
The Potential of the Oil-Producing Oleaginous Yeast Rhodotorula mucilaginosa for Sustainable Production of Bio-Oil Energy

1
Department of Biotechnology, National Formosa University, 64 Wunhua Rd., Huwei Township 632301, Taiwan
2
Department of Food Nutrition and Health Biotechnology, Asia University, 500 Lioufeng Rd., Wufeng, Taichung 41354, Taiwan
3
Office of Environmental Safety and Health, Asia University, 500 Lioufeng Rd., Wufeng, Taichung 41354, Taiwan
4
Department of Medical Research, China Medical University Hospital, China Medical University, 91 Hsueh-Shih Rd., Taichung 40402, Taiwan
*
Authors to whom correspondence should be addressed.
Processes 2022, 10(2), 336; https://doi.org/10.3390/pr10020336
Submission received: 20 January 2022
/
Revised: 8 February 2022
/
Accepted: 8 February 2022
/
Published: 10 February 2022
Abstract
:At the 2021 United Nations Climate Change Conference (COP26), many countries in the world agreed to reach the goal of net-zero greenhouse gas emissions by 2050. This issue constrains energy use, petrochemical use, and related manufacturing production patterns. It is undeniable that the diesel engine of large equipment is still widely used in engineering applications, and it will not disappear in a short time. Many industrial projects still have to face the use of diesel engines. This study was focused on the development of oil-producing microorganisms to generate lipids. The oleaginous yeast Rhodotorula mucilaginosa (R. mucilaginosa) was selected for liquid-state cultivation, and the conditions for growth of the yeast cells were assessed. For the liquid fermentation culture with a fixed inoculation amount of 5%, it was determined that a suitable oil-producing culture was obtained on the sixth day, and the optimum conditions involved a carbon source concentration of 60 g/L, a yeast extract concentration of 0.5 g/L, and a KH2PO4 concentration of 7.0 g/L for each batch of culture experiments. In addition, the extraction method and solvent for the extraction of R. mucilaginosa lipids were chosen. The acid-heat method using the green organic solvent ethyl acetate exhibited the best performance for extraction of yeast lipids under environmentally friendly and safe conditions. The analysis of lipids showed that the fatty acids obtained primarily contained C16:0, C18:1 and C18:2, and especially C18:1 (41%) lipids, indicating that R. mucilaginosa lipids are a good bio-oil source for the production of biodiesel.
1. Introduction
Microbial oil is a renewable resource, so the use of microbial fermentation to produce bio-oil instead of using petroleum resources is of great significance [1]. Oil-producing microorganisms, especially yeast, can produce large amounts of lipids and also have a high growth rate spanning 24 continuous h, which makes the production times short and uniform, with no limiting requirements for cultivating space [2]. Rhodotorula mucilaginosa was used to produce single-cell lipids (SCOs), and the most suitable medium composition and lipid production conditions were found by exploring the different nutrient elements in the medium. The nature of fatty acids synthesized by oil-producing yeast depends on the nutrient content of the provided medium and on culture conditions [3,4,5,6,7,8,9,10]. Yeast lipids mainly include triglycerides, such as oleic acid (C18:1), linoleic acid (C18:2), stearic acid (C18:0), palmitic acid (C16:0) or palmitoleic acid (C16:1) [11,12]. The key factor affecting the biosynthetic pathway of R. mucilaginosa is the ratio of carbon to nitrogen [13]; medium adjustment is essential to the production of SCO from yeast. Therefore, the carbon-nitrogen ratio directly affects the lipid production of the strain. Selecting appropriate carbon and nitrogen sources (yeast extract) to cultivate yeast is important for achieving high lipid production [14]. Different oil-producing microorganisms have their preferred carbon sources, and different carbon sources have different effects on the growth of microorganisms. Compared to other organic carbon sources, when glucose is the main carbon source, it can provide different types of oil-producing microorganisms with higher bacterial organisms. Generally, glucose is the most common medium for the accumulation of high levels of carbon source lipids [15,16,17,18,19,20].
Ethyl acetate, which was used as the main solvent for extracting intracellular yeast lipids in this study, has proven to be an excellent choice for lipid and protein extraction [21,22,23,24]. Lu et al. (2016) [25] studied single-solvent extraction and compared the efficiencies of ethyl acetate, acetone, ethanol, and n-hexane in recovering lipids and observed that ethyl acetate has excellent selectivity in neutral lipid extraction, higher than that of chloroform [26]. Fine et al. (2013) [27] also used different solvents or mixtures and found that ethyl acetate is more polar than hexane, can improve the extraction rate of lipids and is less toxic than a chloroform-methanol blended solvent. Ethyl acetate has been used to extract lipids from various plant seeds [28,29]. Although it has similar properties to n-hexane, it is cheaper and safer when subjected to waste-treatment processes [28].
Lipid extraction is carried out with dry yeast powder or directly with wet bacteria [30]. To release lipids in bacteria, solvents can be used directly, or auxiliary solvents can be used for cell destruction (mechanical, chemical or biological methods) to increase the efficiency of lipid extraction [31]. The cell wall is first physically destroyed so that the lipid molecules mix easily with the solvent. In addition, there are many factors that affect the efficiency and cost of lipid extraction, including solvent selection, extraction time, temperature, and cell disruption method. The optimization of these factors is key for efficient and low-cost oil extraction [32]. The chemical acid-heat treatment of bacterial cells is mainly based on the acid hydrolysis of sugar polymers and proteins in the cell wall by hydrochloric acid or sulfuric acid, which makes the original compact cell wall loosen; then the cell wall is further destroyed with a boiling water bath and quick freezing treatment, and the organic solvent can extract the lipids in the cells effectively. This process has been effectively used as a pretreatment method for extracting bio-oil [33].
The storage-related thermal stability and physical response to low-temperature conditions for R. mucilaginosa lipids are also used as references for the safe production, storage, and transportation conditions of biodiesel. It is also known that yeast lipids have good thermal stability, high calorific value, and high kinematic viscosity [34]. Therefore, R. mucilaginosa lipids have certain prospects that are useful in the production of biodiesel. Scale-up cultivation can be used to quantify lipid production by R. mucilaginosa and ensure the energy sustainability and environmental sustainability of biofuel production by oil-producing microorganisms.
The main objective of this study was the development of a fermentation bioprocess to produce microbial oil-derived biodiesel. Microbial oil has been produced using the oleaginous yeast R. mucilaginosa cultivated in carbon sources, different glucose, yeast extract and potassium dihydrogen phosphate concentration in shake flask cultures. Furthermore, the several parameters of acid hydrolysis and solvent extraction on the cell disruption and lipid recovery from R. mucilaginosa oleaginous yeast were determined. Hence, microbial oil for production of lipids is an alternative to industrial production of biodiesel.
