
Effect of Pre-Fermentative Maceration and Fining Agents on Protein Stability, Macromolecular, and Phenolic Composition of Albariño White Wines: Comparative Efficiency of Chitosan, k-Carrageenan and Bentonite as Heat Stabilisers

 , and
, and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Winemaking Technology and Wine Composition
2.2. Fining Experiments
2.3. Macromolecular Material Isolation
2.4. Electrophoresis (Sodium Dodecyl Sulphate Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.5. Protein Quantification
2.6. Reversed Phase-High Performance Liquid Chromatography (RP-HPLC) Quantitative Protein Analysis
2.7. Protein Heat-Stability Test
2.8. Filterability Index
2.9. Quantification of Non-Flavonoids, Flavonoids and Total Phenols
2.10. Chromatic Characteristics and Colour (A420 nm)
2.11. High-Performance Liquid Chromatography (HPLC) Analysis of Catechin and Phenolic Acids
2.12. Polysaccharides Sugar Composition and Content
2.13. Statistical Analysis
3. Results and Discussion
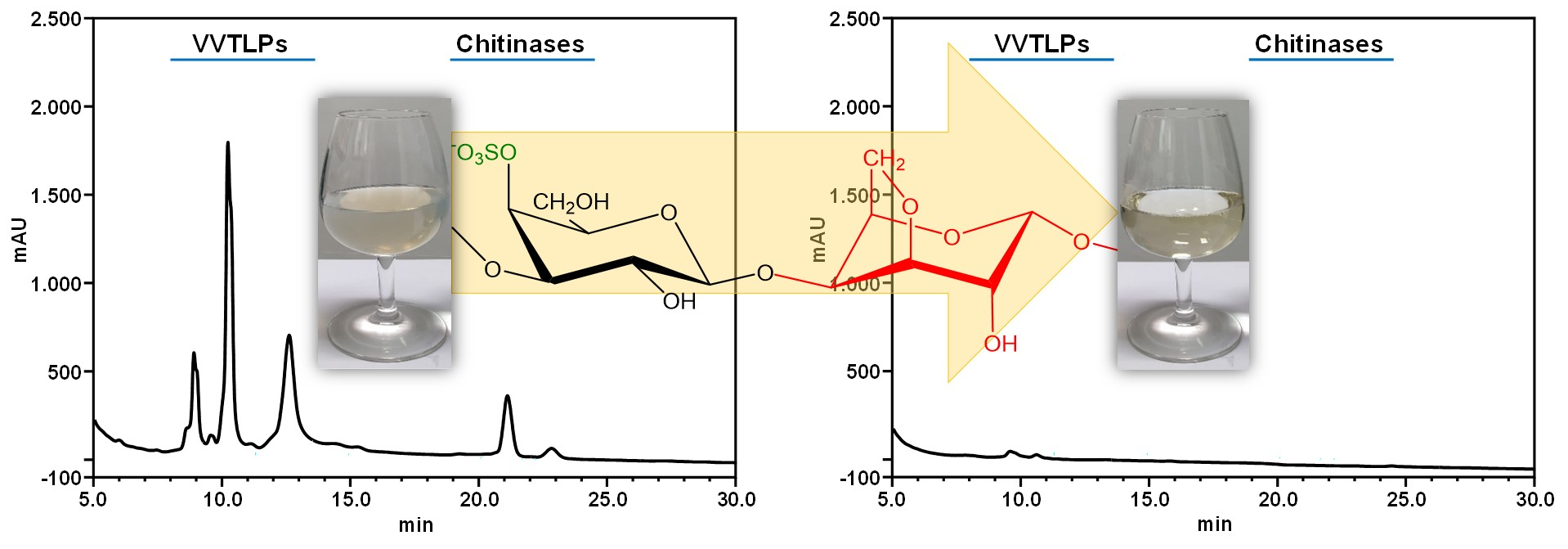
3.1. Protocol for the Isolation of the White Wine Macromolecular Components
3.2. Effect of the Skin Contact on the Albariño White Wine Protein Content, Heat Stability, and Efficiency of Protein Stabilisation by Fungal Chitosan, k-Carrageenan, and Bentonite
3.3. Effect of k-Carrageenan Addition on Wine Filterability
3.4. Effect of the Skin Contact on the Albariño White Wine Phenolic Composition and Chromatic Characteristics, and Effect of the Addition of Fungal Chitosan, k-Carrageenan and Bentonite
3.5. Effect of the Skin Contact on the Albariño Wine Polysaccharides Composition and Effect of the Addition of Fungal Chitosan, k-Carrageenan and Bentonite
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Darias-Martı́n, J.J.; Rodrı́guez, O.; Dı́az, E.; Lamuela-Raventós, R.M. Effect of skin contact on the antioxidant phenolics in white wine. Food Chem. 2000, 71, 483–487. [Google Scholar] [CrossRef]
- Selli, S. Effect of skin contact on the aroma composition of the musts of Vitis vinifera L. cv. Muscat of Bornova and Narince grown in Turkey. Food Chem. 2003, 81, 341–347. [Google Scholar] [CrossRef]
- Ramey, D.; Bertrand, A.; Ough, C.S.; Singleton, V.L.; Sanders, E. Effect of skin contact temperature on Chardonnay must and wine composition. Am. J. Enol. Vitic. 1986, 37, 99–106. [Google Scholar]
- Selli, S.; Canbas, A.; Cabaroglu, T.; Erten, H.; Günata, Z. Aroma components of cv. Muscat of Bornova wines and influence of skin contact treatment. Food Chem. 2006, 94, 319–326. [Google Scholar] [CrossRef]
- Tian, B.; Harrison, R.; Morton, J.; Jaspers, M. Influence of skin contact and different extractants on extraction of proteins and phenolic substances in Sauvignon Blanc grape skin. Aust. J. Grape Wine Res. 2020, 26, 180–186. [Google Scholar] [CrossRef]
- Ough, C.S. Substances extracted during skin contact with white musts. I. General wine composition and quality changes with contact time. Am. J. Enol. Vitic. 1969, 20, 93–100. [Google Scholar]
- Boulton, R. The general relationship between potassium, sodium and pH in grape juice and wine. Am. J. Enol. Vitic. 1980, 31, 182–186. [Google Scholar]
- Sokolowsky, M.; Rosenberger, A.; Fischer, U. Sensory impact of skin contact on white wines characterized by descriptive analysis, time–intensity analysis and temporal dominance of sensations analysis. Food Qual. Prefer. 2015, 39, 285–297. [Google Scholar] [CrossRef]
- Ferreira, R.B.; A Piçarra-Pereira, M.; Monteiro, S.; Loureiro, V.B.; Teixeira, A.R. The wine proteins. Trends Food Sci. Technol. 2001, 12, 230–239. [Google Scholar] [CrossRef]
