Towards Microbial Food Safety of Sprouts: Photodynamic Decontamination of Seeds



1
Laboratory of Photodynamic Inactivation of Microorganisms, Department of Biosciences and Medical Biology, Paris Lodron University of Salzburg, Hellbrunnerstr. 34, 5020 Salzburg, Austria
2
Suncor AgroScience, 2489 North Sheridan Way, Mississauga, ON L5K1A8, Canada
*
Author to whom correspondence should be addressed.
Photonics 2023, 10(3), 239; https://doi.org/10.3390/photonics10030239
Submission received: 30 January 2023
/
Revised: 16 February 2023
/
Accepted: 18 February 2023
/
Published: 22 February 2023
(This article belongs to the Special Issue Light-Based Technologies and Spectroscopic Techniques for Photo-Sensing and Photoinactivation of Microorganisms, Virus, and Cancer Cells)
Abstract
:The climate crisis is one of the biggest challenges for humanity in the 21st century. Production and consumption of meat contributes to global warming by causing emissions of climate-relevant gases. Freshly grown sprouts are part of an alternative, as they are less polluting but still a nutritious food. However, warm humid sprouting conditions may cause pathogenic microorganisms to thrive. Decontamination methods for raw sprouts are therefore relevant. Photodynamic Inactivation (PDI) is a novel approach that uses photoactivatable molecules (photosensitisers, PS) and visible or near-infrared light to produce reactive oxygen species (ROS). These ROS kill microorganisms by oxidative processes. Here, we test the application of PDI based on sodium-magnesium-chlorophyllin (Chl, approved as food additive E140) for photo-decontamination of mung bean, radish, and buckwheat seeds. Seeds were contaminated with Listeria innocua, serving as a model system for Listeria monocytogenes, subjected to PDI using an LED array with 395 nm and tested for remaining bacterial contamination by CFU counting. PDI based on 100 µM Chl reduces the bacterial load of mung bean and radish seeds by 99.9% (radiant exposure 56.4 J/cm2 and 28.2 J/cm2, respectively), and of buckwheat seeds by <90% reduction after illumination with 28.2 J/cm2. Neither weight nor the germination rates of seeds are affected by PDI. Interestingly, the effect of PDI on seeds is partially maintained on stored sprouts after germination: The bacterial load on mung bean sprouts is reduced by more than 99.9% after phototreatment of seeds with 100 µM Chl and illumination at 56.4 J/cm2. In conclusion, we suggest PDI based on Chl as an effective and biocompatible method for the decontamination of seeds and sprouts for human consumption from Listeria.
Keywords:
photodynamic inactivation; seeds; sprouts; chlorophyllin; Listeria innocua; shelf life; decontamination; food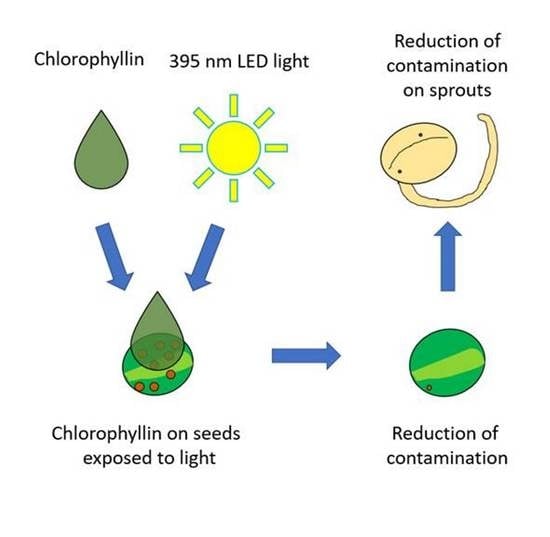
1. Introduction
The ever-worsening climate and biodiversity crisis pose the greatest challenges to humanity in the 21st century. Agriculture is responsible for about one-third of global greenhouse gas emissions [1], with the production of meat being the main driver [2]. A shift to more plant-based diets is helpful to minimize the impact of agriculture on the climate, but replacing nutritious meat is not always easy [3]. Freshly grown sprouts offer an alternative to meat regarding various nutrients [4] while keeping a small environmental footprint [5]. However, as with meat, hygiene is obligatory when handling seeds and sprouts to prevent the growth of pathogenic bacteria. A potential lack of hygiene was the most likely cause of a major outbreak of the foodborne disease in 2011 caused by a pathogenic variant of Escherichia coli. In Germany, the outbreak caused 838 cases of hemolytic uretic syndrome and 2987 cases of gastroenteritis that claimed the lives of 47 people [6]. Tracing the source led to shipments of fenugreek seeds from Egypt [7], demonstrating the relevance of this food for food safety. Various pathogens may occur along the whole food production chain, where they are combated by antibiotics (fed to livestock) [8], heat treatment [9], gamma-irradiation [10], ozone [11], chlorine dioxide [12,13], or simple rinsing with water [14]. Even though these methods to lower pathogen levels are known and widely used, they still might not be sufficient, considering that in 2020 there were 180,000 confirmed cases of food and waterborne zoonosis-caused diseases in Europe [15]. In addition, the above-mentioned use of antibiotics in animal husbandry promotes resistance to conventional antibiotics [8]. These antibiotic-resistant bacteria can be transmitted from animals [16] as well as non-animal foodstuff [17] to humans, while other treatments can adversely the impact physical properties of food, or its by-products can be toxic [14].
The causal agent of at least 1876 cases of listeriosis in the EU is the ubiquitously found pathogenic bacterial species Listeria monocytogenes. It is a Gram-positive bacterium, that can cause severe disease and death in susceptible individuals, such as pregnant or old people [18]. Listeria monocytogenes can survive and reproduce in such a wide variety of environmental conditions that reproduction can take place within the cooling chain of foodstuff at 4 °C [19]. This explains why outbreaks still occur given the high food safety standards in the European Union [20]. Listeriosis is the disease with the highest case fatality rate (17%) of all food-borne-diseases. Furthermore, between 2008 and 2015 an increase in listeriosis cases were observed [21]. Ready-to-eat (RTE) foodstuff, such as salads, however, cannot be treated with most conventional methods, either due to legal constraints, customer preferences, or the risk of inflicting damage to the products [22,23]. This can lead to RTE foodstuff being contaminated with pathogens, including Listeria monocytogenes, in developed countries such as Finland [24]. Since the treatments already in place are not sufficient to prevent outbreaks of food-borne diseases development of new antimicrobial approaches for food decontamination is required.
One of these potential methods is Photodynamic Inactivation (PDI). It is based on light-sensitive compounds, called photosensitisers (PS), which produce reactive oxygen species (ROS) when illuminated with the PSs corresponding wavelength, typically in the visible light spectrum, which, in contrast to UV light, prevents harm to handlers of the light sources. The short-lived ROS, on the other hand, destroys the PS itself by photo-auto-destruction, which should dampen fears of bioaccumulation of the compounds in the human body. The production of ROS by PDI is efficient in an aqueous environment, which is usually given by the solvent of the PS [25]. PDI has already seen success in fighting fungi [26,27], bacteria with [28] and without resistance towards antibiotics [29,30,31,32,33] as well as multicellular organisms [34,35]. An additional benefit of multi-compound damage by the ROS produced by PDI, is a lower chance of evolutionary resistance, in contrast to regular antibiotics [36].
