Effects of Temperature on Seed Germination of Plantago lanceolata and Management in Carya illinoinensis Production



Abstract
:1. Introduction
2. Results
2.1. Seed Morphology
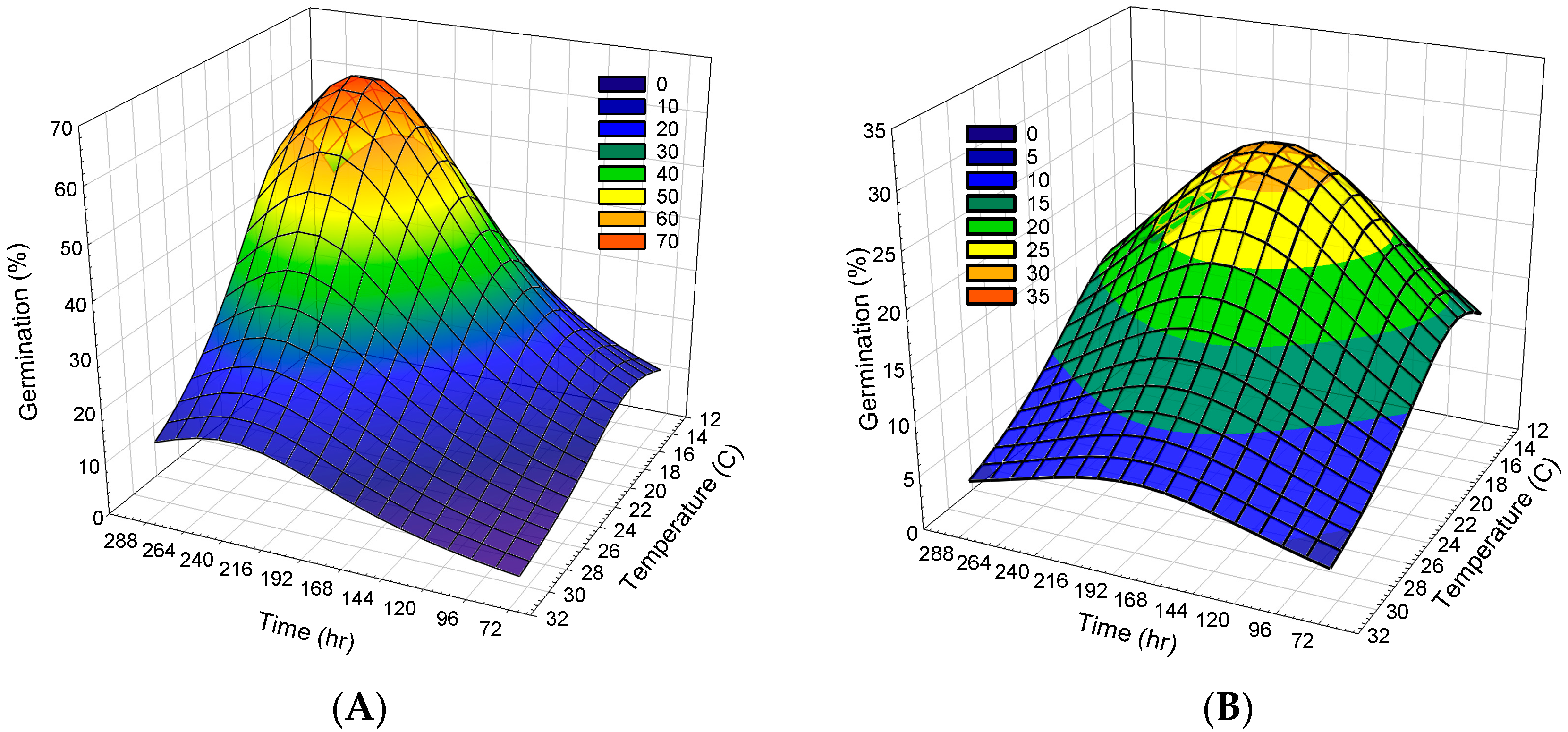
2.2. Germination Evaluation
2.3. Greenhouse and Field Experiments
3. Discussion
3.1. Seed Morphology and Germination
3.2. Herbicide Response
3.3. Conclusions
4. Materials and Methods
4.1. Seed Collection
4.2. Temperature and Germination Evaluation
4.3. Greenhouse Experiments
4.4. Field Experiments
4.5. Statistical Analysis
4.5.1. Germination Evaluation
4.5.2. Cumulative and Maximum P. lanceolata Seed Germination
4.5.3. Greenhouse and Field Experiment
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Grey, T.L.; Turpin, R.S., III; Wells, L.; Webster, T.M. A Survey of weeds and herbicides in Georgia pecan. Weed Technol. 2014, 28, 552–559. [Google Scholar] [CrossRef]
- Bryson, C.T.; DeFelice, M.S. Weeds of the South; University of Georgia Press: Athens, GA, USA, 2009; p. 30602. [Google Scholar]
- Primack, R.B. Reproductive effort in annual and perennial species of Plantago. Am. Nat. 1979, 114, 51–62. [Google Scholar] [CrossRef]
- Webster, T.M. Weed Survey-Southern States. In Proceedings of the Southern Weed Science Society, San Antonio, TX, USA, 23–25 January 2006; pp. 260–277. Available online: http://www.swss.ws/wp-content/uploads/docs/2006%20Proceedings-SWSS.pdf (accessed on 22 April 2019).
