Genetic, Morphological, and Biochemical Diversity of Argan Tree (Argania spinosa L.) (Sapotaceae) in Tunisia

 , ,
, ,
Abstract
1. Introduction
2. Results
2.1. Morphological Analysis of Trees and Fruits
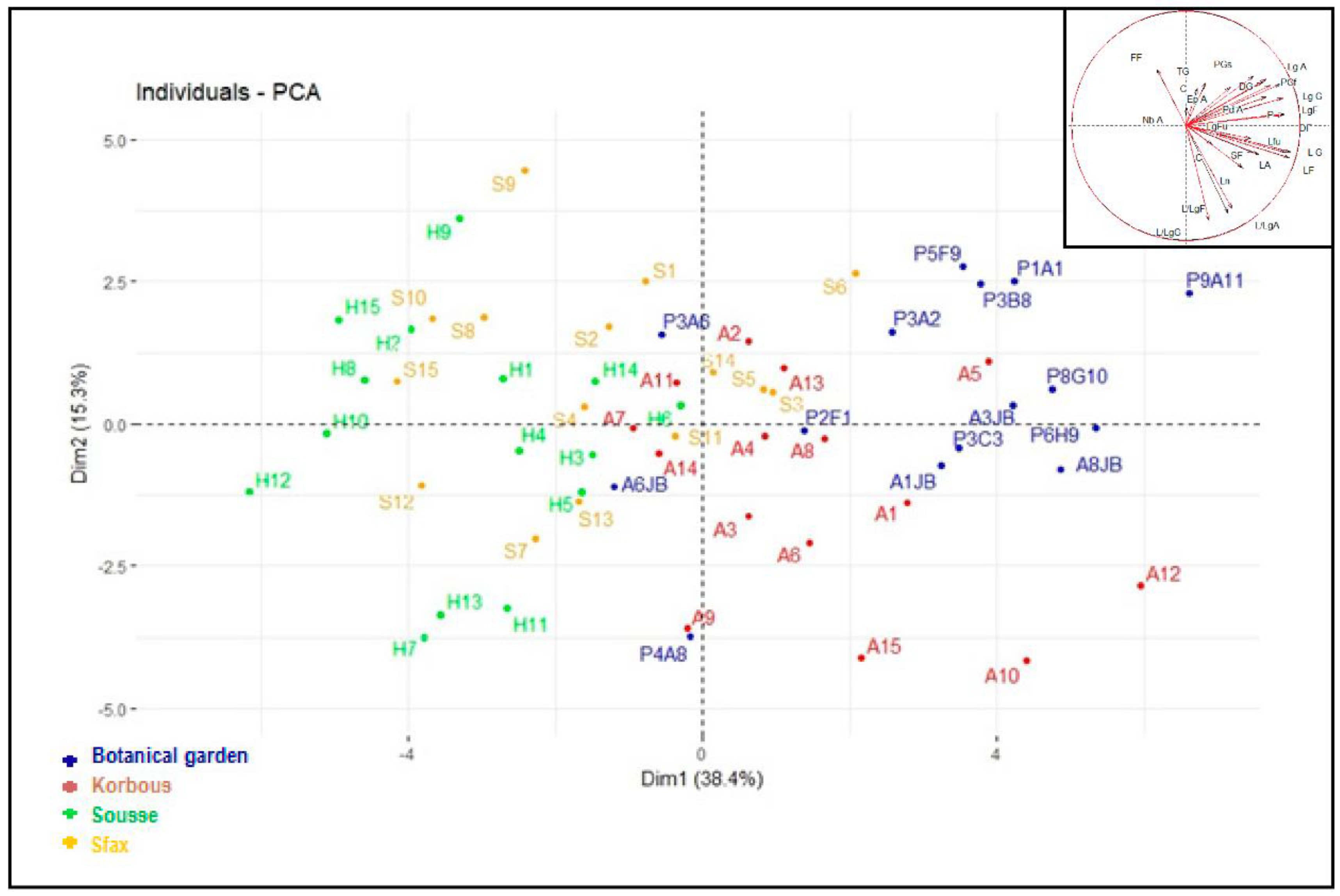
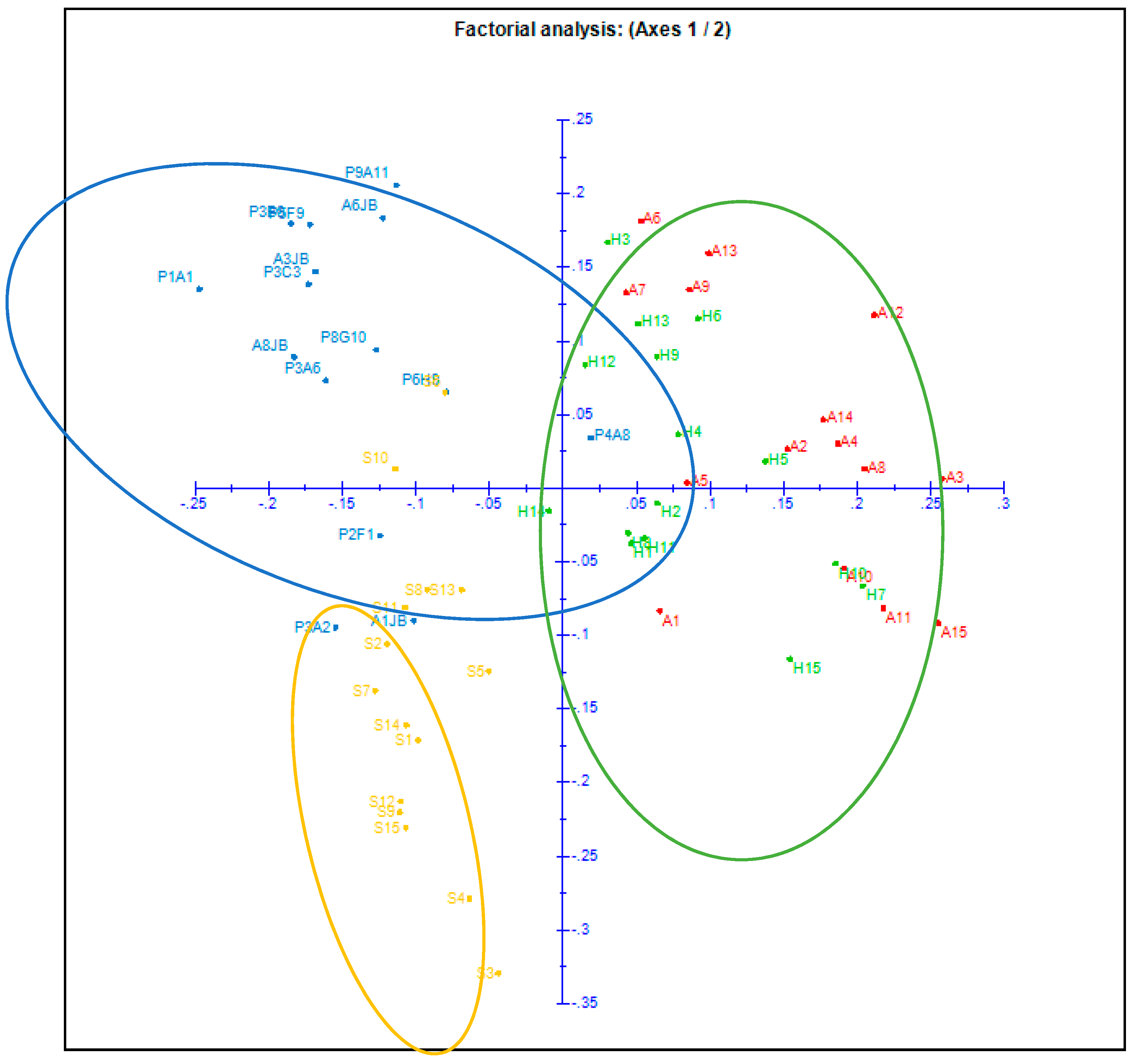
2.2. Principal Component Analysis (PCA)
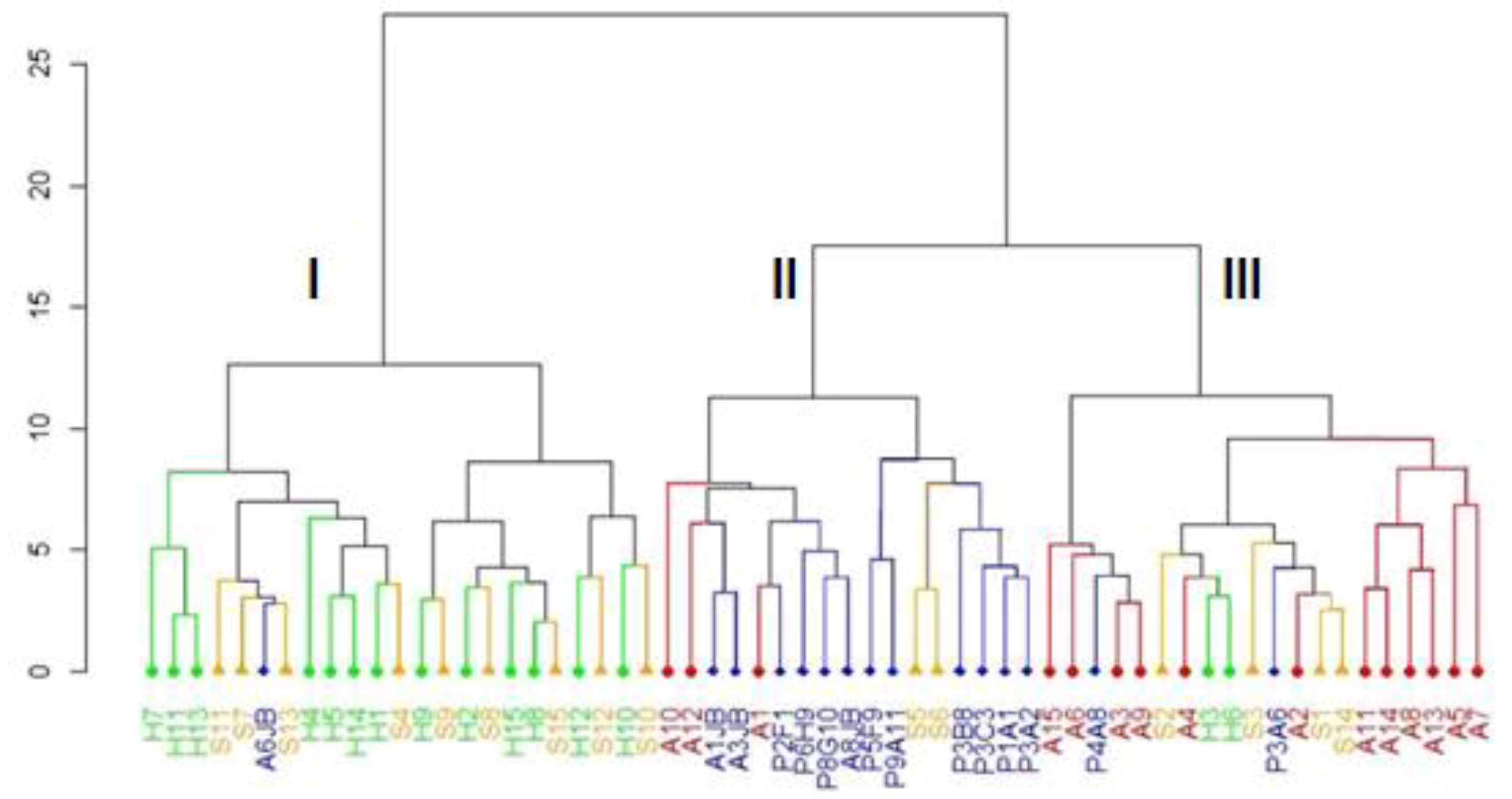
2.3. Cluster Analysis—CA
2.4. Optimization of the ISSR-PCR Reaction System
2.5. Genetic Diversity
