In Vitro Regeneration Potential of White Lupin (Lupinus albus) from Cotyledonary Nodes

 , ,
, ,
Abstract
:1. Introduction
2. Results
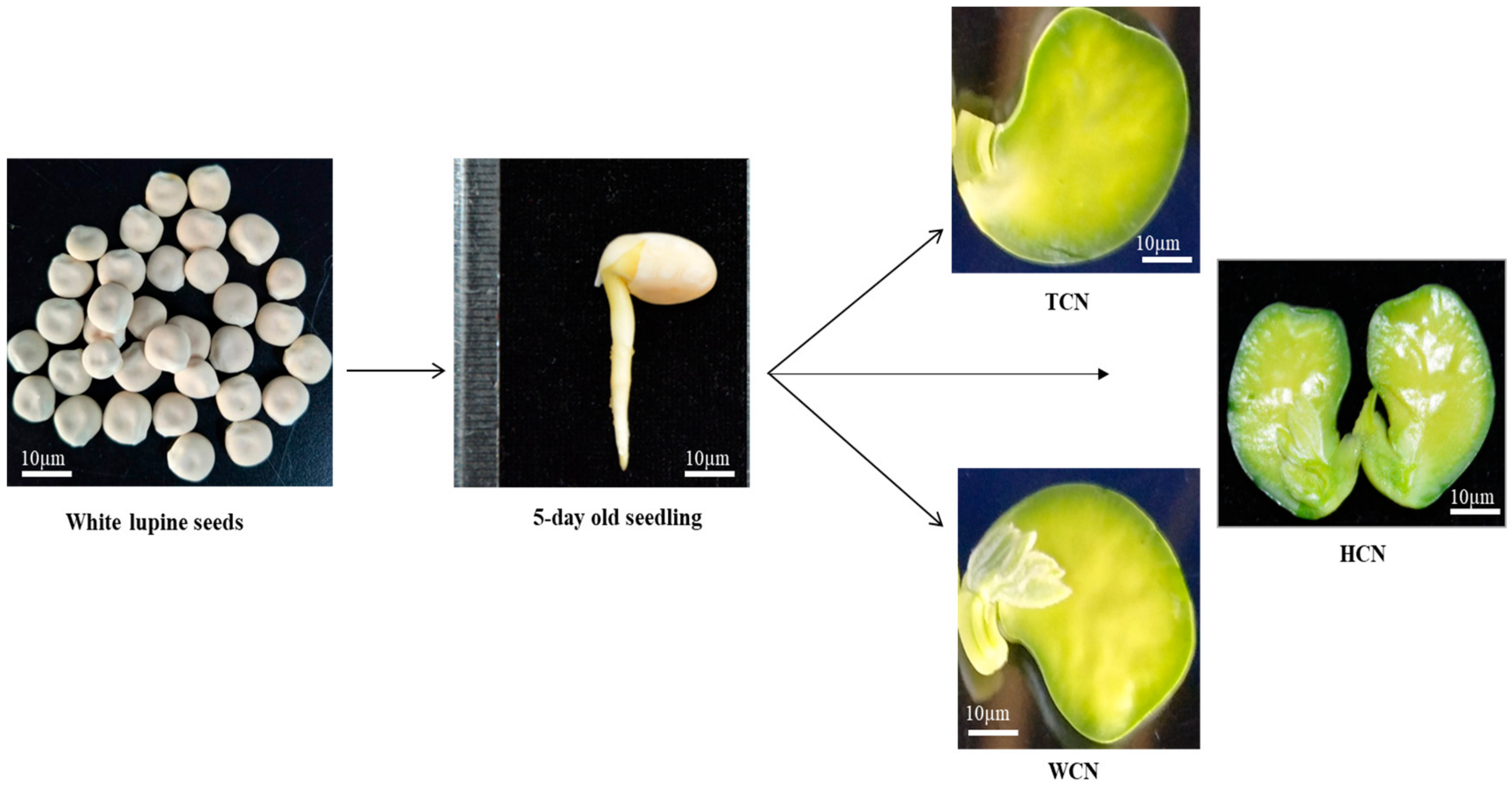
2.1. Efficiency of Shoot Regeneration from Three Cotyledonary Nodes
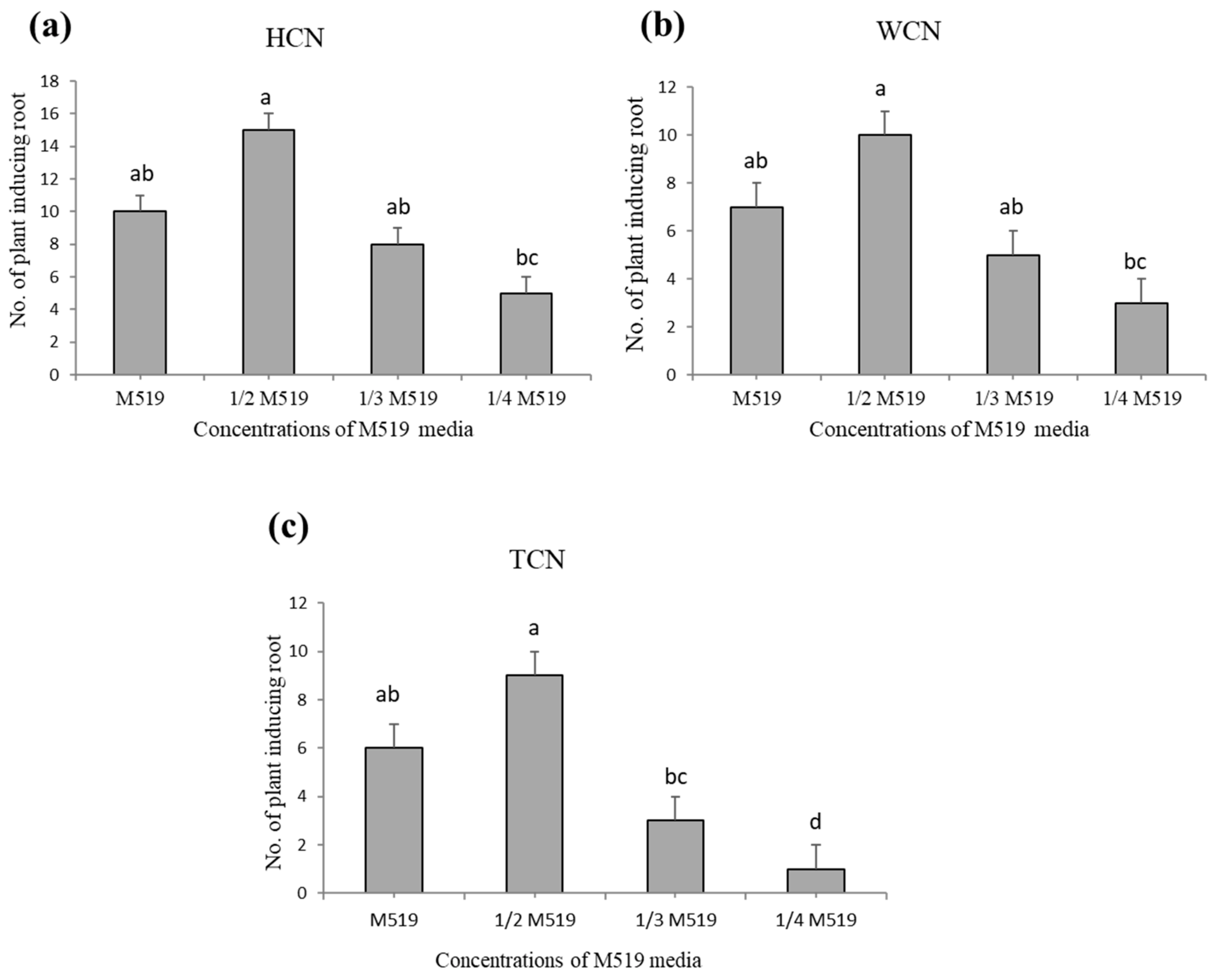
2.2. Concentration of M519 Medium-Affected Shoot Regeneration
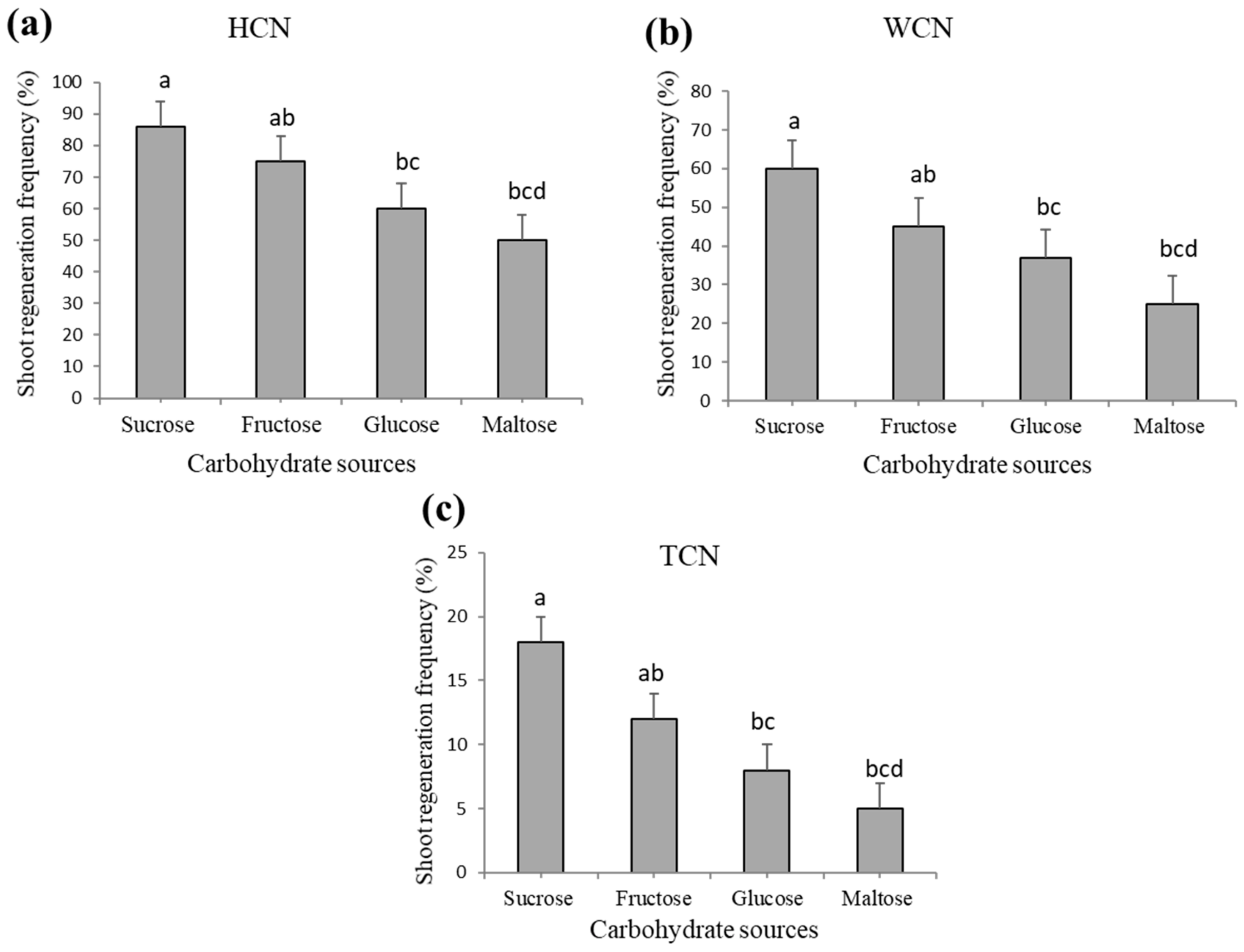
2.3. Effect of Carbohydrate Source on Shoot Regeneration
2.4. Shoot Regeneration and Propagation
2.5. Root Induction
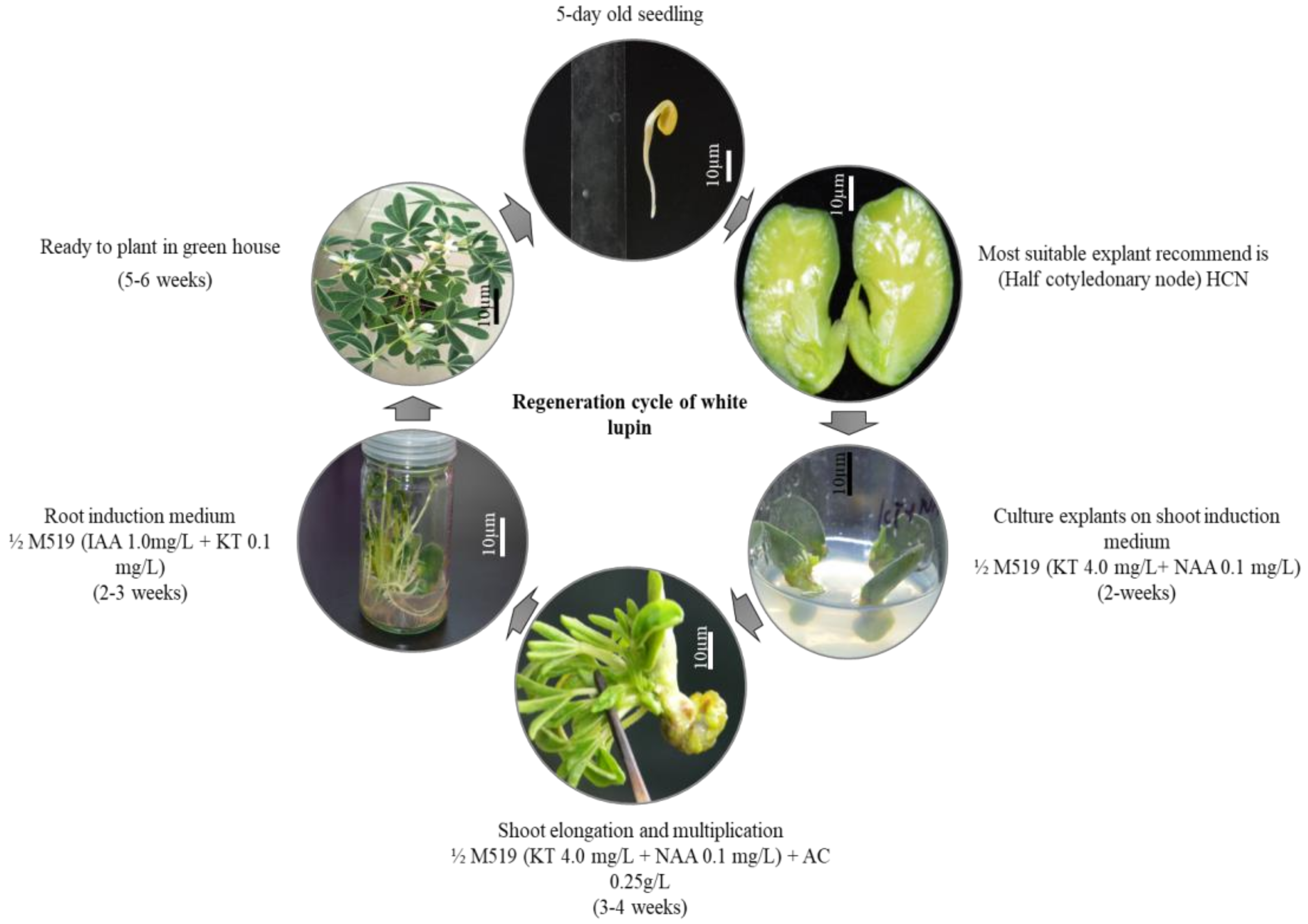
2.6. Regeneration Cycle of White Lupin
3. Discussion
4. Material and Methods
4.1. Plant Material and Seed Germination
4.2. Culture Medium and Preparation
4.3. Initiation of Shoot Induction
4.4. Shoot Induction and Elongation
4.5. Root Induction
4.6. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Bähr, M.; Fechner, A.; Hasenkopf, K.; Mittermaier, S.; Jahreis, G. Chemical composition of dehulled seeds of selected lupin cultivars in comparison to pea and soya bean. LWT Food Sci. Technol. 2014, 59, 587–590. [Google Scholar]
- Lucas, M.M.; Stoddard, F.L.; Annicchiarico, P.; Frias, J.; Martinez-Villaluenga, C.; Sussmann, D.; Duranti, M.; Seger, A.; Zander, P.M.; Pueyo, J.J. The future of lupin as a protein crop in Europe. Front. Plant Sci. 2015, 6, 705. [Google Scholar] [CrossRef]
- Cazzato, E.; Laudadio, V.; Stellacci, A.; Ceci, E.; Tufarelli, V. Influence of sulphur application on protein quality, fatty acid composition and nitrogen fixation of white lupin (Lupinus albus L.). Eur. Food Res. Technol. 2012, 235, 963–969. [Google Scholar] [CrossRef]
- Müller, J.; Gödde, V.; Niehaus, K.; Zörb, C. Metabolic adaptations of white lupin roots and shoots under phosphorus deficiency. Front. Plant Sci. 2015, 6, 1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordell, D.; White, S. Tracking phosphorus security: Indicators of phosphorus vulnerability in the global food system. Food Secur. 2015, 7, 337–350. [Google Scholar] [CrossRef]
- Tomasi, N.; Weisskopf, L.; Renella, G.; Landi, L.; Pinton, R.; Varanini, Z.; Nannipieri, P.; Torrent, J.; Martinoia, E.; Cesco, S. Flavonoids of white lupin roots participate in phosphorus mobilization from soil. Soil Biol. Biochem. 2008, 40, 1971–1974. [Google Scholar] [CrossRef]
- Marschner, P.; Neumann, G.; Kania, A.; Weiskopf, L.; Lieberei, R. Spatial and temporal dynamics of the microbial community structure in the rhizosphere of cluster roots of white lupin (Lupinus albus L.). Plant Soil 2002, 246, 167–174. [Google Scholar] [CrossRef]