- Robinson, S.P.; Davies, C. Molecular biology of grape berry ripening. Aust. J. Grape Wine Res. 2000, 6, 175–188. [Google Scholar] [CrossRef]
- Cosme, F.; Filipe-Ribeiro, L.; Nunes, F.M. Wine Stabilisation: An Overview of Defects and Treatments. In Chemistry and Biochemistry of Winemaking, Wine Stabilization and Aging [Working Title]; IntechOpen: London, UK, 2021. [Google Scholar]
- Sauvage, F.-X.; Bach, B.; Moutounet, M.; Vernhet, A. Proteins in white wines: Thermo-sensitivity and differential adsorbtion by bentonite. Food Chem. 2010, 118, 26–34. [Google Scholar] [CrossRef]
- Dambrouck, T.; Marchal, R.; Marchal-Delahaut, L.; Parmentier, M.; Maujean, A.A.; Jeandet, P. Immunodetection of Proteins from Grapes and Yeast in a White Wine. J. Agric. Food Chem. 2003, 51, 2727–2732. [Google Scholar] [CrossRef] [PubMed]
- Waters, E.; Alexander, G.; Muhlack, R.; Pocock, K.; Colby, C.; O’Neill, B.; Høj, P.; Jones, P. Preventing protein haze in bottled white wine. Aust. J. Grape Wine Res. 2005, 11, 215–225. [Google Scholar] [CrossRef]
- Marangon, M.; Stockdale, V.J.; Munro, P.; Trethewey, T.; Schulkin, A.; Holt, H.E.; Smith, P.A. Addition of Carrageenan at Different Stages of Winemaking for White Wine Protein Stabilization. J. Agric. Food Chem. 2013, 61, 6516–6524. [Google Scholar] [CrossRef]
- Waters, E.J.; Wallace, W.; Williams, P.J. Identification of heat-unstable wine proteins and their resistance to peptidases. J. Agric. Food Chem. 1992, 40, 1514–1519. [Google Scholar] [CrossRef]
- Marangon, M.; Sauvage, F.-X.; Waters, E.J.; Vernhet, A. Effects of Ionic Strength and Sulfate upon Thermal Aggregation of Grape Chitinases and Thaumatin-like Proteins in a Model System. J. Agric. Food Chem. 2011, 59, 2652–2662. [Google Scholar] [CrossRef]
- Cosme, F.; Fernandes, C.; Ribeiro, T.; Filipe-Ribeiro, L.; Nunes, F.M. White Wine Protein Instability: Mechanism, Quality Control and Technological Alternatives for Wine Stabilisation—An Overview. Beverages 2020, 6, 19. [Google Scholar] [CrossRef] [Green Version]
- Blade, W.; Boulton, R. Adsorption of protein by bentonite in a model wine solution. Am. J. Enol. Vitic. 1988, 39, 193–199. [Google Scholar]
- Achaerandio, I.; Pachova, V.; Güell, C.; López, F. Protein adsorption by bentonite in a white wine model solution: Efect of protein molecular weight and ethanol concentration. Am. J. Enol. Vitic. 2001, 52, 122–126. [Google Scholar]
- Lambri, M.; Dordoni, R.; Silva, A.; De Faveri, D.M. Comparing the impact of bentonite addition for both must clarification and wine fining on the chemical profile of wine from Chambave Muscat grapes. Int. J. Food Sci. Technol. 2011, 47, 1–12. [Google Scholar] [CrossRef]
- Waters, E.J.; Shirley, N.J.; Williams, P.J. Nuisance Proteins of Wine Are Grape Pathogenesis-Related Proteins. J. Agric. Food Chem. 1996, 44, 3–5. [Google Scholar] [CrossRef]
- Lambri, M.; Dordoni, R.; Silva, A.; De Faveri, D.M. Odoractive compound adsorption onto bentonite in a model white wine solution. Chem. Eng. Trans. 2013, 32, 1741–1746. [Google Scholar] [CrossRef]
- Dordoni, R.; Colangelo, D.; Giribaldi, M.; Giuffrida, M.G.; De Faveri, D.M.; Lambri, M. Effect of Bentonite Characteristics on Wine Proteins, Polyphenols, and Metals under Conditions of Different pH. Am. J. Enol. Vitic. 2015, 66, 518–530. [Google Scholar] [CrossRef]
- Vincenzi, S.; Panighel, A.; Gazzola, D.; Flamini, R.; Curioni, A. Study of Combined Effect of Proteins and Bentonite Fining on the Wine Aroma Loss. J. Agric. Food Chem. 2015, 63, 2314–2320. [Google Scholar] [CrossRef] [PubMed]