- Webster, T.M. Weed Survey-Southern States. In Proceedings of the Southern Weed Science Society, Little Rock, AR, USA, 25–27 January 2010; pp. 246–257. Available online: http://www.swss.ws/wp-content/uploads/docs/2010%20Proceedings-SWSS.pdf (accessed on 22 April 2019).
- Webster, T.M. Weed Survey-Southern States. In Proceedings of the Southern Weed Science Society, Birmingham, AL, USA, 27–29 January 2014; pp. 282–293. Available online: https://www.swss.ws/wp-content/uploads/2013/09/2014-Proceedings-Final.pdf (accessed on 22 April 2019).
- Mitich, L.W. White man’s foot: Broadleaf plantain. Weed Technol. 1987, 3, 250–251. [Google Scholar] [CrossRef]
- Blom, C. Germination, seedling emergence and establishment of some Plantago species under laboratory and field conditions. Acta Bot. Neerl. 1978, 27, 257–271. [Google Scholar] [CrossRef]
- Cresswell, E.G.; Grime, J.P. Induction of light requirement during seed development and its ecological consequences. Nature 1981, 291, 583–585. [Google Scholar] [CrossRef]
- Gorski, T.; Gorska, K.; Nowicki, J. Germination of seeds of various herbaceous species under leaf canopy. Flora 1977, 166, 249–259. [Google Scholar] [CrossRef]
- Pons, T.L.; van der Toorn, J. Establishment of Plantago lanceolata and Plantago major among grass. Oecologia 1988, 75, 394–399. [Google Scholar] [CrossRef]
- Thompson, K.; Grime, J.P. A comparative study of germination responses to diurnally-fluctuating temperatures. J. Appl. Ecol. 1983, 20, 141–156. [Google Scholar] [CrossRef]
- McCullough, P.E.; Johnston, C.R.; Reed, T.V.; Yu, J. Indaziflam enhances buckhorn plantain (Plantago lanceolata) control from postemergence herbicides. Weed Technol. 2015, 29, 147–153. [Google Scholar] [CrossRef]
- Anonymous. 2,4-D Amine Herbicide Label. 2019. Available online: http://www.cdms.net/ldat/ldEL8000.pdf (accessed on 19 February 2019).
- Mitchem, W.; Grey, T.L. Commercial Pecan Weed Control in Georgia Pest Management Handbook; University of Georgia Cooperative Extension: Athens, GA, USA, 2019; Available online: http://extension.uga.edu/programs-services/integrated-pest-management/publications/handbooks.html#commercial (accessed on 19 February 2019).
- Chen, C.Y.; Nelson, R.L. Variation in early plant height in wild soybean. Crop Sci. 2006, 46, 865–869. [Google Scholar] [CrossRef]
- Knox, P. Georgia Automated Environmental Monitoring Network; University of Georgia: Athens, GA, USA, 2019; Available online: http://www.griffin.uga.edu/bae/ (accessed on 17 August 2019).
