Herbicide Resistance Traits in Maize and Soybean: Current Status and Future Outlook


Abstract
:1. Introduction
2. Early Efforts
3. Current Transgenic HRCs
4. Maize
5. Soybean
6. Future HRCs and Related Technologies
7. Worldwide Use of Transgenic Crops
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Crop Loss. Available online: http://wssa.net/wssa/weed/croploss-2/ (accessed on 9 August 2019).
- Timmons, F.L. A history of weed control in the United States and Canada. Weed Sci. 2005, 53, 748–761. [Google Scholar] [CrossRef]
- Marrone, P.G. Pesticidal natural products—Status and future potential. Pest Manag. Sci. 2019. [Google Scholar] [CrossRef] [PubMed]
- Duke, S.O. Taking stock of herbicide-resistant crops ten years after introduction. Pest Manag. Sci. 2005, 61, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Green, J.M.; Castle, L.A. Transitioning from single to multiple herbicide-resistant crops. In Glyphosate Resistance in Crops and Weeds: History, Development, and Management; Nandula, V.K., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2010; pp. 67–92. [Google Scholar]
- Green, J.M. Evolution of glyphosate-resistant crop technology. Weed Sci. 2009, 57, 108–117. [Google Scholar] [CrossRef]
- Duke, S.O.; Powles, S.B. Glyphosate: A once-in-a-century herbicide. Pest Manag. Sci. 2008, 64, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Feng, P.C.C.; CaJacob, C.A.; Martino-Catt, S.J.; Cerny, R.E.; Elmore, G.A.; Heck, G.R.; Huang, J.; Kruger, W.M.; Malven, M.; Miklos, J.A.; et al. Glyphosate-resistant crops: Developing the next generation products. In Glyphosate Resistance in Crops and Weeds: History, Development, and Management; Nandula, V.K., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2010; pp. 45–66. [Google Scholar]
- Herouet, C.; Esdaile, D.J.; Mallyon, B.A.; Debruyne, E.; Schulz, A.; Currier, T.; Hendicks, K.; van der Klis, R.J.; Rouan, D. Safety evaluation of the phosphinothricin acetyltransferase proteins encoded by the pat and bar sequences that confer tolerance to glufosinate-ammonium herbicide in transgenic plants. Regul. Toxicol. Pharmacol. 2005, 41, 134–149. [Google Scholar] [CrossRef] [PubMed]
- Wright, T.R.; Lira, J.M.; Merlo, D.J.; Hopkins, N. Novel Herbicide Resistance Genes. World Intellectual Property Organization Patent WO2005107437A2, 17 November 2005. [Google Scholar]
- Wright, T.R.; Lira, J.M.; Walsh, T.A.; Merlo, D.J.; Jayakumar, P.S.; Lin, G. Novel Herbicide Resistance Genes. World Intellectual Property Organization Patent WO/2007/053482, 5 October 2007. [Google Scholar]
- Picking Beans: A Look at the Many 2019 Herbicide-Tolerant Soybean Options. Available online: https://www.dtnpf.com/agriculture/web/ag/crops/article/2018/10/02/look-many-2019-herbicide-tolerant (accessed on 9 August 2019).
- Behrens, M.R.; Mutlu, N.; Chakraborty, S.; Dumitru, R.; Jiang, W.Z.; Lavallee, B.J.; Herman, P.L.; Clemente, T.E.; Weeks, D.P. Dicamba resistance: Enlarging and preserving biotechnology-based weed management strategies. Science 2007, 316, 1185–1188. [Google Scholar] [CrossRef] [PubMed]
- A Final Report on Dicamba-Injured Soybean Acres. Available online: https://ipm.missouri.edu/IPCM/2017/10/final_report_dicamba_injured_soybean/ (accessed on 9 August 2019).