Natural photosensitisers, which are already classified as food additives, are ideally suited for PDI of foodstuff, as the compounds are likely to be ingested by consumers. Sodium magnesium chlorophyllin (Chl), for example, is listed as a food additive E140 [37,38,39,40]. Both, the absorption spectrum and chemical structure of Chl have been previously published [26]. Given the fact that potential pathogens are present in food and can reproduce even during refrigeration [19], PDI based on such photosensitisers could ensure long-lasting food safety without affecting other properties of foodstuff [39]. Getting rid of bacteria on foodstuff with PDI has already seen positive results on flat surfaces [33,37,38,39,40], however a limitation so far has been the fact, that two-dimensional light sources might be insufficient for complex geometries such as sprouts [30].
This study shall elucidate whether PDI based on Chl can reduce listeria on mung bean, buckwheat, and radish seeds. Possible effects on their germination rate and the weight of the sprouts shall be determined. Potential long-lasting effects of the PDI treatment of seeds on the bacterial load of germinated and stored mung bean sprouts shall be assessed. As an already established surrogate for Listeria monocytogenes, Listeria innocua will be used as a model system [41,42,43,44].
2. Material and Methods
2.1. Chemicals, Bacteria, and Seeds
Mung beans (Vigna radiata), buckwheat seeds (Fagopyrum esculentum), and radish seeds (Raphanus sativus var. sativus) were obtained from the local organic supermarket (all manufactured by Biosnacky Das Original, Rapunzel Naturkost, Legau, Germany). Sodium magnesium chlorophyllin was obtained from Carl Roth GmBH + Co KG (Karlsruhe, Germany). It was diluted using ddH2O (ultrapure water) to obtain a 10 mmol stock solution. It was stored in the dark frozen at −20 °C. Dulbecco’s modified Phosphate Buffered Saline (DPBS) was bought from Sigma-Aldrich (St Louis, MI, USA). The medium for the Petri dishes consisted of 37 g/L Brain-Heart-Infusion-Broth (BHI-Broth) supplemented with 15 g/L Agar-Agar Kobe I and were obtained from Roth.
Listeria innocua (DSM 20649; ATCC 33090) was grown in liquid media in Brain-Heart-Infusion broth (BHI, Roth) overnight at 37 °C at constant agitation with a MaxQ™ 4450 Benchtop Orbital Shaker (Thermo Scientific, Waltham, MA, USA).
2.2. PDI-Disinfection of Seeds
To remove any pre-existing fungal or bacterial contamination of seeds, they were placed in a 50 mL centrifuge tube and disinfected by addition of enough 15% H2O2 (Roth) to submerge the seeds. The seeds were mixed with H2O2 for 1 min (three-seed-species experiment) or 5 min (mung bean-only experiments). Afterwards, H2O2 was carefully aspirated. A single washing step with ddH2O using twice the volume of H2O2 used for initial seed disinfection was performed. As inoculum, 10 mL of a bacterial overnight culture of L. innocua was added into the centrifuge tube, resulting in total seed submersion in the bacterial culture. Afterwards, the liquid phase was carefully discarded. Three contaminated seeds were then transferred into one well of a sterile flat-bottomed 24-well plate. Three wells for each treatment were used, and thus nine seeds per treatment of one species (mung bean, buckwheat, radish). After the addition of 500 µL DPBS containing the respective photosensitiser, the 24-well plates were covered with Aluminium foil to exclude illumination with ambient light. The plates were placed on a shaker (Flow Laboratories, DSG Titertek, microplate shaker) and constantly agitated during the incubation (5 or 30 min). Illumination (10, 20, or 40 min) was provided by two LED arrays (480 diodes of diode type L-7113UVC, Kingbright Electronic Europe GmbH, Issum Germany) with the main emission wavelength at 395 nm. Samples were constantly agitated during illumination with a laboratory shaker. Radiant exposure was measured by using a spectrometer (LI-180, Licor, Li-Cor Biosciences. Lincoln, Nebraska, USA). Using the single-sided LED setup 20 min of illumination resulted in a radiant exposure of 27.8 J/cm2. Ten minutes from the double-sided LED-array illumination setup resulted in 28.2 J/cm2, 20 min in 56.4 J/cm2, and 40 min in 112.9 J/cm2 radiant exposure. After Illumination, the seeds were transferred into 2 mL reaction tubes containing 1 mL DPBS and subsequently shaken. This mixture was the basis for serial dilution in DPBS. If the PS/DPBS solution without the seed in the well was analysed, the remaining solution in the wells was the basis for the dilution series (visible by the prefix supernatant). Counting of colony forming units (CFU) on agar plates was performed after incubation for 48 h in a Heraeus Typ B 5042 E incubator (Hanau, Germany) at 37 °C in the dark. A summary of the workflow can be seen in Figure 1.
2.3. PDI Parameters
Details of the treatment protocol are shown in Table 1. Samples not illuminated were kept dark by wrapping the 24-well plates in aluminium foil and placing them on the shaker for the duration of the illumination. Samples with the prefix supernatant were based on the PS/DPBS supernatant of the respective seed treatments.
2.4. Germination of Seeds and Determination of Germination Rate
The supernatant for the bacterial assay was aspirated from the seeds. The seeds were placed in a 9 cm Petri dish and 10 mL of autoclaved tap water was added (~1.1 mL per seed). Petri dishes were covered with parafilm and aluminium foil to prevent water loss and further illumination. Seeds were left to germinate for 3 days at room temperature (22–24 °C). Seeds were counted as germinated when the growth of the germling was apparent. Seeds that showed fungal infections were excluded from the experiment.
2.5. Storage and Treatment of Sprouts
The germinated seeds were drained of the remaining water in the Petri dishes. The still-moist mung bean sprouts were kept in Petri dishes, sealed with parafilm, and stored in the fridge at 4–5 °C for 10 days in moist conditions. After 10 days, the sprouts were transferred into a 50 mL centrifuge tube containing 15 mL DPBS. The sprouts were shaken vigorously for one minute. As the sprouts remained intact, the DPBS containing the bacteria served as the basis for the dilution series. Counting of colony forming units (CFU) on agar plates was performed after incubation for 48 h in a Heraeus Typ B 5042 E incubator at 37 °C in the dark.
A different experimental setup was used to test whether the PDI with less contact time of the seeds with aqueous solutions would cause germination of the seeds. Thus, a setup without seed pre-treatment (H2O2 & bacteria) and without post-illumination treatment (storage in DPBS/PS solution during dilution series) was established For these experiments, 27 seeds per treatment were dipped into 100 µM Chl solution. As above three seeds, each were placed in one single well of a 24-well plate. Thus, nine wells were used per treatment. Treatments consisted of a light control (seeds dipped in DPBS) and PDI treatment (seeds dipped in 100 µM Chl solution). Incubation of seeds was carried out in a 24-well plate covered in aluminium foil for 30 min using constant agitation. Illumination was performed as described above. Illumination from top and bottom with the LED arrays resulted in a radiant exposure of 28.2 J/cm2. Seeds, without additional water, were sealed in Petri dishes with parafilm, covered in aluminium foil, and stored for 7 days at lab temperature (22–24 °C). After 7 days seeds were watered with 1.1 mL of autoclaved tap water per seed and again stored for 3 days, covered in aluminium foil, at lab temperature. Seeds were counted as germinated when the growth of the germling was apparent.
2.6. Determination of Sprout Mass
The moist sprouts were then weighed in a centrifuge tube on a 440-47N scale (Kern und Sohn GmbH, Balingen, Germany, ±0.1 g).
2.7. Data Analysis
Each experiment consisted of at least three biological replicates. Relative inactivation (r.i.) was calculated by dividing the CFUs in the double negative control with the CFUs of the respective treatment. The percentual germination rate was calculated by dividing the number of germinated seeds by the total amount of seeds per treatment and multiplying this result by 100. Column height indicates the mean of all replicates. Error bars represent the standard deviation. If no CFUs in the undiluted samples were visible in all replicates, bacterial densities greater than those of the double negative control (Control −/−) were assumed to have been inactivated and r.i. was plotted as the number of untreated controls (Control −/−). These total kills were classified as wipeout. The CFUs per gram of sprouts were calculated by:
The density of DPBS was assumed as 1.0 g/mL.
