Variations of Secondary Metabolites among Natural Populations of Sub-Antarctic Ranunculus Species Suggest Functional Redundancy and Versatility


Abstract
1. Introduction
2. Results
2.1. Populations Differ in Total Metabolite Contents and Metabolite Composition Partly Independently of the Environment
2.2. Environments Partly Explain Variation of Total Metabolite Contents and Metabolite Composition across Populations
2.3. Morphological Phenotypes Partly Explain Variation of Total Metabolite Contents and Metabolite Composition across Populations
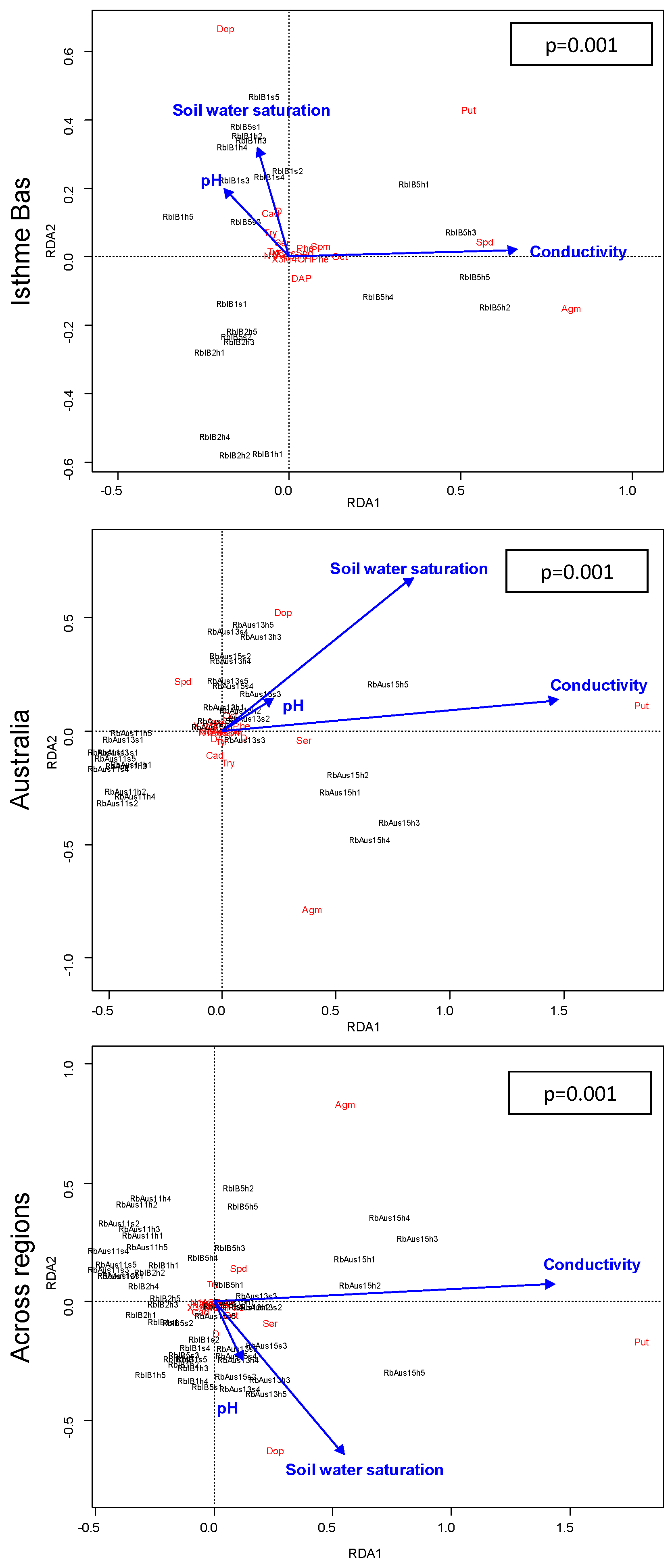
2.4. Regions and Metabolite-Environment Relationships
2.5. Regions and Metabolite-Phenotype Relationships
3. Discussion
3.1. Environments and Morphological Traits Partly Explained Variation of Total Metabolite Contents and Metabolite Composition across Populations
3.2. Environment Did Not Explain All Differences in Total Metabolite Contents and Metabolite Composition among Populations
3.3. Differences in Relationship of Metabolites with Environment or Traits between Regions; Patterns Consistent with Metabolite Redundancy
3.4. Differences in Relationship of Metabolites with Environment or Traits between Regions; Patterns Consistent with Metabolite Versatility
3.5. Metabolite Redundancy and Versatility as a Result of Microevolution Driven by Distance rather than Environment
4. Materials and Methods
4.1. Plant Collection
4.2. Plant Measurements
4.3. Determination and Quantitation of Free Amines and Acetylated Polyamines
4.4. Determination and Quantitation of Quercetins
4.5. Amines Characterized
4.6. Quercetins Characterized
4.7. Environmental Measurements
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wink, M. Evolution of secondary metabolites from an ecological and molecular phylogenetic perspective. Phytochemistry 2003, 64, 3–19. [Google Scholar] [CrossRef]
- Croteau, R.; Kutchan, T.M.; Lewis, N.G. Natural products (secondary metabolites). Biochem. Mol. Biol. Plants 2000, 24, 1250–1319. [Google Scholar]
- Groppa, M.D.; Benavides, M.P. Polyamines and abiotic stress: Recent advances. Amino Acids 2008, 34, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Wahid, A.; Ghazanfar, A. Possible involvement of some secondary metabolites in salt tolerance of sugarcane. J. Plant Physiol. 2006, 163, 723–730. [Google Scholar] [CrossRef] [PubMed]
- Wink, M. Evolution of secondary metabolites in legumes (Fabaceae). S. Afr. J. Bot. 2013, 89, 164–175. [Google Scholar] [CrossRef]
- Ober, D. Seeing double: Gene duplication and diversification in plant secondary metabolism. Trends Plant Sci. 2005, 10, 444–449. [Google Scholar] [CrossRef] [PubMed]
- Tiburcio, A.F.; Altabella, T.; Bitrian, M.; Alcazar, R. The roles of polyamines during the lifespan of plants: From development to stress. Planta 2014, 240, 1–18. [Google Scholar] [CrossRef]
- Hartmann, T. From waste products to ecochemicals: Fifty years research of plant secondary metabolism. Phytochemistry 2007, 68, 2831–2846. [Google Scholar] [CrossRef]
- Tohge, T.; Watanabe, M.; Hoefgen, R.; Fernie, A.R. The evolution of phenylpropanoid metabolism in the green lineage. Crit. Rev. Biochem. Mol. Biol. 2013, 48, 123–152. [Google Scholar] [CrossRef]
