Morphological and Molecular Characterization of Zanthoxylum zanthoxyloides (Rutaceae) from Burkina Faso


Abstract
1. Introduction
2. Materials and Methods
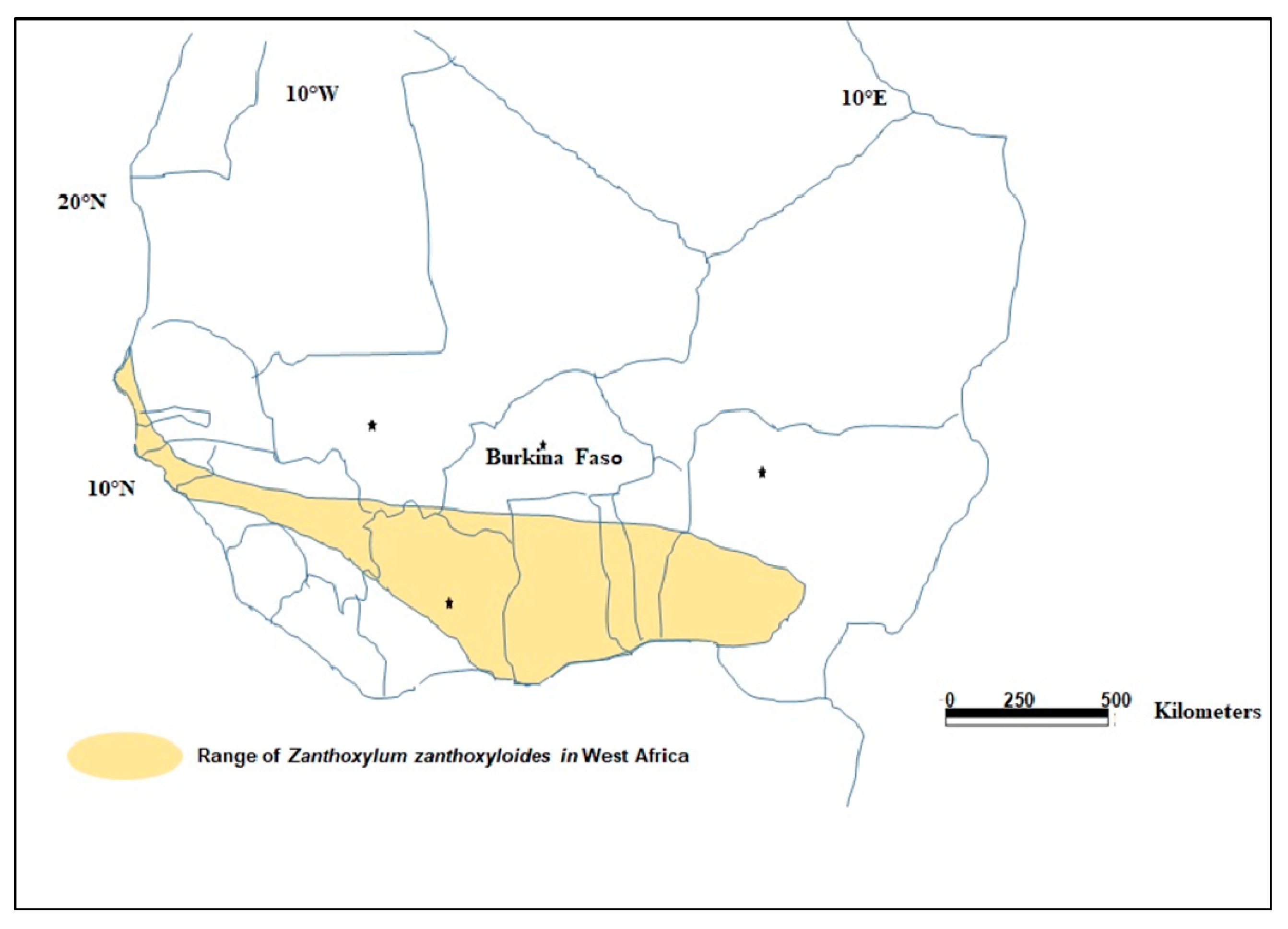
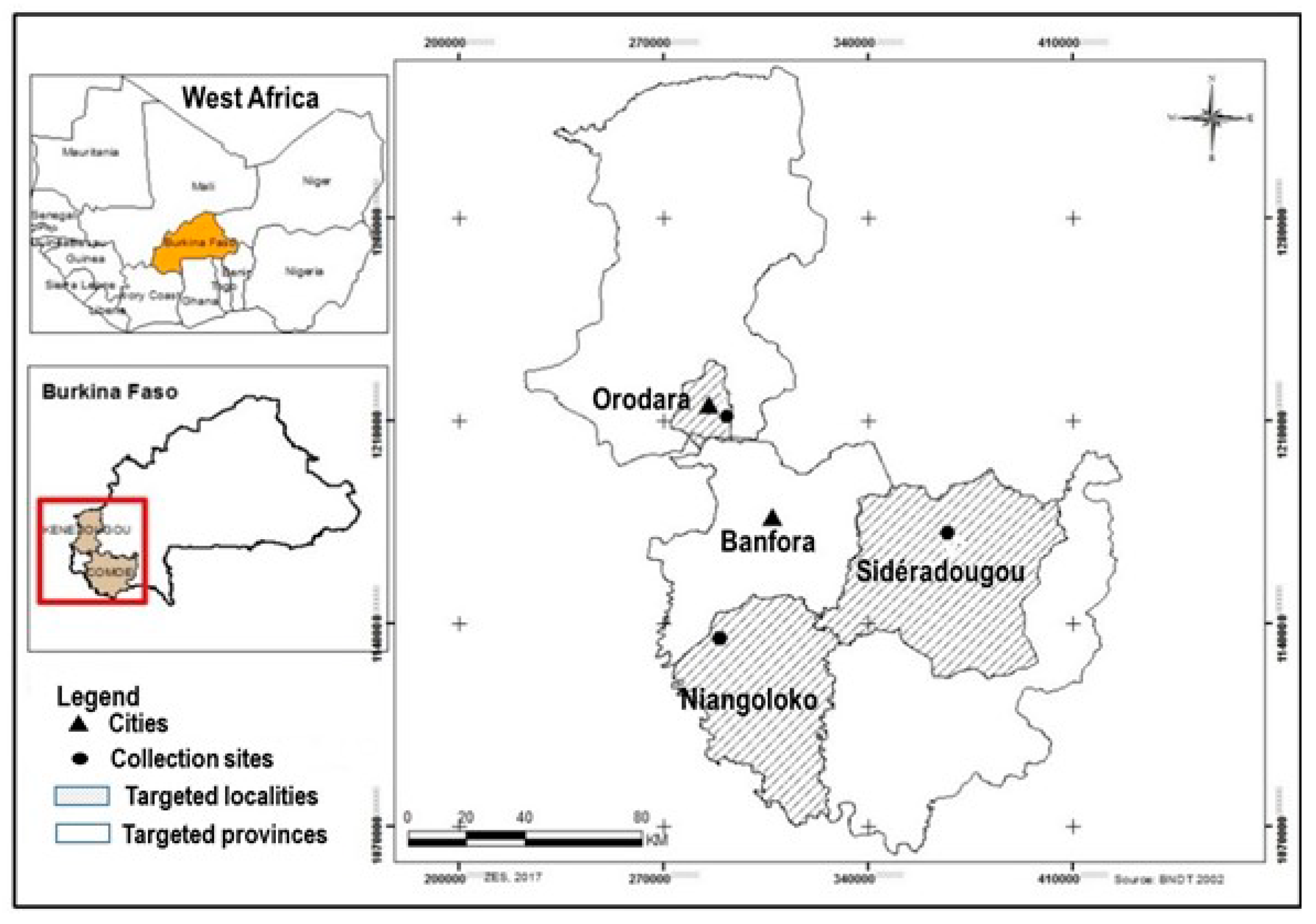
2.1. Sites and Sampling
2.2. Morphological Characterization
2.2.1. Quantitative and Qualitative Assessment
2.2.2. Morphological Data Analysis
2.3. Molecular Characterization
2.3.1. DNA Extraction and Quantification
2.3.2. Purifying the PCR Products and Sequencing
2.3.3. Editing and Alignment
3. Results
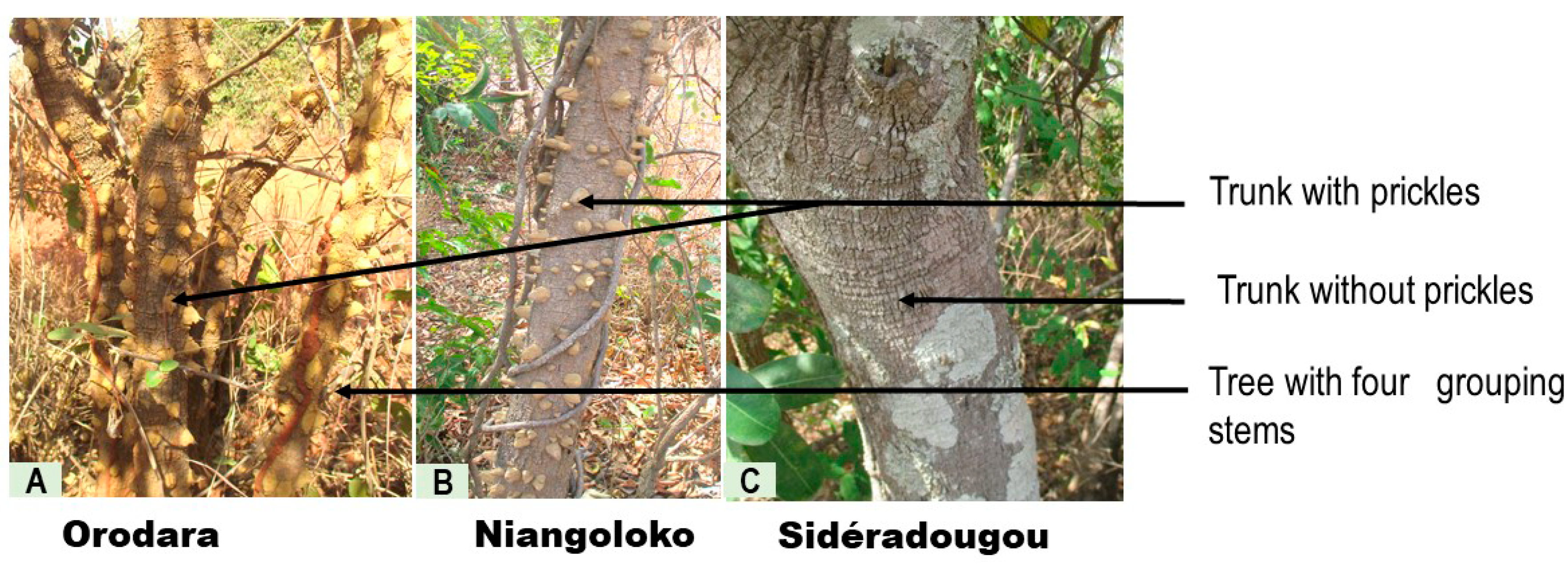
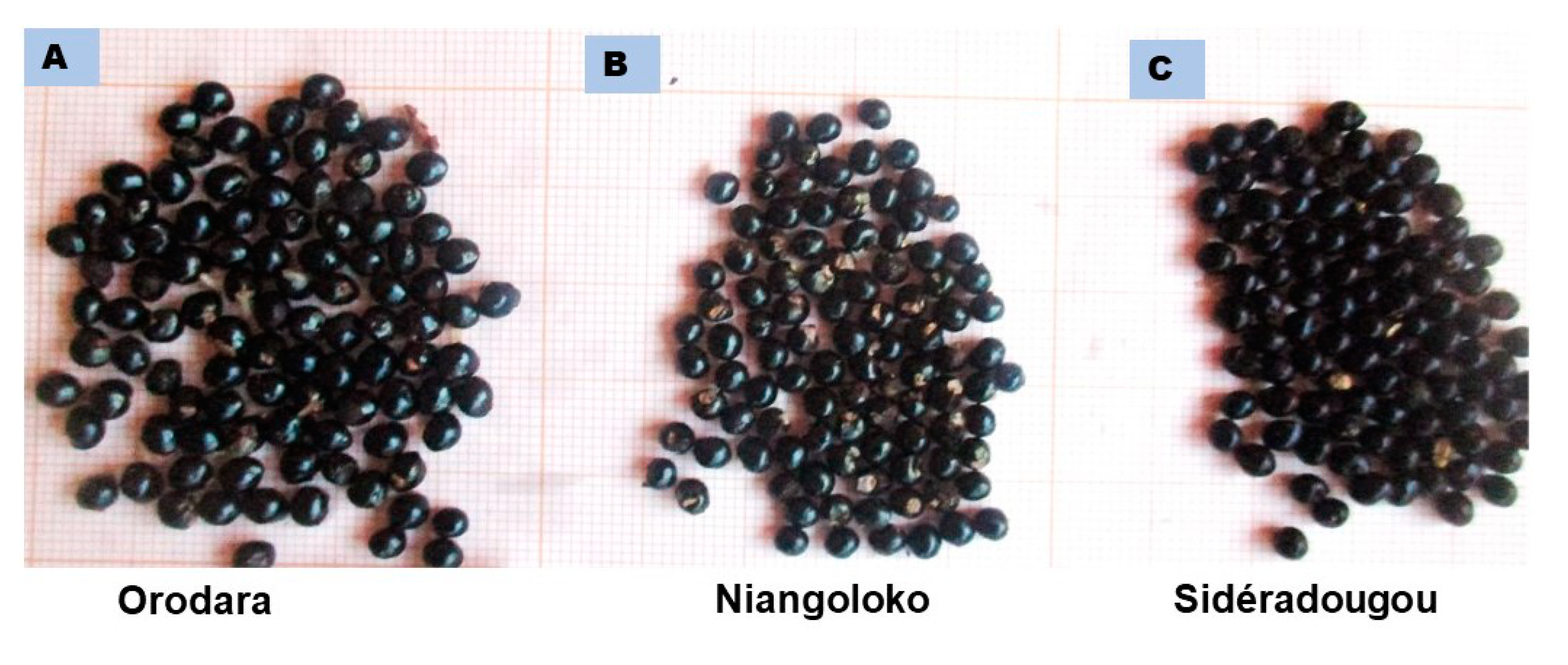
3.1. Morphological Characterization
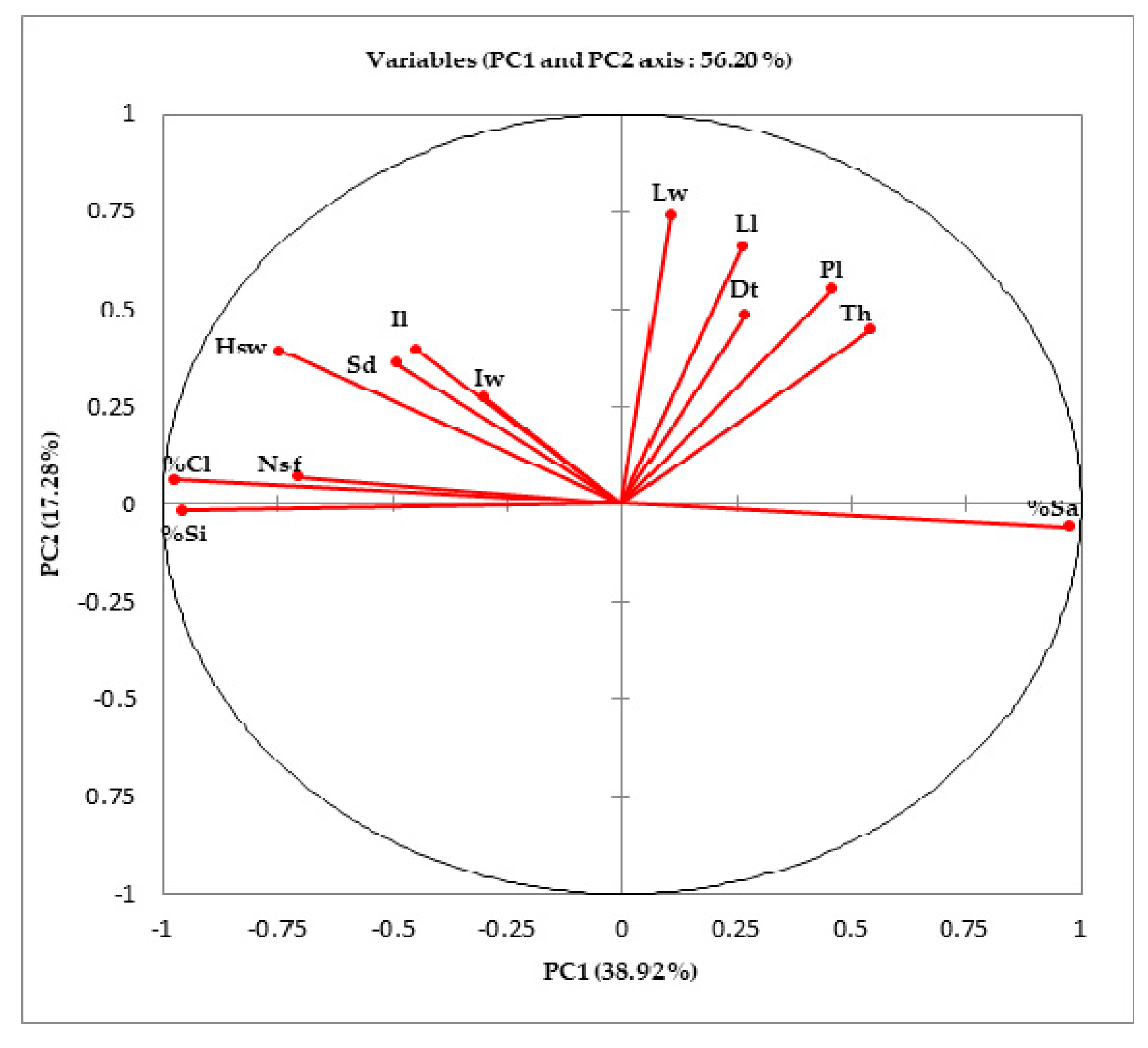
3.2. Correlation of Morphological Traits and Physical Parameters of Soil
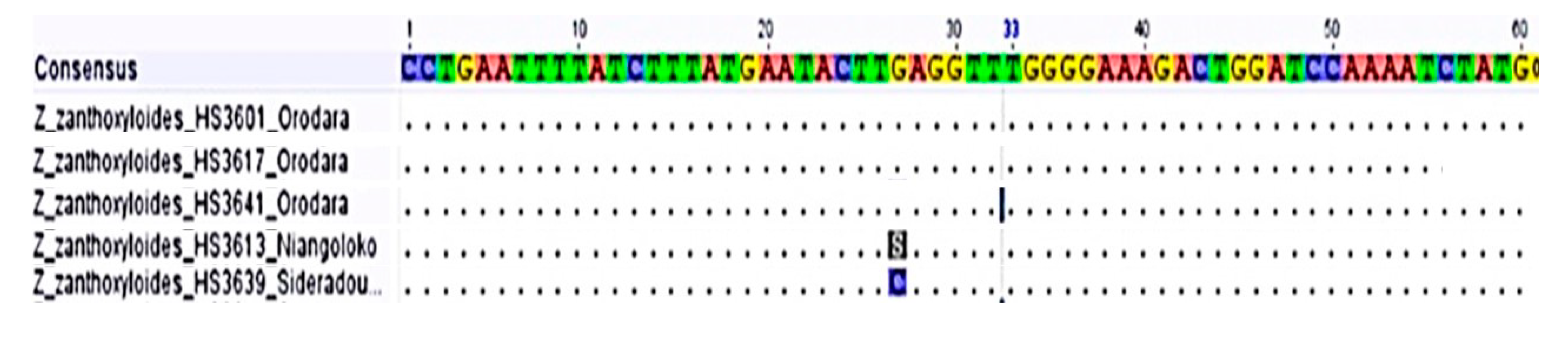
3.3. Molecular Characterization
4. Discussion
4.1. Morphological Characterization