- Lambri, M.; Dordoni, R.; Silva, A.; Faveri, D.M. Efect of bentonite fining on odor-active compounds in two diferent white wine styles. Am. J. Enol. Vitic. 2010, 61, 225–233. [Google Scholar]
- Tattersall, D.B.; Pocock, K.F.; Hayasaka, Y.; Adams, K.; Van Heeswijck, R.; Waters, E.J.; Høj, P.B. Pathogenesis Related Proteins—Their Accumulation in Grapes during Berry Growth and Their Involvement in White Wine Heat Instability. Current Knowledge and Future Perspectives in Relation to Winemaking Practices. Mol. Biol. Biotechnol. Grapevine 2001, 183–201. [Google Scholar] [CrossRef]
- Majewski, P.; Barbalet, A.; Waters, E.J. $1 billion hidden cost of bentonite fining. Aust. N.Z. Grapegrow. Winemak. 2011, 569, 61–62. [Google Scholar]
- Vincenzi, S.; Polesani, M.; Curioni, A. Removal of specific protein compounds by chitin enhanced protein stability in white wine. Am. J. Enol. Vitic. 2005, 56, 246–254. [Google Scholar]
- Colangelo, D.; Torchio, F.; De Faveri, D.M.; Lambri, M. The use of chitosan as alternative to bentonite for wine fining: Effects on heat-stability, proteins, organic acids, colour, and volatile compounds in an aromatic white wine. Food Chem. 2018, 264, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Ratnayake, S.; Stockdale, V.; Grafton, S.; Munro, P.; Robinson, A.; Pearson, W.; McRae, J.; Bacic, A. Carrageenans as heat stabilisers of white wine. Aust. J. Grape Wine Res. 2019, 25, 439–450. [Google Scholar] [CrossRef]
- Ribeiro, T.; Fernandes, C.; Nunes, F.; Filipe-Ribeiro, L.; Cosme, F. Influence of the structural features of commercial mannoproteins in white wine protein stabilization and chemical and sensory properties. Food Chem. 2014, 159, 47–54. [Google Scholar] [CrossRef]
- Gonzalez-Ramos, D.; Cebollero, E.; Gonzalez, R. A Recombinant Saccharomyces cerevisiae Strain Overproducing Mannoproteins Stabilizes Wine against Protein Haze. Appl. Environ. Microbiol. 2008, 74, 5533–5540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Union (EU). Commission regulation (EU) 53/2011 of 21 January 2011. Available online: https://eurlex.euro-pa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2011:019:0001:0006:EN:PDF (accessed on 18 December 2020).
- OIV. International Code of Oenological Practices. International Organisation of Vine and Wine. 2019. Available online: http://www.oiv.int/fr/normes-et-documents-techniques (accessed on 20 December 2020).
- Bowman, S.M.; Free, S.J. The structure and synthesis of the fungal cell wall. BioEssays 2006, 28, 799–808. [Google Scholar] [CrossRef]
- Hamed, I.; Özogul, F.; Regenstein, J.M. Industrial applications of crustacean by-products (chitin, chitosan, and chitooligosaccharides): A review. Trends Food Sci. Technol. 2016, 48, 40–50. [Google Scholar] [CrossRef]
- Vincenzi, S.; Mosconi, S.; Zoccatelli, G.; Pellegrina, C.D.; Veneri, G.; Chignola, R.; Peruo, A.; Curioni, A.; Rizzi, C. Development of a new procedure for protein recovery and quantification in wine. Am. J. Enol. Vitic. 2005, 56, 182–187. [Google Scholar]
- Cabello-Pasini, A.; Victoria-Cota, N.; Macias-Carranza, V.; Hernandez-Garibay, E.; Muñiz-Salazar, R. Clarification of wines using polysaccharides extracted from seaweeds. Am. J. Enol. Vitic. 2005, 56, 52–59. [Google Scholar]
- Marangon, M.; Lucchetta, M.; Duan, D.; Stockdale, V.; Hart, A.; Rogers, P.; Waters, E. Protein removal from a Chardonnay juice by addition of carrageenan and pectin. Aust. J. Grape Wine Res. 2012, 18, 194–202. [Google Scholar] [CrossRef]