- Pathania, S.; Bagler, G.; Ahuja, P.S. Differential Network Analysis Reveals Evolutionary Complexity in Secondary Metabolism of Rauvolfia serpentina over Catharanthus roseus. Front. Plant Sci. 2016, 7, 17. [Google Scholar] [CrossRef]
- Peng, M.; Gao, Y.Q.; Chen, W.; Wang, W.S.; Shen, S.Q.; Shi, J.; Wang, C.; Zhang, Y.; Zou, L.; Wang, S.C.; et al. Evolutionarily Distinct BAHD N-Acyltransferases Are Responsible for Natural Variation of Aromatic Amine Conjugates in Rice. Plant Cell 2016, 28, 1533–1550. [Google Scholar] [CrossRef]
- Xie, L.L.; Liu, P.L.; Zhu, Z.X.; Zhang, S.F.; Zhang, S.J.; Li, F.; Zhang, H.; Li, G.L.; Wei, Y.X.; Sun, R.F. Phylogeny and Expression Analyses Reveal Important Roles for Plant PKS III Family during the Conquest of Land by Plants and Angiosperm Diversification. Front. Plant Sci. 2016, 7, 15. [Google Scholar] [CrossRef]
- Bundy, J.G.; Davey, M.P.; Viant, M.R. Environmental metabolomics: A critical review and future perspectives. Metabolomics 2009, 5, 3–21. [Google Scholar] [CrossRef]
- Moore, B.D.; Andrew, R.L.; Kulheim, C.; Foley, W.J. Explaining intraspecific diversity in plant secondary metabolites in an ecological context. New Phytol. 2014, 201, 733–750. [Google Scholar] [CrossRef]
- Arany, A.M.; de Jong, T.J.; Kim, H.K.; van Dam, N.M.; Choi, Y.H.; Verpoorte, R.; van der Meijden, E. Glucosinolates and other metabolites in the leaves of Arabidopsis thaliana from natural populations and their effects on a generalist and a specialist herbivore. Chemoecology 2008, 18, 65–71. [Google Scholar] [CrossRef]
- Benetis, R.; Radusiene, J.; Janulis, V. Variability of phenolic compounds in flowers of Achillea millefolium wild populations in Lithuania. Med. Lith. 2008, 44, 775–781. [Google Scholar] [CrossRef]
- Davey, M.P.; Burrell, M.M.; Woodward, F.I.; Quick, W.P. Population-specific metabolic phenotypes of Arabidopsis lyrata ssp petraea. New Phytol. 2008, 177, 380–388. [Google Scholar] [CrossRef]
- Kim, J.; Kang, K.; Gonzales-Vigil, E.; Shi, F.; Jones, A.D.; Barry, C.S.; Last, R.L. Striking Natural Diversity in Glandular Trichome Acylsugar Composition Is Shaped by Variation at the Acyltransferase2 Locus in the Wild Tomato Solanum habrochaites. Plant Physiol. 2012, 160, 1854–1870. [Google Scholar] [CrossRef]
- Nkomo, M.M.; Katerere, D.D.R.; Vismer, H.H.F.; Cruz, T.T.; Balayssac, S.S.; Malet-Martino, M.M.; Makunga, N.N.P. Fusarium inhibition by wild populations of the medicinal plant Salvia africana-lutea L. linked to metabolomic profiling. BMC Complement. Altern. Med. 2014, 14, 9. [Google Scholar] [CrossRef]
- El-Bakry, A.A.; Hammad, I.A.; Rafat, F.A. Polymorphism in Calotropis procera: Preliminary genetic variation in plants from different phytogeographical regions of Egypt. Rend. Lincei-Sci. Fis. E Nat. 2014, 25, 471–477. [Google Scholar] [CrossRef]
- Jandova, K.; Dostal, P.; Cajthaml, T.; Kamenik, Z. Intraspecific variability in allelopathy of Heracleum mantegazzianum is linked to the metabolic profile of root exudates. Ann. Bot. 2015, 115, 821–831. [Google Scholar] [CrossRef]
- Nazem, V.; Sabzalian, M.R.; Saeidi, G.; Rahimmalek, M. Essential oil yield and composition and secondary metabolites in self- and open-pollinated populations of mint (Mentha spp.). Ind. Crop. Prod. 2019, 130, 332–340. [Google Scholar] [CrossRef]