4.2. Molecular Characterization
4.3. Conservation
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fuchs, D. Evaluation of the genetic diversity in Zanthoxylum zanthoxyloides combined with a phylogenetic and biogeographic study of the genus Zanthoxylum. Master’s Thesis, Technical University of Munich, Munich, Germany, 2018, unpublished work. [Google Scholar]
- Tan, S.; Guo, T.; Tang, X.; Song, T.; Wang, Y. Chemical constituents of Zanthoxylum armatum. II. Chem. Nat. Compd. 2018, 54, 869–870. [Google Scholar] [CrossRef]
- Perichet, C.; Philippe, F.; Dupouyet, A.; Marteaux, B.; Schnaebele, N.; Dubrulle, N.; Lavoine-Hanneguelle, S.; Giraud, N. Study of some Zanthoxylum species by chemical and DNA analysis approaches. Chem. Biodivers. 2018, 15, e1800251. [Google Scholar] [CrossRef] [PubMed]
- Arbonnier, M. Trees, Shrubs and Lianas of West African Dry Zones; Paris; Margraf Publishers: Weikersheim, Germany, 2004. [Google Scholar]
- Ouédraogo, L.; Nacoulma, A.P.; Compaoré, M.; Lagnika, L.; Kiendrebeogo, M. Stem bark of Zanthoxylum zanthoxyloïdes a possible substitute of root bark for the conservation of the species in Burkina Faso. African J. Biotechnol. 2019, 18, 197–205. [Google Scholar]
- Famobuwa, O.; Adekunbi, E.; Akinnifesi, T.; Hassan, G. In vitro Antioxidant and Anti-bacterial Properties of a Contraceptive Herbal Mixture of Zanthoxylum zanthoxyloides L. (Rutaceae), Euphorbia hirta L. (Euphorbiaceae) and Abrus precatorius L. (Leguminosae). J. Adv. Med. Pharm. Sci. 2016, 7, 1–7. [Google Scholar] [CrossRef]
- Iwu, M.M. Handbook of African Medicinal Plants; CRC Press Inc.: London, UK, 1993. [Google Scholar]
- Kakpo, S. Caractéristiques Structurales et écologiques des forêts de Bonou et d’Itchèdè au sud est du Bénin; The University of Abomey-Calavi: Cotonou, Benin, 2012. [Google Scholar]