2. Materials and Methods
2.1. Strain and Growth Conditions
Rhodotorula mucilaginosa (BCRC 22360) was purchased from the Bioresources Collection and Research Center (BCRC) of the Food Industry Research and Development Institute (FIRDI) in Hsinchu, Taiwan. The yeast was maintained on YPD medium containing (g/L): glucose, 20.0; yeast extract, 10.0; peptone, 20.0; agar, 20.0 at 4 °C. The initial pre-inoculum of cells was grown for six days at 25 °C and 100 rpm in seed medium containing (g/L): glucose, 10.0; yeast extract, 3.0; peptone, 5.0; malt extract, 3.0. The cultivation medium for growth biomass contained (g/L): glucose, 60.0; yeast extract, 0.5; (NH4) 2SO4, 2.0, KH2PO4, 7.0; MgSO4·7H2O, 1.5. Cells were grown at 25 °C in an incubation shaker (100 rpm) for six days and then harvested. Then the liquid containing the well-growing strain was inoculated into the fermentation medium with 5% inoculum, 200 mL liquid medium (each liter contains: glucose 40 g, yeast extract 1.5 g, (NH4) 2SO4 2 g, KH2PO4 7 g and MgSO4·7H2O 1.5 g) in a 500 mL Erlenmeyer flask and shaken at 100 rpm and 25 °C for 15 days [15,16,17,18,19].
2.2. Suitable Liquid Culture Conditions
A 200 mL sample of liquid culture medium (glucose 40 g/L, yeast extract 1.5 g/L, (NH4)2SO4 2 g/L, KH2PO4 7 g/L and MgSO4·7H2O 1.5 g/L) was added to 500 mL Erlenmeyer flasks and sterilized, the inoculum 10 mL was inserted into the sterile box and covered with culture paper, and then the flask was shaken at 100 rpm and 25 °C for 0–8 days. The control carbon source concentration was 40, 60, 80 or 100 g/L, and 200 mL of liquid medium (with the chosen carbon source concentration, yeast extract 15 g/L, (NH4)2SO4 2 g/L, KH2PO4 7 g/L and MgSO4·7H2O 15 g/L) was put it into a 500 mL Erlenmeyer flask and sterilized; 10 mL of the inoculum was inserted into the sterile box, covered with culturing paper, and shaken for culturing at 100 rpm for six days at a culture temperature of 25 °C [13,14,15,16,17,18,19,20]. Next, the control nitrogen content source concentration was 0.5, 1.0, 1.5, or 2.0 g/L, and 200 mL of liquid medium (containing 60 g/L of the carbon source, the chosen yeast extract concentration, (NH4)2SO4 2 g/L, KH2PO4 7 g/L and MgSO4·7H2O 1.5 g/L) was put into a 500 mL Erlenmeyer flask and sterilized; then 10 mL of the inoculum was inserted into a sterile box, covered with culture paper, and shaken for culturing at 100 rpm at the optimum culture temperature for eight days. All experiments were performed in triplicate [15,16,17,18,19].
2.3. Biomass and Lipid Analysis
The cultured yeast fermentation broth was centrifuged for 10 min with 50 mL of fermentation broth, the supernatant was removed, and the sediment was washed several times with deionized water. The cells were collected by centrifugation, then dried at 50 °C to a constant weight, and the dry weight of the cells was measured (the dry weight is expressed in g/100 mL). We took 50 mL of fermentation broth to collect yeast cells by centrifugation, added 20 mL of 1 M hydrochloric acid solution, and placed the sample in a water bath at 60 °C for 10 min to obtain broken cells. Ten milliliters of ethyl acetate was mixed with the yeast cells, ultrasonically shaken for 30 min, and then centrifuged for 20 min to collect the extract. Then, 10 mL of ethyl acetate was added to repeat the extraction of yeast lipids, and the two extracts were combined and isothermally concentrated to dryness at 50 °C [30].
2.4. Selection of Extraction Conditions and Solvent
The aim was to study different organic solvents, such as dichloromethane and methanol, n-hexane, and ethyl acetate, to evaluate the lipid extraction of R. mucilaginosa and compare the effects of different solvents. Acid hydrolysis used hydrochloric acid concentrations of 1, 3 and 4 M on the amount of extracted lipid. Next, the influence of the acid hydrolysis temperature 60 and 100 °C on lipid extraction was assessed. A fixed acid hydrolysis extraction with selected hydrochloric acid concentration and the hot-water bath temperature could be used consistently for subsequent yeast lipid extraction. All experiments were carried out in triplicate.
2.5. Fatty Acid Composition Analysis
The composition of fatty acids was determined by fatty acid methyl esters through transesterification of lipid extracts followed by a gas chromatography analysis. For transesterification, to approximately 100 mg of yeast lipid extracts we added 1 mL to 0.5 mL of internal standard solution (heptadecanoic acid, C17), put the solution in a brown glass bottle, mixed in 1 mL of a 1 N solution of potassium hydroxide in methanol, filled the bottle with nitrogen and mixed with a vortex mixer for 30 s, placed the bottle in an 80 °C water bath for 15 min, removed it and allowed it to cool, added 1 mL of boron trifluoride-methanol solution, filled the bottle with nitrogen, mixed the solution with a vortex mixer for 30 s, and then placed the bottle in a 100 °C water bath for 15 min. After removing and cooling it, we added 1 mL of n-hexane, mixed the solution with a vortex mixer for 1 min, added 5 mL of saturated sodium chloride solution and shook gently, let the mixture stand for layering, then collected the supernatant and injected it into the gas chromatography/mass spectrometer for detection of the fatty acid composition by gas chromatography/mass (GC/Mass) with an Agilent 8975/7890A system (Agilent Technologies Inc., Santa Clara, CA, USA) and a capillary column. The measurement conditions were as follows: chromatographic column initial temperature: 170 °C for 40 min; heating rate: 3 °C/min; final temperature: 200 °C for 50 min; detector temperature: 300 °C; injector temperature: 250 °C; injection volume: 1 μL; mobile phase gas helium flow rate: 0.75 mL/min; and split ratio: 40:1 [34]. The analyzed fatty acid composition was identified and quantified by comparing the internal standard peak areas.
2.6. Differential Scanning Calorimetry (DSC) Thermal Analysis
Differential scanning calorimetry (TA Q20-RCS90, TA Instruments, Newcastle, DE, USA) was used to measure the thermal stability and low-temperature characteristics of lipids. The sample preparation conditions were kept as consistent as possible to reduce the fluctuation of the baseline, and the instrument was calibrated before the test. Approximately 1.5 mg of the yeast lipid and biodiesel was sealed in 20 μL aluminum lips, hermetic 900,794.901, and a special tool for the TA DSC instrument was used to tighten the seal. In all experiments, high-purity nitrogen was used as the purge gas, with a flow rate of 50 mL/min. The samples were heated at rates of 4, 6 or 8 °C/min, with a temperature range of 30–300 °C; the initial temperature was 30 to 70 °C, the sample was heated at a rate of 4, 6, or 8 °C/min, then held at 70 °C for 3 min, and next cooled to −60 °C for determining the low-temperature characteristics and thermal stability of the samples [35].