- Demasi, S.; Caser, M.; Lonati, M.; Cioni, P.L.; Pistelli, L.; Najar, B.; Scariot, V. Latitude and Altitude Influence Secondary Metabolite Production in Peripheral Alpine Populations of the Mediterranean Species Lavandula angustifolia Mill. Front. Plant Sci. 2018, 9, 983. [Google Scholar] [CrossRef]
- Iwanycki Ahlstrand, N.; Reghev, N.H.; Markussen, B.; Hansen, H.C.B.; Eiriksson, F.F.; Thorsteinsdottir, M.; Ronsted, N.; Barnes, C.J. Untargeted metabolic profiling reveals geography as the strongest predictor of metabolic phenotypes of a cosmopolitan weed. Ecol. Evol. 2018, 8, 6812–6826. [Google Scholar] [CrossRef]
- Davey, M.P.; Woodward, F.I.; Quick, W.P. Intraspecific variation in cold-temperature metabolic phenotypes of Arabidopsis lyrata ssp petraea. Metabolomics 2009, 5, 138–149. [Google Scholar] [CrossRef]
- Hennion, F.; Bouchereau, A.; Gauthier, C.; Hermant, M.; Vernon, P.; Prinzing, A. Variation in amine composition in plant species: How it integrates macroevolutionary and environmental signals. Am. J. Bot. 2012, 99, 36–45. [Google Scholar] [CrossRef]
- Taft, S.; Najar, A.; Godbout, J.; Bousquet, J.; Erbilgin, N. Variations in foliar monoterpenes across the range of jack pine reveal three widespread chemotypes: Implications to host expansion of invasive mountain pine beetle. Front. Plant Sci. 2015, 6, 12. [Google Scholar] [CrossRef]
- Sulmon, C.; van Baaren, J.; Cabello-Hurtado, F.; Gouesbet, G.; Hennion, F.; Mony, C.; Renault, D.; Bormans, M.; El Amrani, A.; Wiegand, C.; et al. Abiotic stressors and stress responses: What commonalities appear between species across biological organization levels? Environ. Pollut. 2015, 202, 66–77. [Google Scholar] [CrossRef]
- Pichersky, E.; Gang, D.R. Genetics and biochemistry of secondary metabolites in plants: An evolutionary perspective. Trends Plant Sci. 2000, 5, 439–445. [Google Scholar] [CrossRef]
- Hanada, K.; Sawada, Y.; Kuromori, T.; Klausnitzer, R.; Saito, K.; Toyoda, T.; Shinozaki, K.; Li, W.-H.; Hirai, M.Y. Functional compensation of primary and secondary metabolites by duplicate genes in Arabidopsis thaliana. Mol. Biol. Evol. 2011, 28, 377–382. [Google Scholar] [CrossRef]
- Lehmann, S.; Funck, D.; Szabados, L.; Rentsch, D. Proline metabolism and transport in plant development. Amino Acids 2010, 39, 949–962. [Google Scholar] [CrossRef]
- Di Ferdinando, M.; Brunetti, C.; Agati, G.; Tattini, M. Multiple functions of polyphenols in plants inhabiting unfavorable Mediterranean areas. Environ. Exp. Bot. 2014, 103, 107–116. [Google Scholar] [CrossRef]
- Wagstaff, S.J.; Hennion, F. Evolution and biogeography of Lyallia and Hectorella (Portulacaceae), geographically isolated sisters from the Southern Hemisphere. Antarct. Sci. 2007, 19, 417–426. [Google Scholar] [CrossRef]
- Bouchereau, A.; Aziz, A.; Larher, F.; Martin-Tanguy, J. Polyamines and environmental challenges: Recent development. Plant Sci. 1999, 140, 103–125. [Google Scholar] [CrossRef]
- Agati, G.; Azzarello, E.; Pollastri, S.; Tattini, M. Flavonoids as antioxidants in plants: Location and functional significance. Plant Sci. 2012, 196, 67–76. [Google Scholar] [CrossRef]
- Hennion, F.; Fiasson, J.L.; Gluchoff-Fiasson, K. Morphological and phytochemical relationships between Ranunculus species from Iles Kerguelen. Biochem. Syst. Ecol. 1994, 22, 533–542. [Google Scholar] [CrossRef]
