga-Junior, V.F. Amazon acai: Chemistry and biological activities: A review. Food Chem. 2015, 179, 137–151. [Google Scholar] [CrossRef] [PubMed]
- IBGE. The Brazilian Institute of Geography and Statistics. Production of Plant Extraction and Forestry. Vegetable Extraction: Table 1—Amount Produced and the Production Value of Brazil, Great Regions and Federal Units, According to the Extractive Products. 2021. Available online: https://www.ibge.gov.br/estatisticas/economicas/agricultura-e-pecuaria/9105-producao-da-extracao-vegetal-e-da-silvicultura.html?=&t=resultados (accessed on 10 October 2022).
- Oliveira, M.S.P.; Schwartz, G. oleraceae. In Exotic Fruits; Academic Press: Cambridge, MA, USA, 2018; pp. 1–5. [Google Scholar]
- Kang, J.; Xie, C.H.; Li, Z.M.; Nagarajan, S.; Schauss, A.G.; Wu, T.; Wu, X.L. Flavonoids from acai (Euterpe oleraceae Mart.) pulp and their antioxidant and anti-inflammatory activities. Food Chem. 2011, 128, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Embrapa. Empresa Brasileira de Pesquisa Agropecuária, Sistemas de Produção 4—Açaí; Embrapa Amazônia Oriental: Belém, PA, USA, 2005; p. 137. [Google Scholar]
- Melo, P.S.; Selani, M.M.; Gonçalves, R.H.; Paulino, J.d.O.; Massarioli, A.P.; de Alencar, S.M. Açaí seeds: An unexplored agro-industrial residue as a potential source of lipids, fibers, and antioxidant phenolic compounds. Ind. Crop. Prod. 2021, 161, 113204. [Google Scholar] [CrossRef]
- Yi, C.; Shi, J.; Kramer, J.; Xue, S.; Jiang, Y.; Zhang, M.; Ma, Y.; Pohorly, J. Fatty acid composition and phenolic antioxidants of winemaking pomace powder. Food Chem. 2009, 114, 570–576. [Google Scholar] [CrossRef]
- Hogan, S.; Chung, H.; Zhang, L.; Li, J.; Lee, Y.; Dai, Y.; Zhou, K. Antiproliferative and antioxidant properties of anthocyanin-rich extract from açai. Food Chem. 2010, 118, 208–214. [Google Scholar] [CrossRef]
- Schreckinger, M.E.; Lotton, J.; Lila, M.A.; De Mejia, E.G. Berries from South America: A Comprehensive Review on Chemistry, Health Potential, and Commercialization. J. Med. Food 2010, 13, 233–246. [Google Scholar] [CrossRef]
- Rufino, M.D.S.M.; Pérez-Jiménez, J.; Arranz, S.; Alves, R.E.; de Brito, E.S.; Oliveira, M.S.; Saura-Calixto, F. Açaí (Euterpe oleraceaee) ‘BRS Pará’: A tropical fruit source of antioxidant dietary fiber and high antioxidant capacity oil. Food Res. Int. 2011, 44, 2100–2106. [Google Scholar] [CrossRef] [Green Version]
- Wong, D.Y.S.; Musgrave, I.F.; Harvey, B.S.; Smid, S.D. Açaí (Euterpe oleraceaee Mart.) berry extract exerts neuroprotective effects against β-amyloid exposure in vitro. Neurosci. Lett. 2013, 556, 221–226. [Google Scholar] [CrossRef]
- Bonomo, L.D.F.; Silva, D.N.; Boasquivis, P.F.; Paiva, F.A.; Guerra, J.F.D.C.; Martins, T.A.F.; Torres, G.D.J.; de Paula, I.T.B.R.; Caneschi, W.L.; Jacolot, P.; et al. Açaí (Euterpe oleraceae Mart.) Modulates Oxidative Stress Resistance in Caenorhabditis elegans by Direct and Indirect Mechanisms. PLoS ONE 2014, 9, e89933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, D.S.; Gomes, A.L.; Da Silva, M.G.; Alves, A.B.; Agnol, W.H.D.; Ferrari, R.A.; Carvalho, P.R.N.; Pacheco, M.T.B. Antioxidant Capacity and Chemical Characterization of Açaí (Euterpe oleraceae Mart.) Fruit Fractions. Food Sci. Technol. 2016, 4, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Linan, L.Z.; Cidreira, A.C.M.; da Rocha, C.Q.; de Menezes, F.F.; de Moraes Rocha, G.J.; Paiva, A.E.M. Utilization of Acai Berry Residual Biomass for Extraction of Lignocellulosic Byproducts. J. Bioresour. Bioprod. 2021, 6, 323–337. [Google Scholar] [CrossRef]