- Ohtake, H.; Tsuneda, S. Phosphorus Recovery and Recycling; Springer: New York, NY, USA, 2019. [Google Scholar]
- Lambers, H.; Finnegan, P.; Jost, R.; Plaxton, W.; Shane, M.; Stitt, M. Phosphorus nutrition in Proteaceae and beyond. Nat. Plants 2015, 1, 1–9. [Google Scholar] [CrossRef]
- Wang, Z.; Straub, D.; Yang, H.; Kania, A.; Shen, J.; Ludewig, U.; Neumann, G. The regulatory network of cluster-root function and development in phosphate-deficient white lupin (Lupinus albus) identified by transcriptome sequencing. Physiol. Plant. 2014, 151, 323–338. [Google Scholar] [CrossRef]
- Manman, T.; Qian, L.; Huaqiang, T.; Yongpeng, Z.; Jia, L.; Huanxiu, L. A review of regeneration and genetic transformation in cowpea (Vigna unguiculata L. Walp). Afr. J. Agric. Res. 2013, 8, 1115–1122. [Google Scholar]
- Nguyen, A.H.; Hodgson, L.M.; Erskine, W.; Barker, S.J. An approach to overcoming regeneration recalcitrance in genetic transformation of lupins and other legumes. Plant Cell Tissue Organ Cult. (PCTOC) 2016, 127, 623–635. [Google Scholar] [CrossRef]
- Kumar, P.; Gambhir, G.; Gaur, A.; Srivastava, D. Molecular analysis of genetic stability in in vitro regenerated plants of broccoli (Brassica oleracea L. var. italica). Curr. Sci. 2015, 109, 1470–1475. [Google Scholar]
- Kumar, P.; Srivastava, D. High frequency organogenesis in hypocotyl, cotyledon, leaf and petiole explants of broccoli (Brassica oleracea L. var. italica), an important vegetable crop. Physiol. Mol. Biol. Plants 2015, 21, 279–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorpe, T.A. History of plant tissue culture. Mol. Biotechnol. 2007, 37, 169–180. [Google Scholar] [CrossRef]
- Hossain, M.A.; Hossain, M.T.; Ali, M.R.; Rahman, S.M. Effect of different carbon sources on in vitro regeneration of Indian pennywort (Centella asiatica L.); Springer: New York, NY, USA, 2005. [Google Scholar]
- Maher, M.F.; Nasti, R.A.; Vollbrecht, M.; Starker, C.G.; Clark, M.D.; Voytas, D.F. Plant gene editing through de novo induction of meristems. Nat. Biotechnol. 2020, 38, 84–89. [Google Scholar] [CrossRef]
- Lowe, K.; La Rota, M.; Hoerster, G.; Hastings, C.; Wang, N.; Chamberlin, M.; Wu, E.; Jones, T.; Gordon-Kamm, W. Rapid genotype “independent” Zea mays L. (maize) transformation via direct somatic embryogenesis. Vitr. Cell. Dev. Biol. Plant 2018, 54, 240–252. [Google Scholar] [CrossRef] [Green Version]