2.7. Heat of Combustion Measurement
One gram of yeast lipid was carefully weighed and placed on the test dish, which was then placed on the measuring device of the Parr 1341 oxygen bomb calorimeter instrument (Parr Instrument Company, Moline, IL, USA). At the end of this period, we recorded the time on the timer of the Parr 6775 digital thermometer and read the temperature [34], and then calculated the heat of combustion for each experiment.
2.8. Measurement of Isothermal 40 °C Kinematic Viscosity
We used a Cannon-Fenske75 U540 viscometer tube (Cannon Instrument Company, State College, PA, USA) fixed in a 40 °C isothermal water tank with a bracket, added 10 mL of yeast lipid to the viscometer tube, placed it in a 40 °C water bath for 10 min, and measured its kinematic viscosity [34].
2.9. Statistical Analyses
The experimental data obtained were expressed as mean ± standard error and subjected to an analysis of variance for a completely random design using SAS 9.4 Software (SAS Institute, Inc., Cary, NC, USA). The difference among means was determined using Duncan’s multiple range tests at the level of α = 0.05 [36].
3. Results and Discussion
3.1. Rhodotorula mucilaginosa Culture Time and Liquid Medium Composition
Figure 1 shows the morphology of the R. mucilaginosa strain for culturing in liquid medium. R. mucilaginosa was cultured for two days and the dry cells were extracted; then, the biomass of the cells and the amount accumulated were evaluated. Figure 2 shows that R. mucilaginosa was in the biomass and lipid production increased rapidly when cultured for 2–4 days. After the fourth day, the yeast began to accumulate secondary metabolite fatty acids; the logarithmic phase occurred in four to six days, during which time the yeast mass multiplied in the rapid division and multiplication stage. The highest mass was observed on the sixth day, and the lipid content began to decline after the eighth day.
3.2. Influence of Liquid Medium Composition on R. mucilaginosa Lipid Production
Determining the influence of different carbon sources on the growth of yeast, we used other carbon source substrates and selected low-cost glycerin and molasses as biomass sources with which we planned to reduce the production cost of bio-oil and improve the market competitiveness of bio-oil. Herein, it is the preliminary analysis method using Section 2.3. Biomass and lipid analysis. The concentrations of the various fixed carbon sources glucose, glycerol, and molasses were 60 g/L, and the experimental results are shown in Figure 3A. The results show that molasses at 7.62 g/L had the highest biomass content, followed by glucose and glycerol, but the use of glucose as a carbon source resulted in the highest lipid content, with lipid accumulation above 13.45%. According to these results, the concentration of glucose biomass was lower than that of molasses; glucose was the best carbon source for cell lipid synthesis, so glucose will be used as the main carbon source. We also confirmed that low-cost carbon sources such as glycerol and molasses can be used as competitive substitutes for microbial lipid production.
Glucose was used as the main carbon source, and four carbon source concentrations of 40, 60, 80, and 100 g/L were selected to evaluate the accumulation of biomass and lipids. Figure 3B shows that the yeast-cell lipid content tended to increase as the carbon source concentration was increased. In terms of the accumulation of lipids by yeast, the 60 g/L carbon source with glucose provided the highest lipid content, which accounted for approximately 14.86% of the dry yeast cell weight. For the yeast lipid content, when the carbon source concentration was 40 g/L, the carbon source was in short supply, and the yeast could no longer grow after the carbon source was depleted. In addition, if the carbon source concentration was higher than 60 g/L, the lipid content gradually decreased, which should have been due to the “oversupply” of carbon sources.
The fixed carbon source concentration was 60 g/L, and the effects of the different yeast extract concentrations 0.5, 1.0, 1.5, and 2.0 g/L on the growth and lipid accumulation of R. mucilaginosa were explored. Figure 3C shows that the initial yeast-extract concentration of 1.5 g/L resulted in the highest biomass, 7.68 g/L, but the lipid content was only 10.25%. When the initial yeast-extract concentration was 0.5 g/L, the biomass obtained was only 6.03 g/L, but the lipid content reached its highest level at 16.11%. Under culture conditions with excessive carbon sources and limited yeast extract, yeast grows slowly and R. mucilaginosa can continue to accumulate lipids produced from glucose. When the yeast was used for lipid production, the carbon source concentration was fixed at 60 g/L, and a suitable yeast extract concentration of 0.5 g/L was selected for the subsequent amplification experiment.
Due to the influence of KH2PO4 concentration on the growth of yeast, phosphorus is known to be responsible for regulating most cell activity and metabolism. Figure 3D shows that increasing the KH2PO4 concentration increased biomass and lipid content. When the KH2PO4 concentration reached 7.0 g/L, the yeast biomass reached 5.97 g/mL and the lipid production rate from multiple extractions was as high as 25.51%. A fixed carbon source concentration of 60 g/L, a yeast extract concentration of 0.5 g/L, and a KH2PO4 concentration of 7.0 g/L were used for subsequent amplification of lipid production.
3.3. Influence of Different Extraction Methods on R. mucilaginosa Lipids
Since solvents account for a large part of the extraction cost, it is necessary to find a cheaper and safer extraction solvent. The different organic solvents, dichloromethane-methanol (1:1), n-hexane, and ethyl acetate, were also studied to evaluate the lipid extraction of R. mucilaginosa, which was conducted by comparing different extraction methods of Table 1. Using the extraction results, a suitable extraction solvent was selected after consideration of environmental protection factors. The R. mucilaginosa lipid is an intracellular product that is wrapped by the yeast cell wall and cell membrane. To achieve improved production efficiency, it is necessary to break the yeast cell walls first. Acid hydrolysis was used to break the yeast cell walls, mainly by using hydrochloric acid and the sugar and protein in the cells to loosen the originally intact cell wall; then, during boiling in a water bath and a rapid cooling treatment, the yeast cell wall was further damaged. Table 1 shows data for the use of the acid hydrolysis method with a 4 M HCl solution in determining the lipid content obtained by extraction with different organic solvents. The three organic solvents were compared using the same acid hydrolysis method, and the extraction efficiency of ethyl acetate was the highest. The results of this study also proved that ethyl acetate can replace other toxic solvents and operational costs can be reduced by recycling.
3.4. Analysis of Fatty Acid Composition of Yeast Lipid
The fatty acid composition of yeast lipids was measured by GC/mass analysis (see Table 2). The fatty-acid composition of the R. mucilaginosa strain comprises mainly oleic acid (C18:1), palmitic acid (C16:0) and linoleic acid (C18:2), which is 41, 16, and 10% of fatty-acid composition, respectively. In particular, C18:1 exceeds 41%; this means that the fatty-acid composition of R. mucilaginosa is a good lipid source for the production of biodiesel. In addition, yeast lipids contain only trace amounts of long-chain fatty acids with C20 and C24 chains, which is conducive to the development of biodiesel. Certainly, as biodiesel, the composition of fatty acids affects the physical and chemical properties (saponification value and iodine value) and economic value of the lipid, and its composition also affects the quality of the biodiesel (oxidation stability, heat of combustion, and lubricity).