- Rossetto, R.; Maciel, G.M.; Bortolini, D.G.; Ribeiro, V.R.; Haminiuk, C.W.I. Acai pulp and seeds as emerging sources of phenolic compounds for enrichment of residual yeasts (Saccharomyces cerevisiae) through biosorption process. LWT 2020, 128, 109447. [Google Scholar] [CrossRef]
- Tyug, T.S.; Prasad, K.N.; Ismail, A. Antioxidant capacity, phenolics and isoflavones in soybean by-products. Food Chem. 2010, 123, 583–589. [Google Scholar] [CrossRef]
- Zamindar, N.; Bashash, M.; Khorshidi, F.; Serjouie, A.; Shirvani, M.A.; Abbasi, H.; Sedaghatdoost, A. Antioxidant efficacy of soybean cake extracts in soy oil protection. J. Food Sci. Technol. 2017, 54, 2077–2084. [Google Scholar] [CrossRef]
- Halliwell, B.; Aeschbach, R.; Löliger, J.; Aruoma, O. The characterization of antioxidants. Food Chem. Toxicol. 1995, 33, 601–617. [Google Scholar] [CrossRef]
- Gomes, A.; Fernandes, E.; Silva, A.M.; Santos, C.M.; Pinto, D.C.; Cavaleiro, J.A.; Lima, J.L. 2-Styrylchromones: Novel strong scavengers of reactive oxygen and nitrogen species. Bioorganic Med. Chem. 2007, 15, 6027–6036. [Google Scholar] [CrossRef]
- Li, H.; Christman, L.M.; Li, R.; Gu, L. Synergic interactions between polyphenols and gut microbiota in mitigating inflammatory bowel diseases. Food Funct. 2020, 11, 4878–4891. [Google Scholar] [CrossRef]
- Romão, M.H.; de Bem, G.; Santos, I.B.; Soares, R.D.A.; Ognibene, D.; de Moura, R.S.; da Costa, C.A.; Resende, C. Açaí (Euterpe oleraceae Mart.) seed extract protects against hepatic steatosis and fibrosis in high-fat diet-fed mice: Role of local renin-angiotensin system, oxidative stress and inflammation. J. Funct. Foods 2019, 65, 103726. [Google Scholar] [CrossRef]
- Martinez, R.M.; Guimarães, D.D.A.B.; Berniz, C.R.; de Abreu, J.P.; da Rocha, A.P.M.; de Moura, R.S.; Resende, A.C.; Teodoro, A.J. Açai (Euterpe oleraceae Mart.) Seed Extract Induces Cell Cycle Arrest and Apoptosis in Human Lung Carcinoma Cells. Foods 2018, 7, 178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wedick, N.M.; Pan, A.; Cassidy, A.; Rimm, E.B.; Sampson, L.; Rosner, B.; Willett, W.; Hu, F.B.; Sun, Q.; van Dam, R.M. Dietary flavonoid intakes and risk of type 2 diabetes in US men and women. Am. J. Clin. Nutr. 2012, 95, 925–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Circu, M.L.; Aw, T.Y. Reactive oxygen species, cellular redox systems, and apoptosis. Free Radic. Biol. Med. 2010, 48, 749–762. [Google Scholar] [CrossRef] [Green Version]
- Carpes, S.T.; Pereira, D.; De Moura, C.; Dos Reis, A.S.; Da Silva, L.D.; Oldoni, T.L.C.; Almeida, J.F.; Plata-Oviedo, M.V.S. Lyophilized and microencapsulated extracts of grape pomace from winemaking industry to prevent lipid oxidation in chicken pâté. Braz. J. Food Technol. 2020, 23, e2019112. [Google Scholar] [CrossRef]
- Silva, A.P.D.S.; Rosalen, P.L.; de Camargo, A.C.; Lazarini, J.G.; Rocha, G.; Shahidi, F.; Franchin, M.; de Alencar, S.M. Inajá oil processing by-product: A novel source of bioactive catechins and procyanidins from a Brazilian native fruit. Food Res. Int. 2021, 144, 110353. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Meth. Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Melo, P.S.; Massarioli, A.P.; Denny, C.; dos Santos, L.F.; Franchin, M.; Pereira, G.E.; Vieira, T.M.F.D.S.; Rosalen, P.L.; de Alencar, S.M. Winery by-products: Extraction optimization, phenolic composition and cytotoxic evaluation to act as a new source of scavenging of reactive oxygen species. Food Chem. 2015, 181, 160–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CLSI Document M27-A3; Clinical and Laboratory Standards Institute. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts. CLSI: Wayne, PA, USA, 2008.
- CLSI document M07-A9; Reference Method for Broth Dilution antibacterial Susceptibility Testing of Bacterial. CLSI: Wayne, PA, USA, 2012.