- Bouton, J.H.; Woodfield, D.; Caradus, J.R.; Wood, D.T. Registration of ‘Durana’ white clover. Crop Sci. 2005, 45, 797. [Google Scholar] [CrossRef]
- McCurdy, J.D.; McElroy, J.; Flessner, M.L.; Hoyle, J.A.; Parker, E.T. Tolerance of three clovers (Trifolium spp.) to common herbicides. Weed Technol. 2016, 30, 478–485. [Google Scholar] [CrossRef]
- Lacey, E.P. Parental effects in Plantago lanceolata L. I: A growth chamber experiment to examine pre- and postzygotic temperature effects. Evolution 1996, 50, 865–878. [Google Scholar] [CrossRef] [PubMed]
- Lacey, E.P.; Roach, D.A.; Herr, D.; Kincaid, S.; Perrott, R. Multigenerational effects of flowering and fruiting phenology in Plantago lanceolata. Ecology 2003, 84, 2462–2475. [Google Scholar] [CrossRef]
- Lee, K.H.; Jose, S. Soil respiration and microbial biomass in a pecan-cotton alley cropping system in southern US. Agroforest Syst. 2003, 58, 45–54. [Google Scholar] [CrossRef]
- Benvenuti, S.; Macchia, M.; Sergio, M. Quantitative analysis of emergence of seedlings from buried weed seeds with increasing soil depth. Weed Sci. 2001, 49, 528–535. [Google Scholar] [CrossRef]
- Derr, J.F. Broadleaf weed control with sulfonylurea herbicides in cool-season turfgrass. Weed Technol. 2012, 26, 582–586. [Google Scholar] [CrossRef]
- Hancock, D. White Clover Establishment and Management Guide; University of Georgia Cooperative Extension Bulletin 1251: Athens, GA, USA, 2017; Available online: http://www.extension.uga.edu (accessed on 22 April 2019).
- Cardina, J.; Hook, J.E. Factors influencing germination and emergence of Florida beggarweed (Desmodium totruosum). Weed Technol. 1989, 3, 402–407. [Google Scholar] [CrossRef]
- Chatterton, N.; Kadish, A. A temperature gradient germinator. Agron. J. 1969, 61, 643–644. [Google Scholar] [CrossRef]
- Evans, R.A.; Young, J.A.; Kay, B.L. Germination of winter annual species from a rangeland community treated with paraquat. Weed Sci. 1974, 22, 185–187. [Google Scholar] [CrossRef]
- Frans, R.; Talbert, R.; Marx, D.; Crowley, H. Experimental Design and Techniques for Measuring and Analyzing Plant Responses to Weed Control Practices in Research Methods in Weed Science, 3rd ed.; Southern Weed Science Society: Westminster, CO, USA, 1986; pp. 29–46. [Google Scholar]
- Leon, R.G.; Knapp, A.D.; Owen, M.D.K. Effect of temperature on the germination of common waterhemp, giant foxtail, and velvetleaf. Weed Sci. 2004, 52, 67–73. [Google Scholar] [CrossRef]
- Motulsky, H.J.; Christopoulos, A. Fitting models to biological data using linear and nonlinear regression. In A Practical Guide to Curve Fitting; GraphPad Software Inc.: San Diego, CA, USA, 2003; Available online: http://www.graphpad.com (accessed on 30 June 2019).




{kind=link}
{kind=link}
{kind=link}
| Treatment a | Rate | 2,4-D Amine | Greenhouse Experiments b,d | Grove Experiments c,d | |
|---|---|---|---|---|---|
| g ai ha−1 | % control | dry weight mg plant−1 | % control | ||
| Nontreated | - | - | 0 c | 1647 a | 0 c |
| Indaziflam | 37 | 0 | 81 a | 10 e | 82 ab c |
| Indaziflam | 37 | 280 | 99 a | 0 e | 90 a |
| Halosulfuron | 26 | 0 | 56 b | 1456 ab | 48 b |
| Halosulfuron | 26 | 280 | 59 b | 1063 bc | 56 b |
| Simazine | 1120 | 0 | 78 a | 653 cd | 62 b |
| Simazine | 1120 | 280 | 83 a | 240 de | 67 b |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grey, T.L.; Eason, K.M.; Wells, L.; Basinger, N.T. Effects of Temperature on Seed Germination of Plantago lanceolata and Management in Carya illinoinensis Production. Plants 2019, 8, 308. https://doi.org/10.3390/plants8090308
Grey TL, Eason KM, Wells L, Basinger NT. Effects of Temperature on Seed Germination of Plantago lanceolata and Management in Carya illinoinensis Production. Plants. 2019; 8(9):308. https://doi.org/10.3390/plants8090308
Chicago/Turabian StyleGrey, Timothy L., Kayla M. Eason, Lenny Wells, and Nicholas T. Basinger. 2019. "Effects of Temperature on Seed Germination of Plantago lanceolata and Management in Carya illinoinensis Production" Plants 8, no. 9: 308. https://doi.org/10.3390/plants8090308
APA StyleGrey, T. L., Eason, K. M., Wells, L., & Basinger, N. T. (2019). Effects of Temperature on Seed Germination of Plantago lanceolata and Management in Carya illinoinensis Production. Plants, 8(9), 308. https://doi.org/10.3390/plants8090308