3.5. DSC Thermal Analysis of Yeast Lipid
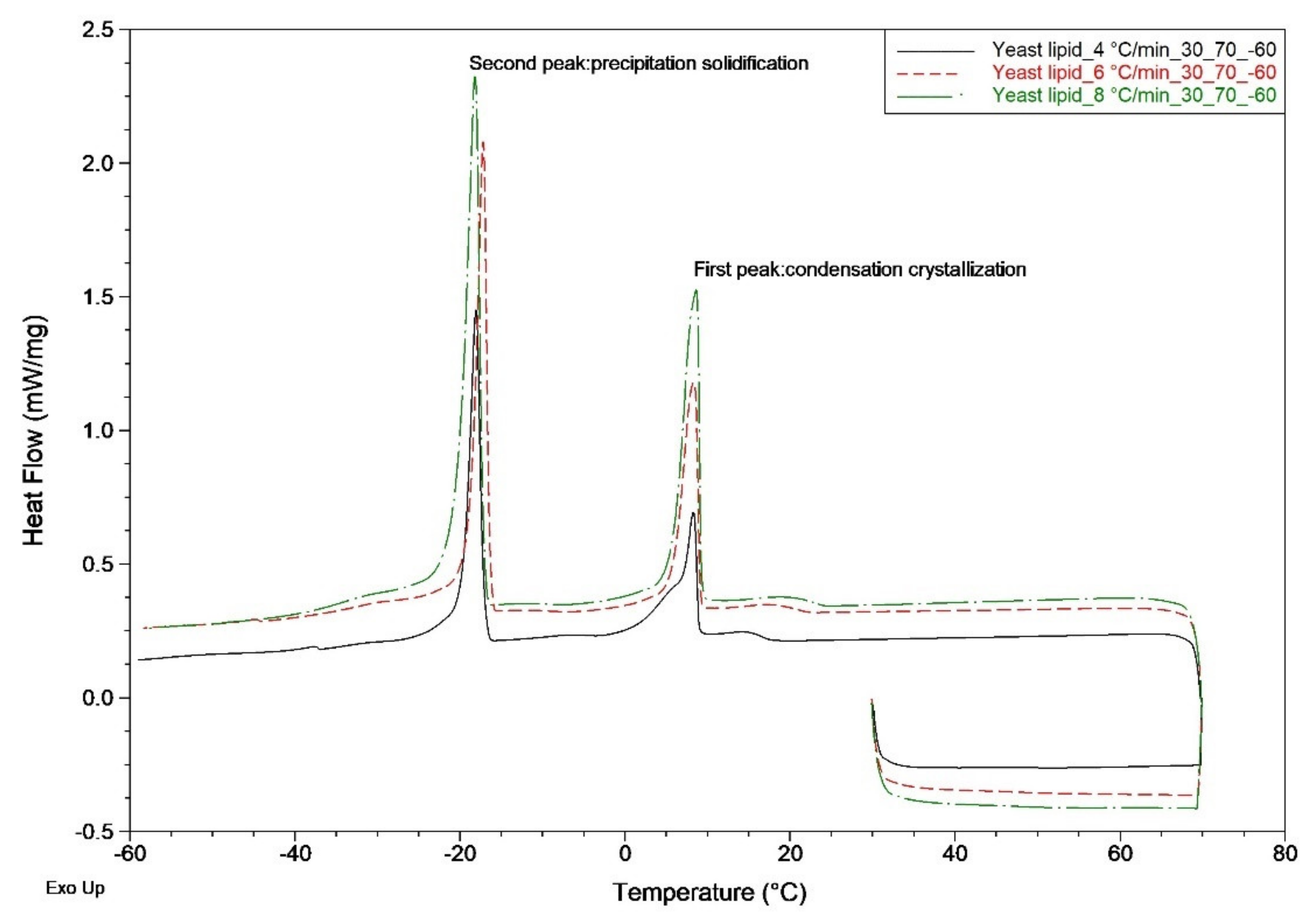
Using DSC and the initial temperatures to analyze the thermal characteristics and phase changes of yeast lipids under different thermal conditions, we estimated the peak maximum temperatures of endothermic and exothermic reactions and enthalpies of endothermic and exothermic reactions. Figure 4 shows that yeast lipids have good thermal properties. We found that during heating from 30 to 300 °C, the yeast lipid exhibited almost no obvious exothermic or endothermic reaction. In addition, DSC thermal analysis was used to assess the exothermic reaction characteristics of yeast lipids at general operating temperatures. This was done to observe the influence of summer or high operating temperatures on lipids and determine whether condensation would occur at low temperature. DSC nonthermal analysis conditions were conducted with increasing rates of 4, 6, and 8 °C/min, and the temperature rose from 30 to 70 °C at the beginning of the experiment, stayed at 70 °C for three minutes to reach a stable state, and then decreased to −60 °C, as displayed and listed in Table 3. Moreover, from Figure 5, we obtained data regarding cooling from 70 to −60 °C. Two obvious exothermic peaks were observed during cooling condensation; the first peak, for the condensation crystallization, was at approximately 9 °C and the enthalpy of crystallization was 24.51 J/g, and the second peak, for the precipitation solidification, was at ca. −17 °C and the enthalpy of solidification was 38.23 J/g (at cooling rate of 4 °C/min). From the abovementioned results, we learned that the saturated fatty acids contained in yeast lipids still displayed the low-temperature characteristics of lipids. Therefore, after the yeast lipid is transesterified, it still needs to be blended with petrochemical fuel to allow the lipid cycles of engines to operate normally at low temperature.
3.6. Measurement of Yeast Lipid Calorific Value and Isothermal 40 °C Kinematic Viscosity
A good fuel has a suitable heat of combustion. We put the yeast lipid into an oxygen bomb calorimeter to measure the heat of combustion, averaged the values from triplicate determinations, and then obtained the calorific value of yeast lipid as 38.16 MJ/kg. The carbon chain lengths in gasoline ranged from C4–C12, the carbon chain lengths in diesel ranged from C10–C22, and the carbon chain lengths in yeast lipid ranged from C14–C24. The difference in chain length causes the yeast lipid to have a higher calorific value of combustion. Kinematic viscosity is one of the important properties of diesel. If the carbon chain length of the lipid and the molecular weight increase, the kinematic viscosity will increase accordingly. Based on the results of the kinematic viscosity measurements run in triplicate, the kinematic viscosity of the yeast lipid (not yet transesterified) was 49.04 mm2/s.