- Garzón, G.A.; Narváez-Cuenca, C.-E.; Vincken, J.-P.; Gruppen, H. Polyphenolic composition and antioxidant activity of açai (Euterpe oleraceae Mart.) from Colombia. Food Chem. 2017, 217, 364–372. [Google Scholar] [CrossRef]
- Fernandes, I.; Faria, A.; Azevedo, J.; Soares, S.; Calhau, C.; De Freitas, V.; Mateus, N. Influence of Anthocyanins, Derivative Pigments and Other Catechol and Pyrogallol-Type Phenolics on Breast Cancer Cell Proliferation. J. Agric. Food Chem. 2010, 58, 3785–3792. [Google Scholar] [CrossRef]
- Tan, C.; Kong, Y.; Tong, Y.; Deng, H.; Wang, M.; Zhao, Y.; Wan, M.; Lin, S.; Liu, X.; Meng, X.; et al. Anti-apoptotic effects of high hydrostatic pressure treated cyanidin-3-glucoside and blueberry pectin complexes on lipopolysaccharide-induced inflammation in Caco-2 cells. J. Funct. Foods 2021, 86, 104709. [Google Scholar] [CrossRef]
- Ma, M.-M.; Li, Y.; Liu, X.-Y.; Zhu, W.-W.; Ren, X.; Kong, G.-Q.; Huang, X.; Wang, L.-P.; Luo, L.-Q.; Wang, X.-Z. Cyanidin-3-O-glucoside Ameliorates Lipopolysaccharide-Induced Injury Both In Vivo and In Vitro Suppression of NF-κB and MAPK Pathways. Inflammation 2015, 38, 1669–1682. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.-H.; Guo, H.; Xu, W.-B.; Ge, J.; Li, X.; Alimu, M.; He, D.-J. Rapid Identification of Flavonoid Constituents Directly from PTP1B Inhibitive Extract of Raspberry (Rubus idaeus L.) Leaves by HPLC–ESI–QTOF–MS-MS. J. Chromatogr. Sci. 2016, 54, 805–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Rosso, V.; Hillebrand, S.; Montilla, E.C.; Bobbio, F.O.; Winterhalter, P.; Mercadante, A.Z. Determination of anthocyanins from acerola (Malpighia emarginata DC.) and açai (Euterpe oleraceae Mart.) by HPLC–PDA–MS/MS. J. Food Compos. Anal. 2008, 21, 291–299. [Google Scholar] [CrossRef]
- Zhang, Y.; Seeram, N.P.; Lee, R.; Feng, L.; Heber, D. Isolation and Identification of Strawberry Phenolics with Antioxidant and Human Cancer Cell Antiproliferative Properties. J. Agric. Food Chem. 2008, 56, 670–675. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, Y.; Zaima, N.; Moriyama, T.; Kawamura, Y. Different Localization Patterns of Anthocyanin Species in the Pericarp of Black Rice Revealed by Imaging Mass Spectrometry. PLoS ONE 2012, 7, e31285. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, H.; Zhang, G.; Meng, J.; Deng, K.; Zhou, W.; Wang, H.; Wang, Z.; Hu, N.; Suo, Y. Anthocyanins from Lycium ruthenicum Murr. Ameliorated d-galactose-inducedmemory impairment, oxidative stress, and neuroinfammation in adult rats. J. Agric. Food Chem. 2019, 67, 3140–3149. [Google Scholar] [CrossRef]
- Jugran, A.K.; Rawat, S.; Devkota, H.P.; Bhatt, I.D.; Rawal, R.S. Diabetes andplant-derived natural products: From ethnopharmacological approaches to theirpotential for modern drug discovery and development. Phytoth. Res. 2022, 35, 223–245. [Google Scholar] [CrossRef]
- Wei, J.; Zhang, G.; Zhang, X.; Xu, D.; Gao, J.; Fan, J.; Zhou, Z. Anthocyanins from Black Chokeberry (Aroniamelanocarpa Elliot) Delayed Aging-Related Degenerative Changes of Brain. J. Agric. Food Chem. 2017, 65, 5973–5984. [Google Scholar] [CrossRef]
- Liu, F.; Zhao, F.; Wang, W.; Sang, J.; Jia, L.; Li, L.; Lu, F. Cyanidin-3-O-glucoside inhibits Aβ40 fibrillogenesis, disintegrates preformed fibrils, and reduces amyloid cytotoxicity. Food Funct. 2020, 11, 2573–2587. [Google Scholar] [CrossRef]
- Li, L.; Zhou, P.; Wang, Y.; Pan, Y.; Chen, M.; Tian, Y.; Zhou, H.; Yang, B.; Meng, H.; Zheng, J. Antimicrobial activity of cyanidin-3-O-glucoside–lauric acid ester against Staphylococcus aureus and Escherichia coli. Food Chem. 2022, 383, 132410. [Google Scholar] [CrossRef]