3. Results
Figure 2A shows the germination rate of mung bean, buckwheat, and radish seeds treated with 100 µM Chl, to elucidate the potential effects of PDI treatment. Figure 2B depicts the relative inactivation of mung bean, buckwheat, and radish seeds treated with 100 µM Chl. All numerical values can be found in the Supplementary Materials. Both Figure 2A,B are based on data from the same experiment. Seeds were incubated for 30 min and subsequently illuminated for 10 min with the LED arrays from the top and bottom of the well plate resulting in a radiant exposure of 28.2 J/cm2. The germination rate for mung bean and radish seeds is 100% in the double negative control (Control −/−) and the 100 µM Chl PDI treatment. The mean germination rate of buckwheat was 85% in the double negative control (Control −/−), 85% in the dark control, 81% in the light control, and 74% in the 100 µM Chl PDI treatment.
The relative inactivation of Listeria innocua of the 3 seed types (Figure 2B) is 1.1 log step of dark toxicity for mung bean samples. The relative inactivation of the other seeds and controls remained below 0.5 log steps, while the PDI treatment with 100 µM Chl resulted in 2.7, 3.1, and 0.8 log steps reduction on mung bean, radish, and buckwheat seeds, respectively, with the 10 min illumination setup.
Figure 3 shows the effect of the reduced contact time with aqueous solutions (to control for unwanted germination induced by disinfection and dilution steps) on the germination rate of the mung bean seeds. The PDI-treatment with 30 min dark incubation and the 10 min illumination (28.2 J/cm2 radiant exposure) did not cause mung bean seeds to sprout in the light control (DPBS instead of Chl) nor did the treatment with 100 µM Chl. Seeds sprouted within 3 days when moistened with water with a mean of 98.7% and 96.2% in the light control and the PDI treatment respectively.
In Figure 4 the relative inactivation of L. innocua after PDI treatment with a radiant exposure of 27.8 J/cm2 is plotted. Means of the relative inactivation of the light on dark control is below 0.3 log steps. PDI with 10 µM Chl achieves a relative inactivation of 3.2 log steps. The higher concentration of 100 µM Chl PDI-treatment resulted in a 4.2 log reduction of CFUs on the seeds. The bacteria-PS supernatant in the wells itself also achieves more than a 4.0 log reduction in bacterial densities with the 10 µM and 100 µM treatments.
PDI of mung beans which were exposed to 56.4 J/cm2 radiant exposure from the two 395 nm LED-arrays and subsequently grown to sprouts and stored in the fridge for 10 days can be seen in Figure 5A–D. The germination rate of mung beans (Figure 5A) remained at 100% in all six replicates of the PDI treatment with 30 min dark incubation and 100 µM Chl. The antimicrobial effectivity of 56.4 J/cm2 PDI treatment of the mung bean seeds (Figure 5B) shows a 4.6 log step reduction of L. innocua with 100 µM Chl and a photokilling below the limit of the detection (LOD) with an effect greater than 6.1 log steps in the remaining photosensitiser-liquid (supernatant) of the same treatment. The remaining reduction in bacterial load of the mung bean seed with the PDI treatment on the 10-day fridge-stored mung bean sprouts (Figure 5C) was still 3.4 log steps in the 100 µM Chl 30 min incubation treatment. The outcome 13 days after the PDI treatment regarding the mass of the sprouts shows, that the treatment did not alter the mean mass of the sprouts within the natural distribution (Figure 5D). The mean mass of the sprouts of the double negative, dark, and light control were 2.2, 2.2, and 2.0 g per nine sprouts, respectively. PDI treatment with 100 µM Chl of the former seeds, resulted in a mean mass of 2.1 g per nine sprouts. A 100% germination rate was achieved in all replicates in several experiments.
The average bacterial load with L. innocua of wet sprouts, calculated from Figure 5C,D, was 7.5 ∗ 106 CFU/g for the double negative control.
PDI treatment of mung bean seeds using illumination from above and below (112.9 J/cm2), with 10 or 100 µM Chl, as well as 5 min or 30 min dark incubation, and did not inhibit the germination rate of the seeds (Figure 6A). The relative inactivation of L. innocua is plotted in Figure 6B. Two-sided illumination (112.9 J/cm2) induced a 4.6 log step reduction for 100 µM Chl/30 min. incubation. The remaining supernatant in the same treatment showed a total kill of CFU, resulting in at least 6.5 log step inactivation.
4. Discussion
Environmentally benign, sustainable, nutritious, and healthy foods are required to tackle climate change. However, bacterial contaminations can make sprouts, which would fulfil these criteria dangerous foods. The experiments conducted in this study show that a simple two-sided LED-illumination Chl-based PDI can be used for effective decontamination of seeds (4.6 log steps reduction in CFUs at 56.4 J/cm2) and sprouts, without affecting the mass of the sprouts or the germination rates of the seeds. Illumination from two sides ensures better all-round activation of the PS and by this increases the effectiveness of PDI when compared to single-sided illumination [30]. After three days of room temperature, the sprouting of PDI-treated seeds, and subsequent storage at 4 °C for 10 days in a fridge, the decontamination efficiency is still at 3.4 log steps. Even though the bacterial load on seeds applied in this study is very high—several million CFUs per mL and about 7.5 ∗ 106 CFUs per gram of sprout—PDI with 100 µM Chl corresponding to 68.49 mg/L Chl is very effective. As prolonged illumination (40 min, 112.9 J/cm2) does not affect the mung bean seed germination rate and does not yield a higher decontamination rate, the potential to use higher Chl concentrations could be tested in follow-up studies. We hypothesize that, at the lowest concentration (1 µM), the smaller antibacterial effect of the 30 min when compared to the 5 min incubation is due to additional degradation of Chl during longer incubation, which is critical at very low concentrations.
One study on the shelf-life of PDI-treated food has been published so far: PDI treatment of Gram-negative Pseudomonas reinekei and Pseudomonas palleroniana on leafy greens (Pak Choi) caused a decrease in CFUs/g with a one-time 100 µM Chl based spray, followed by a constant 405 nm illumination with a radiant exposure of 22.27 J/cm2 per day. After 12 days of cooled and illuminated storage, resulting in a 267.25 J/cm2 radiant exposure, there were remarkable effects, leading to an >8.0 log step reduction in comparison to the control, while blue light alone only achieved a 3.0 log reduction in comparison with the control [39]. In our study, no constant illumination was used during storage periods. Even though the decontamination on Pak Choi is higher when compared to our study, constant illumination during storage and or transport is rarely possible due to stacked transportation and storage or a simple lack of illumination capabilities in the cooling chain. Additionally, constant illumination could influence nutrient levels in food [45,46]. Furthermore, since natural PS can bleach and degrade rather rapidly [25], it is not clear, how long the used PS would remain active in this setup with long illumination periods.
PDI based on other natural photosensitisers or their derivatives of mung bean germlings contaminated with Escherichia coli also demonstrated the applicability of PDI to increase food safety. Relative inactivation of E. coli on ungerminated mung bean with the curcumin derivate SACUR-3 at 100 µM showed a 3.0 log steps of relative inactivation. However, the effect on germlings was less pronounced (below 1.0 log r.i.). Smaller fenugreek seeds revealed a 5.6 log steps relative inactivation [30].