- Vidal, S.; Williams, P.; Doco, T.; Moutounet, M.; Pellerin, P. The polysaccharides of red wine: Total fractionation and characterization. Carbohydr. Polym. 2003, 54, 439–447. [Google Scholar] [CrossRef]
- Gawel, R.; Smith, P.A.; Waters, E.J. Influence of polysaccharides on the taste and mouthfeel of white wine. Aust. J. Grape Wine Res. 2016, 22, 350–357. [Google Scholar] [CrossRef]
- Jones, P.; Gawel, R.; Francis, I.; Waters, E. The influence of interactions between major white wine components on the aroma, flavour and texture of model white wine. Food Qual. Prefer. 2008, 19, 596–607. [Google Scholar] [CrossRef]
- Martínez-Lapuente, L.; Guadalupe, Z.; Ayestarán, B. Properties of Wine Polysaccharides. In Pectins-Extraction, Purification, Characterization and Applications; InTech: London, UK, 2020. [Google Scholar]
- Waters, E.J.; Pellerin, P.; Brillouet, J.-M. A Saccharomyces mannoprotein that protects wine from protein haze. Carbohydr. Polym. 1994, 23, 185–191. [Google Scholar] [CrossRef]
- Moine-Ledoux, V.; Dubourdieu, D. An invertase fragment responsible for improving the protein stability ofdry white wines. J. Sci. Food Agric. 1999, 79, 537–543. [Google Scholar] [CrossRef]
- Gawel, R.; Day, M.; Van Sluyter, S.C.; Holt, H.; Waters, E.J.; Smith, P.A. White Wine Taste and Mouthfeel As Affected by Juice Extraction and Processing. J. Agric. Food Chem. 2014, 62, 10008–10014. [Google Scholar] [CrossRef]
- Jaeckels, N.; Meier, M.; Dietrich, H.; Will, F.; Decker, H.; Fronk, P. Influence of polysaccharides on wine protein aggregation. Food Chem. 2016, 200, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Read, S.; Northcote, D. Minimization of variation in the response to different proteins of the Coomassie blue G dye-binding assay for protein. Anal. Biochem. 1981, 116, 53–64. [Google Scholar] [CrossRef]
- Pocock, K.F.; Hayasaka, Y.; McCarthy, A.M.G.; Waters, E.J. Thaumatin-like Proteins and Chitinases, the Haze-Forming Proteins of Wine, Accumulate during Ripening of Grape (Vitis vinifera) Berries and Drought Stress Does Not Affect the Final Levels per Berry at Maturity. J. Agric. Food Chem. 2000, 48, 1637–1643. [Google Scholar] [CrossRef] [PubMed]
- Salazar, F.N.; López, F.; Chiffelle, I.; Lopez, R.; Peña-Neira, Á. Evaluation of pathogenesis-related protein content and protein instability of seven white grape (Vitis vinifera L.) clones from Casablanca Valley, Chile. Eur. Food Res. Technol. 2012, 234, 509–515. [Google Scholar] [CrossRef]
- Dubourdieu, D.; Serrano, M.; Vannier, A.C.; Ribéreau-Gayon, P. Étude comparée des tests de stabilité protéique. Conn. Vigne Vin 1988, 22, 261–273. [Google Scholar] [CrossRef] [Green Version]
- Descout, J.J.; Bordier, J.L.; Laurenty, J.; Guimberteau, G. Contribution a l’etude des phenomenes de colmatage lors de la filtration des vins sur filtre ecran. Conn. Vigne Vin 1976, 10, 93–123. [Google Scholar] [CrossRef]
- Kramling, T.; Singleton, V.L. An estimate of the nonflavonoid phenols in wines. Am. J. Enol. Vitic. 1969, 20, 86–92. [Google Scholar]
- Ribéreau-Gayon, P.; Peynaud, E.; Sudraud, P. Traité d’OEnologie. In Science et Techniques du Vin; Tome 4; Dunod: Paris, France, 1982; p. 643. [Google Scholar]
- OIV. Organisation International de la Vigne et du Vin Récueil deMéthodes Internationales d’Analyse des Vins et des Moûts. Edition O/methodes-danalyse. Available online: https://www.oiv.int/fr/normes-et-documents-techniques/methodes-danalyse/recueil-des-methodes-internationales-danalyse-des-vins-et-des-mouts-2-vol (accessed on 20 December 2020).