- Soares, M.J.; Sampaio, G.R.; Guizellini, G.M.; Figueira, M.S.; Pinaffi, A.C.D.C.; Freitas, R.A.M.S.; Shahidi, F.; de Camargo, A.C.; Torres, E.A.F.D.S. Regular and decaffeinated espresso coffee capsules: Unravelling the bioaccessibility of phenolic compounds and their antioxidant properties in milk model system upon in vitro digestion. LWT 2020, 135, 110255. [Google Scholar] [CrossRef]
- Naveed, M.; Hejazi, V.; Abbas, M.; Kamboh, A.A.; Khan, G.J.; Shumzaid, M.; Ahmad, F.; Babazadeh, D.; Xia, F.F.; Modarresi-Ghazani, F.; et al. Chlorogenic acid (CGA): A pharmacological review and call for further research. Biomed. Pharmacother. 2018, 97, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xiong, H.; Xu, X.; Xue, X.; Liu, M.; Xu, S.; Liu, H.; Gao, Y.; Zhang, H.; Li, X. Compounds Identification in Semen Cuscutae by Ultra-High-Performance Liquid Chromatography (UPLCs) Coupled to Electrospray Ionization Mass Spectrometry. Molecules 2018, 23, 1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolniak-Ostek, J.; Oszmiański, J. Characterization of phenolic compounds in different anatomical pear (Pyrus communis L.) parts by ultra-performance liquid chromatography photodiode detector-quadrupole/time of flight-mass spectrometry (UPLC-PDA-Q/TOF-MS). Int. J. Mass Spectrom. 2015, 392, 154–163. [Google Scholar] [CrossRef]
- Brunschwig, C.; Leba, L.-J.; Saout, M.; Martial, K.; Bereau, D.; Robinson, J.-C. Chemical Composition and Antioxidant Activity of Euterpe oleraceae Roots and Leaflets. Int. J. Mol. Sci. 2016, 18, 61. [Google Scholar] [CrossRef] [Green Version]
- Zhong, L.; Lin, Y.; Wang, C.; Niu, B.; Xu, Y.; Zhao, G.; Zhao, J. Chemical Profile, Antimicrobial and Antioxidant Activity Assessment of the Crude Extract and Its Main Flavonoids from Tartary Buckwheat Sprouts. Molecules 2022, 27, 374. [Google Scholar] [CrossRef]
- Ferreres, F.; Silva, B.M.; Andrade, P.B.; Seabra, R.M.; Ferreira, M.A. Approach to the study of C-glycosyl flavones by ion trap HPLC-PAD-ESI/MS/MS: Application to seeds of quince (Cydonia oblonga). Phytochem. Anal. 2003, 14, 352–359. [Google Scholar] [CrossRef]
- Kim, M.K.; Yun, K.J.; Lim, D.H.; Kim, J.; Jang, Y.P. Anti-Inflammatory Properties of Flavone di-C-Glycosides as Active Principles of Camellia Mistletoe, Korthalsella japonica. Biomol. Ther. 2016, 24, 630–637. [Google Scholar] [CrossRef] [Green Version]
- Odontuya, G.; Hoult, J.R.S.; Houghton, P.J. Structure-activity relationship for antiinflammatory effect of luteolin and its derived glycosides. Phytother. Res. 2005, 19, 782–786. [Google Scholar] [CrossRef]
- Francisco, V.; Figueirinha, A.; Costa, G.; Liberal, J.; Lopes, M.C.; García-Rodríguez, C.; Geraldes, C.F.; Cruz, M.T.; Batista, M.T. Chemical characterization and anti-inflammatory activity of luteolin glycosides isolated from lemongrass. J. Funct. Foods 2014, 10, 436–443. [Google Scholar] [CrossRef]
- Jang, D.; Jung, Y.S.; Kim, M.-S.; Oh, S.E.; Nam, T.G.; Kim, D.-O. Developing and Validating a Method for Separating Flavonoid Isomers in Common Buckwheat Sprouts Using HPLC-PDA. Foods 2019, 8, 549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.-L.; Mutie, F.M.; Xu, Y.-B.; Saleri, F.D.; Hu, G.-W.; Guo, M.-Q. Antioxidant, Anti-inflammatory Activities and Polyphenol Profile of Rhamnus prinoides. Pharmaceuticals 2020, 13, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martini, N.; Katerere, D.; Eloff, J. Biological activity of five antibacterial flavonoids from Combretum erythrophyllum (Combretaceae). J. Ethnopharmacol. 2004, 93, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Bodakhe, S.H.; Ram, A.; Verma, S.; Pandey, D.P. Anticataract activity of rhamnocitrin isolated from Bauhinia variegata stem bark. Orient. Pharm. Exp. Med. 2012, 12, 227–232. [Google Scholar] [CrossRef]