- Bonnet, P.; Arbonnier, M. Ligneux du Sahel: Outil Graphique d’identification; Quae.: Versailles, France, 2008. [Google Scholar]
- Schmelzer, G.; Gurib-Fakim, A. Ressources Végétales de L’Afrique Tropicale 11(2). Plante Médicinale 2; Fondation PROTA: Wageningen, The Netherlands, 2013. [Google Scholar]
- Nacoulma, O.G. Plantes médicinales et Pratiques médicales traditionnelles au Burkina Faso: cas du plateau central. Ph.D. Thesis, Université de Ouagadougou, Ouagadougou, Burkina Faso, 1996. [Google Scholar]
- Guendéhou, F.; Djossa, B.A.; Kènou, C.; Assogbadjo, C. Review of Studies on Zanthoxylum zanthoxyloides (L.): availability and ethnomedical, phytochemical, pharmacological uses. Sch. J. Res. Agric. Biol. 2018, 3, 244–254. [Google Scholar]
- Nikiema, J.-B.; Ouattara, B.; Semde, R.; Djierro, K.; Compaore, M.; Guissou, I.P.; Kasilo, O.M. Promotion de la médicine traditionnelle du Burkina Faso: Essai de développement d’un médicament antidrépanocytose, le FACA. African Heal. Monit. 2010, 13, 64–67. [Google Scholar]
- Ouattara, B.; Angenot, L.; Guissou, P.; Fondu, P.; Dubois, J.; Frédérich, M.; Jansen, O.; van Heugen, J.C.; Wauters, J.N. LC/MS/NMR analysis of isomeric divanilloylquinic acids from the root bark of Fagara zanthoxyloides L. Phytochemistry 2004, 65, 1145–1151. [Google Scholar] [CrossRef] [PubMed]
- Ouattara, B.; Jansen, O.; Angenot, L.; Guissou, I.P.; Frédérich, M.; Fondu, P.; Tits, M. Antisickling properties of divanilloylquinic acids isolated from Fagara zanthoxyloides L. (Rutaceae). Phytomedecine 2009, 16, 125–129. [Google Scholar] [CrossRef]
- IUCN. The IUCN Red List of Threatened Species. Version 2019-1. 2019. Available online: http://www.iucnredlist.org (accessed on 16 July 2019).