2.6. Genetic Differentiation and Gene Flow
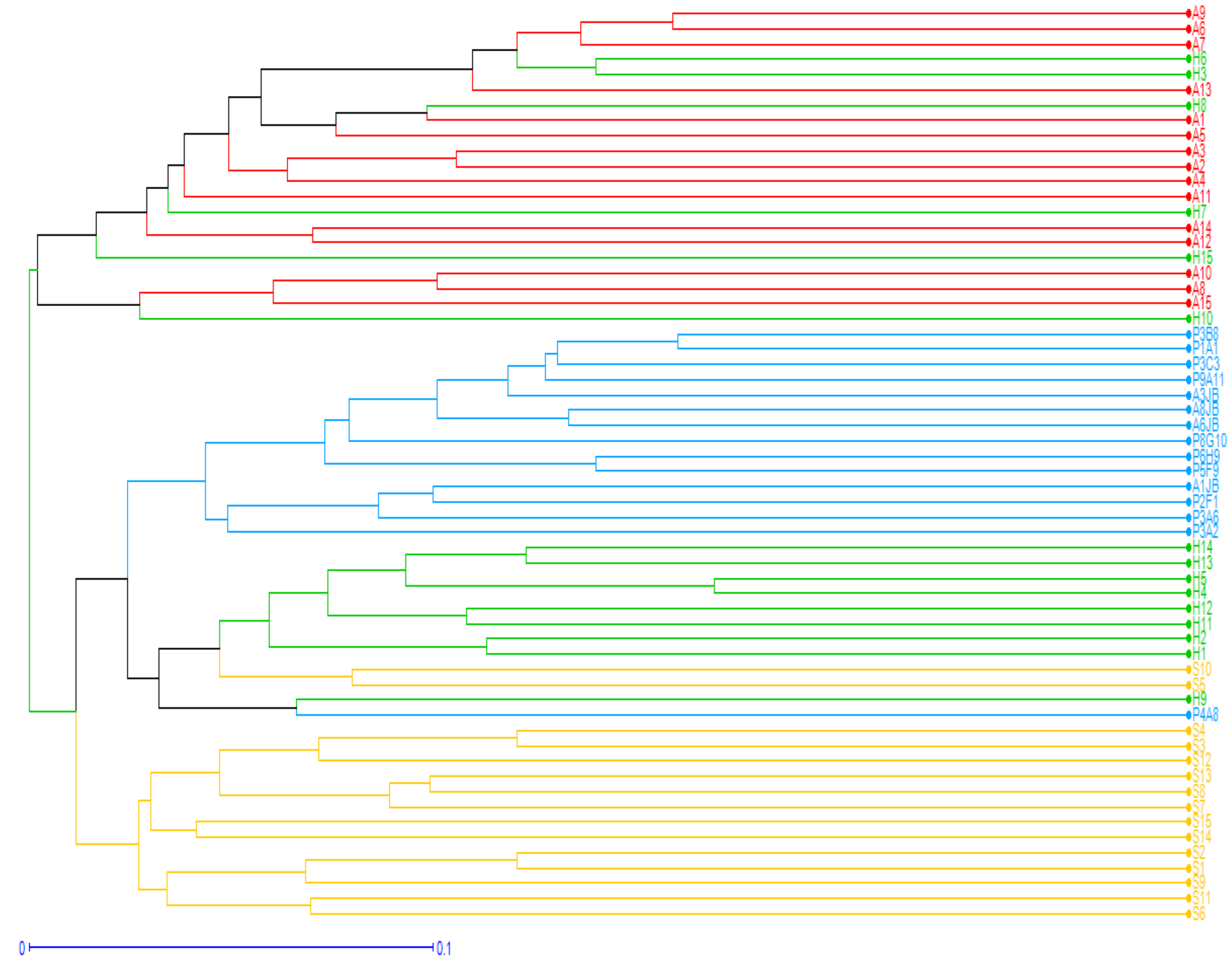
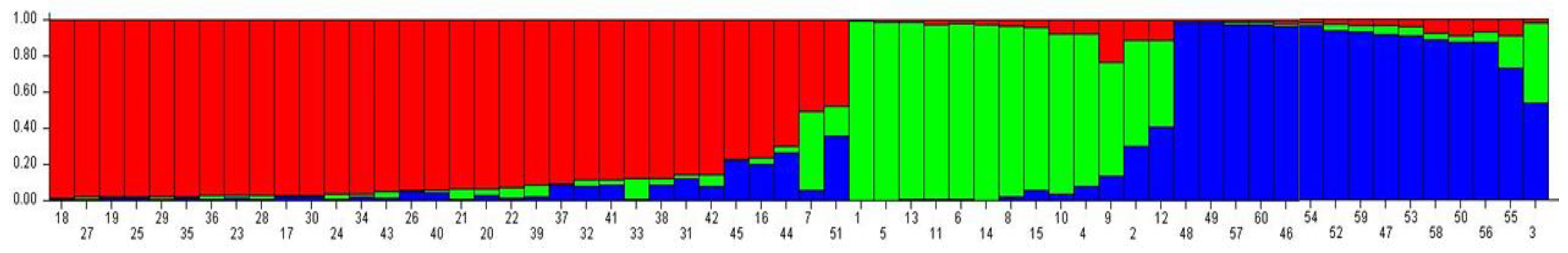
2.7. Clustering and Genetic Relationships among Argan Provenances
2.8. High Resolution Melting (HRM) Analysis
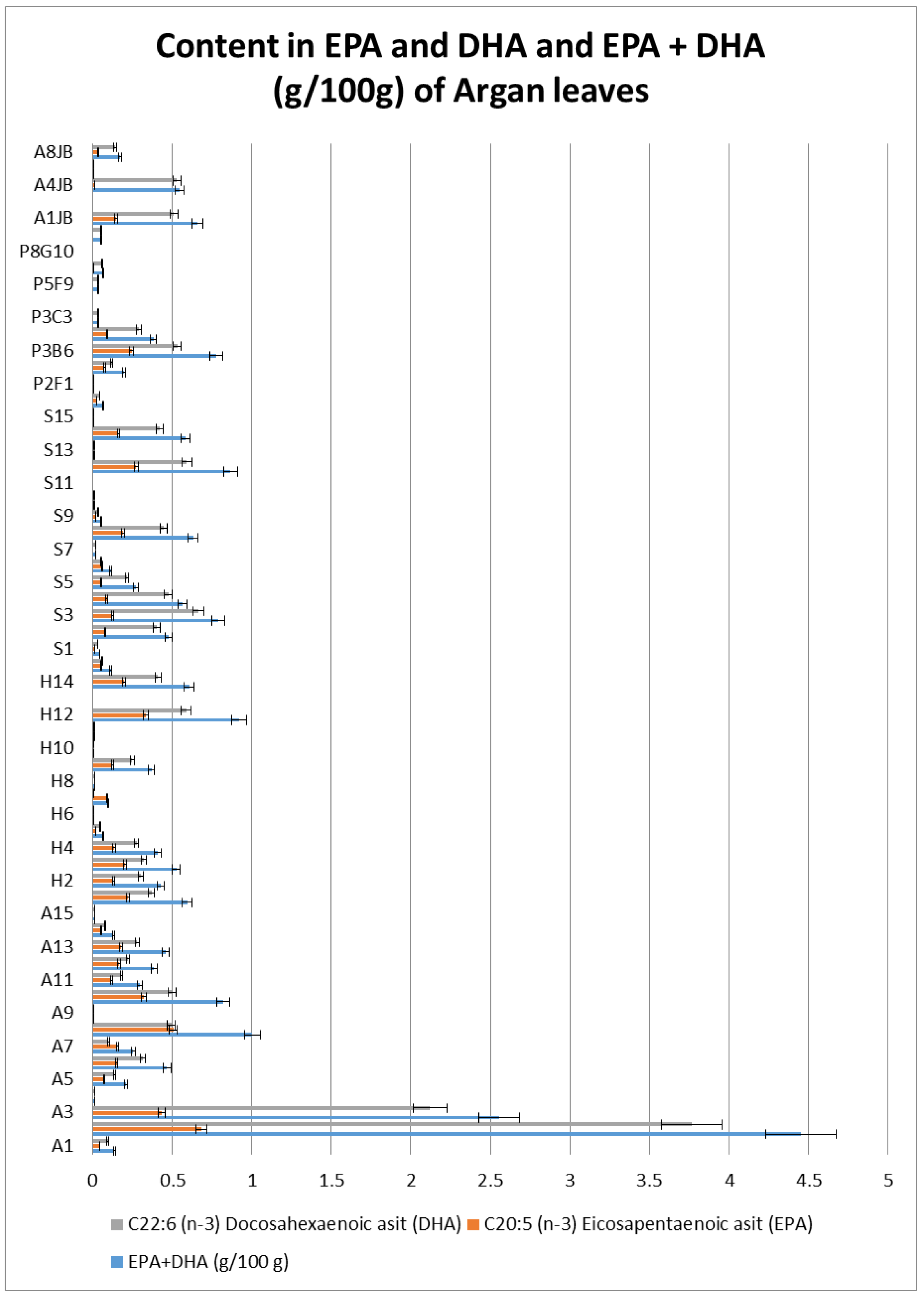
2.9. Fatty Acid Analysis
3. Discussion
3.1. Morphological Diversity
3.2. Genetic Diversity
3.3. Genetic Differentiation and Gene Flow
3.4. Genetic Structure
3.5. HRM
3.6. Fatty Acid Analysis