Decontamination of wheat seeds by PDI with Chl concentrations of 500 µM achieved about 1.5 log reduction after 50 min incubation and a radiant exposure of 30 J/cm2 (405 nm) [37], which is better than reported effects obtained with standard washing procedures with water [14]. The microstructure, length of root, and shoot of seed germlings were not influenced at 18 J/cm2 [37]. These results are comparable to our study indicated with respect to the low photokilling of L. innocua on buckwheat. As a working hypothesis, the lack of seed hulls on buckwheat and wheat seeds causes components of the seeds to be partially dissolved in aqueous solutions. This could adversely influence Chl from reaching the surface of the seed and still being activated because the dissolved seed content absorbs the relevant wavelengths. A set of experiments on L. monocytogenes biofilm using Chl-based PDI was published by Luksiene and Paskeviciute [40]. Biofilms grown on polyolefin packaging were treatable by PDI with 150 µM Chl with a 405 nm LED-based light source (12 mW/cm2) illuminated for just 5 min. This PDI treatment resulted in at least 4.5 log steps of relative inactivation of L. monocytogenes.
The comparison of PDI-treatment to alternative decontamination methods helps to assess the potential of PDI-based treatments for food safety. An established treatment is the use of peroxyacetic acid (PAA) (<70 ppm) ranging from 90 to 180 s and treatment with chlorine (<170 ppm) resulting in less than 2.0 logs reduction of Listeria monocytogenes on mung bean sprouts [47]. Both approaches are less effective than PDI. Decontamination of mung bean sprouts from Salmonella enteritidis (1.89 log reduction) and Escherichia coli (1.23 log reduction) by ultrasound alone was also less effective than the method presented in this study. A combination of ultrasound and chlorine (3 mg/L) achieved only a 2.0 log reduction [12]. Other studies tested a sequential combination of treatments against L. monocytogenes on mung beans. Here, a 2.0-to-5.0 log step reduction was achieved (20 min of 60 °C water +3–4% H2O2 + 0.2% acetic acid +1 min water rinsing) on mung bean seeds. The germination rate after the treatment was above 90% [48]. This treatment, though not applicable by organic standards, shows that a combination of treatments might be useful and could probably enhance the photokilling efficiency of PDI. Another, relatively new method to decontaminate foodstuff, is cold plasma. It achieves an approximate 2.0 to 5.0 log step reduction after a 180 s treatment. However, it reduces radish seed germination to 36%. Considering that more than 60% of the seeds would theoretically be lost because they do not sprout, and thus cannot be consumed, cold plasma treatment is inferior to PDI in this regard [49]. Fortunately, PDI and cold plasma treatment can kill a wide variety of pathogens [26,28,29,30,33,50,51]. Other seed disinfection methods usually achieve between 3.0 and 3.7 log steps relative inactivation of different microorganisms [52]. Only disinfection by applying high pressure has achieved 5.09 log steps of reduction on microorganisms [52], thus theoretically surpassing the effect size of PDI.
Organic acids are permitted in the EU for application in organic food treatment. However, only 0.6–2.3 log steps reductions were achievable without negative effects on the physical properties of the greens. Contact times of around 15 min required for a notable effect were however already deemed too long for economic decontamination [13]. Chlorine dioxide gas (4.1–4.3 mg/L) achieved between 3.0 and 6.0 log steps reduction after 30 min incubation, with the drawback of altering the sensory properties of the food [13]. Incubation periods of 20 or 30 min achieved between 1.2 and 5.8 log step reduction of Listeria monocytogenes at 2.7–4.1 g/mL chlorine dioxide in carrot, lettuce, and cabbage [13], which is slightly better in terms of duration and effectivity of the PDI treatment in this study. In the EU neither chlorine, chlorine dioxide, ozone, electrolyzed water, hydrogen peroxide nor peroxyacetic acid are approved for application on organic fresh-cut vegetables. Organic acids, such as citric or acetic acid, are however permitted according to EU guidelines [53,54]. PDI with PS already approved as food additives could therefore show potential even under strict EU guidelines for organic produce.
In general, the most widely used way of reducing the bacterial load of produce is rinsing with water. It is capable of providing a 1.0 log step CFU reduction of various microorganisms, mainly due to mechanical and turbulent effects [14]. Water consumption, often cooled under high energy use, is in the range of several tonnes per tonne of finished product [55], which of course can be detrimental to energy and water conservation plans, potentially hampering sustainability targets. The discussion of effect sizes must take realistic bacterial loads on alimentaria into account. Considering that the microbial load of freshly harvested food can range from 103 to 107.5 CFU per gram [23,55] rinsing with water is not always sufficient to extend shelf life. Ready-to-eat salads show up to 109 CFU/g at the end of shelf life [23] and spoilage of food becomes visible at above 108 CFU/g [14,23]. This particular microbial load is usually present after 8 days of storage. For example, after 14 days of storage at 3 °C, L. innocua densities on treated and untreated foodstuff are equal. Less ideal storage conditions of 8 °C reduce this timeframe to eight days [56]. This would mean, that even with a high initial reduction of bacterial load by PDI or other treatments, L. monocytogenes could still replicate, and thus prevent an even longer shelf life. The upper limit of contaminations with L. monocytogenes according to EU guidelines is 100 CFU/g and must be below the limit of detection which is 25 g of tested material [57,58]. This threshold might still be achievable on highly contaminated foodstuff (105–107 logs CFU/g) with PDI by treatment protocols with higher PS concentration and increased radiant exposure.
5. Conclusions