- Farooq, N.; Nawaz, M.A.; Mukhtar, Z.; Ali, I.; Hundleby, P.; Ahmad, N. Investigating the In Vitro Regeneration Potential of Commercial Cultivars of Brassica. Plants 2019, 8, 558. [Google Scholar] [CrossRef] [Green Version]
- Grozeva, S.; Velkov, N. In vitro plant regeneration of two cucumber (Cucumis sativum L.) genotypes: effects of explant types and culture medium. Genetika 2014, 46, 485–493. [Google Scholar] [CrossRef] [Green Version]
- Croser, J.; Lülsdorf, M.; Davies, P.; Clarke, H.; Bayliss, K.; Mallikarjuna, N.; Siddique, K. Toward doubled haploid production in the Fabaceae: Progress, constraints, and opportunities. Crit. Rev. Plant Sci. 2006, 25, 139–157. [Google Scholar] [CrossRef]
- Raza, G.; Singh, M.B.; Bhalla, P.L. In vitro plant regeneration from commercial cultivars of soybean. BioMed Res. Int. 2017, 2017, 7379693. [Google Scholar] [CrossRef] [Green Version]
- Che, P.; Chang, S.; Simon, M.K.; Zhang, Z.; Shaharyar, A.; Ourada, J.; O’Neill, D.; Torres-Mendoza, M.; Guo, Y.; Marasigan, K.M. Developing a rapid and highly efficient cowpea regeneration and transformation system using embryonic axis explants. bioRxiv 2019, 738971. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, A.; Outka, D.; Cocking, E.; Davey, M. Organogenesis and somatic embryogenesis in tissues derived from leaf protoplasts and leaf expiants of Medicago sativa. Zeitschrift Für Pflanzenphysiologie 1980, 99, 261–270. [Google Scholar] [CrossRef]
- Crocomo, O.; Sharp, W.; Peters, J. Plantlet morphogenesis and the control of callus growth and root induction of Phaseolus vulgaris with the addition of a bean seed extract. Zeitschrift Für Pflanzenphysiologienphysiol. 1976, 78, 456–460. [Google Scholar] [CrossRef]
- Bajaj, Y.; KS, L. Regeneration of plants from seedling-explants and callus cultures of Arachis hypogaea L. Indian J. Exp. Biol. 1981, 19, 1026–1029. [Google Scholar]
- Oelck, M.; Schieder, O. Genotypic differences in some legume species affecting the redifferentiation ability from callus to plants. Zeitschrift Für Pflanzenphysiologie 1983, 91, 312–321. [Google Scholar]
- Gharyal, P.; Maheshwari, S. Genetic and physiological influences on differentiation in tissue cultures of a legume, Lathyrus sativus. Theor. Appl. Genet. 1983, 66, 123–126. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, K.; Godbole, D.; Mascarenhas, A. Studies on a drought resistant legume: The moth bean, Vigna aconitifolia (Jacq) marechal. I. Protoplast culture and organogenesis. Plant Cell Rep. 1984, 3, 30–32. [Google Scholar] [CrossRef]