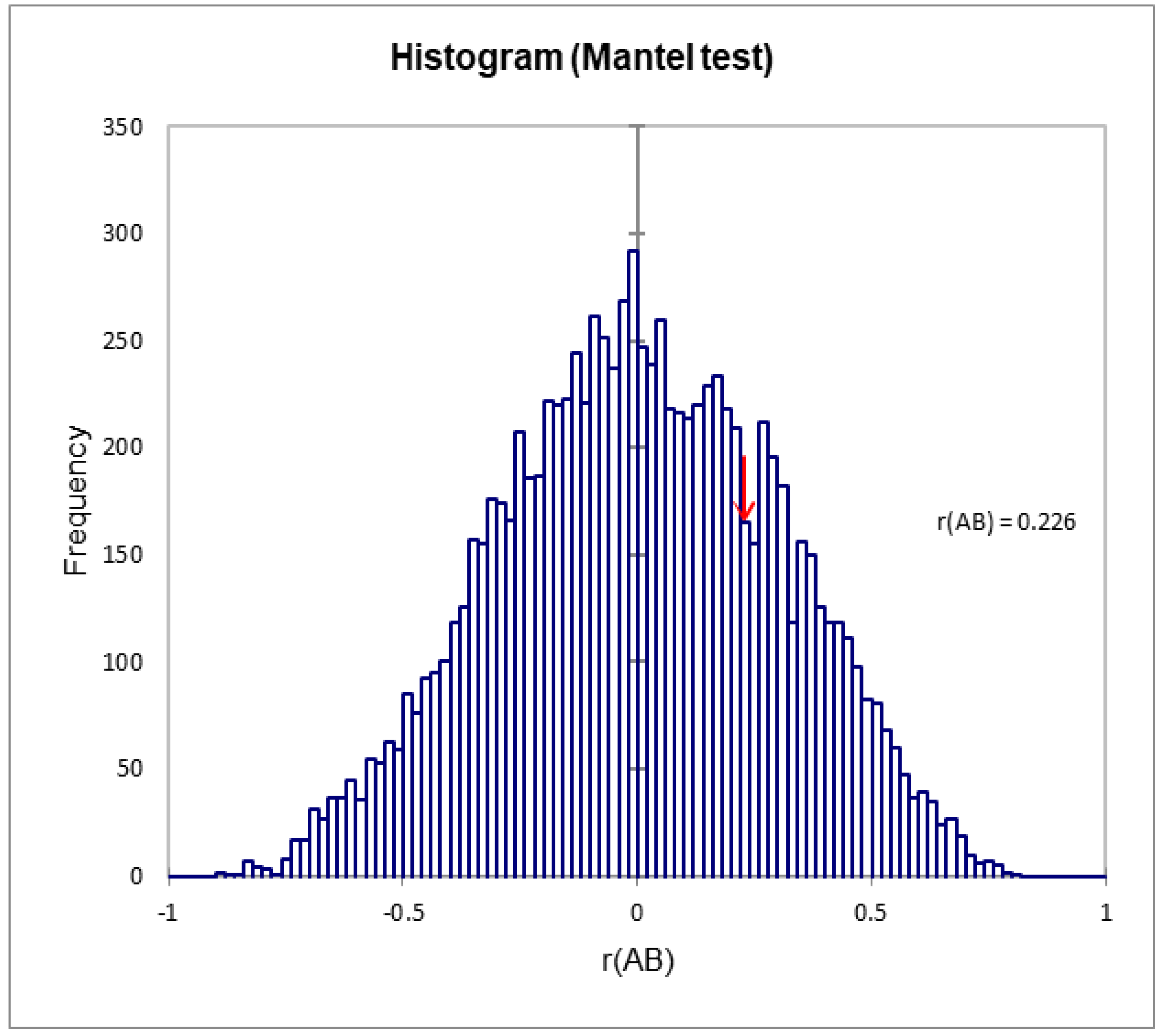
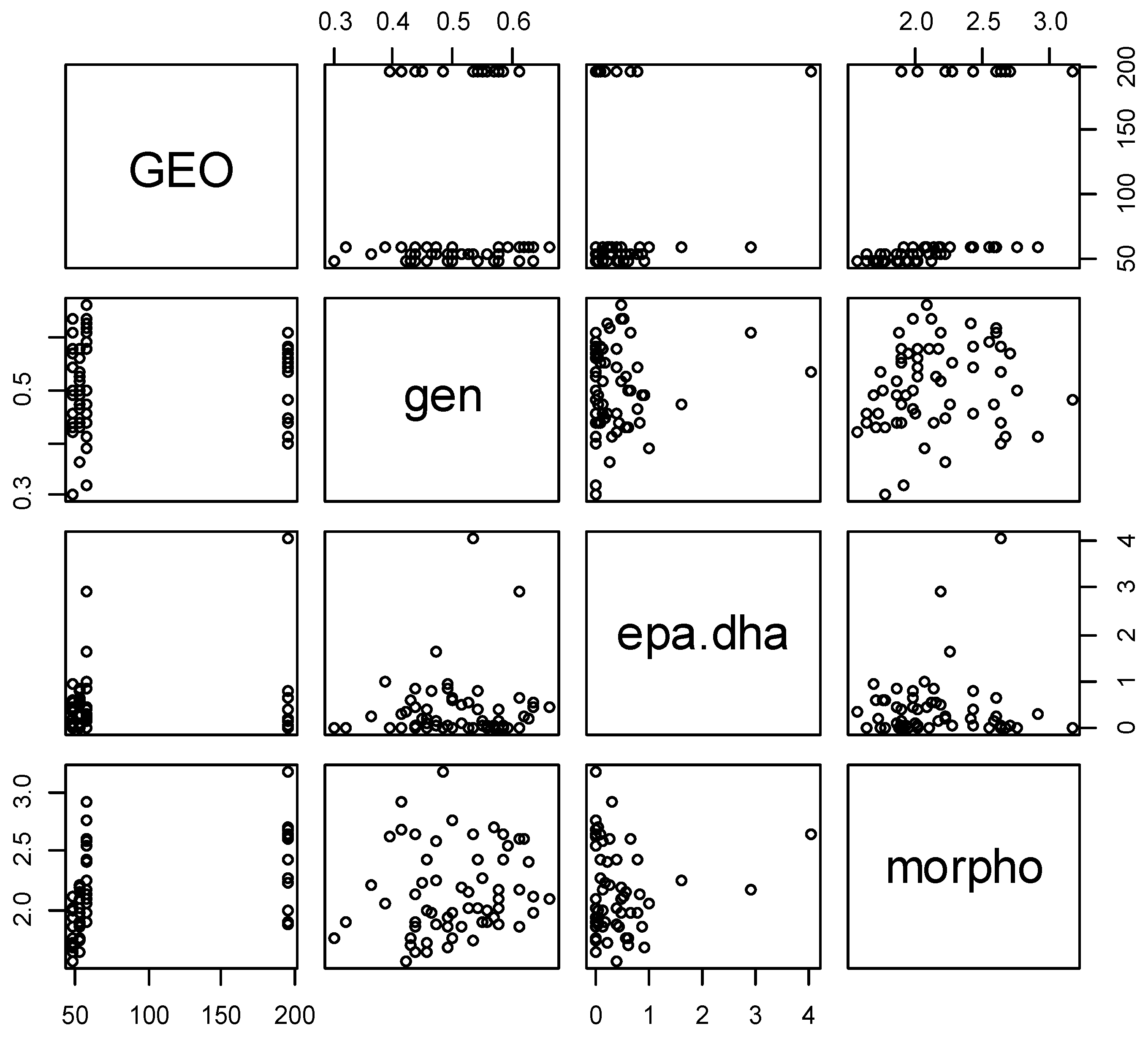
3.7. Correlation between the Different Analyses
4. Materials and Methods
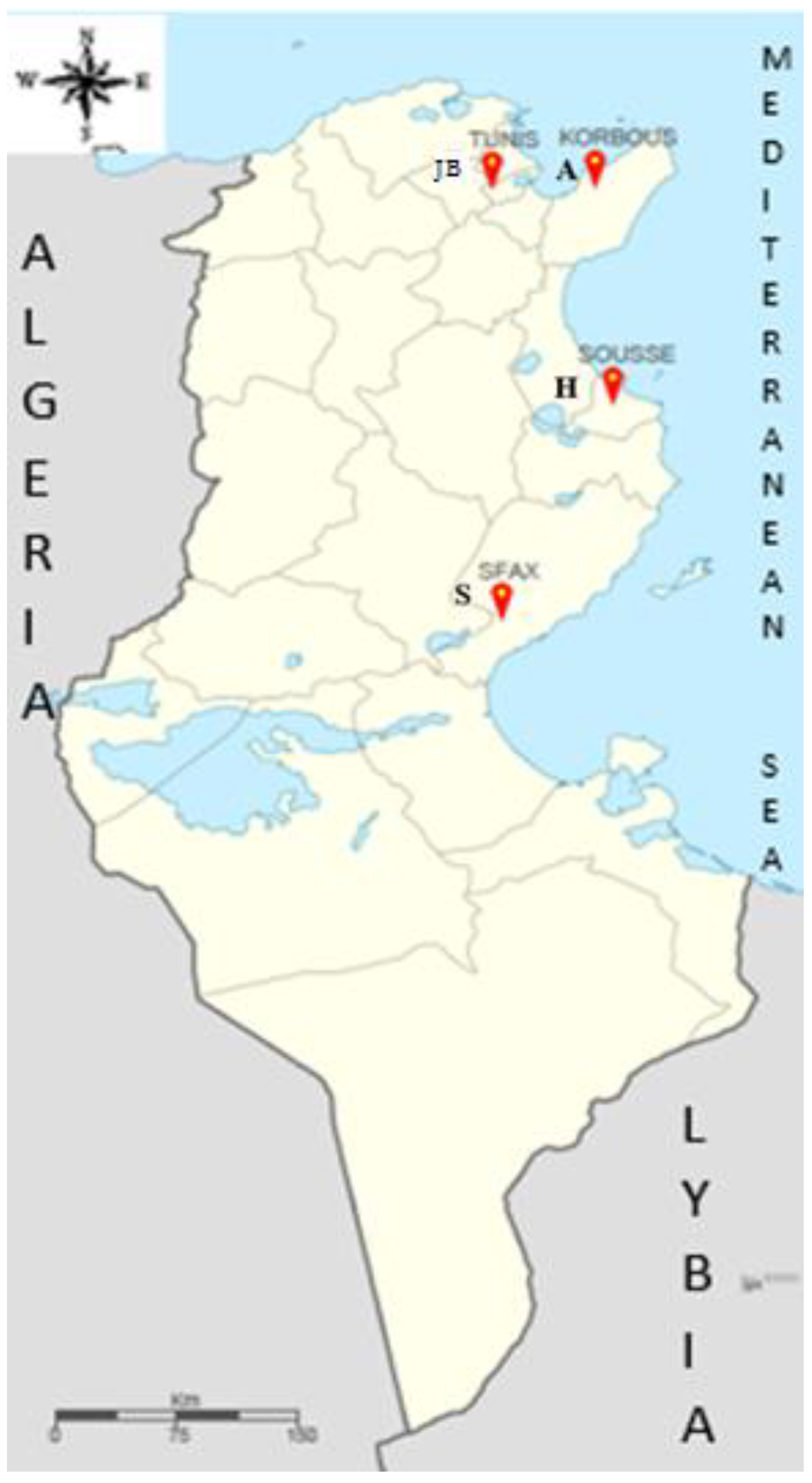
4.1. The Geographical Area of the Study
4.2. Morphological and Pomological Characterization
4.3. Extraction and Purification of DNA
4.4. ISSR Analysis
4.5. PCR and HRM Conditions
4.6. Fatty Acid Analysis
4.7. Statistical Analysis
5. Conclusions and Perspective
- -
- The first most important management practice is to increase awareness about the importance of this species and start exploiting these trees with moderation especially trees with high genetic diversity. At the same time, we should allow populations to increase in size through natural regeneration and also try to grow argan trees in different locations in Tunisia, especially locations under the threat of desertification since it tolerates high temperatures and salinity and fights desertification.