- Spagna, G.; Pifferi, P.G.; Rangoni, C.; Mattivi, F.; Nicolini, G.; Palmonari, R. The stabilization of white wines by adsorption of phenolic compounds on chitin and chitosan. Food Res. Int. 1996, 29, 241–248. [Google Scholar] [CrossRef]
- Guise, R.; Filipe-Ribeiro, L.; Nascimento, D.; Bessa, O.; Nunes, F.; Cosme, F. Comparison between different types of carboxylmethylcellulose and other oenological additives used for white wine tartaric stabilization. Food Chem. 2014, 156, 250–257. [Google Scholar] [CrossRef]
- Filipe-Ribeiro, L.; Milheiro, J.; Matos, C.C.; Cosme, F.; Nunes, F.M. Reduction of 4-ethylphenol and 4-ethylguaiacol in red wine by activated carbons with different physicochemical characteristics: Impact on wine quality. Food Chem. 2017, 229, 242–251. [Google Scholar] [CrossRef]
- Filipe-Ribeiro, L.; Milheiro, J.; Matos, C.C.; Cosme, F.; Nunes, F.M. Data on changes in red wine phenolic compounds, headspace aroma compounds and sensory profile after treatment of red wines with activated carbons with different physicochemical characteristics. Data Brief. 2017, 12, 188–202. [Google Scholar] [CrossRef] [PubMed]
- Fraga, S.M.; Nunes, F.M. Agaricus bisporus By-Products as a Source of Chitin-Glucan Complex Enriched Dietary Fibre with Potential Bioactivity. Appl. Sci. 2020, 10, 2232. [Google Scholar] [CrossRef] [Green Version]
- Batista, L.; Monteiro, S.; Loureiro, V.B.; Teixeira, A.R.; Ferreira, R.B. The complexity of protein haze formation in wines. Food Chem. 2009, 112, 169–177. [Google Scholar] [CrossRef]
- Mesquita, P.R.; Piçarra-Pereira, M.A.; Monteiro, S.; Loureiro, V.B.; Teixeira, A.R.; Ferreira, R.B. Effect of wine composition on protein stability. Am. J. Enol. Vitic. 2001, 52, 324–330. [Google Scholar]
- Pellerin, P.; Cabanis, J.-C. Les Glucides. In Œnologie. Fondements Scientifiques et Technologiques; Flanzy, C., Ed.; Lavoisier- Tec & Doc: Paris, France, 1998. [Google Scholar]
- Doco, T.; Williams, P.; Moutounet, M.; Pellerin, P. Les polysaccharides du vin. Bulletin l’O.I.V. 2000, 73, 785–792. [Google Scholar]
- Tian, B.; Harrison, R.; Morton, J.D.; Jaspers, M.V.; Hodge, S.; Grose, C.; Trought, M.C. Extraction of Pathogenesis-Related Proteins and Phenolics in Sauvignon Blanc as Affected by Grape Harvesting and Processing Conditions. Molecules 2017, 22, 1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marangon, M.; Van Sluyter, S.C.; Haynes, P.A.; Waters, E.J. Grape and Wine Proteins: Their Fractionation by Hydrophobic Interaction Chromatography and Identification by Chromatographic and Proteomic Analysis. J. Agric. Food Chem. 2009, 57, 4415–4425. [Google Scholar] [CrossRef] [PubMed]
- Sęczyk, Ł.; Świeca, M.; Kapusta, I.; Gawlik-Dziki, U. Protein–Phenolic Interactions as a Factor Affecting the Physicochemical Properties of White Bean Proteins. Molecules 2019, 24, 408. [Google Scholar] [CrossRef] [Green Version]
- Gabrielli, M.; Fracassetti, D.; Tirelli, A. Release of phenolic compounds from cork stoppers and its effect on protein-haze. Food Control. 2016, 62, 330–336. [Google Scholar] [CrossRef]
- Falconer, R.J.; Marangon, M.; Van Sluyter, S.C.; Neilson, K.A.; Chan, C.; Waters, E.J. Thermal Stability of Thaumatin-Like Protein, Chitinase, and Invertase Isolated from Sauvignon blanc and Semillon Juice and Their Role in Haze Formation in Wine. J. Agric. Food Chem. 2010, 58, 975–980. [Google Scholar] [CrossRef]
- Bayly, F.C.; Berg, H.W. Grape and wine proteins of white wine varietals. Am. J. Enol. Vitic. 1967, 24, 18–32. [Google Scholar]