- Arcioni, S. Tissue culture and plant regeneration in Onobrychis viciaefolia Scop. Z. Fuer Pflanz. 1983, 90, 192–197. [Google Scholar]
- Meijer, E. High-frequency plant regeneration from hypocotyl-and leaf-derived tissue cultures of the tropical pasture legume Stylosanthes humilis. Physiol. Plant. 1982, 56, 381–385. [Google Scholar] [CrossRef]
- Nadolska-Orczyk, A. Somatic embryogenesis of agriculturally important lupin species (Lupinus angustifolius, L. albus, L. mutabilis). Plant Cell Tissue Organ Cult. 1992, 28, 19–25. [Google Scholar] [CrossRef]
- Surma, M.; Adamski, T.; Swiecicki, W.; Barzyk, P.; Kaczmarek, Z.; Kuczynska, A.; Krystkowiak, K.; Mikolajczak, K.; Ogrodowicz, P. Preliminary results of in vitro culture of pea and lupin embryos for the reduction of generation cycles in single seed descent technique. Acta Soc. Bot. Pol. 2013, 82, 231–236. [Google Scholar] [CrossRef] [Green Version]
- Święcicki, W.; Rybczyński, J.; Święcicki, W.K. Domestication and genetics of the yellow lupin (Lupinus luteus L.) and the biotechnological improvement of lupins. J. Appl. Genet. 2000, 41, 11–34. [Google Scholar]
- Bayliss, K.; Wroth, J.; Cowling, W. Pro-embryos of Lupinus albus produced from isolated microspore culture. In Proceedings of the 12th Australasian Plant Breeding Conference, Perth, Australia, 15–20 September 2002; pp. 57–61. [Google Scholar]
- Uhde-Stone, C.; Liu, J.; Zinn, K.E.; Allan, D.L.; Vance, C.P. Transgenic proteoid roots of white lupin: A vehicle for characterizing and silencing root genes involved in adaptation to P stress. Plant J. 2005, 44, 840–853. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.-M.; Xu, C.; Kim, C.-H.; Um, Y.-C.; Bah, A.A.; Guo, D.-P. Effects of explant type, culture media and growth regulators on callus induction and plant regeneration of Chinese jiaotou (Allium chinense). Sci. Hortic. 2009, 123, 124–128. [Google Scholar] [CrossRef]
- Dobránszki, J.; da Silva, J.A.T. Micropropagation of apple—a review. Biotechnol. Adv. 2010, 28, 462–488. [Google Scholar] [CrossRef]