- Cuoco, G.; Mathe, C.; Vieillescazes, C. Liquid chromatographic analysis of flavonol compounds in green fruits of three Rhamnus species used in Stil de grain. Microchem. J. 2014, 115, 130–137. [Google Scholar] [CrossRef]
- Kim, B.-R.; Paudel, S.B.; Han, A.-R.; Park, J.; Kil, Y.-S.; Choi, H.; Jeon, Y.G.; Park, K.Y.; Kang, S.-Y.; Jin, C.H.; et al. Metabolite Profiling and Dipeptidyl Peptidase IV Inhibitory Activity of Coreopsis Cultivars in Different Mutations. Plants 2021, 10, 1661. [Google Scholar] [CrossRef]
- Chen, M.; Zhang, X.; Wang, H.; Lin, B.; Wang, S.; Hu, G. Determination of Rutin in Rat Plasma by Ultra Performance Liquid Chromatography Tandem Mass Spectrometry and Application to Pharmacokinetic Study. J. Chromatogr. Sci. 2015, 53, 519–525. [Google Scholar] [CrossRef] [Green Version]
- Ben Sghaier, M.; Pagano, A.; Mousslim, M.; Ammari, Y.; Kovacic, H.; Luis, J. Rutin inhibits proliferation, attenuates superoxide production and decreases adhesion and migration of human cancerous cells. Biomed. Pharmacother. 2016, 84, 1972–1978. [Google Scholar] [CrossRef]
- Zhou, C.; Xie, Z.; Lei, Z.; Huang, Y.; Wei, G. Simultaneous identification and determination of flavonoids in Dendrobium officinale. Chem. Central J. 2018, 12, 40. [Google Scholar] [CrossRef] [Green Version]
- Dias, A.L.S.; Rozet, E.; Larondelle, Y.; Hubert, P.; Rogez, H.; Quetin-Leclercq, J. Development and validation of an UHPLC-LTQ-Orbitrap MS method for non-anthocyanin flavonoids quantification in Euterpe oleraceae juice. Anal. Bioanal. Chem. 2013, 405, 9235–9249. [Google Scholar] [CrossRef]
- Ho, K.-V.; Lei, Z.; Sumner, L.W.; Coggeshall, M.V.; Hsieh, H.-Y.; Stewart, G.C.; Lin, C.-H. Identifying Antibacterial Compounds in Black Walnuts (Juglans nigra) Using a Metabolomics Approach. Metabolites 2018, 8, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.K. Anti-inflammatory effects of eriodictyol in lipopolysaccharidestimulated raw 264.7 murine macrophages. Arch. Pharmacal Res. 2011, 34, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Mulabagal, V.; Calderón, A.I. Liquid chromatography/mass spectrometry based fingerprinting analysis and mass profiling of Euterpe oleraceae (açaí) dietary supplement raw materials. Food Chem. 2012, 134, 1156–1164. [Google Scholar] [CrossRef] [PubMed]
- Prior, R.L.; Cao, G.; Martin, A.; Sofic, E.; McEwen, J.; O’Brien, C.; Lischner, N.; Ehlenfeldt, M.; Kalt, W.; Krewer, A.G.; et al. Antioxidant Capacity As Influenced by Total Phenolic and Anthocyanin Content, Maturity, and Variety of Vaccinium Species. J. Agric. Food Chem. 1998, 46, 2686–2693. [Google Scholar] [CrossRef]
- de Camargo, A.C.; Regitano-D’Arce, M.A.B.; Biasoto, A.C.T.; Shahidi, F. Low Molecular Weight Phenolics of Grape Juice and Winemaking Byproducts: Antioxidant Activities and Inhibition of Oxidation of Human Low-Density Lipoprotein Cholesterol and DNA Strand Breakage. J. Agric. Food Chem. 2014, 62, 12159–12171. [Google Scholar] [CrossRef]
- Ayoub, M.; de Camargo, A.C.; Shahidi, F. Antioxidants and bioactivities of free, esterified and insoluble-bound phenolics from berry seed meals. Food Chem. 2016, 197, 221–232. [Google Scholar] [CrossRef]
- Butkevičiūtė, A.; Urbštaitė, R.; Liaudanskas, M.; Obelevičius, K.; Janulis, V. Phenolic Content and Antioxidant Activity in Fruit of the Genus Rosa L. Antioxidants 2022, 11, 912. [Google Scholar] [CrossRef]