- Compaoré, M.; Roland, M.N.T.; Zerbo, P.; Karama, I.; Traoré, O.; Méda, A.L.; Kiendrebeogo, M.; Novak, J. Availability evaluation of twelve antimalarial medicinal plants from western regions of Burkina Faso. J. Dis. Med. Plants 2018, 4, 80–88. [Google Scholar]
- Etsè, K.D.; Aïdam, A.V.; de Souza, C.; Crêche, J.; Lanoue, A. In vitro propagation of Zanthoxylum zanthoxyloides L., an endangered African medicinal plant. Acta Bot. Gall. 2011, 158, 47–55. [Google Scholar] [CrossRef]
- Traoré, L.; Ouédraogo, A.; Thombiano, A. Perceptions, usages et vulnérabilité des ressources végétales ligneuses dans le Sud-Ouest du Burkina Faso. Int. J. Biol. Chem. Sc. 2011, 5, 258–278. [Google Scholar] [CrossRef]
- Mansfelds, R. Mansfeld’s Encyclopedia of Agricultural Crops: (Except Ornamentals); Springer Sciences & Business Media: New York, NY, USA, 2001; Volume 1. [Google Scholar]
- Neya, T.; Daboue, E.; Neya, O.; Ouedraogo, I.; Sena, K.Y. Tolerance à la dessiccation des semences de Parinari curatellifolia planch. ex benth, Vitex doniana Sweet et Zanthoxylum zanthoxyloides (L.) Watermann au Burkina Faso. Int. J. Biol. Chem. Sci. 2017, 11, 2730–2744. [Google Scholar] [CrossRef][Green Version]
- Ouôba, P.; Boussim, J.; Guinko, S. Le potentiel fruitier de la forêt classée de Niangoloko au Burkina Faso. Fruits 2005, 61, 71–81. [Google Scholar] [CrossRef]
- Ouôba, P. Fonctions écologiques et potentialités agroforestières de Maranthes polyandra [(Benth.)Prance] dans la Forêt Classée de Niangoloko. Bachelor’s Thesis, Polytechnic University of Bobo-Dioulasso, Bobo-Dioulasso, Burkina Faso, 2016. [Google Scholar]
- Chehade, A.; Chalak, L.; Elbitar, A.; Cosson, P.; Zanetto, A.; Dirlewanger, E. Caractérisation préliminaire morphologique et moléculaire de clones de cerisier cultivés au Liban (Prunus avium L.). Leban. Sci. J. 2005, 6, 29–40. [Google Scholar]
- Eyog-Matig, O.; Gaoué, O.G.; Obel-Lawson, E. (Eds.) Development of Appropriate Conservation Strategies for African Forest Trees Identified as Priority Species by SAFORGEN Member Countries; International Plant Genetic Resources Institute: Nairobi, Kenya, 2002. [Google Scholar]
- Appelhans, M.S.; Reichelt, N.; Groppo, M.; Paetzold, C.; Wen, J. Phylogeny and biogeography of the pantropical genus Zanthoxylum and its closest relatives in the proto-Rutaceae group (Rutaceae). Mol. Phylogenet. Evol. 2018, 126, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Shin, J.; Cho, S.; Hwang, Y.; Choi, C. Species-specific InDel markers for authentication of the Korean herbs Zanthoxylum schinifolium and Zanthoxylum piperitum. BioRxiv 2019. [Google Scholar] [CrossRef]
- Kamiya, K.; Moritsuka, E.; Yoshida, T.; Yahara, T.; Tachida, H. High population differentiation and unusual haplotype structure in a shade-intolerant pioneer tree species, Zanthoxylum ailanthoides (Rutaceae) revealed by analysis of DNA polymorphism at four nuclear loci. Mol. Ecol. 2008, 17, 2329–2338. [Google Scholar] [CrossRef]
- Taberlet, P.; Gielly, L.; Pautou, G.; Bouvet, J. Universal primers for amplification of three non-coding regions of chloroplast DNA. Plant Mol. Biol. 1991, 17, 11051109. [Google Scholar] [CrossRef]
- Balthazar, M.V.; Endress, P.K.; Qiu, L.-Y. Phylogenetic relationships in Buxaceae based on nuclear internal transcribed spacers and plastid ndhF sequences. Int. J. Plant Sci. 2000, 161, 785–792. [Google Scholar] [CrossRef]
- Sang, T.; Crawford, D.J.; Stuessy, T.F. Chloroplast DNA phylogeny reticulate evolution, and Biogeography of Paeonia ( Paeoniaceae). Am. J. Bot. 1997, 84, 1120–1136. [Google Scholar] [CrossRef]