- Lyon, B.R.; Cousins, Y.L.; Llewellyn, D.J.; Dennis, E.S. Cotton plants transformed with a bacterial degradation gene are protected from accidental spray drift damage by the herbicide 2,4-dichlorophenoxyacetic acid. Transgenic Res. 1993, 3, 162–169. [Google Scholar] [CrossRef]
- Streber, W.R.; Willmitzer, L. Transgenic tobacco expressing a bacterial etoxifying enzyme are resistant to 2,4-D. Bio/technology 1989, 8, 811–816. [Google Scholar]
- Laurent, F.; Debrauwer, L.; Rathahao, E.; Scalla, R. 2,4-Dichlorophenoxyacetic acid metabolism in transgenic tolerant cotton (Gossypium hirsutum). J. Agric. Food Chem. 2000, 48, 5307–5311. [Google Scholar] [CrossRef] [PubMed]
- Skirvin, R.M.; Norton, M.A.; Farrand, S.K.; Mulwa, R.M.S. Grape plant named “Improved Chancellor”. U.S. Patent Application USPP20428P3, 20 October 2009. [Google Scholar]
- Simpson, D.M.; Wright, T.R.; Chambers, R.S.; Peterson, M.A.; Cui, C.; Robinson, A.E.; Richburg, J.S.; Ruen, D.C.; Ferguson, S.; Maddy, B.E. Introduction to Dow AgroSciences herbicide tolerance traits. In Proceedings of the 2008 Annual Meeting of the Weed Science Society of America, Chicago, IL, USA, 4–7 February 2008; Volume 48, p. 115. [Google Scholar]
- Enlist Weed Control System. Available online: https://www.enlist.com/en/news.html (accessed on 9 August 2019).
- Corn, Soybean Trait Options for 2019 and Beyond. Available online: https://farmweeknow.com/blogs-corn-soybean-trait-options-2019-beyond-19929 (accessed on 9 August 2019).
- More Dicamba to Come? Dicamba-Stacked Corn on the Horizon. Available online: https://www.dtnpf.com/agriculture/web/ag/crops/article/2019/03/21/dicamba-stacked-corn-horizon (accessed on 5 September 2019).