- Lipavská, H.; Konrádová, H. Somatic embryogenesis in conifers: The role of carbohydrate metabolism. Vitr. Cell. Dev. Biol. Plant 2004, 40, 23–30. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Fuentes, S.R.; Calheiros, M.B.; Manetti-Filho, J.; Vieira, L.G. The effects of silver nitrate and different carbohydrate sources on somatic embryogenesis in Coffea canephora. Plant Cell Tissue Organ Cult. 2000, 60, 5–13. [Google Scholar] [CrossRef]
- Skrebsky, E.C.; Nicoloso, F.T.; Ferrão, G.D.E. Sacarose e período de cultivo in vitro na aclimatização ex vitro de ginseng brasileiro (Pfaffia glomerata Spreng. Pedersen). Ciência Rural 2004, 34, 1471–1477. [Google Scholar] [CrossRef] [Green Version]
- Pati, P.K.; Rath, S.P.; Sharma, M.; Sood, A.; Ahuja, P.S. In vitro propagation of rose—a review. Biotechnol. Adv. 2006, 24, 94–114. [Google Scholar] [CrossRef]
- Vasudevan, A.; Selvaraj, N.; Ganapathi, A.; Choi, C.; Manickavasagam, M.; Kasthurirengan, S. Direct plant regeneration from cucumber embryonal axis. Biol. Plant. 2007, 51, 521–524. [Google Scholar] [CrossRef]
- Selvaraj, N.; Vasudevan, A.; Manickavasagam, M.; Kasthurirengan, S.; Ganapathi, A. High frequency shoot regeneration from cotyledon explants of cucumber via organogenesis. Sci. Hortic. 2007, 112, 2–8. [Google Scholar] [CrossRef]
- Walden, R.; Wingender, R. Gene-transfer and plant-regeneration (techniques). Trends Biotechnol. 1995, 13, 324–331. [Google Scholar] [CrossRef]
- Sivanesan, I.; Kim, D.H. EFFECT OF SILICON ON BULBLET REGENERATION FROM BULBSCALE EXPLANTS OF LiLium LongifLorum THUNB. Propag. Ornam. Plants 2015, 15, 107–112. [Google Scholar]
- Née Sparrow, P.A.H.; Irwin, J.A. Brassica oleracea and B. napus. In Agrobacterium Protocols; Springer: New York, NY, USA, 2015; pp. 287–297. [Google Scholar]
- Pniewski, T.; Kapusta, J.; Legocki, A.B. In vitro micropropagation of four lupin species. Acta Physiol. Plant. 2002, 24, 417–424. [Google Scholar] [CrossRef]
- Hundleby, P.; Irwin, J.; Wang, K. Brassica oleracea and B. napus. Methods Mol. Biol 2015, 1223, 287–297. [Google Scholar]
- Paz, M.M.; Martinez, J.C.; Kalvig, A.B.; Fonger, T.M.; Wang, K. Improved cotyledonary node method using an alternative explant derived from mature seed for efficient Agrobacterium-mediated soybean transformation. Plant Cell Rep. 2006, 25, 206–213. [Google Scholar] [CrossRef]
- Shivanand, N.; Rana, M.; Yadav, R.C.; Yadav, N.R. Effect of explants and media on plant regeneration in different tomato genotypes. Indian J. Hortic. 2011, 68, 484–489. [Google Scholar]