- -
- The second possible strategy is to focus on the establishment of an ex situ conservation program by seed storage from all the four sites to preserve most of the genetic variation.
- -
- The third possible strategy consists of building a gene bank with a large number of plantations in a natural reserve, taking into consideration all the populations and genetic groups so that we can increase the possibility of gene exchange and ensure long-term conservation of this genetic variation. Knowing the economical, ecological, nutritional, and pharmaceutical value of this tree, it is important to increase awareness in Tunisia about the importance of this tree in order to preserve it. In addition, it would be interesting to combine the studies on the distribution of genetic diversity with other information such as oil composition and morphological characters per population.
Author Contributions
Funding
Conflicts of Interest
References
- Berrougui, H.; Cloutier, M.; Isabelle, M.; Khalil, A. Phenolic-extract from argan oil (Argania spinosa L.) inhibits human low-density lipoprotein (LDL) oxidation and enhances cholesterol efflux from human THP-1 macrophages. Atherosclerosis. 2006, 184, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Majourhat, K.; Jabbar, Y.; Araneda, L.; Zeinalabedini, M.; Hafidi, A.; Martínez-Gómez, P. Karyotype characterization of Argania spinosa (L.) Skeel (Sapotaceae). S. Afr. J. Bot. 2007, 73, 661–663. [Google Scholar] [CrossRef]
- Msanda, F.; El Aboudi, A.; Peltier, J.P. Biodiversité et biogéographie de l’arganeraie marocaine. Cah. Agric. 2005, 14, 357–364. [Google Scholar]
- Thompson, J.D. Population differentiation in Mediterranean plants: insights into colonization history and the evolution and conservation of endemic species. Heredity 1999, 82, 229–236. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hamrick, J.L. The distribution of genetic variation within and among natural plant populations, in Genetics and Conservation (eds C. M. Schonewald-Cox, S.M. Chambers, B. MacBryde and W. L. Thomas), Benjamin/Cummings, London, 1983, pp. 335–44. Hamrick, J.L.; Godt, M.W. Effects of life history traits on genetic diversity in plant species. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1996, 351, 1291–1298. [Google Scholar]
- El Mousadik, A.; Petit, R.J. High level of genetic differentiation for allelic richness among populations of the argan tree [Argania spinosa (L.) Skeels] endemic to Morocco. Theor. Appl. Genet. 1996, 92, 832–839. [Google Scholar] [CrossRef]
- El Mousadik, A.; Petit, R.J. Chloroplast DNA phylogeography of the argan tree of Morocco. Mol. Ecol. 1996, 5, 547–555. [Google Scholar] [CrossRef]
- Nouaim, R.; Chaussod, R.; El Aboudi, A.; Schnabel, C.; Peltier, J.P. L’arganier. Essai de synthèse des connaissances sur cet arbre. Groupe d’étude de l’arbre, Editor. Physiologie des Arbres et Arbustes en Zones Arides et Semi Arides 1991, 373–388. (In French) [Google Scholar]
- Qian, W.; Ge, S.; Hong, D.Y. Genetic variation within and among populations of a wild rice Oryza granulata from China detected by RAPD and ISSR markers. Theor. Appl. Genet. 2001, 102, 440–449. [Google Scholar] [CrossRef]
- Shokri, E.; Nematzadeh, G.; Zolala, J.; Nasiri, N.; Kazemi-Tabar, S.K.; HosseinpourAzad, N. Delta-6 desaturase makes a different fatty acid profile of seed oil in Iranian borage (Echium amoenum Fisch. and Mey.). Acta Physiol. Plant. 2012, 34, 2413–2418. [Google Scholar] [CrossRef]
- Adiguzel, A.; Agar, G.; Baris, O.; Gulluce, M.; Sahin, F.; Sengul, M. RAPD and FAME analyses of Astragalus species growing in eastern Anatolia region of Turkey. Biochem. Syst. Ecol. 2006, 34, 424–432. [Google Scholar] [CrossRef]
- Krauss, R.M.; Eckel, R.H.; Howard, B.; Appel, L.J.; Daniels, S.R.; Deckelbaum, R.J.; Erdman, J.W., Jr.; Kris-Etherton, P.; Goldberg, I.J.; Kotchen, T.A.; et al. AHA Dietary Guidelines: Revision 2000: A statement for healthcare professionals from the Nutrition Committee of the American Heart Association. Circulation 2000, 102, 2284–2299. [Google Scholar] [CrossRef] [PubMed]
- Ursin, V.M. Modification of plant lipids for human health: Development of functional land-based omega-3 fatty acids. J. Nutr. 2003, 133, 4271–4274. [Google Scholar] [CrossRef] [PubMed]
- Majourhat, K.; Jabbar, Y.; Hafidi, A.; Martínez-Gómez, P. Molecular characterization and genetic relationships among most common identified morphotypes of critically endangered rare Moroccan species Argania spinosa (Sapotaceae) using RAPD and SSR markers. Ann. For. Sci. 2008, 65, 1. [Google Scholar] [CrossRef]
- Mouhaddab, J.; Msanda, F.; Filali-Maltouf, A.; Belkadi, B.; Ferradouss, A.; El Modafar, C.; El Mousadik, A. Using microsatellite markers to map genetic diversity and population structure of an endangered Moroccan endemic tree (Argania spinosa L. Skeels) and development of a core collection. Plant Gene 2017, 10, 51–59. [Google Scholar] [CrossRef]
- Aabd, N.A.; Msanda, F.; Mousadik, A.E. Genetic diversity of the endangered argan tree (Argania spinosa L.) (sapotaceae) revealed by ISSR analysis. Basic Res. J. Agric. Sci. Rev. 2015, 4, 176–186. [Google Scholar]
- Mouhaddab, J.; Naima, A.I.T.; Achtak, H.; Msanda, F.; Zahidi, A.; Filali-Maltouf, A.; Ferradouss, A.; El Modafar, C.; Nejmeddine, M.; El Mousadik, A. Patterns of genetic diversity and structure at fine scale of an endangered Moroccan endemic tree (Argania spinosa L. Skeels) based on ISSR polymorphism. Not. Bot. Horti Agrobot. Cluj-Napoca 2015, 43, 528–535. [Google Scholar] [CrossRef]
- Yatrib, C.; Belkadi, B.; Pakhrou, O.; Alami, M.; Medraoui, L.; El, A.; Mousadik, A.F.; Msanda, F.; El Modafar, C.; Souda-Kouraichi, S.I.; et al. Assessment of genetic diversity of Argania spinosa L. growing in arid and semi-arid areas of Morocco as revealed by Inter-Simple Sequence Repeats. J. Agric. Sci. Technol. B 2015, 5, 336–346. [Google Scholar]
- Pakhrou, O.; Medraoui, L.; Yatrib, C.; Alami, M.; Souda-kouraichi, S.I.; Ferradous, A.; Msanda, F.; Filali-Maltouf, A.; Belkadi, B. Study of genetic diversity and differentiation of argan tree population (Argania spinosa L.) using AFLP markers. Aust. J. Crop Sci. 2016, 10, 990. [Google Scholar] [CrossRef]
- Pakhrou, O.; Medraoui, L.; Yatrib, C.; Alami, M.; Filali-maltouf, A.; Belkadi, B. Assessment of genetic diversity and population structure of an endemic Moroccan tree (Argania spinosa L.) based in IRAP and ISSR markers and implications for conservation. Physiol. Mol. Boil. Plants 2017, 23, 651–661. [Google Scholar] [CrossRef]
- Zietkiewicz, E.; Rafalski, A.; Labuda, D. Genome fingerprinting by simple sequence repeat (SSR)-anchored polymerase chain reaction amplification. Genomics 1994, 20, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Ganopoulos, I.; Kalivas, A.; Kavroulakis, N.; Xanthopoulou, A.; Mastrogianni, A.; Koubouris, G.; Madesis, P. Genetic diversity of Barbary fig (Opuntia ficus-indica) collection in Greece with ISSR molecular markers. Plant Gene 2015, 2, 29–33. [Google Scholar] [CrossRef]
- Scotti-Saintagne, C.; Mariette, S.; Porth, I.; Goicoechea, P.G.; Barreneche, T.; Bodénes, C.; Burg, K.; Kremer, A. Genome scanning for interspecific differentiation between two closely related oak species [Quercus robur L. and Quercus petraea (Matt.) Liebl.]. Genetics 2004, 168, 1615–1626. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, S.; Das, A.B.; Ghosh, N.; Panda, B.B.; Smith, D.W. Genetic diversity of 28 wild species of fodder legume Cassia using RAPD, ISSR and SSR markers: a novel breeding strategy. J. Biotech Res. 2010, 2, 44. [Google Scholar]