- Kang, J.; Thakali, K.M.; Xie, C.; Kondo, M.; Tong, Y.; Ou, B.; Jensen, G.; Medina, M.B.; Schauss, A.G.; Wu, X. Bioactivities of açaí (Euterpe precatoria Mart.) fruit pulp, superior antioxidant and anti-inflammatory properties to Euterpe oleraceae Mart. Food Chem. 2012, 133, 671–677. [Google Scholar] [CrossRef]
- Batista, C.D.C.R.; de Oliveira, M.S.; Araújo, M.E.; Rodrigues, A.M.C.; Botelho, J.R.S.; da Silva Souza Filho, A.P.; Machado, N.; Carvalho Junior, R.N. Supercritical CO 2 extraction of açaí (Euterpe oleraceae) berry oil: Global yield, fatty acids, allelopathic activities, and determination of phenolic and anthocyanins total compounds in the residual pulp. J. Supercrit. Fluids 2016, 107, 364–369. [Google Scholar] [CrossRef]
- Sette, P.; Fernandez, A.; Soria, J.; Rodriguez, R.; Salvatori, D.; Mazza, G. Integral valorization of fruit waste from wine and cider industries. J. Clean. Prod. 2020, 242, 118486. [Google Scholar] [CrossRef]
- Kumar, M.; Dahuja, A.; Tiwari, S.; Punia, S.; Tak, Y.; Amarowicz, R.; Bhoite, A.G.; Singh, S.; Joshi, S.; Panesar, P.S.; et al. Recent trends in extraction of plant bioactives using green technologies: A review. Food Chem. 2021, 353, 129431. [Google Scholar] [CrossRef] [PubMed]
- Tremocoldi, M.A.; Rosalen, P.L.; Franchin, M.; Massarioli, A.P.; Denny, C.; Daiuto, E.R.; Paschoal, J.A.R.; Melo, P.S.; de Alencar, S.M. Exploration of avocado by-products as natural sources of bioactive compounds. PLoS ONE 2018, 13, e0192577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Moura, C.; dos Reis, A.S.; da Silva, L.D.; de Lima, V.A.; Oldoni, T.L.C.; Pereira, C.; Carpes, S.T. Optimization of phenolic compounds extraction with antioxidant activity from acai, blueberry and goji berry using response surface methodology. Emir. J. Food Agric. 2018, 180–189. [Google Scholar] [CrossRef] [Green Version]
- Silva, F.; De Miranda, D.; Carnier, M.; Maza, P.; Boldarine, V.; Rischiteli, A.S.; Avila, F.; Pontes, L.; Hachul, A.; Neto, N.; et al. Low dose of Juçara pulp (Euterpe edulis Mart.) minimizes the colon inflammatory milieu promoted by hypercaloric and hyperlipidic diet in mice. J. Funct. Foods 2021, 77, 104343. [Google Scholar] [CrossRef]
- Morais, D.R.; Rotta, E.M.; Sargi, S.C.; Schmidt, E.M.; Bonafe, E.G.; Eberlin, M.N.; Sawaya, A.C.; Visentainer, J.V. Antioxidant activity, phenolics and UPLC–ESI(–)–MS of extracts from different tropical fruits parts and processed peels. Food Res. Int. 2015, 77, 392–399. [Google Scholar] [CrossRef]
- Tiveron, A.P.; Melo, P.S.; Bergamaschi, K.B.; Vieira, T.M.F.D.S.; Regitano-D’Arce, M.A.B.; De Alencar, S.M. Antioxidant Activity of Brazilian Vegetables and Its Relation with Phenolic Composition. Int. J. Mol. Sci. 2012, 13, 8943–8957. [Google Scholar] [CrossRef] [Green Version]
- Infante, J.; Rosalen, P.L.; Lazarini, J.G.; Franchin, M.; De Alencar, S.M. Antioxidant and Anti-Inflammatory Activities of Unexplored Brazilian Native Fruits. PLoS ONE 2016, 11, e0152974. [Google Scholar] [CrossRef] [Green Version]
- Lazarini, J.G.; Franchin, M.; Soares, J.C.; Nani, B.D.; Massarioli, A.P.; De Alencar, S.M.; Rosalen, P.L. Anti-inflammatory and antioxidant potential, in vivo toxicity, and polyphenolic composition of Eugenia selloi B.D.Jacks. (pitangatuba), a Brazilian native fruit. PLoS ONE 2020, 15, e0234157. [Google Scholar] [CrossRef]
- Soares, J.C.; Rosalen, P.L.; Lazarini, J.G.; Massarioli, A.P.; da Silva, C.F.; Nani, B.D.; Franchin, M.; de Alencar, S.M. Comprehensive characterization of bioactive phenols from new Brazilian superfruits by LC-ESI-QTOF-MS, and their ROS and RNS scavenging effects and anti-inflammatory activity. Food Chem. 2019, 281, 178–188. [Google Scholar] [CrossRef]