We here show that PDI using the food additive Chl E140 in concentrations up to 100 µM can achieve a substantial reduction (3–4.6 log steps) of L. innocua on mung bean seeds following illumination (56.4–112.9 J/cm2). Furthermore, the germination rate of the mung bean seeds was close to 100% even in very high radiant exposure settings of 112.9 J/cm2. The power of this approach on shelf life is proven by the lasting antilisterial effect on sprouted seeds of PDI-treated mung beans. Despite high standard deviations, PDI was able to maintain more than a 3.0 log reduction of bacterial load after treatment of seeds with 56.4 J/cm2 even after 13 days. No relevant effects on the mass of the sprouts after the PDI treatment have been observed. Germination of mung beans was not triggered by a PDI. Despite the relevant reduction of the bacterial load of radish (3.1 log reduction) and mung bean seeds (2.7 log reduction) by just 10 min of two-sided illumination, only a weak antibacterial effect on buckwheat seeds (0.8 log reduction) was evident. For mung bean and radish seeds, no adverse effects on the germination rate have been observed. For Buckwheat seeds germination rate was already not ideal in the controls and slightly decreased due to treatment. With improvements in PDI treatment protocols with near-natural photosensitisers, as shown in this study, increased food safety could be achieved.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/photonics10030239/s1, Table S1: Germination rate [%] of mung bean, buckwheat and radish seeds after PDI treatment via two-sided illumination from LED-arrays (radiant) exposure of 28.2 J/cm2. Control −/− and Dark control were not illuminated. Light control contained no Chl. n = 3. Table S2: Relative inactivation of L. innocua on mung bean, buckwheat and radish seeds after PDI treatment with 28.2 J/cm2. Dark control was not illuminated but contained 100 µM Chl. Light control was illuminated but did not contain Chl. PDI 100 µL contained 100 µM Chl and was illuminated. n = 3. Table S3: Germination rate [%] of PDI with shortened aqueous contact time with single-sided illumination for 20 minutes resulted in 28.2 J/cm2 radiant exposure on mung bean seeds. “Dry” indicates the germination rate after treatment, but without addition tap water to the seeds. “Wet” samples were the same mung bean seeds as ”Dry” of the respective control or PDI-treatment, but the germination rate was assessed after seven days of dry storage and subsequent germination post watering with 1.1 mL sterile tap water per seed. n = 3. Table S4: Relative inactivation of L. innocua on mung bean seeds after PDI treatment with one sided illumination for 27.8 J/cm2. Dark control was not illuminated but contained 100 µM Chl. Light control was illuminated but did not contain Chl. PDI 100 µM contained 100 µM Chl and was illuminated. n ≥ 5. Table S5: Germination rate [%] of 56.4 J/cm2 PDI treated mung bean seeds. Control −/− contained no Chl and was not illuminated. Dark control contained 100 µM Chl but was not illuminated. Light control was illuminated but contained no Chl. n = 6. Table S6: Relative inactivation of mung bean seeds of L. innocua after 56.4 J/cm2 of PDI treatment with illumination from two sides. Mean CFUs/mL: 1.2 ∗ 106. Liquid indicates the relative inactivation of the seedless PS/DPBS Liquid of the respective samples. Control −/− contained no Chl and was not illuminated. Dark control contained 100 µM Chl but was not illuminated. Light control was illuminated but contained no Chl. n ≥ 4. Table S7: Relative inactivation of L. innocua on mung bean sprouts, 13 days after PDI treatment with LED-array illumination from two sides resulting in a radiant exposure of 56.4 J/cm2. Mean CFUs/mL: 1.1 ∗ 106. Supernatant indicates the relative inactivation of the seedless PS/DPBS liquid of the respective samples. Control −/− contained no Chl and was not illuminated. Dark control contained 100 µM Chl but was not illuminated. Light control was illuminated but contained no Chl. n ≥ 3. Table S8: Mass of mung bean sprouts [g] that were treated as seeds with PDI from two-sided illumination by LED-arrays with 56.4 J/cm2 radiant exposure. Control −/− contained no Chl and was not illuminated. Dark control contained 100 µM Chl but was not illuminated. Light control was illuminated but contained no Chl. n = 6. Table S9: Germination rate [%] of 112.9 J/cm2 PDI treated mung bean seeds. Control −/− contained no Chl and was not illuminated. Dark control contained 100 µM Chl but was not illuminated. Light control was illuminated but contained no Chl. n ≥ 3. Table S10: Relative inactivation of mung bean seeds of L. innocua after 112.9 J/cm2 of PDI treatment with illumination from two sides. Mean CFUs/mL: 1.27 ∗ 106. Supernatant indicates the relative inactivation of the seedless PS/DPBS liquid of the respective samples. Control −/− contained no Chl and was not illuminated. Dark control contained 100 µM Chl but was not illuminated. Light control was illuminated but contained no Chl. n ≥ 3.
Author Contributions
Conceptualization, A.F. and K.P.; methodology, A.F., C.H. and K.P.; validation, A.F., C.H. and K.P.; investigation, A.F.; resources, J.L., M.F. and K.P.; data curation, A.F.; writing—original draft preparation, A.F.; writing—review and editing, A.F., C.H., J.L., M.F. and K.P.; visualization, A.F.; supervision, K.P.; project administration, K.P.; funding acquisition, K.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors are grateful to Annette Wimmer, Department of Biosciences and Medical Biology, Paris Lodron University of Salzburg, for proof-reading the manuscript.
Conflicts of Interest
The authors declare no competing interest.
Abbreviations
CFU: Colony forming unit(s); Chl: Sodium magnesium chlorophyllin (Na-Chlorophyllin); DPBS: Dulbecco’s modified phosphate buffered saline; PDI: Photodynamic Inactivation; PS: Photosensitiser; ROS: Reactive oxygen species; RPM: Revolutions per minute; SD: Standard deviation; BHI: Brain-heart-infusion broth; L. innocua: Listeria innocua; L. monocytogenes: Listeria monocytogenes; ddH2O: Ultrapure water; H2O2: Hydrogen peroxide; PAA: peroxyacetic acid; RTE: Read-to-eat; r.i.: relative inactivion.
References
- Tubiello, F.N.; Rosenzweig, C.; Conchedda, G.; Karl, K.; Gütschow, J.; Xueyao, P.; Obli-Laryea, G.; Wanner, N.; Qiu, S.Y.; de Barros, J.; et al. Greenhouse Gas Emissions from Food Systems: Building the Evidence Base. Environ. Res. Lett. 2021, 16, 65007. [Google Scholar] [CrossRef]
- Westhoek, H.; Lesschen, J.P.; Rood, T.; Wagner, S.; de Marco, A.; Murphy-Bokern, D.; Leip, A.; van Grinsven, H.; Sutton, M.A.; Oenema, O. Food Choices, Health and Environment: Effects of Cutting Europe’s Meat and Dairy Intake. Glob. Environ. Chang. 2014, 26, 196–205. [Google Scholar] [CrossRef] [Green Version]
- Irz, X.; Jensen, J.D.; Leroy, P.; Réquillart, V.; Soler, L.G. Promoting Climate-Friendly Diets: What Should We Tell Consumers in Denmark, Finland and France? Environ. Sci. Policy 2019, 99, 169–177. [Google Scholar] [CrossRef]
- Ebert, A.W. Sprouts and Microgreens—Novel Food Sources for Healthy Diets. Plants 2022, 11, 571. [Google Scholar] [CrossRef]
- Poore, J.; Nemecek, T. Reducing Food’s Environmental Impacts through Producers and Consumers. Science 2018, 360, 987–992. [Google Scholar] [CrossRef] [Green Version]
- Robert-Koch-Institut Abschließende Darstellung Und Bewertung Der Epidemiologischen Erkenntnisse Im EHEC O104:H4 Ausbruch, Deutschland.2011.1–23. Available online: https://www.rki.de/DE/Content/InfAZ/E/EHEC/EHEC_O104/EHEC-Abschlussbericht.pdf (accessed on 30 January 2023).