- Daud, N.; Hasbullah, N.; Azis, N.; Rasad, F.; Amin, M.; Lassim, M. In vitro regeneration of Brassica oleracea var. capitata trough steams, roots, leaves and petioles cultures. In Proceedings of the International Conference on Agricultural, Ecological and Medical Sciences (AEMS-2015), Dubai, UAE, 23–24 December 2015; pp. 7–8. [Google Scholar]
- Gambhir, G.; Kumar, P.; Srivastava, D. High frequency regeneration of plants from cotyledon and hypocotyl cultures in Brassica oleracea cv. Pride of India. Biotechnol. Rep. 2017, 15, 107–113. [Google Scholar] [CrossRef]
- Gerszberg, A.; Hnatuszko-Konka, K.; Kowalczyk, T. In vitro regeneration of eight cultivars of Brassica oleracea var. capitata. Vitr. Cell. Dev. Biol. Plant 2015, 51, 80–87. [Google Scholar] [CrossRef] [Green Version]
- Jana, S.; Sivanesan, I.; Jeong, B.R. Effect of cytokinins on in vitro multiplication of Sophora tonkinensis. Asian Pac. J. Trop. Biomed. 2013, 3, 549–553. [Google Scholar] [CrossRef] [Green Version]
- Takatsuka, H.; Umeda, M. Hormonal control of cell division and elongation along differentiation trajectories in roots. J. Exp. Bot. 2014, 65, 2633–2643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Y.H.; Zhang, X.S. The hormonal control of regeneration in plants. In Current Topics in Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2014; Volume 108, pp. 35–69. [Google Scholar]
- Ren, X.; Liu, Y.; Jeong, B.R. Enhanced Somatic Embryo Induction of a Tree Peony, Paeonia ostii ‘Fengdan’, by a Combination of 6-benzylaminopurine (BA) and 1-naphthylacetic Acid (NAA). Plants 2020, 9, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaspar, T.; Kevers, C.; Penel, C.; Greppin, H.; Reid, D.M.; Thorpe, T.A. Plant hormones and plant growth regulators in plant tissue culture. Vitr. Cell. Dev. Biol. Plant 1996, 32, 272–289. [Google Scholar] [CrossRef]
- Gubis, J.; Lajchova, Z.; Farago, J.; Jurekova, Z. Effect of genotype and explant type on shoot regeneration in tomato (Lycopersicon esculentum) in vitro. Czech J. Genet. Plant Breed. Uzpi (Czech Republic) 2003, 39, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Bhatia, S.; Sharma, K.; Dahiya, R.; Bera, T. Modern Applications of Plant Biotechnology in Pharmaceutical Sciences; Academic Press: Cambridge, MA, USA, 2015. [Google Scholar]
- Yaseen, M.; Ahmad, T.; Sablok, G.; Standardi, A.; Hafiz, I.A. Role of carbon sources for in vitro plant growth and development. Mol. Biol. Rep. 2013, 40, 2837–2849. [Google Scholar] [CrossRef]