- Prakash, L.; Middha, S.K.; Mohanty, S.K.; Swamy, M.K. Micropropagation and validation of genetic and biochemical fidelity among regenerants of Nothapodytes nimmoniana (Graham) Mabb. employing ISSR markers and HPLC. 3 Biotech 2016, 6, 171. [Google Scholar] [CrossRef]
- Santana, J.G.S.; Nascimento, A.L.S.; Costa, T.S.; de Almeida, T.M.B.; Rabbani, A.R.C.; MUNIZ, A.D.S. Estimation of genetic diversity in a natural population of cambui tree (Myrciaria tenella O. Berg) using ISSR markers. Embrapa Tabuleiros Costeiros-Artigo em Periódico Indexado (ALICE) 2016. [Google Scholar] [CrossRef] [PubMed]
- Ringnér, M. What is principal component analysis? Nat. Biotechnol. 2008, 26, 303. [Google Scholar] [CrossRef]
- Nei, M. Analysis of gene diversity in subdivided populations. Proc. Natl. Acad. Sci. USA 1973, 70, 3321–3323. [Google Scholar] [CrossRef]
- Nei, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 1978, 89, 583–590. [Google Scholar]
- Zahidi, A.; Bani-Aameur, F.; Mousadık, E. Morphological variability of the fruiting branches in Argania spinosa: Effects of seasonal variations, locality and genotype. J. Hortic. For. 2013, 5, 168–182. [Google Scholar]
- Bani-Aameur, F.; Ferradous, A. Fruits and stone variability in three argan (Argania spinosa (L.) Skeels) populations. For. Genet. 2001, 8, 39–43. [Google Scholar]
- Hartl, D.L.; Clark, A.G. Principles of population genetics. Sunderland, MA. Sinauer Assoc. 1997, 116, 95–109. [Google Scholar]
- Aabd, N.A.; El-Ayadi, F.; Msanda, F.; El-Mousadik, A. Evaluation of agromorphological variability of argan tree under different environmental conditions in Morocco: Implication for selection. Int. J. Biodivers. Conserv. 2011, 3, 73–82. [Google Scholar]
- Huang, Y.; Ji, K.; Jiang, Z.; Tang, G. Genetic structure of Buxus sinica var. parvifolia, a rare and endangered plant. Sci. Hortic. 2008, 116, 324–329. [Google Scholar] [CrossRef]
- Verma, K.; Kumar, A.G. Studies of Genetic Diversity in the Natural Population of Cassia Fistula L. Using RAPD and ISSR Markers. Ph.D. Thesis, Thapar Institute of Engineering and Technology, Patiala, India, 2017. [Google Scholar]
- Nimbalkar, S.D.; Jade, S.S.; Kauthale, V.K.; Agale, S.; Bahulikar, R.A. Genetic diversity in the candidate trees of Madhuca indica JF Gmel. (Mahua) revealed by inter-simple sequence repeats (ISSRs). 3 Biotech 2018, 8, 143. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Al-Qurainy, F.; Nadeem, M. Biotechnological approaches for conservation and improvement of rare and endangered plants of Saudi Arabia. Saudi J. Biol. Sci. 2012, 19, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Maquia, I.; Ribeiro, N.S.; Silva, V.; Bessa, F.; Goulao, L.F.; Ribeiro, A.I. Genetic diversity of Brachystegia boehmii Taub. and Burkea africana Hook. f. across a fire gradient in Niassa National Reserve, Northern Mozambique. Biochem. Syst. Ecol. 2013, 48, 238–247. [Google Scholar] [CrossRef]
- Liu, J.; Sun, H.G.; Jiang, J.M.; Shao, W.H.; Luan, Q. Genetic diversity of natural populations of Machilus thunbergii, an endangered tree species in eastern China, determined with ISSR analysis. Genet. Mol. Res. 2013, 12, 3689–3697. [Google Scholar] [CrossRef]
- Hedrick, P.W. Book reviews: Genetics and conservation. Evolution 1985, 39, 1180–1181. [Google Scholar] [CrossRef][Green Version]
- Ellstrand, N.C.; Elam, D.R. Population genetic consequences of small population size: Implications for plant conservation. Annu. Rev. Ecol. Syst. 1993, 24, 217–242. [Google Scholar] [CrossRef]
- Kremer, A.; Kleinschmit, J.; Cottrell, J.; Cundall, E.P.; Deans, J.D.; Ducousso, A.; König, A.O.; Lowe, A.J.; Munro, R.C.; Petit, R.J.; et al. Is there a correlation between chloroplastic and nuclear divergence, or what are the roles of history and selection on genetic diversity in European oaks? For. Ecol. Manag. 2002, 156, 75–87. [Google Scholar] [CrossRef]
- Hui-yu, L.; Jing, J.; Gui-feng, L.; Xu-jun, M.; Jing-xiang, D.; Shi-jie, L. Genetic variation and division of Pinus sylvestris provenances by ISSR markers. J. For. Res. 2005, 16, 216–218. [Google Scholar] [CrossRef]
- Xia, T.; Meng, L.; Mao, K.; Tian, B.; Miehe, G.; Liu, J. Genetic variation in the Qinghai-Tibetan plateau endemic and endangered conifer Cupressus gigantea, detected using RAPD and ISSR markers. Silvae Genet. 2008, 57, 85–92. [Google Scholar] [CrossRef]
- Wang, X.; Yang, R.; Feng, S.; Hou, X.; Zhang, Y.; Li, Y.; Ren, Y. Genetic variation in Rheum palmatum and Rheum tanguticum (Polygonaceae), two medicinally and endemic species in China using ISSR markers. PLoS ONE 2012, 7, 51667. [Google Scholar] [CrossRef] [PubMed]
- Hogbin, P.M.; Peakall, R. Evaluation of the contribution of genetic research to the management of the endangered plant Zieria prostrata. Conserv. Biol. 1999, 13, 514–522. [Google Scholar] [CrossRef]
- Slatkin, M. Gene flow and the geographic structure of natural populations. Science 1987, 236, 787–792. [Google Scholar] [CrossRef] [PubMed]
- Gabrielsen, T.M.; Bachmann, K.; Jakobsen, K.S.; Brochmann, C. Glacial survival does not matter: RAPD phylogeography of Nordic Saxifraga oppositifolia. Mol. Ecol. 1997, 6, 831–842. [Google Scholar] [CrossRef]
- Schaal, B.A.; Hayworth, D.A.; Olsen, K.M.; Rauscher, J.T.; Smith, W.A. Phylogeographic studies in plants: Problems and prospects. Mol. Ecol. 1998, 7, 465–474. [Google Scholar] [CrossRef]
- Wright, S. The genetical structure of populations. Ann. Eugen. 1949, 15, 323–354. [Google Scholar] [CrossRef]
- Rodrigues, L.; van den Berg, C.; Póvoa, O.; Monteiro, A. Low genetic diversity and significant structuring in the endangered Mentha cervina populations and its implications for conservation. Biochem. Syst. Ecol. 2013, 50, 51–61. [Google Scholar] [CrossRef]
- Nettleton, J.A. Omega-3 fatty acids: Comparison of plant and seafood sources in human nutrition. J. Am. Diet. Assoc. 1991, 91, 331–337. [Google Scholar] [PubMed]
- Simopoulos, A.P. Omega-3 fatty acids in health and disease and in growth and development. Am. J. Clin. Nutr. 1991, 54, 438–463. [Google Scholar] [CrossRef] [PubMed]
- Qi, B.; Fraser, T.; Mugford, S.; Dobson, G.; Sayanova, O.; Butler, J.; Napier, J.A.; Stobart, A.K.; Lazarus, C.M. Production of very long chain polyunsaturated omega-3 and omega-6 fatty acids in plants. Nat. Biotechnol. 2004, 22, 739. [Google Scholar] [CrossRef] [PubMed]
- Omara-Alwala, T.R.; Mebrahtu, T.; Prior, D.E.; Ezekwe, M.O. Omega-three fatty acids in purslane (Portulaca oleracea) Tissues. J. Am. Oil Chem. Soc. 1991, 68, 198–199. [Google Scholar] [CrossRef]
- Guil, J.L.; Torija, M.E.; Gimenez, J.J.; Rodriguez, I. Identification of fatty acids in edible wild plants by gas chromatography. J. Chromatogr. A 1996, 719, 229–235. [Google Scholar] [CrossRef]
- Kris-Etherton, P.M.; Taylor, D.S.; Yu-Poth, S.; Huth, P.; Moriarty, K.; Fishell, V.; Hargrove, R.L.; Zhao, G.; Etherton, T.D. Polyunsaturated fatty acids in the food chain in the United States. Am. J. Clin. Nutr. 2000, 71, 179S–188S. [Google Scholar] [CrossRef] [PubMed]
- Krzynowek, J.; Panunzio, L.J. Cholesterol and fatty acids in several species of shrimp. J. Food Sci. 1989, 54, 237–239. [Google Scholar] [CrossRef]
- RStudio. R. RStudio: Integrated Development Environment for R (version 0.95.262)[computer software] Boston, MA, USA. 2012. Available online: http://www.rstudio.org/ (accessed on 19 August 2013).