The higher kinematic viscosity value, which indicates lower fluidity, may cause the fuel to stick in the fuel pipe or stick to the wall of the fuel cylinder, which will have an adverse effect on the engine. The solution is to blend the biodiesel with petrochemical fuel, but the kinematic viscosity of the diesel should not be too low. Because parts of the high-pressure fuel injection system are lubricated by diesel, poor lubrication will cause the parts to wear excessively. Biodiesel with a high kinematic viscosity must be blended with petrochemical diesel in a certain proportion to reduce the kinematic viscosity value to within the range specified in biodiesel regulations (1.9 to 6.0 mm2/s). According to Tsai et al. (2015) (previous research) [34], transesterifying yeast lipids to be made into biodiesel reduces the kinematic viscosity value of yeast lipid biodiesel to 6.37 mm2/s.
Therefore, R. mucilaginosa lipids have excellent prospects for the production of biodiesel. However, to reduce production costs, the use of the low-price molasses and the recycling of food processing and of agricultural waste as a carbon source for cultivation, and finding a suitable yeast oil extraction method, they will be improved to develop the prospect of producing bio-oil from oleaginous yeast.
4. Conclusions
We investigated the relationship between different nutrient elements and the growth of R. mucilaginosa. R. mucilaginosa exhibited higher lipid accumulation under nitrogen-restricted conditions, but the increase in carbon-source concentration promoted the accumulation of lipids. However, a further increase in the concentration of the carbon source inhibited the growth of yeast. Therefore, the shake flask fermentation was sampled on the sixth day, and culturing was done with glucose as the carbon source, an initial carbon source concentration of 60 g/L, an initial yeast extract concentration of 0.5 g/L, and a KH2PO4 concentration of 7.0 g/L. With these conditions the accumulation of yeast lipids reached a maximum. In addition, the selection of extraction method and solvent affected the extraction of R. mucilaginosa lipids. We determined that the acid-thermal extraction method had the best effect (lipid content 25.5%). Ethyl acetate had a higher lipid extraction efficiency than n-hexane and was also more efficient than a methylene chloride:methanol mixture; moreover, it is less toxic and safer to operate. From the analysis of fatty-acid composition of yeast lipids, the fatty acids mainly included C16:0, C18:1 (41%), and C18:2, which showed that R. mucilaginosa lipids are a good raw material for the production of biodiesel. Furthermore, the thermal stability and low-temperature physical characteristics of R. mucilaginosa lipids were also used as a reference to determine safe conditions for production, storage, and transportation of biodiesel. It is also known that yeast lipids have good thermal stability, high calorific value and suitable kinematic viscosity (biodiesel). We believe that with additional research, cultivation technology and methods will become more mature. As a result of larger-scale cultivation, increases in the production of lipids by R. mucilaginosa will be gradually realized, ensuring that the biofuels produced from oil-producing microbial raw materials are cost-effective, environmentally friendly, and sustainable.
Author Contributions
Conceptualization, S.-Y.T.; methodology, C.-P.L. and H.-T.Y.; writing—original draft, C.-P.L. and H.-T.Y.; writing—review and editing, S.-Y.T. and C.-P.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors are grateful to the Ministry of Science and Technology (MOST) for providing funding under contract No. MOST107-2221-E-468-007-MY2 and 109-2221-E-468-002-MY2.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bandhu, S.; Bansal, N.; Dasgupta, D.; Junghare, V.; Sidana, A.; Kalyan, G.; Hazra, S.; Ghosh, D. Overproduction of single cell oil from xylose rich sugarcane bagasse hydrolysate by an engineered oleaginous yeast Rhodotorula mucilaginosa IIPL32. Fuel 2019, 254, 115653. [Google Scholar] [CrossRef]
- Li, M.; Liu, G.L.; Chi, Z.; Chi, Z.M. Single cell oil production from hydrolysate of cassava starch by marine-derived yeast Rhodotorula mucilaginosa TJY15a. Biomass Bioenergy 2010, 34, 101–107. [Google Scholar] [CrossRef]
- Kitcha, S.; Cheirsilp, B. Screening of oleaginous yeasts and optimization for lipid production using crude glycerol as a carbon source. Energy Procedia 2011, 9, 274–282. [Google Scholar] [CrossRef] [Green Version]
- Bandhu, S.; Dasgupta, D.; Akhter, J.; Kanaujia, P.; Suman, S.K.; Agrawal, D.; Kaul, S.; Adhikari, D.K.; Ghosh, D. Statistical design and optimization of single cell oil production from sugarcane bagasse hydrolysate by an oleaginous yeast Rhodotorula sp. IIP-33 using response surface methodology. SpringerPlus 2014, 3, 691. [Google Scholar] [CrossRef] [Green Version]
- Santamauro, F.; Whiffin, F.M.; Scott, R.J.; Chuck, C.J. Low-cost lipid production by an oleaginous yeast cultured in non-sterile conditions using model waste resources. Biotechnol. Biofuels 2014, 7, 34. [Google Scholar] [CrossRef] [Green Version]
- Annamalai, N.; Sivakumar, N.; Oleskowicz-Popiel, P. Enhanced production of microbial lipids from waste office paper by the oleaginous yeast Cryptococcus curvatus. Fuel 2018, 217, 420–426. [Google Scholar] [CrossRef]
- Saenge, C.; Cheirsilp, B.; Suksaroge, T.T.; Bourtoom, T. Potential use of oleaginous red yeast Rhodotorula glutinis for the bioconversion of crude glycerol from biodiesel plant to lipids and carotenoids. Process Biochem. 2011, 46, 210–218. [Google Scholar] [CrossRef]
- Saran, S.; Mathur, A.; Dalal, J.; Saxena, R.K. Process optimization for cultivation and oil accumulation in an oleaginous yeast Rhodosporidium toruloides A29. Fuel 2017, 188, 324–331. [Google Scholar] [CrossRef]
- Subramaniam, R.; Dufreche, S.; Zappi, M.; Bajpai, R. Microbial lipids from renewable resources: Production and characterization. J. Ind. Microbiol. Biotechnol. 2010, 37, 1271–1287. [Google Scholar] [CrossRef] [PubMed]