- da Silveira, T.F.F.; de Souza, T.C.L.; Carvalho, A.V.; Ribeiro, A.B.; Kuhnle, G.G.; Godoy, H.T. White açaí juice (Euterpe oleraceae): Phenolic composition by LC-ESI-MS/MS, antioxidant capacity and inhibition effect on the formation of colorectal cancer related compounds. J. Funct. Foods 2017, 36, 215–223. [Google Scholar] [CrossRef]
- Rodrigues, E.; Mariutti, L.R.B.; Mercadante, A.Z. Carotenoids and Phenolic Compounds from Solanum sessiliflorum, an Unexploited Amazonian Fruit, and Their Scavenging Capacities against Reactive Oxygen and Nitrogen Species. J. Agric. Food Chem. 2013, 61, 3022–3029. [Google Scholar] [CrossRef] [PubMed]
- Sprenger, L.K.; Giese, E.G.; Dos Santos, J.N.; Molento, M.B. In vitro antibacterial effect of Euterpe oleraceae Mart. and Theobroma grandiflorum hydroalcoholic extracts. Arch. Vet. Sci. 2016, 21, 21–32. [Google Scholar] [CrossRef] [Green Version]
- Dias-Souza, M.V.; dos Santos, R.M.; Cerávolo, I.P.; Cosenza, G.; Marçal, P.H.F.; Figueiredo, F.J.B. Euterpe oleraceae pulp extract: Chemical analyses, antibiofilm activity against Staphylococcus aureus, cytotoxicity and interference on the activity of antimicrobial drugs. Microb. Pathog. 2018, 114, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Golovinskaia, O.; Wang, C.-K. Review of Functional and Pharmacological Activities of Berries. Molecules 2021, 26, 3904. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Tsao, R. Dietary polyphenols, oxidative stress and antioxidant and anti-inflammatory effects. Curr. Opin. Food Sci. 2016, 8, 33–42. [Google Scholar] [CrossRef]
- Xie, C.; Kang, J.; Li, Z.; Schauss, A.; Badger, T.M.; Nagarajan, S.; Wu, T.; Wu, X. The açaí flavonoid velutin is a potent anti-inflammatory agent: Blockade of LPS-mediated TNF-α and IL-6 production through inhibiting NF-κB activation and MAPK pathway. J. Nutr. Biochem. 2012, 23, 1184–1191. [Google Scholar] [CrossRef]
- Brito, C.; Stavroullakis, A.; Oliveira, T.; Prakki, A. Cytotoxicity and potential anti-inflammatory activity of velutin on RAW 264.7 cell line differentiation: Implications in periodontal bone loss. Arch. Oral Biol. 2017, 83, 348–356. [Google Scholar] [CrossRef]
- Daglia, M. Polyphenols as antimicrobial agents. Curr. Opin. Biotechnol. 2012, 23, 174–181. [Google Scholar] [CrossRef]
- Nile, S.H.; Park, S.W. Edible berries: Bioactive components and their effect on human health. Nutrition 2014, 30, 134–144. [Google Scholar] [CrossRef]
- Xue, J.; Davidson, P.M.; Zhong, Q. Thymol Nanoemulsified by Whey Protein-Maltodextrin Conjugates: The Enhanced Emulsifying Capacity and Antilisterial Properties in Milk by Propylene Glycol. J. Agric. Food Chem. 2013, 61, 12720–12726. [Google Scholar] [CrossRef]
- Ultee, A.; Bennik, M.H.J.; Moezelaar, R. The Phenolic Hydroxyl Group of Carvacrol Is Essential for Action against the Food-Borne Pathogen Bacillus cereus. Appl. Environ. Microbiol. 2002, 68, 1561–1568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freires, I.A.; Denny, C.; Benso, B.; De Alencar, S.M.; Rosalen, P.L. Antibacterial Activity of Essential Oils and Their Isolated Constituents against Cariogenic Bacteria: A Systematic Review. Molecules 2015, 20, 7329–7358. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Retention Time (min) | Molecular Formula | Exact Mass | Parent Ion (m/z) | Error (ppm) | Fragments (MS2) | |
|---|---|---|---|---|---|---|---|
| Anthocyanins [M+H]+ | Cyanidin 3-O-rutinoside | 11.4 | C27H30O15 | 594.1574 | 595.1652 | −1.346 | 287.0556 |