- Reddy, M.P.; Sarla, N.; Siddiq, E.A. Inter simple sequence repeats (ISSR) polymorphism and its application in plant breeding. Euphytica 2002, 120, 9–16. [Google Scholar] [CrossRef]
- Louati, M.; Khouja, A.; Ben Abdelkrim, A.; Hannachi, A.S.; Baraket, G. Adaptation of Argania spinosa L. in Northern Tunisia: Soil analysis and morphological traits variability. Sci. Hortic. 2019, 255, 220–230. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Ray, B.; Loutelier-Bourhis, C.; Lange, C.; Condamine, E.; Driouich, A.; Lerouge, P. Structural investigation of hemicellulosic polysaccharides from Argania spinosa: Characterisation of a novel xyloglucan motif. Carbohydr. Res. 2004, 339, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Souto Alves, T.; Vanusa da Silva, M.; Alves de Almeida, C.M.; Oliveira Jordão do Amaral, D.; Cordeiro dos Santos, D.; Farias, I.; Tenório Sabino Donato, V.M.; Da Costa, A.F. Genetic diversity in cactus clones using ISSR markers. Acta Hortic. 2009, 811, 55–58. [Google Scholar] [CrossRef]
- Verma, S.; Rana, T.S. Genetic diversity within and among the wild populations of Murraya koenigii (L.) Spreng., as revealed by ISSR analysis. Biochem. Syst. Ecol. 2011, 39, 139–144. [Google Scholar] [CrossRef]
- Su, Y.J.; Zan, Q.J.; Wang, T.; Ying, Z.M.; Ye, H.G. High ISSR variation in 24 surviving individuals of Apterosperma oblata (Theaceae) endemic to China. Biochem. Syst. Ecol. 2008, 36, 619–625. [Google Scholar] [CrossRef]
- Ucarli, C.; Poyraz, N.; Gürel, A.F. Discovery of Dhn3 variants in wild barley (Hordeum spontaneum) byhigh-resolution melting (HRM) technology. Turk. J. Biol. 2015, 39, 758–764. [Google Scholar] [CrossRef]
- Ichihara, K.I.; Waku, K.; Yamaguchi, C.; Saito, K.; Shibahara, A.; Miyatani, S.; Yamamoto, K. A convenient method for determination of the C20–22 PUFA composition of glycerolipids in blood and breast milk. Lipids 2002, 37, 523–526. [Google Scholar] [CrossRef] [PubMed]
- Dagnelie, P. Analyse Statistique à Plusieurs Variables; Presses agronomiques de Gembloux: Gembloux, Belgium, 1999. [Google Scholar]
- Abdelkefi, A.; Boussaid, M.; Biborchi, A.; Haddioui, A.; Salhi-Hanachi, A.; Marrakchi, M. Genetic diversity inventory and valuation of spontaneous species belonging to Medicago L. genus in Tunisia. AGRIS 1996, 18, 185. [Google Scholar]
- Yeh, F.C.; Yang, R.; Boyle, T.J.; Ye, Z.; Xiyan, J.M. PopGene32, Microsoft Windows-Based Freeware for Population Genetic Analysis, Version 1.32; Molecular Biology and Biotechnology Centre, University of Alberta: Edmonton, AB, Canada, 2000. [Google Scholar]
- McDermott, J.M.; McDonald, B.A. Gene flow in plant pathosystems. Annu. Rev. Phytopathol. 1993, 31, 353–373. [Google Scholar] [CrossRef]
- Yeh, F.C.; Yang, R.C.; Boyle, T.J.B. POPGENE VERSION 1.31 Microsoft window-based freeware for population genetic analysis. Available online: http://www.ualberta.ca/~fyeh/popgene.html (accessed on 12 August 2012).
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Boil. Evolut. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Perrier, X.; Jacquemoud-Collet, J.P. DARwin Software. 2006. Available online: http://darwin.cirad.fr/darwin (accessed on 30 January 2019).
- Peakall, R.O.D.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Matthew, S.; Peter, D. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Earl, D.A. Structure Harvester: A website and program for visualizing Structure output and implementing the Evanno method. Conserv. Genet. Res. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Mantel, N. The detection of disease clustering and a generalized regression approach. Cancer Res. 1967, 27, 209–220. [Google Scholar] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Codes | DF | Sum of sq | Average of sq | F | P > F |
|---|---|---|---|---|---|
| LF | 11 | 17.398 | 1.582 | 6.297 | <0.0001 |
| LgF | 11 | 2.305 | 0.210 | 1.803 | 0.080 |
| L/LgF | 11 | 2.168 | 0.197 | 7.453 | <0.0001 |
| P | 11 | 184.492 | 16.772 | 3.955 | <0.0001 |
| Ln | 11 | 2.590 | 0.235 | 9.305 | <0.0001 |
| DF | 11 | 30.207 | 2.746 | 2.817 | 0.006 |
| L G | 11 | 8.328 | 0.757 | 4.331 | <0.0001 |
| LgG | 11 | 1.236 | 0.112 | 1.616 | 0.124 |
| L/LgG | 11 | 2.043 | 0.186 | 2.608 | 0.011 |
| PGf | 11 | 11.180 | 1.016 | 1.919 | 0.060 |
| PGs | 11 | 5.750 | 0.523 | 1.433 | 0.189 |
| DG | 11 | 5.937 | 0.540 | 1.387 | 0.210 |
| TG | 11 | 1.017 | 0.092 | 2.611 | 0.011 |
| Nb A | 11 | 1.140 | 0.104 | 1.356 | 0.224 |
| Pd A | 11 | 0.048 | 0.004 | 1.205 | 0.309 |
| LA | 11 | 5.211 | 0.474 | 3.717 | 0.001 |
| Lg A | 11 | 0.287 | 0.026 | 0.711 | 0.722 |
| L/Lg A | 11 | 3.445 | 0.313 | 2.725 | 0.008 |
| Ep A | 11 | 0.037 | 0.003 | 0.944 | 0.508 |
| Lfu | 11 | 20.742 | 1.886 | 1.928 | 0.059 |
| LgFu | 11 | 0.473 | 0.043 | 1.604 | 0.128 |
| SF | 11 | 9.696 | 0.881 | 1.721 | 0.097 |
| Primers | Motif | Ta (°C) | MgCl2 (mM) | Size (bp) | TNB | NPB | PPB% | PIC | No. of Bands in Each Population | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| JB | A | H | S | |||||||||
| ISSR 1/8 | (AG)8C | 50 °C | 2 | 200–1000 | 10 | 8 | 80% | 54% | 10/10 | 10/10 | 10/10 | 10/10 |
| ISSR 3/8 | (GA)8YG | 50.3 °C | 1.5 | 200–900 | 8 | 6 | 75% | 47.6% | 8/8 | 8/8 | 8/8 | 8/8 |