- Tkachenko, A.F.; Tigunova, O.A.; Shulga, S.M. Microbial lipids as a source of biofuel. Cytol. Genet. 2013, 47, 343–348. [Google Scholar] [CrossRef] [Green Version]
- Leiva-Candia, D.E.; Pinzi, S.; Redel-Macías, M.D.; Koutinas, A.; Webb, C.; Dorado, M.P. The potential for agro-industrial waste utilization using oleaginous yeast for the production of biodiesel. Fuel 2014, 123, 33–42. [Google Scholar] [CrossRef]
- Prabhu, A.A.; Gadela, R.; Bharali, B.; Deshavath, N.N.; Dasu, V.V. Development of high biomass and lipid yielding medium for newly isolated Rhodotorula mucilaginosa. Fuel 2019, 239, 874–885. [Google Scholar] [CrossRef]
- Pacia, M.Z.; Pukalski, J.; Turnau, K.; Baranska, M.; Kaczor, A. Lipids, hemoproteins and carotenoids in alive Rhodotorula mucilaginosa cells under pesticide decomposition–Raman imaging study. Chemosphere 2016, 164, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Anschau, A. Lipids from oleaginous yeasts: Production and encapsulation. Nutr. Deliv. 2017, 5, 749–794. [Google Scholar]
- Ageitos, J.M.; Vallejo, J.A.; Veiga-Crespo, P.; Villa, T.G. Oily yeasts as oleaginous cell factories. Appl. Microbiol. Biotechnol. 2011, 90, 1219–1227. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.; Song, H.; Cao, Y.; Yang, H.; Hua, S.; Xia, C. The characteristics of biomass production, lipid accumulation and chlorophyll biosynthesis of Chlorella vulgaris under mixotrophic cultivation. Afr. J. Biotechnol. 2011, 10, 11620–11630. [Google Scholar]
- Leong, W.H.; Lim, J.W.; Lam, M.K.; Uemura, Y.; Ho, Y.C. Third generation biofuels: A nutritional perspective in enhancing microbial lipid production. Renew. Sustain. Energy Rev. 2018, 91, 950–961. [Google Scholar] [CrossRef]
- Liang, M.H.; Jiang, J.G. Advancing oleaginous microorganisms to produce lipid via metabolic engineering technology. Prog. Lipid Res. 2013, 52, 395–408. [Google Scholar] [CrossRef]
- Xing, D.; Wang, H.; Pan, A.; Wang, J.; Xue, D. Assimilation of corn fiber hydrolysates and lipid accumulation by Mortierella isabellina. Biomass Bioenergy 2012, 39, 494–501. [Google Scholar] [CrossRef]
- Quigg, A. Micronutrients. In The Physiology of Microalgae; Springer: Cham, Switzerland, 2016; Volume 6, pp. 211–223. [Google Scholar]
- Henderson, R.K.; Jiménez-González, C.; Constable, D.J.; Alston, S.R.; Inglis, G.G.; Fisher, G.; Sherwood, J.; Binks, S.P.; Curzons, A.D. Expanding GSK’s solvent selection guide–embedding sustainability into solvent selection starting at medicinal chemistry. Green Chem. 2011, 13, 854–862. [Google Scholar] [CrossRef]
- Alder, C.M.; Hayler, J.D.; Henderson, R.K.; Redman, A.M.; Shukla, L.; Shuster, L.E.; Sneddon, H.F. Updating and further expanding GSK’s solvent sustainability guide. Green Chem. 2016, 18, 3879–3890. [Google Scholar] [CrossRef]
- Cseri, L.; Razali, M.; Pogany, P.; Szekely, G. Organic solvents in sustainable synthesis and engineering. Green Chem. 2018, 513–553. [Google Scholar]
- Park, J.; Kim, B.; Chang, Y.K.; Lee, J.W. Wet in situ transesterification of microalgae using ethyl acetate as a co-solvent and reactant. Bioresour. Technol. 2017, 230, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Alam, M.A.; Pan, Y.; Nock, W.J.; Wang, Z.; Yuan, Z. Optimization of algal lipid extraction by mixture of ethyl acetate and ethanol via response surface methodology for biodiesel production. Korean J. Chem. Eng. 2016, 33, 2575–2581. [Google Scholar] [CrossRef]
- Lu, W.; Wang, Z.; Yuan, Z. Characteristics of lipid extraction from Chlorella sp. cultivated in outdoor raceway ponds with mixture of ethyl acetate and ethanol for biodiesel production. Bioresour. Technol. 2015, 191, 433–437. [Google Scholar] [CrossRef]
- Fine, F.; Vian, M.A.; Tixier, A.S.F.; Carre, P.; Pages, X.; Chemat, F. Les agro-solvants pour l’extraction des huiles vegetales issues de graines oléagineuses. Oilseeds Fats Crops Lipids 2013, 20, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Lohani, U.C.; Fallahi, P.; Muthukumarappan, K. Comparison of ethyl acetate with hexane for oil extraction from various oilseeds. J. Am. Oil Chem. Soc. 2015, 92, 743–754. [Google Scholar] [CrossRef]
- Jiyane, P.C.; Tumba, K.; Musonge, P. Optimisation of Croton gratissimus oil extraction by n-hexane and ethyl acetate using response surface methodology. J. Oleo Sci. 2018, 67, 369–377. [Google Scholar] [CrossRef]
- Xu, L.; Brilman, D.W.W.; Withag, J.A.; Brem, G.; Kersten, S. Assessment of a dry and a wet route for the production of biofuels from microalgae: Energy balance analysis. Bioresour. Technol. 2011, 102, 5113–5122. [Google Scholar] [CrossRef]
- Teo, C.L.; Idris, A. Enhancing the various solvent extraction method via microwave irradiation for extraction of lipids from marine microalgae in biodiesel production. Bioresour. Technol. 2014, 171, 477–481. [Google Scholar] [CrossRef]
- Deshmukh, S.; Kumar, R.; Bala, K. Microalgae biodiesel: A review on oil extraction, fatty acid composition, properties and effect on engine performance and emissions. Fuel Process. Technol. 2019, 191, 232–247. [Google Scholar] [CrossRef]
- Lee, I.; Han, J.I. The effects of waste-activated sludge pretreatment using hydrodynamic cavitation for methane production. Ultrason. Sonochem. 2013, 20, 1450–1455. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.Y.; Lin, H.Y.; Lu, G.Y.; Lin, C.P. Solid byproducts of Aurantiochytrium sp. oil made into the biodiesel. J. Therm. Anal. Calorim. 2015, 120, 563–572. [Google Scholar] [CrossRef]
- Tsai, S.Y.; Tsay, G.J.; Li, C.Y.; Hung, Y.T.; Lin, C.P. Assessment of melting kinetics of sugar-reduced silver ear mushroom ice cream under various additive models. Appl. Sci. 2020, 10, 2664. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.P.; Tsai, S.Y. Differences in the moisture capacity and thermal stability of Tremella fuciformis polysaccharides obtained by various drying processes. Molecules 2019, 24, 2856. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Flow diagram showing the oil-producing yeast R. mucilaginosa for sustainable production of bio-oil.
Figure 1.
Flow diagram showing the oil-producing yeast R. mucilaginosa for sustainable production of bio-oil.