- EFSA. Tracing Seeds, in Particular Fenugreek (Trigonella Foenum-graecum) Seeds, in Relation to the Shiga Toxin-producing E. Coli (STEC) O104:H4 2011 Outbreaks in Germany and France. EFSA Support. Publ. 2011, 8, 7. [Google Scholar] [CrossRef] [Green Version]
- Manyi-Loh, C.; Mamphweli, S.; Meyer, E.; Okoh, A. Antibiotic Use in Agriculture and Its Consequential Resistance in Environmental Sources: Potential Public Health Implications. Molecules 2018, 23, 795. [Google Scholar] [CrossRef] [Green Version]
- Tompkin, R.B. Control of Listeria Monocytogenes in the Food-Processing Environment. J. Food Prot. 2002, 65, 709–725. [Google Scholar] [CrossRef] [Green Version]
- Farkas, J. Irradiation as a Method for Decontaminating Food: A Review. Int. J. Food Microbiol. 1998, 44, 189–204. [Google Scholar] [CrossRef]
- Brodowska, A.J.; Nowak, A.; Śmigielski, K. Ozone in the Food Industry: Principles of Ozone Treatment, Mechanisms of Action, and Applications: An Overview. Crit. Rev. Food Sci. Nutr. 2018, 58, 2176–2201. [Google Scholar] [CrossRef]
- Millan-Sango, D.; Sammut, E.; van Impe, J.F.; Valdramidis, V.P. Decontamination of Alfalfa and Mung Bean Sprouts by Ultrasound and Aqueous Chlorine Dioxide. LWT—Food Sci. Technol. 2017, 78, 90–96. [Google Scholar] [CrossRef]
- Sy, K.V.; Murray, M.B.; Harrison, M.D.; Beuchat, L.R. Evaluation of Gaseous Chlorine Dioxide as a Sanitizer for Killing Salmonella, Escherichia Coli O157:H7, Listeria Monocytogenes, and Yeasts and Molds on Fresh and Fresh-Cut Produce. J. Food Prot. 2005, 68, 1176–1187. [Google Scholar] [CrossRef]
- Allende, A.; Selma, M.V.; López-Gálvez, F.; Villaescusa, R.; Gil, M.I. Role of Commercial Sanitizers and Washing Systems on Epiphytic Microorganisms and Sensory Quality of Fresh-Cut Escarole and Lettuce. Postharvest Biol. Technol. 2008, 49, 155–163. [Google Scholar] [CrossRef]
- EFSA; ECDC. The European Union One Health 2020 Zoonoses Report. EFSA J. 2021, 19, e06971. [Google Scholar] [CrossRef]
- Kirbis, A.; Krizman, M. Spread of Antibiotic Resistant Bacteria from Food of Animal Origin to Humans and Vice Versa. Procedia Food Sci. 2015, 5, 148–151. [Google Scholar] [CrossRef] [Green Version]
- Founou, L.L.; Founou, R.C.; Essack, S.Y. Antibiotic Resistance in the Food Chain: A Developing Country-Perspective. Front. Microbiol. 2016, 7, 1881. [Google Scholar] [CrossRef]
- Buchanan, R.L.; Gorris, L.G.M.; Hayman, M.M.; Jackson, T.C.; Whiting, R.C. A Review of Listeria Monocytogenes: An Update on Outbreaks, Virulence, Dose-Response, Ecology, and Risk Assessments. Food Control. 2017, 75, 1–13. [Google Scholar] [CrossRef]
- Ramaswamy, V.; Cresence, V.M.; Rejitha, J.S.; Lekshmi, U.; Dharsana, K.S.; Prasad, P.; Vijila, M. Listeria-Review of Epidemiology and Pathogenesis. J. Microbiol. Immunol. Infect. 2007, 40, 4–13. Available online: www.academia.edu/download/31249641/listeria.pdf (accessed on 20 February 2023).
- Halbedel, S.; Wilking, H.; Holzer, A.; Kleta, S.; Fischer, M.A.; Lüth, S.; Pietzka, A.; Huhulescu, S.; Lachmann, R.; Krings, A.; et al. Large Nationwide Outbreak of Invasive Listeriosis Associated with Blood Sausage, Germany, 2018–2019. Emerg. Infect. Dis. 2020, 26, 1456–1464. [Google Scholar] [CrossRef]
- EFSA; ECDC. The European Union Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-borne Outbreaks in 2015. EFSA J. 2016, 14, e04634. [Google Scholar] [CrossRef]
- Mogren, L.; Windstam, S.; Boqvist, S.; Vågsholm, I.; Söderqvist, K.; Rosberg, A.K.; Lindén, J.; Mulaosmanovic, E.; Karlsson, M.; Uhlig, E.; et al. The Hurdle Approach-A Holistic Concept for Controlling Food Safety Risks Associated with Pathogenic Bacterial Contamination of Leafy Green Vegetables. A Review. Front. Microbiol. 2018, 9, 1965. [Google Scholar] [CrossRef] [Green Version]
- Barth, M.; Hankinson, T.R.; Zhuang, H.; Breidt, F. Microbiological Spoilage of Fruits and Vegetables. In Compendium of the Microbiological Spoilage of Foods and Beverages; Springer: New York, NY, USA, 2009; pp. 135–183. [Google Scholar]
- Nousiainen, L.L.; Joutsen, S.; Lunden, J.; Hänninen, M.L.; Fredriksson-Ahomaa, M. Bacterial Quality and Safety of Packaged Fresh Leafy Vegetables at the Retail Level in Finland. Int. J. Food Microbiol. 2016, 232, 73–79. [Google Scholar] [CrossRef]
- Plaetzer, K.; Krammer, B.; Berlanda, J.; Berr, F.; Kiesslich, T. Photophysics and Photochemistry of Photodynamic Therapy: Fundamental Aspects. Lasers Med. Sci. 2009, 24, 259–268. [Google Scholar] [CrossRef]
- Hamminger, C.; Glueck, M.; Fefer, M.; Ckurshumova, W.; Liu, J.; Tenhaken, R.; Plaetzer, K. Photodynamic Inactivation of Plant Pathogens Part II: Fungi. Photochem. Photobiol. Sci. 2022, 21, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Pereira Gonzales, F.; Maisch, T. Photodynamic Inactivation for Controlling Candida Albicans Infections. Fungal Biol. 2012, 116, 1–10. [Google Scholar] [CrossRef]
- Wimmer, A.; Glueck, M.; Ckurshumova, W.; Liu, J.; Fefer, M.; Plaetzer, K. Breaking the Rebellion: Photodynamic Inactivation against Erwinia Amylovora Resistant to Streptomycin. Antibiotics 2022, 11, 544. [Google Scholar] [CrossRef]
- Glueck, M.; Hamminger, C.; Fefer, M.; Liu, J.; Plaetzer, K. Save the Crop: Photodynamic Inactivation of Plant Pathogens I: Bacteria. Photochem. Photobiol. Sci. 2019, 18, 1700–1708. [Google Scholar] [CrossRef]
- Glueck, M.; Schamberger, B.; Eckl, P.; Plaetzer, K. New Horizons in Microbiological Food Safety: Photodynamic Decontamination Based on a Curcumin Derivative. Photochem. Photobiol. Sci. 2017, 16, 1784–1791. [Google Scholar] [CrossRef] [Green Version]
- Ryu, A.R.; Han, C.S.; Oh, H.K.; Lee, M.Y. Chlorin E6-Mediated Photodynamic Inactivation with Halogen Light against Microbes and Fungus. Toxicol. Environ. Health Sci. 2015, 7, 231–238. [Google Scholar] [CrossRef]
- Hanakova, A.; Bogdanova, K.; Tomankova, K.; Pizova, K.; Malohlava, J.; Binder, S.; Bajgar, R.; Langova, K.; Kolar, M.; Mosinger, J.; et al. The Application of Antimicrobial Photodynamic Therapy on S. Aureus and E. Coli Using Porphyrin Photosensitizers Bound to Cyclodextrin. Microbiol. Res. 2014, 169, 163–170. [Google Scholar] [CrossRef]
- Tortik, N.; Spaeth, A.; Plaetzer, K. Photodynamic Decontamination of Foodstuff from Staphylococcus Aureus Based on Novel Formulations of Curcumin. Photochem. Photobiol. Sci. 2014, 13, 1402–1409. [Google Scholar] [CrossRef]
- El-Tayeb, T.A.; El-Aziz, N.M.A.; Awad, H.H. A Study on the Dynamics of Aedes Caspius Larval Uptake and Release of Novel Haematoporphyrin. Afr. Entomol. 2013, 21, 15–23. [Google Scholar] [CrossRef]
- Lima, A.R.; Silva, C.M.; Caires, C.S.A.; Prado, E.D.; Rocha, L.R.P.; Cabrini, I.; Arruda, E.J.; Oliveira, S.L.; Caires, A.R.L. Evaluation of Eosin-Methylene Blue as a Photosensitizer for Larval Control of Aedes Aegypti by a Photodynamic Process. Insects 2018, 9, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maisch, T. Resistance in Antimicrobial Photodynamic Inactivation of Bacteria. Photochem. Photobiol. Sci. 2015, 14, 1518–1526. [Google Scholar] [CrossRef] [Green Version]