- Barpete, S.; Özcan, S.; Khawar, K.; Özcan, S. Effect of plant growth regulators and physical factors on in vitro high frequency regeneration of Grass pea. J. Anim. Plant Sci. 2016, 26, 1087–1093. [Google Scholar]
- Santana, J.R.F.d.; Paiva, R.; Souza, A.V.d.; Oliveira, L.M.D. Effect of different carbon sources on the in vitro multiplication of Annona sp. Ciência E Agrotecnologia 2011, 35, 487–493. [Google Scholar] [CrossRef] [Green Version]
- Begum, N.; Zenat, E.A.; Sarkar, M.K.; K Roy, C.; Munshi, J.L.; Jahan, A.; Miskat, A. Micro Propagation of Soybean () BARI-5 Variety. Open Microbiol. J. 2019, 13, 177–187. [Google Scholar] [CrossRef]
- Alam, N.; Anis, M. Influence of silver nitrate in enhancing the in vitro shoot regeneration in mucuna pruriens (L.) Dc.-Amultipurpose medicinal legume. Res. J. Life Sci. Bioinform. Pharm. Chem. Sci 2019. [Google Scholar] [CrossRef]
- Ahmad, T.; Abbasi, N.A.; Hafiz, I.A.; Ali, A. Comparison of sucrose and sorbitol as main carbon energy sources in microprogation of peach rootstock GF-677. Pak. J. Bot. 2007, 39, 1269–1275. [Google Scholar]
- Asande, L.; Indieka, A.; Adero, M.; Kiboi, S.; Amugune, N. In vitro regeneration of pigeon pea using leaf explants. Afr. Crop Sci. J. 2016, 24, 191–201. [Google Scholar] [CrossRef] [Green Version]
- Saha, P.; Afrin, M.; Mohiuddin, A.; Shohael, A. In vitro regeneration of grass pea (Lathyrus sativus L.). Jahangirnagar Univ. J. Biol. Sci. 2015, 4, 1–8. [Google Scholar] [CrossRef]
- Jo, E.-A.; Tewari, R.K.; Hahn, E.-J.; Paek, K.-Y. In vitro sucrose concentration affects growth and acclimatization of Alocasia amazonica plantlets. Plant Cell Tissue Organ Cult. (PCTOC) 2009, 96, 307. [Google Scholar] [CrossRef]
- Venkatachalam, P.; Jinu, U.; Sangeetha, P.; Geetha, N.; Sahi, S.V. High frequency plant regeneration from cotyledonary node explants of Cucumis sativus L. cultivar ‘Green Long’via adventitious shoot organogenesis and assessment of genetic fidelity by RAPD-PCR technology. 3 Biotech 2018, 8, 60. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effect of Different Concentration of Plant Growth Regulators on Shoot Regeneration (mg/L) | Length of Shoot after 14 days of Initiation | Length of Shoot after 21 days of Initiation | Length of Shoot after 28 days of Initiation | No. of Shoot Explant−1 (Mean ± SE) | Percentage of Shoot (%) | ||
|---|---|---|---|---|---|---|---|
| KT | NAA | BAP | |||||