- FMC to Introduce Two New Herbicide Modes of Action in Next Decade. Available online: https://www.agprofessional.com/article/fmc-introduce-two-new-herbicide-modes-action-next-decade (accessed on 12 August 2019).
- New Corn and Soybean Herbicide Site of Action May be Coming in Late 2020s. Available online: https://www.agriculture.com/crops/new-corn-and-soybean-herbicide-site-of-action-may-be-coming-in-late-2020s (accessed on 12 August 2019).
- Biotech Crop Adoption Surges as Economic Benefits Accumulate in 22 Years. Available online: http://www.isaaa.org/resources/publications/briefs/53/executivesummary/default.asp (accessed on 12 August 2019).
- Palmer Pigweed Resistance to 2,4-D and Dicamba Confirmed in Kansas. Available online: https://web.archive.org/web/20190905153145/https://agfaxweedsolutions.com/2019/03/05/palmer-pigweed-resistant-to-24-d-and-dicamba-confirmed-in-kansas/ (accessed on 5 September 2019).
- International Survey of Herbicide Resistant Weeds. Available online: https://web.archive.org/web/20190905154516/http://weedscience.com/Summary/MOA.aspx?MOAID=24 (accessed on 5 September 2019).
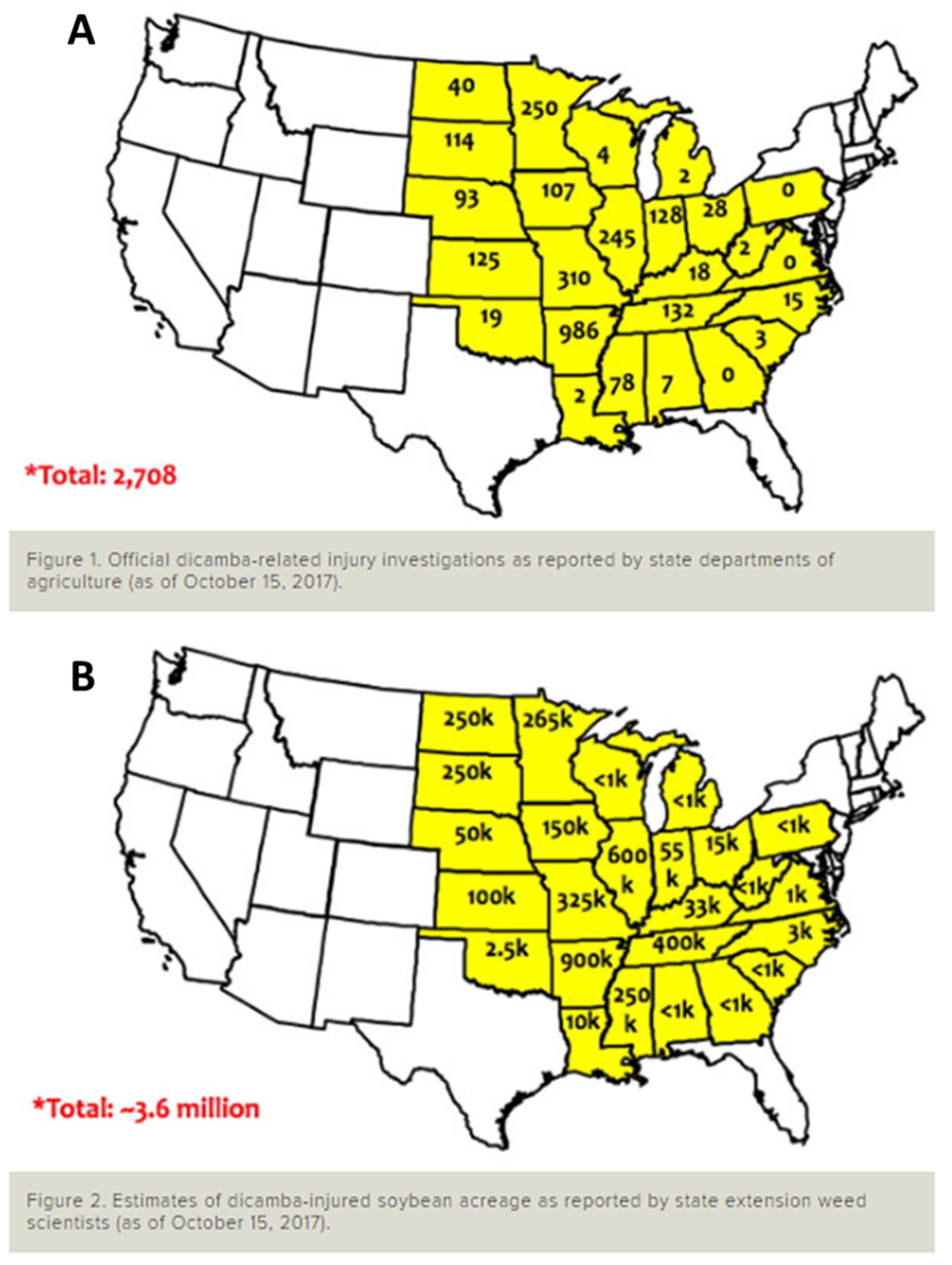

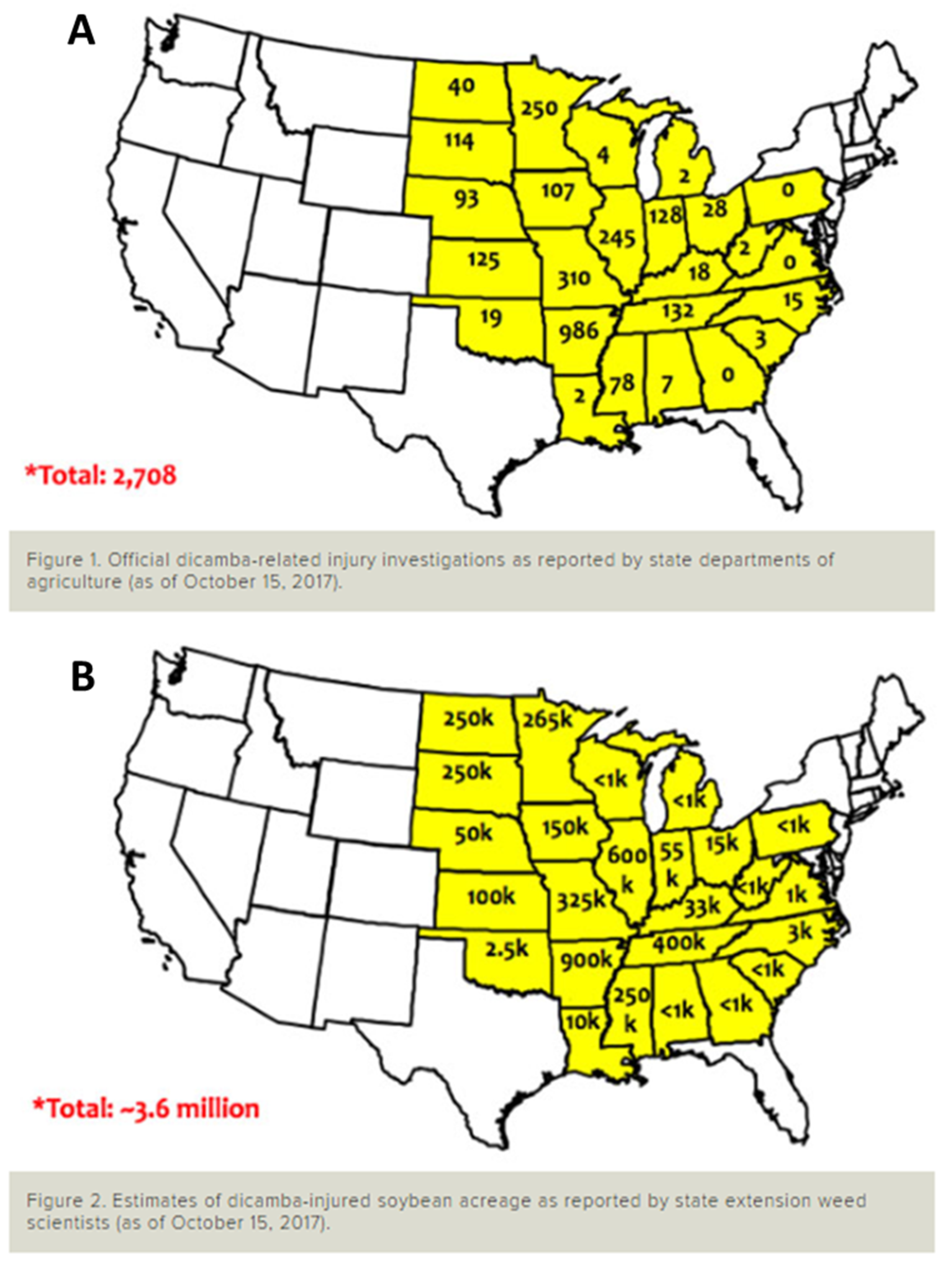{kind=link}
| Selection Method | Herbicide Family | Crop | Year of Disclosure |
|---|---|---|---|
| Seed mutagenesis | Sulfonylurea | Soybean | 1987 |
| Pollen mutagenesis | Imidazolinone | Maize | 1992 |
| Tissue culture | ACCase inhibitor | Maize | 1992 |
| Imidazolinone | Maize | 1991 | |
| Triazine | Soybean | 1996 |
| Crop | Resistance Trait | Trait Gene | Trait Designation | First Sales | Trade Name |
|---|---|---|---|---|---|
| Maize | Glyphosate | Three modified maize epsps | GA21 | 1998 | Roundup Ready® |