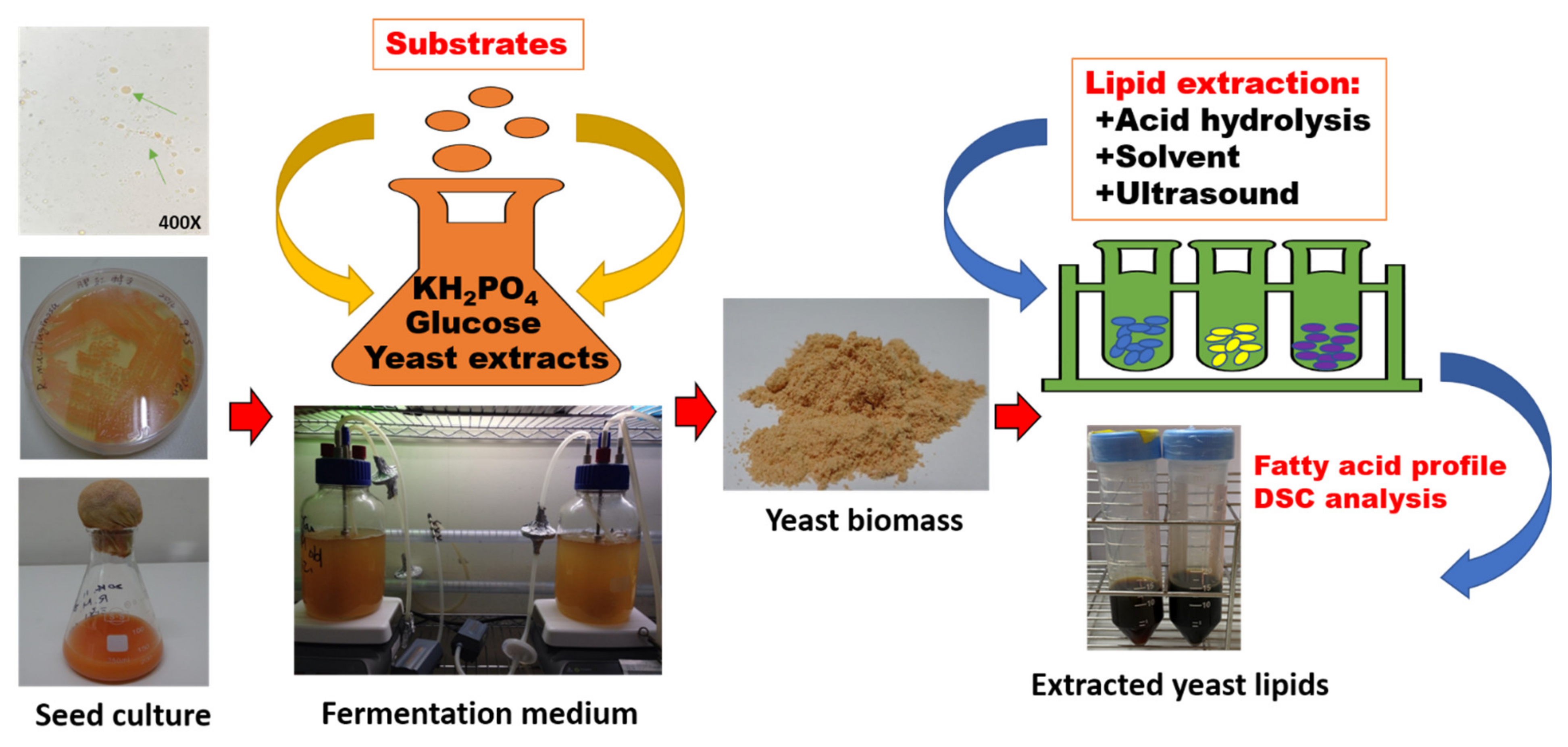
Figure 2.
R. mucilaginosa lipid accumulation and cell growth curves.
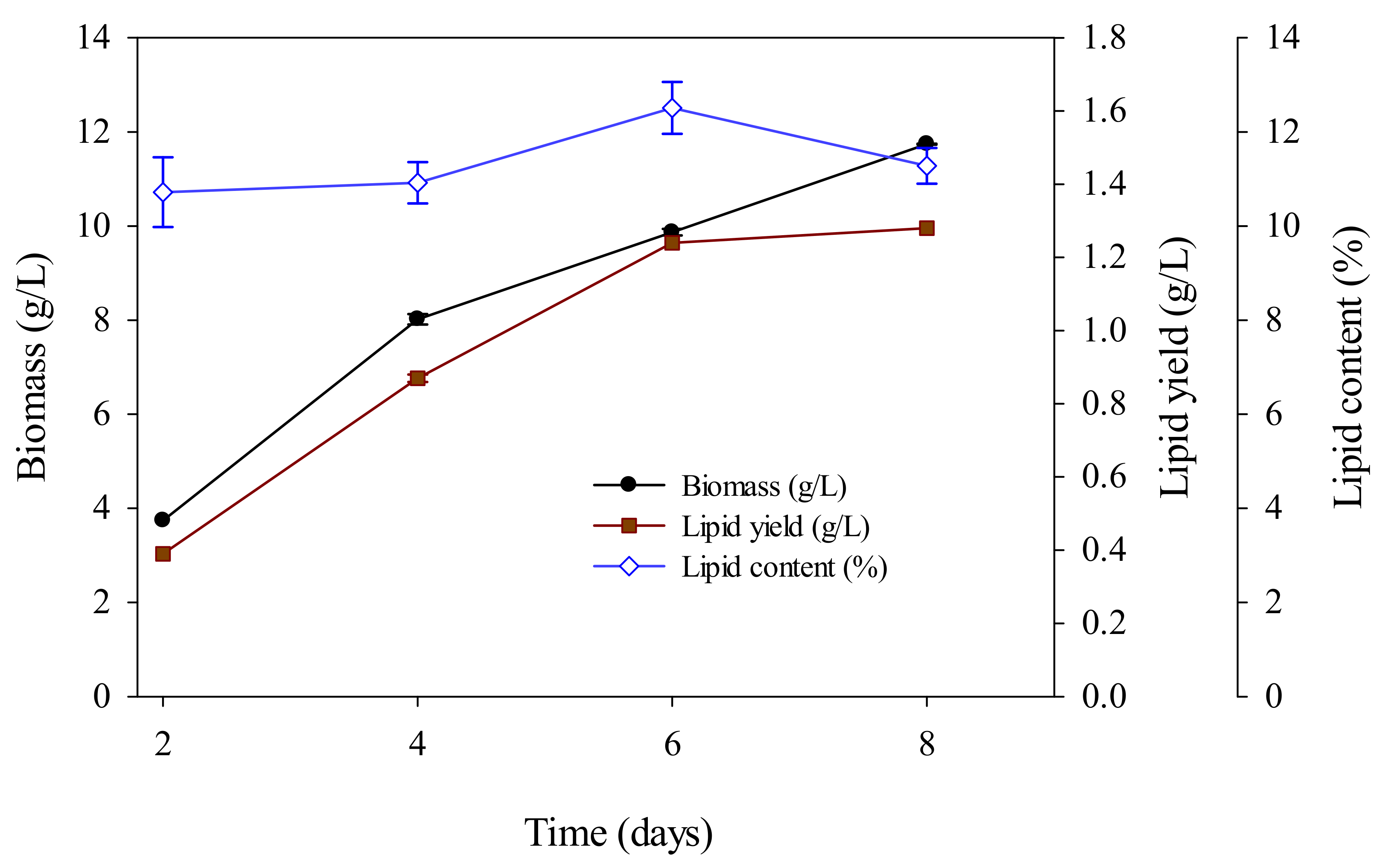
Figure 3.