- Žudyte, B.; Lukšiene, Ž. Toward Better Microbial Safety of Wheat Sprouts: Chlorophyllin-Based Photosensitization of Seeds. Photochem. Photobiol. Sci. 2019, 18, 2521–2530. [Google Scholar] [CrossRef]
- Lukseviciute, V.; Luksiene, Z. Inactivation of Molds on the Surface of Wheat Sprouts by Chlorophyllin-Chitosan Coating in the Presence of Visible LED-Based Light. J. Photochem. Photobiol. B 2020, 202, 111721. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ding, Z.; Shao, C.; Xie, J. Chlorophyllin-Based 405 Nm Light Photodynamic Improved Fresh-Cut Pakchoi Quality at Postharvest and Inhibited the Formation of Biofilm. Foods 2022, 11, 2541. [Google Scholar] [CrossRef]
- Luksiene, Z.; Paskeviciute, E. Microbial Control of Food-Related Surfaces: Na-Chlorophyllin-Based Photosensitization. J. Photochem. Photobiol. B 2011, 105, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Hammer, P.; Bockelmann, W.; Hoffmann, W. Fate of Listeria Innocua during Production and Ripening of Smeared Hard Cheese Made from Raw Milk. J. Dairy Sci. 2017, 100, 7846–7856. [Google Scholar] [CrossRef] [Green Version]
- Girardin, H.; Morris, C.E.; Albagnac, C.; Dreux, N.; Glaux, C.; Nguyen-The, C. Behaviour of the Pathogen Surrogates Listeria Innocua and Clostridium Sporogenes during Production of Parsley in Fields Fertilized with Contaminated Amendments. FEMS Microbiol. Ecol. 2005, 54, 287–295. [Google Scholar] [CrossRef] [Green Version]
- Friedly, E.C.; Crandall, P.G.; Ricke, S.; O’Bryan, C.A.; Martin, E.M.; Boyd, L.M. Identification of Listeria Innocua Surrogates for Listeria Monocytogenes in Hamburger Patties. J. Food Sci. 2008, 73, M174–M178. [Google Scholar] [CrossRef]
- Sommers, C.H.; Cooke, P.H.; Fan, X.; Sites, J.E. Ultraviolet Light (254 Nm) Inactivation of Listeria Monocytogenes on Frankfurters That Contain Potassium Lactate and Sodium Diacetate. J. Food Sci. 2009, 74, M114–M119. [Google Scholar] [CrossRef]
- Brazaitytė, A.; Miliauskienė, J.; Vaštakaitė-Kairienė, V.; Sutulienė, R.; Laužikė, K.; Duchovskis, P.; Małek, S. Effect of Different Ratios of Blue and Red Led Light on Brassicaceae Microgreens under a Controlled Environment. Plants 2021, 10, 801. [Google Scholar] [CrossRef] [PubMed]
- Sheraz, M.A.; Kazi, S.H.; Ahmed, S.; Anwar, Z.; Ahmad, I. Photo, Thermal and Chemical Degradation of Riboflavin. Beilstein J. Org. Chem. 2014, 10, 1999–2012. [Google Scholar] [CrossRef]
- Neo, S.Y.; Lim, P.Y.; Phua, L.K.; Khoo, G.H.; Kim, S.J.; Lee, S.C.; Yuk, H.G. Efficacy of Chlorine and Peroxyacetic Acid on Reduction of Natural Microflora, Escherichia Coli O157: H7, Listeria Monocyotgenes and Salmonella Spp. on Mung Bean Sprouts. Food Microbiol. 2013, 36, 475–480. [Google Scholar] [CrossRef]
- Trząskowska, M.; Dai, Y.; Delaquis, P.; Wang, S. Pathogen Reduction on Mung Bean Reduction of Escherichia Coli O157:H7, Salmonella Enterica and Listeria Monocytogenes on Mung Bean Using Combined Thermal and Chemical Treatments with Acetic Acid and Hydrogen Peroxide. Food Microbiol. 2018, 76, 62–68. [Google Scholar] [CrossRef]
- Puligundla, P.; Kim, J.W.; Mok, C. Effects of Nonthermal Plasma Treatment on Decontamination and Sprouting of Radish (Raphanus Sativus L.) Seeds. Food Bioproc. Technol. 2017, 10, 1093–1102. [Google Scholar] [CrossRef]
- Wohllebe, S.; Ulbrich, C.; Grimm, D.; Pietsch, J.; Erzinger, G.; Richter, R.; Lebert, M.; Richter, P.R.; Häder, D.P. Photodynamic Treatment of Chaoborus Crystallinus Larvae with Chlorophyllin Induces Necrosis and Apoptosis. Photochem. Photobiol. 2011, 87, 1113–1122. [Google Scholar] [CrossRef]
- de Menezes, H.D.; Rodrigues, G.B.; Teixeira, S.d.P.; Massola, N.S.; Bachmann, L.; Wainwright, M.; Braga, G.U.L. In Vitro Photodynamic Inactivation of Plant-Pathogenic Fungi Colletotrichum Acutatum and Colletotrichum Gloeosporioides with Novel Phenothiazinium Photosensitizers. Appl. Environ. Microbiol. 2014, 80, 1623–1632. [Google Scholar] [CrossRef] [Green Version]
- Ding, H.; Fu, T.J.; Smith, M.A. Microbial Contamination in Sprouts: How Effective Is Seed Disinfection Treatment? J. Food Sci. 2013, 78, R495–R501. [Google Scholar] [CrossRef] [Green Version]
- European Commission Commission Regulation (EC). No 889/2008 of 5 September 2008 Laying Down Detailed Rules for the Implementation of Council Regulation (EC) No 834/2007 on Organic Production and Labelling of Organic Products With Regard to Organic Production, Labelling and Control; European Commission Commission Regulation (EC): Brussels, Belgium, 2008. [Google Scholar]
- Ölmez, H.; Kretzschmar, U. Potential Alternative Disinfection Methods for Organic Fresh-Cut Industry for Minimizing Water Consumption and Environmental Impact. LWT—Food Sci. Technol. 2009, 42, 686–693. [Google Scholar] [CrossRef]
- Lehto, M.; Sipilä, I.; Alakukku, L.; Kymäläinen, H.-R. Water Consumption and Wastewaters in Fresh-Cut Vegetable Production. Agric. Food Sci. 2014, 23, 246–256. [Google Scholar] [CrossRef]
- Gómez-López, V.M.; Ragaert, P.; Debevere, J.; Devlieghere, F. Decontamination Methods to Prolong the Shelf-Life of Minimally Processed Vegetables, State-of-the-Art. Crit. Rev. Food Sci. Nutr. 2008, 48, 487–495. [Google Scholar] [CrossRef]
- European Commission Commission Regulation (EC). No 1441/2007 of 5 December 2007 Amending Regulation (EC) No 2073/2005 on Microbiological Criteria for Foodstuffs; European Commission Commission Regulation (EC): Brussels, Belgium, 2007. [Google Scholar]
- European Commission Commission Regulation (EC). No 2073/2005 of 15 November 2005 on Microbiological Criteria for Foodstuffs; European Commission Commission Regulation (EC): Brussels, Belgium, 2005. [Google Scholar]
Figure 1.
The overall workflow of the seed and sprout experiments. Only mung beans in the 56.4 J/cm2 were processed after being grown into sprouts.
Figure 1.
The overall workflow of the seed and sprout experiments. Only mung beans in the 56.4 J/cm2 were processed after being grown into sprouts.

Figure 2.
(A) Germination rate [%] of mung bean, buckwheat, and radish seeds after PDI treatment using two-sided illumination with LED arrays, resulting in a radiant exposure of 28.2 J/cm2. Control −/− and Dark control were not illuminated. Light control contained no Chl. n = 3. (B) Relative inactivation of L. innocua on mung bean, buckwheat, and radish seeds after PDI treatment with 28.2 J/cm2. Mean CFUs/mL in Control −/−: Mung bean: 2.9 ∗ 106; Buckwheat: 8.0 ∗ 106; Radish: 2.3 ∗ 106. n = 3. Green bar: mung bean seeds, yellow bar: buckwheat seeds, red bar: radish seeds. Column height shows the mean, error bars show the standard deviation.