- Tate, J.A.; Simpson, B.B. Paraphyly of Tarasa (Malvaceae) and diverse origins of the polyploid species. Syst. Bot. 2003, 28, 723–737. [Google Scholar]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M.W.; Caro, T.M. Effect of selective logging on tree and understory regeneration in miombo woodland in western Tanzania. Afr. J. Ecol. 2003, 41, 75–82. [Google Scholar] [CrossRef]
- Eyog Matig, O.; Ndoye, O.; Kengue, J.; Awono, A. Les Fruitiers Forestiers Comestibles du Cameroun; IPGRI: Rome, Italy, 2006. [Google Scholar]
- Yang, Y.; Watanabe, M.; Li, F.; Zhang, J.; Zhang, W.; Zhai, J. Factors affecting forest growth and possible effects of climate change in the Taihang Mountains, northern China. Forestry 2006, 79, 135–147. [Google Scholar] [CrossRef]
- Gupta, B.S.; Jelle, B.P.; Hovde, P.J.; Gao, T. Wood facade materials ageing analysis by FTIR spectroscopy. Constr. Mater. 2015, 168, 219–231. [Google Scholar] [CrossRef]
- Medhi, K.; Sarmah, D.; Deka, M.; Bhau, B. High gene flow and genetic diversity in three economically important Zanthoxylum spp. of Upper Brahmaputra Valley Zone of NE India using molecular markers. Meta Gene 2014, 2, 706–721. [Google Scholar] [CrossRef] [PubMed]
- Jump, A.S.; Carr, M.; Ahrends, A.; Marchant, R. Genetic divergence during long-term isolation in highly diverse populations of tropical trees across the Eastern Arc mountains of Tanzania. Biotropica 2014, 46, 565–574. [Google Scholar] [CrossRef]
- Sam, J.B. Etude de L’effet de Prétraitement sur la Germination et la Croissance de Zanthoxylum Zanthoxyloides (L.) Zepernick & Timler de Quatre Provenances en Pépinère; École nationale des eaux et forêts: Bobo-Dioulasso, Burkina Faso, 2017. [Google Scholar]
- FAO. Conservation ex situ de Pollen et de Graines, et de Cultures in vitro de Plantes Ligneuses Perennes; Etude FAO: Rome, Italy, 1994. [Google Scholar]
- Douh, C. Rôle de la banque de graines du sol dans la régénération des forêts denses humides d’Afrique centrale. Ph.D. Thesis, Université de Liège, Liège, Belgium, 2018. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Geographic Coordinates (Recorded with GPS) | Altitude in m a.s.l. (Recorded with GPS) | Average Annual Precipitation from 2016–2017 (mm) (from the National Agency of Meteorology, Burkina Faso) | Average Annual Temperature 2017 (°C) from (mm) (from the National Agency of Meteorology, Burkina Faso) | Soil Type from [5] |
|---|---|---|---|---|---|
| Niangoloko | 30 P 0289150 UTM 1134950 | 342–353 | 979.55 | 26–31 | sand |
| Sidéra dougou | 30P 0368450 UTM 1169500 | 320–343 | 771.5 | 26–31 | sand |
| Orodara | 30 P 0291500 UTM 1217000 | 546–560 | 980.3 | 26–32 | clay |
| Site | Depth (cm) | % Clay | % Silt | % Sand |
|---|---|---|---|---|
| Niangoloko | 0–25 | 6.1 | 23.49 | 70.41 |
| 25.1–50 | 15.2 | 25 | 59.8 | |
| 50.1–100 | 23.53 | 19.61 | 56.86 | |
| Sidéradougou | 0–25 | 10.2 | 22.7 | 67.1 |
| 25.1–50 | 16.4 | 25.2 | 58.4 | |
| 50.1–100 | 26.4 | 19.5 | 54.1 | |
| Orodara | 0–25 | 63 | 25.8 | 11.2 |
| 25.1–50 | 62.75 | 27.45 | 9.8 | |
| 50.1–100 | 49.6 | 19.61 | 30.79 |
| Primers | Sequence (All Given in Orientation 5’-3’) | References |
|---|---|---|
| trnLc | CGA AAT CGG TAG ACG CTA CG | [29] |
| trnLd | GGG GAT AGA GGG ACT TGA AC | [29] |
| trnLFe | GGT TCA AGT CCC TCT ATC CC | [29] |
| trnLFf | ATT TGA ACT GGT GAC ACG AG | [29] |
| ITS-F1 | CCT GCC CTT TGT ACA CAC C | [30] |
| ITS-F2 | TCT CGG CAA CGG ATA TCT CG | [30] |
| ITS-R1 | GCT TCT NCA GAC TAC AAT TC | [30] |
| ITS-R2 | CGT TCA AAG ACT CGA TGG TTC | [30] |
| psbA | GTT ATG CAT GAA CGT AAT GCT C | [31] |
| trnH | CGC GCA TGG TGG ATT CAC AAT CC | [32] |
| GBSSI f156 | GCT CCT CGC TAT GAC CAG TA | [28] |
| GBSSI r642 | ACT CAA CAC CTT TAT CTT CC | [28] |
| Site | Trunk Height (cm) | Diameter of Trunk C50 | Number of Stems Per Feet |
|---|---|---|---|
| Sidéradougou (24) | 203.7 ± 57.61 a | 60.67 ±13,40 a | 1.00 ± 0.000 b |