Optimization of the medium for R. mucilaginosa lipid production. Data are given as the mean SD, n = 3. (A) Impact of carbon sources on cell growth and lipid accumulation. (B) Lipid content and biomass of the yeast growth at different initial glucose concentration. (C) Lipid content and biomass of the yeast growth at different initial yeast extract concentration. (D) Lipid content and biomass of the yeast growth at different initial potassium dihydrogen phosphate concentration.
Figure 3.
Optimization of the medium for R. mucilaginosa lipid production. Data are given as the mean SD, n = 3. (A) Impact of carbon sources on cell growth and lipid accumulation. (B) Lipid content and biomass of the yeast growth at different initial glucose concentration. (C) Lipid content and biomass of the yeast growth at different initial yeast extract concentration. (D) Lipid content and biomass of the yeast growth at different initial potassium dihydrogen phosphate concentration.

Figure 4.
R. mucilaginosa lipids analyzed by differential scanning calorimetry (DSC) with heating rates of 4, 6, and 8 °C/min, with the temperature ranging from 30 to 300 °C.
Figure 4.
R. mucilaginosa lipids analyzed by differential scanning calorimetry (DSC) with heating rates of 4, 6, and 8 °C/min, with the temperature ranging from 30 to 300 °C.

Figure 5.
R. mucilaginosa lipids measured with DSC heating rates of 2, 4, 6, and 8 °C/min, with the operating temperature ranging from 30 to 70 °C, and then cooling to −60 °C.
Figure 5.
R. mucilaginosa lipids measured with DSC heating rates of 2, 4, 6, and 8 °C/min, with the operating temperature ranging from 30 to 70 °C, and then cooling to −60 °C.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Comparison of R. mucilaginosa lipid content obtained with the acid hydrolysis and different solvents of extraction conditions.
Table 1.
Comparison of R. mucilaginosa lipid content obtained with the acid hydrolysis and different solvents of extraction conditions.
| Disruption and Extraction Method | Solvent | Conditions | Lipid Yield (g/L) | Lipid Content (%) |
|---|---|---|---|---|
| Soxhlet | Ethyl oxide | 1 g/150 mL, 50 °C, 6 h | 0.40 ± 0.10 e | 7.90 ± 0.15 f |
| Solvent | Dichloromethane: methanol (1:1) | 1 g/10 mL, sonication 30 min | 0.61 ± 0.02 d | 11.38 ± 0.49 e |
| Acid hydrolysis + Solvent | Dichloromethane:methanol (1:1) | 1 g/1 M HCl, 10 min, 60 °C; 10 mL, sonication 30 min | 0.70 ± 0.01 cd | 14.85 ± 0.26 d |
| Acid hydrolysis + Solvent | n-Hexane | 1 g/1 M HCl, 10 min, 60 °C; 10 mL, sonication 30 min | 0.80 ± 0.01 cd | 15.80 ± 0.25 d |
| Acid hydrolysis + Solvent | Ethyl acetate | 1 g/1 M HCl, 10 min, 60 °C; 10 mL, sonication 30 min | 0.91 ± 0.01 c | 18.80 ± 0.03 c |
| Acid hydrolysis + Solvent | Ethyl acetate | 1 g/3 M HCl, 10 min, 60 °C; 10 mL, sonication 30 min | 1.14 ± 0.10 b | 18.29 ± 0.46 c |
| Acid hydrolysis + Solvent | Ethyl acetate | 1 g/4 M HCl, 10 min, 60 °C; 10 mL, sonication 30 min | 1.24 ± 0.01 b | 20.29 ± 0.32 b |
| Acid hydrolysis + Solvent | Ethyl acetate | 1 g/4 M HCl, 10 min, 100 °C; 10 mL, sonication 30 min | 1.50 ± 0.30 a | 25.51 ± 2.07 a |
Remarks: Each value is expressed as mean ± SD (n ≥ 3). Means with same capital letter or small letter within a column do not differ significantly (p > 0.05).
Table 2.
Fatty acid profile of single-cell oil from R. mucilaginosa.
| Fatty Acids | w/w, % |
|---|---|
| Myristic acid (C14:0) | 1.08 |
| Palmitic acid (C16:0) | 15.85 |
| Palmitoleic acid (C16:1) | 1.92 |
| Stearic acid (C18:0) | 0.86 |
| Oleic acid (C18:1) | 41.44 |
| Linoleic acid (C18:2) | 10.35 |
| Linolenic acid (C18:3) | 0.97 |
| Eicosenoic acid (C20:1) | 0.25 |
| Lignoceric acid (C24:0) | 0.65 |
| Total saturation | 18.45 |
| Total monounsaturated | 43.61 |
| Total polyunsaturated | 11.32 |
Table 3.
DSC analysis results for R. mucilaginosa lipids with heating rates of 4, 6 and 8 °C/min, with operating temperature ranging from 30 to 70 °C, and then cooling to −60 °C.
Table 3.
DSC analysis results for R. mucilaginosa lipids with heating rates of 4, 6 and 8 °C/min, with operating temperature ranging from 30 to 70 °C, and then cooling to −60 °C.
| Mass (mg) | Cooling Rate (°C/min) | 1exoTonset (°C) | 1exoTpeak (°C) | 1Enthalpy (J/g) | 2exoTonset (°C) | 2exoTpeak (°C) | 2Enthalpy (J/g) |
|---|---|---|---|---|---|---|---|
| 1.50 | 4 | 8.94 | 8.30 | 24.51 | −17.15 | −18.06 | 38.23 |
| 1.45 | 6 | 9.08 | 8.34 | 21.16 | −16.43 | −17.19 | 37.94 |
| 1.48 | 8 | 9.20 | 8.61 | 26.25 | −17.19 | −18.19 | 42.54 |
Remarks: Standard deviation: temperature accuracy: +/− 0.1; temperature precision: +/− 0.05. Calorimetric reproducibility: +/− 1 mass%; sensitivity: 1.0 μW.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tsai, S.-Y.; Yu, H.-T.; Lin, C.-P. The Potential of the Oil-Producing Oleaginous Yeast Rhodotorula mucilaginosa for Sustainable Production of Bio-Oil Energy. Processes 2022, 10, 336. https://doi.org/10.3390/pr10020336
AMA Style
Tsai S-Y, Yu H-T, Lin C-P. The Potential of the Oil-Producing Oleaginous Yeast Rhodotorula mucilaginosa for Sustainable Production of Bio-Oil Energy. Processes. 2022; 10(2):336. https://doi.org/10.3390/pr10020336
Chicago/Turabian StyleTsai, Shu-Yao, Hsuan-Ti Yu, and Chun-Ping Lin. 2022. "The Potential of the Oil-Producing Oleaginous Yeast Rhodotorula mucilaginosa for Sustainable Production of Bio-Oil Energy" Processes 10, no. 2: 336. https://doi.org/10.3390/pr10020336
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.