| Two cp4 epsps | NK603 | 2001 | Roundup Ready® 2 | ||
| Glufosinate | pat | T14, T25 | 1996 | LibertyLink System® | |
| 2,4-D | tfdA | DHT1 | 2019 | Enlist™ Weed Control System | |
| AOPP | aad | DHT1 | 2019 | Enlist™ Weed Control System | |
| Soybean | Glyphosate | cp4 epsps | GTS 40-3-2 | 1996 | Roundup Ready® |
| cp4 epsps | MON89788 | 2009 | Roundup Ready® 2 Yield | ||
| Glufosinate | pat | A2704-12 | 2009 | LibertyLink System® | |
| Dicamba | dmo | MON87708 | 2017 | Roundup Ready 2 Xtend® | |
| 2,4-D | tfdA | DHT2 | 2019 | Enlist™ Weed Control System |
| Event | Crop | # of approvals |
|---|---|---|
| NK603 | Maize | 55 approvals in 26 non-EU countries plus 28 EU countries |
| GTS 40-3-2 | Soybean | 54 approvals in 27 non-EU countries plus 28 EU countries |
| GA21 | Maize | 50 approvals in 24 non-EU countries plus 28 EU countries |
| A2704-12 | Soybean | 43 approvals in 23 non-EU countries plus 28 EU countries |
| T25 | Maize | 41 approvals in 20 non-EU countries plus 28 EU countries |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nandula, V.K. Herbicide Resistance Traits in Maize and Soybean: Current Status and Future Outlook. Plants 2019, 8, 337. https://doi.org/10.3390/plants8090337
Nandula VK. Herbicide Resistance Traits in Maize and Soybean: Current Status and Future Outlook. Plants. 2019; 8(9):337. https://doi.org/10.3390/plants8090337
Chicago/Turabian StyleNandula, Vijay K. 2019. "Herbicide Resistance Traits in Maize and Soybean: Current Status and Future Outlook" Plants 8, no. 9: 337. https://doi.org/10.3390/plants8090337
APA StyleNandula, V. K. (2019). Herbicide Resistance Traits in Maize and Soybean: Current Status and Future Outlook. Plants, 8(9), 337. https://doi.org/10.3390/plants8090337