| Cyanidin 3-O-glucoside | 16.9 | C21H20O11 | 448.1018 | 449.1096 | −13.836 | 287.0548 | |
| Delphinidin 3-rutinoside | 19.0 | C27H30O16 | 610.1528 | 611.1606 | −0.656 | 303.0505 | |
| Pelargonidin 3-O-glucoside | 19.3 | C21H20O10 | 432.1058 | 433.1136 | 1.389 | 271.0607 | |
| Peonidin 3-O-rutinoside | 19.7 | C28H32O15 | 608.1736 | 609.1814 | −0.987 | 301.0725 | |
| Peonidin 3-O-glucoside | 23.1 | C22H22O11 | 462.1165 | 463.1243 | 0.649 | 301.0722 | |
| Malvidin 3-glucoside | 23.1 | C23H24O12 | 492.1268 | 493.1346 | −0.813 | 331.0822 | |
| Phenolic acids [M-H]− | Caffeoylquinic acid | 12.3 | C16H18O9 | 354.0968 | 353.089 | 5.083 | 191.0562; 179.034 |
| 4-Caffeoylshikimic acid | 15.3 | C16H16O8 | 336.085 | 335.0772 | 2.975 | 179.0347; 161.0319; 135.0464 | |
| Flavones [M-H]− | 6,8-di-C-hexosyl apigenin (vicenin-2) | 14.5 | C27H30O15 | 594.1581 | 593.1503 | 0.168 | 383.0763; 353.0662 |
| 6-C-glycosyl luteolin (isoorientin) | 16.9 | C21H20O11 | 448.1002 | 447.0924 | −1.785 | 447.0924; 357.0610; 327.0505 | |
| 6-C-glycosyl luteolin (isoorientin) | 17.4 | C21H20O11 | 448.1007 | 447.0929 | −0.669 | 447.0929; 357.0617; 327.0499 | |
| 6-C-glycosyl luteolin (isoorientin) | 17.6 | C21H20O11 | 448.0988 | 447.091 | −4.910 | 447.0910; 357.0588; 327.0488 | |
| 6-C-glycosyl apigenin (isovitexin) | 19.0 | C21H20O10 | 432.1053 | 431.0975 | −1.620 | 431.0975; 311.0557; 283.0602; 341.0676 | |
| Rhamnocitrin | 33.2 | C16H12O6 | 300.0633 | 299.0555 | 1.000 | 299; 284; 256; 227 | |
| Flavonoids [M-H]− | Taxifolin 3-O-glucoside | 15.9 | C21H22O12 | 466.1103 | 465.1025 | −1,716 | 285.0409; 151.0035; 125.0728 |
| Rutin | 19.0 | C27H30O16 | 610.1513 | 609.1435 | −2.786 | 255.0312; 271.0206. 301.0328 | |
| Flavanone [M-H]− | Eriodictyol 7-O-glucoside I | 18.4 | C21H22O11 | 450.1157 | 449.1079 | −0.666 | 269.0441; 259.0616 |
| Flavonols [M-H]− | Scoparin | 20.1 | C22H22O11 | 462.1160 | 461.1082 | 0.000 | 341.0667; 371.0776; 298.0483 |
| Scoparin | 20.4 | C22H22O11 | 462.1155 | 461.1077 | 0.000 | 341.0667; 371.0895; 298.0483 |
| Antioxidant Activity | |||||
|---|---|---|---|---|---|
| TPC (mg GAE∙g−1) | FRAP (µmol FS∙g−1) | ABTS•+ (µmol TE∙g−1) | ROO∙ (µmol TE∙g−1) | O2− (EC50 µg∙mL−1) | HOCl (EC50 µg∙mL−1) |
| 88.4 ± 0.4 | 986.0 ± 22.0 | 820.0 ± 36.4 | 975.7 ± 69.0 | 37.1 ± 1.9 | 4.2 ± 0.7 |
| Strain | Gram Staining | MIC/MBC (µg∙mL−1) |
|---|---|---|
| Staphylococcus aureus (ATCC 25923) | + | 1250/1250 |
| S. aureus (MRSA, ATCC 33591) | + | 2500/2500 |
| Pseudomonas aeruginosa (ATCC 27853) | − | 2500/2500 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Souza Silva, A.P.; de Camargo, A.C.; Lazarini, J.G.; Franchin, M.; Sardi, J.d.C.O.; Rosalen, P.L.; de Alencar, S.M. Phenolic Profile and the Antioxidant, Anti-Inflammatory, and Antimicrobial Properties of Açaí (Euterpe oleracea) Meal: A Prospective Study. Foods 2023, 12, 86. https://doi.org/10.3390/foods12010086
de Souza Silva AP, de Camargo AC, Lazarini JG, Franchin M, Sardi JdCO, Rosalen PL, de Alencar SM. Phenolic Profile and the Antioxidant, Anti-Inflammatory, and Antimicrobial Properties of Açaí (Euterpe oleracea) Meal: A Prospective Study. Foods. 2023; 12(1):86. https://doi.org/10.3390/foods12010086
Chicago/Turabian Stylede Souza Silva, Anna Paula, Adriano Costa de Camargo, Josy Goldoni Lazarini, Marcelo Franchin, Janaina de Cassia Orlandi Sardi, Pedro Luiz Rosalen, and Severino Matias de Alencar. 2023. "Phenolic Profile and the Antioxidant, Anti-Inflammatory, and Antimicrobial Properties of Açaí (Euterpe oleracea) Meal: A Prospective Study" Foods 12, no. 1: 86. https://doi.org/10.3390/foods12010086