| ISSR 4/8 | (AC)8YG | 49 °C | 2 | 300–1500 | 14 | 14 | 100% | 66.4% | 14/14 | 14/14 | 14/14 | 14/14 |
| ISSR 6/8 | (AC)8YT | 52 °C | 1.5 | 300–2500 | 22 | 21 | 95% | 78.1% | 20/22 | 19/22 | 22/22 | 21/22 |
| ISSR 7/8 | (AGC)6 | 60 °C | 2 | 200–1000 | 6 | 5 | 83% | 69.5% | 6/6 | 6/6 | 5/6 | 6/6 |
| ISSR 8/8 | (CTC)6 | 57 °C | 2 | 400–2000 | 11 | 10 | 90% | 69.9% | 10/11 | 11/11 | 10/11 | 10/11 |
| ISSR 807 | (GA)8C | 48.5 °C | 1.7 | 300–1000 | 8 | 6 | 75% | 54.6% | 8/8 | 8/8 | 8/8 | 8/8 |
| ISSR 808 | (CT)8A | 50.8 °C | 2 | 400–2000 | 11 | 10 | 90% | 77.1% | 11/11 | 11/11 | 10/11 | 10/11 |
| ISSR 857 | (AC)8G | 53 °C | 2 | 500–2000 | 13 | 12 | 92% | 67.7% | 13/13 | 13/13 | 12/13 | 13/13 |
| FLA 10 | (GA)8CT | 50.3 °C | 1.8 | 300–2000 | 13 | 12 | 92% | 70.4% | 13/13 | 12/13 | 11/13 | 12/13 |
| Mean± Standard Deviation | Polymorphic Loci | ||||||
|---|---|---|---|---|---|---|---|
| Provenance | Size | na | ne | H | I | NPL | PPL% |
| JB | 15 | 1.81 ± 0.39 | 1.44 ± 0.33 | 0.26 ± 0.18 | 0.396 ± 0.25 | 94 | 81.03% |
| A | 15 | 1.80 ± 0.40 | 1.46 ± 0.35 | 0.27 ± 0.18 | 0.41 ± 0.25 | 93 | 80.17% |
| H | 15 | 1.77 ± 0.41 | 1.41 ± 0.35 | 0.25 ± 0.18 | 0.38 ± 0.25 | 90 | 77.59% |
| S | 15 | 1.8 ± 0.39 | 1.43 ± 0.35 | 0.26 ± 0.17 | 0.40 ± 0.24 | 94 | 81.03% |
| Total | 60 | 1.90 ± 0.31 | 1.53 ± 0.32 | 0.32 ± 0.16 | 0.47 ± 0.21 | 104 | 89.66% |
| JB | A | H | S | |
|---|---|---|---|---|
| 0.000 | JB | |||
| 0.272 | 0.000 | A | ||
| 0.247 | 0.115 | 0.000 | H | |
| 0.234 | 0.237 | 0.186 | 0.000 | S |
| Source of Variation | df | SS | MS | VC | % of Variation | p-Value |
|---|---|---|---|---|---|---|
| Among Pops | 3 | 233.983 | 77.994 | 3.967 | 18% | >0.001 |
| Within Pops | 56 | 1035.200 | 18.486 | 18.486 | 82% | >0.001 |
| Total | 59 | 1269.183 | 22.453 | 100% |
| Group Number | Individual | Group Number | Individual |
|---|---|---|---|
| 1 | A1, A2, A5, A6, A7, A10, A11, A12, H2, H7, H9, S1, S3, S9, S11, S12, S14, P3A2 | 6 | H6, H8, S2, S4 |
| 2 | A9, A13 | 7 | A14, H13, H14, H15 |
| 3 | A15, H10, S7, S8, S10, S13, S15, P1A1, P3A6, P5F9, P6H9 | 8 | H12, S6 |
| 4 | A3, A4, H4, H5, P2F1, P3B8, P3C3, P4A8, P8G10, P9A11, A1JB, A3JB, A6JB | 9 | H3, H1 |
| 5 | S5, A8JB | 10 | H11 |
| Codes | Name of Sites (Cities) | Individuals | Soil Texture | pH of Soil | Climate |
|---|---|---|---|---|---|
| JB | Botanical garden (Tunis) | P1A, P2F1, P3A2, P3A6, P3B8, P3C3, P4A8, P5F9, P6H9, P8G10, P9A11, A1JB, A3JB, A6JB, A8JB | Silty | 8.3 | Semi-arid |
| A | Korbous (Nabeul) | A1, A2, A3, A4, A5, A6, A7, A8, A9, A10, A11, A12, A13, A14, A15 | Sand | 6.2 | Semi-arid |
| S | Sfax (Sfax) | S1, S2, S3, S4, S5, S6, S7, S8, S9, S10, S11, S12, S13, S14, S15 | Silty sand | 6 | Arid |
| H | Souss (Souss) | H1, H2, H3, H4, H5, H6, H7, H8, H9, H10, H11, H12, H13, H14, H15 | Clay loam | 8.4 | Semi-Arid |
| Code | Morphological Character | |
|---|---|---|
| Fruit | ||
| 1 | P F F | Fruit weight |
| 2 | L F | Fruit length |
| 3 | lg F | Width of the fruit |
| 4 | LF/lgF | Ratio length width of the fruit |
| 5 | D F | Diameter of the fruit |
| 6 | F | Fruit shape |
| 7 | C F | Fruit color |
| 8 | L N | Nape length |
| 9 | PG | Weight of the seed |
| 10 | PG S | Weight of the seed after drying in the oven |
| 11 | LG | Length of the seed |
| 12 | Lg | Width of the seed |
| 13 | LG/lg G | Ratio length width of the seed |
| 14 | DG | Diameter of the seed |
| 15 | TG | Number of sprouting lines |
| 16 | CL | Color of the seed |
| 17 | Nbr A | Number of kernels per seed |
| 18 | PA | Weight of kernel |
| 19 | LA | Length of the kernel |
| 20 | Lg A | Width of the kernel |
| 21 | LA/LgA | Ratio length width of the kernel |
| 22 | Ep A | Thickness of the kernel |
| Tree | ||
| 23 | Fr Fleurs | Frequency of the flowers |
| 24 | Fr BF | Frequency of flower buds |
| 25 | Fr Fr | Frequency of fruits |
| 26 | Frm de l’arbre | Shape of the tree |
| 27 | L de l’arbre | Length of the tree |
| 28 | D trc | Trunk diameter |
| 29 | Fr ramific | Frequency of ramifications |
| 30 | Fr ep | Frequency of spines |
| Leaves | ||
| 31 | Lf | Length of the leaf |
| 32 | Lf | Width of the sheet |
| 33 | Sf | Leaf area |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Louati, M.; Ucarli, C.; Arikan, B.; Ghada, B.; Salhi Hannachi, A.; Turgut-Kara, N. Genetic, Morphological, and Biochemical Diversity of Argan Tree (Argania spinosa L.) (Sapotaceae) in Tunisia. Plants 2019, 8, 319. https://doi.org/10.3390/plants8090319
Louati M, Ucarli C, Arikan B, Ghada B, Salhi Hannachi A, Turgut-Kara N. Genetic, Morphological, and Biochemical Diversity of Argan Tree (Argania spinosa L.) (Sapotaceae) in Tunisia. Plants. 2019; 8(9):319. https://doi.org/10.3390/plants8090319
Chicago/Turabian StyleLouati, Marwa, Cuneyt Ucarli, Burcu Arikan, Baraket Ghada, Amel Salhi Hannachi, and Neslihan Turgut-Kara. 2019. "Genetic, Morphological, and Biochemical Diversity of Argan Tree (Argania spinosa L.) (Sapotaceae) in Tunisia" Plants 8, no. 9: 319. https://doi.org/10.3390/plants8090319
APA StyleLouati, M., Ucarli, C., Arikan, B., Ghada, B., Salhi Hannachi, A., & Turgut-Kara, N. (2019). Genetic, Morphological, and Biochemical Diversity of Argan Tree (Argania spinosa L.) (Sapotaceae) in Tunisia. Plants, 8(9), 319. https://doi.org/10.3390/plants8090319