Figure 2.
(A) Germination rate [%] of mung bean, buckwheat, and radish seeds after PDI treatment using two-sided illumination with LED arrays, resulting in a radiant exposure of 28.2 J/cm2. Control −/− and Dark control were not illuminated. Light control contained no Chl. n = 3. (B) Relative inactivation of L. innocua on mung bean, buckwheat, and radish seeds after PDI treatment with 28.2 J/cm2. Mean CFUs/mL in Control −/−: Mung bean: 2.9 ∗ 106; Buckwheat: 8.0 ∗ 106; Radish: 2.3 ∗ 106. n = 3. Green bar: mung bean seeds, yellow bar: buckwheat seeds, red bar: radish seeds. Column height shows the mean, error bars show the standard deviation.

Figure 3.
Germination rate [%] of mung bean seeds with reduced contact time to aqueous solutions with one-sided illumination for 20 min resulted in 28.2 J/cm2 radiant exposure. “Dry” indicates the germination rate after treatment, but without the addition of tap water to the seeds. “Wet” samples were the same mung bean seeds as “Dry” of the respective control or PDI-treatment, but the germination rate was assessed after seven days of dry storage and subsequent germination post-watering with 1.1 mL sterile tap water per seed. n = 3. Column height shows the mean. Error bars show the standard deviation.
Figure 3.
Germination rate [%] of mung bean seeds with reduced contact time to aqueous solutions with one-sided illumination for 20 min resulted in 28.2 J/cm2 radiant exposure. “Dry” indicates the germination rate after treatment, but without the addition of tap water to the seeds. “Wet” samples were the same mung bean seeds as “Dry” of the respective control or PDI-treatment, but the germination rate was assessed after seven days of dry storage and subsequent germination post-watering with 1.1 mL sterile tap water per seed. n = 3. Column height shows the mean. Error bars show the standard deviation.

Figure 4.
Relative inactivation of L. innocua after PDI treatment of mung bean seeds. One-sided illumination resulted in 27.8 J/cm2. Mean CFUs/mL in Control −/−: 2.3 ∗ 106. The supernatant terminus indicates the relative inactivation of the seedless PS/DPBS supernatant of the respective samples. Column height shows the mean of 5 independent experiments. Error bars show the standard deviation.
Figure 4.
Relative inactivation of L. innocua after PDI treatment of mung bean seeds. One-sided illumination resulted in 27.8 J/cm2. Mean CFUs/mL in Control −/−: 2.3 ∗ 106. The supernatant terminus indicates the relative inactivation of the seedless PS/DPBS supernatant of the respective samples. Column height shows the mean of 5 independent experiments. Error bars show the standard deviation.

Figure 5.
(A) Germination rate [%] of mung bean seeds PDI treatment using radiant exposure of 56.4 J/cm2. n = 6. (B) Relative inactivation of L. innocua (radiant exposure = 56.4 J/cm2) by PDI treatment of mung beans with illumination from above and below (mean CFUs/mL: 1.2 ∗ 106, n ≥ 4). (C) Relative inactivation of L. innocua on mung bean sprouts 13 days after PDI treatment of seeds (mean CFUs/mL: 1.1 ∗ 106, n ≥ 3). Supernatant indicates the relative inactivation of the seedless PS/DPBS supernatant of the respective samples. The arrow “>” indicates a wipeout. (D) Mass of mung bean sprouts [g] after PDI treatment using two-sided illumination (56.4 J/cm2). n = 6. Column height shows the mean. Error bars show the standard deviation.
Figure 5.
(A) Germination rate [%] of mung bean seeds PDI treatment using radiant exposure of 56.4 J/cm2. n = 6. (B) Relative inactivation of L. innocua (radiant exposure = 56.4 J/cm2) by PDI treatment of mung beans with illumination from above and below (mean CFUs/mL: 1.2 ∗ 106, n ≥ 4). (C) Relative inactivation of L. innocua on mung bean sprouts 13 days after PDI treatment of seeds (mean CFUs/mL: 1.1 ∗ 106, n ≥ 3). Supernatant indicates the relative inactivation of the seedless PS/DPBS supernatant of the respective samples. The arrow “>” indicates a wipeout. (D) Mass of mung bean sprouts [g] after PDI treatment using two-sided illumination (56.4 J/cm2). n = 6. Column height shows the mean. Error bars show the standard deviation.

Figure 6.
(A) Germination rate [%] of PDI-treated mung bean seeds with a radiant exposure of 112.9 J/cm2. n ≥ 3. (B) Relative inactivation of L. innocua on mung bean seeds after the PDI treatment with a radiant exposure of 112.9 J/cm2. Mean CFUs/mL: 1.27 ∗ 106. Supernatant indicates the relative inactivation of the seedless PS/DPBS supernatant of the respective samples. n ≥ 3. “>” indicates a wipeout. Column height shows the mean. Error bars show the standard deviation.
Figure 6.
(A) Germination rate [%] of PDI-treated mung bean seeds with a radiant exposure of 112.9 J/cm2. n ≥ 3. (B) Relative inactivation of L. innocua on mung bean seeds after the PDI treatment with a radiant exposure of 112.9 J/cm2. Mean CFUs/mL: 1.27 ∗ 106. Supernatant indicates the relative inactivation of the seedless PS/DPBS supernatant of the respective samples. n ≥ 3. “>” indicates a wipeout. Column height shows the mean. Error bars show the standard deviation.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Treatment parameters of samples.
| Sample | Incubation (min) | Illumination (min) | Chl Concentration (µM) |
|---|---|---|---|
| Control −/− | 30 | 0 | 0 |
| Dark control | 30 | 0 | 100 |
| Light control | 30 | 10/20/40 | 0 |
| PDI, 1 µM Chl, 5 min | 5 | 10/20/40 | 1 |
| PDI, 10 µM Chl, 5 min | 5 | 10/20/40 | 10 |
| PDI, 100 µM Chl, 5 min | 5 | 10/20/40 | 100 |
| PDI, 1 µM Chl, 30 min | 30 | 10/20/40 | 1 |
| PDI, 10 µM Chl, 30 min | 30 | 10/20/40 | 10 |
| PDI, 100 µM Chl, 30 min | 30 | 10/20/40 | 100 |
| Supernatant Control −/− | 30 | 0 | 0 |
| Supernatant PDI, 10 µM Chl, 30 min | 30 | 10/20/40 | 10 |
| Supernatant PDI, 100 µM Chl, 30 min | 30 | 10/20/40 | 100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Fellner, A.; Hamminger, C.; Fefer, M.; Liu, J.; Plaetzer, K. Towards Microbial Food Safety of Sprouts: Photodynamic Decontamination of Seeds. Photonics 2023, 10, 239. https://doi.org/10.3390/photonics10030239
AMA Style
Fellner A, Hamminger C, Fefer M, Liu J, Plaetzer K. Towards Microbial Food Safety of Sprouts: Photodynamic Decontamination of Seeds. Photonics. 2023; 10(3):239. https://doi.org/10.3390/photonics10030239
Chicago/Turabian StyleFellner, Andreas, Christoph Hamminger, Michael Fefer, Jun Liu, and Kristjan Plaetzer. 2023. "Towards Microbial Food Safety of Sprouts: Photodynamic Decontamination of Seeds" Photonics 10, no. 3: 239. https://doi.org/10.3390/photonics10030239
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.