- Moretti, R.H.; Berg, H.W. Variability among wines to protein clouding. Am. J. Enol. Vitic. 1965, 16, 69–78. [Google Scholar]
- Waters, E.J.; Hayasaka, Y.; Tattersall, D.B.; Adams, K.S.; Williams, P.J. Sequence Analysis of Grape (Vitis vinifera) Berry Chitinases That Cause Haze Formation in Wines. J. Agric. Food Chem. 1998, 46, 4950–4957. [Google Scholar] [CrossRef]
- Esteruelas, M.; Poinsaut, P.; Sieczkowski, N.; Manteau, S.; Fort, M.; Canals, J.; Zamora, F. Characterization of natural haze protein in sauvignon white wine. Food Chem. 2009, 113, 28–35. [Google Scholar] [CrossRef]
- Dufrechou, M.; Poncet-Legrand, C.; Sauvage, F.-X.; Vernhet, A. Stability of White Wine Proteins: Combined Effect of pH, Ionic Strength, and Temperature on Their Aggregation. J. Agric. Food Chem. 2012, 60, 1308–1319. [Google Scholar] [CrossRef]
- Sun, B.; Spranger, I.; Roque-Do-Vale, F.; Leandro, C.; Belchior, P. Effect of Different Winemaking Technologies on Phenolic Composition in Tinta Miúda Red Wines. J. Agric. Food Chem. 2001, 49, 5809–5816. [Google Scholar] [CrossRef]
- Guadalupe, Z.; Martínez, L.; Ayestarán, B. Yeast mannoproteins in red winemaking: Effect on polysaccharide, polyphenolic, and color composition. Am. J. Enol. Vitic. 2010, 61, 191–200. [Google Scholar]
- De Vries, R.P.; Visser, J. Aspergillus Enzymes Involved in Degradation of Plant Cell Wall Polysaccharides. Microbiol. Mol. Biol. Rev. 2001, 65, 497–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guadalupe, Z.; Ayestarán, B.; Williams, P.; Doco, T. Determination of Must and Wine Polysaccharides by Gas. Chromatography-Mass Spectrometry (GC-MS) and Size-Exclusion Chromatography (SEC); Springer International Publishing: Berlin/Heidelberg, Germany, 2014; pp. 1–28. [Google Scholar]
- Thompson, J.E.; Fry, S.C. Evidence for covalent linkage between xyloglucan and acidic pectins in suspension-cultured rose cells. Planta 2000, 211, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Doco, T.; Williams, P.; Pauly, M.; O’Neill, M.A.; Pellerin, P. Polysaccharides from grape berry cell walls. Part II. Structural characterization of the xyloglucan polysaccharides. Carbohydr. Polym. 2003, 53, 253–261. [Google Scholar] [CrossRef]
- Vernhet, A.; Pellerin, P.; Prieur, C.; Osmianski, J.; Moutounet, M. Charge properties of some grape and wine polysaccharide and polyphenolic fractions. Am. J. Enol. Vitic. 1996, 47, 25–30. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Albariño Monovarietal White Wine | With Pre-Fermentative Skin Maceration (+PFSM) | Without Pre-Fermentative Skin Maceration (−PFSM) |
|---|---|---|
| Alcohol content (% v/v) | 13.6 | 14.3 |
| Density at 20 °C (g/cm3) | 0.9888 | 0.9883 |
| Titratable acidity (expressed as g/L tartaric acid) | 6.2 | 7.3 |
| pH | 3.81 | 3.52 |
| Volatile acidity (expressed as g/L acetic acid) | 0.42 | 0.45 |
| Protein instability (Heat-test, ΔNTU) | 26.6 | 22.2 |
| Total Phenols | Flavonoid Phenols | Non-Flavonoid Phenols | Abs 420 nm | L* | a* | b* | ΔE* | |
|---|---|---|---|---|---|---|---|---|
| Without pre-fermentative skin contact | ||||||||
| Control | 39 ± 1 | 22 ± 2 a | 17 ± 1 a | 0.079 ± 0.006 a | 98.2 ± 0.5 a | 0.14 ± 0.08 a | 4.89 ± 0.08 | |
| Fungal Chitosan | 35 ± 0 | 21 ± 0 a,c | 15 ± 1 a | 0.076 ± 0.014 a,b | 99.1 ± 0.4 a,b | 0.12 ± 0.08 a | 3.84 ± 0.96 | 1.43 ± 1.13 |
| k-Carrageenan | 38 ± 1 | 22 ± 0 a | 16 ± 0 a | 0.079 ± 0.002 a | 98.1 ± 0.2 a | 0.16 ± 0.07 a | 4.78 ± 0.03 | 0.13 ± 0.07 |
| Sodium Bentonite | 38 ± 0 | 17 ± 1 b,c | 21 ± 0 b | 0.075 ± 0.002 a,b | 99.3 ± 0.4 a,b | −0.40 ± 0.04 b | 4.00 ± 0.28 | 1.62 ± 0.84 |