| 1.0 | 0.1 | 0 | 2.0 cm | 4.0 cm | 4.6 cm | 9 ± 0.00 cd | 45% |
| 1.0 | 0.2 | 0 | 2.4 cm | 3.8 cm | 4.8 cm | 8 ± 0.33 de | 40% |
| 1.0 | 0.4 | 0 | 2.2 cm | 3.7 cm | 4.2 cm | 9 ± 0.33 cd | 45% |
| 2.0 | 0.1 | 0 | 2.5 cm | 3.2 cm | 4.0 cm | 12 ± 1.15 b | 60% |
| 2.0 | 0.2 | 0 | 2.1 cm | 3.5 cm | 4.5 cm | 10 ± 0.44 bc | 50% |
| 2.0 | 0.4 | 0 | 2.5 cm | 3.4 cm | 4.6 cm | 11 ± 0.33 b | 55% |
| 4.0 | 0.1 | 0 | 3.4 cm | 5.0 cm | 6.1 cm | 16 ± 0.57 a | 80% |
| 4.0 | 0.2 | 0 | 3.0 cm | 5.5 cm | 5.8 cm | 9 ± 0.57 cd | 45% |
| 4.0 | 0.4 | 0 | 2.9 cm | 5.0 cm | 5.6 cm | 10 ± 0.16 bc | 50% |
| 1.0 | 0.1 | 1.67 | 2.0 cm | 4.0 cm | 4.7 cm | 5 ± 0.72 f | 25% |
| 2.0 | 0.1 | 1.67 | 1.5 cm | 3.0 cm | 4.3 cm | 3 ± 0.57 g | 15% |
| 4.0 | 0.1 | 1.67 | 2.2 cm | 3.5 cm | 4.2 cm | 12 ± 1.15 b | 60% |
| Effect of Different Concentration of Plant Growth Regulators on Shoot Regeneration (mg/L) | No. of Explants Used | No. of Root Induced Explant−1 (Mean ± SE) | Percentage of Root Induction | ||
|---|---|---|---|---|---|
| IAA | IBA | KT | |||
| 1.0 | − | − | 15 | 9.66 ± 0.33 bc | 64% |
| 2.0 | − | − | 15 | 7.68 ± 0.33 d | 51% |
| 3.0 | − | − | 15 | 7.00 ± 0.57 de | 46% |
| − | 1.0 | 0.1 | 15 | 5.00 ± 0.00 g | 33% |
| − | 2.0 | 0.2 | 15 | 7.00 ± 0.57 de | 46% |
| − | 3.0 | 0.3 | 15 | 5.33 ± 0.33 | 33% |
| 1.0 | − | 0.1 | 15 | 12.0 ± 0.57 a | 80% |
| 2.0 | − | 0.2 | 15 | 10.0 ± 0.88 b | 66% |
| 3.0 | − | 0.3 | 15 | 10.0 ± 0.57 b | 66% |
| 1.0 | 0.5 | 0 | 15 | 6.0 ± 0.33 f | 40% |
| 2.0 | 0.5 | 0 | 15 | 9.0 ± 0.33 bc | 60% |
| 3.0 | 0.5 | 0 | 15 | 7.0 ± 0.33 de | 46% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aslam, M.M.; Karanja, J.K.; Zhang, Q.; Lin, H.; Xia, T.; Akhtar, K.; Liu, J.; Miao, R.; Xu, F.; Xu, W. In Vitro Regeneration Potential of White Lupin (Lupinus albus) from Cotyledonary Nodes. Plants 2020, 9, 318. https://doi.org/10.3390/plants9030318
Aslam MM, Karanja JK, Zhang Q, Lin H, Xia T, Akhtar K, Liu J, Miao R, Xu F, Xu W. In Vitro Regeneration Potential of White Lupin (Lupinus albus) from Cotyledonary Nodes. Plants. 2020; 9(3):318. https://doi.org/10.3390/plants9030318
Chicago/Turabian StyleAslam, Mehtab Muhammad, Joseph K. Karanja, Qian Zhang, Huifeng Lin, Tianyu Xia, Kashif Akhtar, Jianping Liu, Rui Miao, Feiyun Xu, and Weifeng Xu. 2020. "In Vitro Regeneration Potential of White Lupin (Lupinus albus) from Cotyledonary Nodes" Plants 9, no. 3: 318. https://doi.org/10.3390/plants9030318