| Niangoloko (24) | 151.2 ± 50.58 b | 45.63 ± 27.5 b | 1.25 ± 0.53 b |
| Orodara (24) | 106.5 ± 50.77 c | 42.04 ± 21.75 b | 2.71 ± 1.3 a |
| Min | 27 | 18 | 1 |
| Max | 300 | 133 | 5 |
| Mean | 153.8 | 49.44 | 1.65 |
| SqR | 65.87 | 22.76 | 1.10 |
| P-Value | <0.0001 | 0.009 | <0.0001 |
| Significance | HS | S | HS |
| Site | Leaf Length (cm) | Leaf Width (cm) | Petiole Length (cm) | Inflorescence Length (cm) | Inflorescence Width (cm) | Seed Diameter (mm) | Hundred Seed Weight (g) |
|---|---|---|---|---|---|---|---|
| Sidéradougou (24) | 18.92 ± 4.11 a | 11.4 ± 2.69 a | 4.27 ± 1.01 a | 8.37 ± 2.2 a | 2.91 ± 0.88 a | 3.3 ± 0.28 a | 2.21 ± 0.32 b |
| Niangoloko (24) | 17.02 ± 2.89 ab | 11.2 ± 2.39 a | 4.25 ± 1.15 a | 5.37 ± 1.68 b | 2.63 ± 0.51 a | 3.05 ± 0.07b | 2.01 ± 0.09 c |
| Orodara (24) | 16.48 ± 2.77 b | 11,08 ± 1.76 a | 3.48 ± 0.71 b | 8.83 ± 2.3 a | 3.16 ± 0.88 a | 3.42 ± 0.32 a | 2.78 ± 0.34 a |
| Min | 10 | 7 | 2 | 3 | 2 | 3 | 1.8 |
| Max | 25 | 18 | 6 | 13 | 6 | 4 | 3.5 |
| Mean | 17.47 | 11.22 | 4 | 7.55 | 2.9 | 3.25 | 2.34 |
| SqR | 3.43 | 2.26 | 1.03 | 2.57 | 0.8 | 0.29 | 0.43 |
| P-Value | 0.03 | 0.891 | 0.008 | <0.0001 | 0.074 | <0.0001 | <0.0001 |
| Significance | S | NS | S | HS | NS | HS | HS |
| Site | Flowering Time | Fruiting Time | ||||
|---|---|---|---|---|---|---|
| May–July | August–October | Novemer–Jane | July–September | October–December | Jane–March | |
| Sidéradougou | 4 | 12 | 8 | 4 | 12 | 8 |
| Niangoloko | 9 | 12 | 3 | 9 | 12 | 3 |
| Orodara | 2 | 6 | 16 | 6 | 6 | 16 |
| Total number of trees per season | 15 | 30 | 27 | 15 | 30 | 27 |
| Percentage (%) | 20.83 | 41.67 | 37.5 | 20.83 | 41.67 | 37.5 |
| Variables | Th | Dt | Nsf | Lf | Lw | Pl | Hsw | Sd | Il | Iw | %Cl | %Si | Sa |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Th | 1 | ||||||||||||
| Dt | 0.3107 | 1 | |||||||||||
| Nsf | −0.2741 | −0.2396 | 1 | ||||||||||
| Ll | 0.2419 | 0.1755 | −0.0565 | 1 | |||||||||
| Lw | 0.1449 | 0.2412 | 0.0370 | 0.6249 | 1 | ||||||||
| Pl | 0.5091 | 0.2148 | −0.1982 | 0.3706 | 0.4132 | 1 | |||||||
| Hsw | −0.1665 | −0.0405 | 0.4180 | 0.0388 | 0.1620 | −0.0838 | 1 | ||||||
| Sd | 0.0430 | −0.0045 | 0.3004 | 0.0023 | 0.0382 | −0.1016 | 0.4733 | 1 | |||||
| Il | 0.0082 | 0.1938 | 0.1903 | −0.0373 | 0.0203 | −0.1489 | 0.4015 | 0.4079 | 1 | ||||
| Iw | −0.0919 | 0.1267 | 0.1041 | −0.0564 | 0.0642 | −0.0684 | 0.1733 | 0.1659 | 0.4707 | 1 | |||
| %Cl | −0.4925 | −0.2164 | 0.6756 | −0.1931 | −0.0416 | −0.3596 | 0.7539 | 0.4298 | 0.3880 | 0.2419 | 1 | ||
| %Si | −0.5541 | −0.2694 | 0.6880 | −0.2372 | −0.0488 | −0.3544 | 0.7073 | 0.3541 | 0.2869 | 0.2096 | 0.9793 | 1 | |
| %Sa | 0.4943 | 0.2179 | −0.6762 | 0.1943 | 0.0418 | 0.3596 | −0.7531 | −0.4281 | −0.3856 | −0.2412 | −1.0000 | −0.9804 | 1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ouédraogo, L.; Fuchs, D.; Schaefer, H.; Kiendrebeogo, M. Morphological and Molecular Characterization of Zanthoxylum zanthoxyloides (Rutaceae) from Burkina Faso. Plants 2019, 8, 353. https://doi.org/10.3390/plants8090353
Ouédraogo L, Fuchs D, Schaefer H, Kiendrebeogo M. Morphological and Molecular Characterization of Zanthoxylum zanthoxyloides (Rutaceae) from Burkina Faso. Plants. 2019; 8(9):353. https://doi.org/10.3390/plants8090353
Chicago/Turabian StyleOuédraogo, Lassané, Dominik Fuchs, Hanno Schaefer, and Martin Kiendrebeogo. 2019. "Morphological and Molecular Characterization of Zanthoxylum zanthoxyloides (Rutaceae) from Burkina Faso" Plants 8, no. 9: 353. https://doi.org/10.3390/plants8090353
APA StyleOuédraogo, L., Fuchs, D., Schaefer, H., & Kiendrebeogo, M. (2019). Morphological and Molecular Characterization of Zanthoxylum zanthoxyloides (Rutaceae) from Burkina Faso. Plants, 8(9), 353. https://doi.org/10.3390/plants8090353