| Calcium Bentonite | 39 ± 2 | 24 ± 1 a | 15 ± 1 a | 0.048 ± 0.003 b | 99.9 ± 0.1 b | −0.46 ± 0.02 b | 3.79 ± 0.08 | 1.07 ± 0.30 |
| ANOVA | 0.0752 | 0.0157 | 0.0037 | 0.0279 | 0.0152 | 0.0003 | 0.1450 | 0.3059 |
| With pre-fermentative skin contact | ||||||||
| Control | 59 ± 0 a | 32 ± 0 a | 28 ± 0 a | 0.117 ± 0.008 a | 97.8 ± 0.5 | 0.77 ± 0.02 | 6.78 ± 0.17 a | |
| Fungal Chitosan | 52 ± 0 b,c | 33 ± 1 a | 19 ± 1 b | 0.084 ± 0.008 b | 98.1 ± 0.2 | 0.70 ± 0.12 | 5.32 ± 0.49 b | 1.52 ± 0.31 |
| k-Carrageenan | 54 ± 0 b | 33 ± 1 a | 21 ± 0 b,c | 0.116 ± 0.002 a | 97.6 ± 0.1 | 0.81 ± 0.04 | 6.68 ± 0.11 a | 0.19 ± 0.11 |
| Sodium Bentonite | 48 ± 0 c | 26 ± 1 b,c | 22 ± 0 c | 0.107 ± 0.010 a,b | 97.3 ± 0.7 | 1.03 ± 0.14 | 6.49 ± 0.15 a | 0.74 ± 0.48 |
| Calcium Bentonite | 52 ± 0 b,c | 29 ± 1 a,c | 22 ± 0 c | 0.102 ± 0.005 a,b | 97.6 ± 0.2 | 0.99 ± 0.07 | 6.42 ± 0.23 a | 0.59 ± 0.27 |
| ANOVA | 0.0001 | 0.0023 | 0.0002 | 0.0280 | 0.4670 | 0.0534 | 0.0143 | 0.0575 |
| t-test | 0.0016 | 0.0206 | 0.0051 | 0.0329 | 0.508 | 0.0085 | 0.0049 | |
| Gallic Acid | Catechin | t-Caftaric Acid | Coutaric Acid | Caffeic Acid | p-Coumaric Acid | Ferulic Acid | Ethyl Ester of Caffeate | Ethyl Ester of p-Coumarate | Total | |
|---|---|---|---|---|---|---|---|---|---|---|
| Without pre-fermentative skin maceration | ||||||||||
| Control | 50.0 ± 0.9 | 7.06 ± 1.48 | 23.8 ± 0.3 | 21.7 ± 1.6 | 6.48 ± 0.74 | 1.84 ± 0.80 | 0.06 ± 0.01 | 0.12 ± 0.14 | 0.12 ± 0.00 | 111.4 ± 3.5 a |
| Fungal Chitosan | 46.0 ± 0.1 | 6.78 ± 0.34 | 23.6 ± 0.6 | 15.5 ± 1.8 | 5.89 ± 0.09 | 1.07 ± 0.23 | 0.04 ± 0.02 | 0.03 ± 0.00 | 0.10 ± 0.02 | 99.1 ± 1.1 b |
| k-Carrageenan | 48.3 ± 1.6 | 6.06 ± 0.06 | 23.3 ± 0.3 | 16.2 ± 2.8 | 6.14 ± 0.13 | 1.66 ± 0.22 | 0.05 ± 0.01 | 0.06 ± 0.04 | 0.08 ± 0.06 | 102.0 ± 0.7 b |
| Sodium Bentonite | 49.7 ± 0.2 | 5.58 ± 0.61 | 23.7 ± 1.1 | 14.9 ± 0.1 | 6.53 ± 0.05 | 1.33 ± 0.04 | 0.04 ± 0.01 | 0.03 ± 0.00 | 0.05 ± 0.04 | 101.9 ± 0.6 b |
| Calcium Bentonite | 52.7 ± 6.9 | 3.70 ± 0.99 | 22..9 ± 0.5 | 15.6 ± 0.4 | 6.14 ± 0.41 | 1.19 ± 0.23 | 0.03 ± 0.01 | 0.06 ± 0.06 | 0.06 ± 0.01 | 102.5 ± 2.6 b |
| ANOVA | 0.4201 | 0.0573 | 0.6431 | 0.0428 | 0.5037 | 0.3793 | 0.3008 | 0.7122 | 0.3445 | 0.0122 |
| With pre-fermentative skin maceration | ||||||||||
| Control | 57.4 ± 0.5 a | 22.3 ± 0.6 | 25.0 ± 0.8 | 22.7 ± 4.4 | 6.54 ± 0.80 | 8.12 ± 0.43 | 0.15 ± 0.02 | 0.37 ± 0.01 | 0.48 ± 0.02 | 143.3 ± 2.5 a |
| Fungal Chitosan | 31.5 ± 4.9 b | 22.4 ± 0.2 | 26.0 ± 3.5 | 20.4 ± 5.6 | 6.11 ± 0.44 | 5.29 ± 1.03 | 0.06 ± 0.01 | 0.32 ± 0.01 | 0.44 ± 0.07 | 112.9 ± 1.3 c |
| k-Carrageenan | 50.5 ± 4.6 a | 22.7 ± 1.9 | 25.9 ± 3.2 | 19.7 ± 0.2 | 5.86 ± 0.15 | 5.48 ± 3.58 | 0.07 ± 0.06 | 0.36 ± 0.07 | 0.47 ± 0.06 | 129.6 ± 3.4 b,c |
| Sodium Bentonite | 47.1 ± 2.1 a | 20.7 ± 0.2 | 25.1 ± 0.5 | 18.5 ± 2.1 | 5.83 ± 0.08 | 6.88 ± 0.43 | 0.13 ± 0.08 | 0.36 ± 0.02 | 0.13 ± 0.12 | 124.7 ± 0.4 b |
| Calcium Bentonite | 35.3 ± 0.9 b | 20.4 ± 0.6 | 24.4 ± 0.6 | 19.6 ± 1.1 | 5.70 ± 0.03 | 4.93 ± 0.89 | 0.13 ± 0.07 | 0.29 ± 0.09 | 0.40 ± 0.31 | 118.4 ± 5.3 b |
| ANOVA | 0.0021 | 0.1639 | 0.1883 | 0.7753 | 0.3888 | 0.4206 | 0.4655 | 0.5520 | 0.2718 | 0.0012 |
| t-test | 0.0095 | 0.0054 | 0.1854 | 0.7911 | 0.9450 | 0.0103 | 0.0295 | 0.1280 | 0.0015 | 0.0090 |
| Fuc | Rha | Ara | GlcN | Gal | Glc | Xyl | Man | GalA | GlcA | Total | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Without pre-fermentative skin contact | |||||||||||
| Control | 0.25 ± 0.03 a | 2.79 ± 0.15 a | 2.46 ± 0.10 a | 0.91 ± 0.17 a | 9.17 ± 0.55 a | 2.04 ± 0.06 a | n.d. | 27.8 ± 0.1 a | 12.4 ± 0.2 a | 3.73 ± 0.33 a | 61.6 ± 1.0 a |
| Fungal Chitosan | n.d. c | 0.59 ± 0.13 c | 1.00 ± 0.06 b | 0.66 ± 0.17 a,b | 5.25 ± 0.80 b | 1.87 ± 0.44 a,c | n.d. | 13.4 ± 1.6 b | 1.28 ± 0.53 b | 0.90 ± 0.01 b | 29.0 ± 1.0 c |
| k-Carrageenan | 0.25 ± 0.02 a | 2.79 ± 0.15 a | 2.72 ± 0.33 a | 1.06 ± 0.09 a | 13.3 ± 1.2 c | 3.31 ± 0.39 b | n.d. | 24.8 ± 1.9 a | 10.4 ± 2.3 a | 3.93 ± 0.12 a | 65.6 ± 2.2 a |
| Sodium Bentonite | 0.11 ± 0.01 b | 1.33 ± 0.11 b | 1.38 ± 0.21 b | 0.35 ± 0.05 b | 5.40 ± 0.85 b | 0.89 ± 0.21 c | n.d. | 11.9 ± 1.5 b | 2.18 ± 0.91 b | 1.86 ± 0.06 c | 25.4 ± 3.8 c |
| Calcium Bentonite | 0.17 ± 0.04 a,b | 2.36 ± 0.18 a | 2.64 ± 0.19 a | 0.70 ± 0.06 a,b | 10.4 ± 0.3 a,c | 1.52 ± 0.13 a,c | n.d. | 23.5 ± 1.3 a | 4.83 ± 0.11 b | 3.50 ± 0.25 a | 49.6 ± 2.0 b |
| ANOVA | 0.0007 | <0.0001 | 0.0010 | 0.0129 | 0.0007 | 0.0031 | - | 0.0003 | 0.0006 | <0.0001 | <0.0001 |
| With pre-fermentative skin contact | |||||||||||
| Control | 0.36 ± 0.02 a | 5.68 ± 0.01 a | 5.30 ± 0.15 b | 0.87 ± 0.06 a | 11.7 ± 0.5 a | 2.62 ± 0.55 a | 2.75 ± 0.07 a | 23.2 ± 1.4 a | 15.9 ± 1.0 a | 10.5 ± 1.5 a | 78.8 ± 2.2 a |
| Fungal Chitosan | 0.02 ± 0.01 c | 1.50 ± 0.02 b | 2.25 ± 0.24 c | 0.44 ± 0.07 b | 5.57 ± 0.84 b | 1.97 ± 0.15 a | 0.97 ± 0.16 b | 9.79 ± 1.91 b | 3.02 ± 1.07 b | 1.86 ± 0.22 b | 27.4 ± 2.6 b |
| k-Carrageenan | 0.38 ± 0.02 a | 5.08 ± 0.25 a | 6.34 ± 0.31 a | 0.84 ± 0.02 a | 19.4 ± 0.3 c | 4.93 ± 1.35 b | 3.92 ± 0.48 c | 28.4 ± 1.0 c | 16.0 ± 3.0 a | 10.2 ± 1.5 a | 95.8 ± 1.3 c |
| Sodium Bentonite | 0.27 ± 0.02 b | 4.39 ± 0.29 a | 4.44 ± 0.06 b | 0.46 ± 0.02 b | 10.8 ± 0.5 a | 1.55 ± 0.05 a | 1.57 ± 0.08 b | 15.3 ± 0.4 d | 12.3 ± 0.2 a | 6.56 ± 0.42 a,c | 57.6 ± 0.9 d |
| Calcium Bentonite | 0.30 ± 0.03 a,b | 5.92 ± 0.96 a | 5.23 ± 0.30 b | 0.47 ± 0.08 b | 13.5 ± 0.04 a | 1.86 ± 0.31 a | 2.20 ± 0.05 a | 18.9 ± 0.1 a,d | 11.7 ± 1.1 a | 5.88 ± 0.70 b,c | 65.9 ± 1.9 e |
| ANOVA | <0.0001 | 0.0011 | <0.0001 | 0.0010 | <0.0001 | 0.0073 | 0.0004 | <0.0001 | 0.0021 | 0.0018 | <0.0001 |
| t-test | 0.0497 | 0.0014 | 0.0020 | 0.7834 | 0.0406 | 0.2764 | 0.0001 | 0.0435 | 0.0399 | 0.0248 | 0.0097 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arenas, I.; Ribeiro, M.; Filipe-Ribeiro, L.; Vilamarim, R.; Costa, E.; Siopa, J.; Cosme, F.; Nunes, F.M. Effect of Pre-Fermentative Maceration and Fining Agents on Protein Stability, Macromolecular, and Phenolic Composition of Albariño White Wines: Comparative Efficiency of Chitosan, k-Carrageenan and Bentonite as Heat Stabilisers. Foods 2021, 10, 608. https://doi.org/10.3390/foods10030608
Arenas I, Ribeiro M, Filipe-Ribeiro L, Vilamarim R, Costa E, Siopa J, Cosme F, Nunes FM. Effect of Pre-Fermentative Maceration and Fining Agents on Protein Stability, Macromolecular, and Phenolic Composition of Albariño White Wines: Comparative Efficiency of Chitosan, k-Carrageenan and Bentonite as Heat Stabilisers. Foods. 2021; 10(3):608. https://doi.org/10.3390/foods10030608
Chicago/Turabian StyleArenas, Inma, Miguel Ribeiro, Luís Filipe-Ribeiro, Rafael Vilamarim, Elisa Costa, João Siopa, Fernanda Cosme, and Fernando M. Nunes. 2021. "Effect of Pre-Fermentative Maceration and Fining Agents on Protein Stability, Macromolecular, and Phenolic Composition of Albariño White Wines: Comparative Efficiency of Chitosan, k-Carrageenan and Bentonite as Heat Stabilisers" Foods 10, no. 3: 608. https://doi.org/10.3390/foods10030608