- Hennion, F.; Litrico, I.; Bartish, I.V.; Weigelt, A.; Bouchereau, A.; Prinzing, A. Ecologically diverse and distinct neighbourhoods trigger persistent phenotypic consequences, and amine metabolic profiling detects them. J. Ecol. 2016, 104, 125–137. [Google Scholar] [CrossRef]
- Alcazar, R.; Marco, F.; Cuevas, J.C.; Patron, M.; Ferrando, A.; Carrasco, P.; Tiburcio, A.F.; Altabella, T. Involvement of polyamines in plant response to abiotic stress. Biotechnol. Lett. 2006, 28, 1867–1876. [Google Scholar] [CrossRef]
- Treutter, D. Significance of flavonoids in plant resistance and enhancement of their biosynthesis. Plant Biol. 2005, 7, 581–591. [Google Scholar] [CrossRef]
- Rott, E.; Gross, E.; Schwienbacher, E. Small-scale heterogeneity of Ranunculus trichophyllus in Lake Tovel (microhabitat, morphology, phenolic compounds and molecular taxonomy). Acta Biol. 2004, 81, 359–367. [Google Scholar]
- Hummel, I.; Quemmerais, F.; Gouesbet, G.; El Amrani, A.; Frenot, Y.; Hennion, F.; Couée, I. Characterization of environmental stress responses during early development of Pringlea antiscorbutica in the field at Kerguelen. New Phytol. 2004, 162, 705–715. [Google Scholar] [CrossRef]
- Van der Putten, N.; Verbruggen, C.; Ochyra, R.; Verleyen, E.; Frenot, Y. Subantarctic flowering plants: Pre-glacial survivors or post-glacial immigrants? J. Biogeogr. 2010, 37, 582–592. [Google Scholar] [CrossRef]
- Hennion, F.; Walton, D.W.H. Ecology and seed morphology of endemic species from Kerguelen Phytogeographic Zone. Polar Biol. 1997, 18, 229–235. [Google Scholar] [CrossRef]
- Hennion, F. Etude des Caractéristiques Biologiques et Génétiques de la Flore Endémique des îles Kerguelen. Ph.D. Thesis, Muséum National d’Histoire Naturelle, Paris, France, 1992. [Google Scholar]
- Lebouvier, M.; Frenot, Y. Conservation and management in the french sub-Antarctic islands and surrounding seas. Pap. Proc. R. Soc. Tasman. 2007, 141, 23–28. [Google Scholar] [CrossRef]
- Lebouvier, M.; Laparie, M.; Hulle, M.; Marais, A.; Cozic, Y.; Lalouette, L.; Vernon, P.; Candresse, T.; Frenot, Y.; Renault, D. The significance of the sub-Antarctic Kerguelen Islands for the assessment of the vulnerability of native communities to climate change, alien insect invasions and plant viruses. Biol. Invasions 2011, 13, 1195–1208. [Google Scholar] [CrossRef]
- Lehnebach, C.A.; Winkworth, R.C.; Becker, M.; Lockhart, P.J.; Hennion, F. Around the pole: Evolution of sub-Antarctic Ranunculus. J. Biogeogr. 2017, 44, 875–886. [Google Scholar] [CrossRef]
- Tiburcio, A.F.; Kaur-Sawhney, R.; Galston, A.W. Polyamine metabolism. In The Biochemistry of Plants, Volume 16; Miflin, B.J., Lea, P.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 283–325. [Google Scholar]
- Fujihara, S.; Yoneyama, T. Endogenous levels of polyamines in the organs of cucumber plant (Cucumis sativus) and factors affecting leaf polyamine contents. Physiol. Plant. 2001, 113, 416–423. [Google Scholar] [CrossRef]
- Hermant, M.; Prinzing, A.; Vernon, P.; Convey, P.; Hennion, F. Endemic species have highly integrated phenotypes, environmental distributions and phenotype–environment relationships. J. Biogeogr. 2013, 40, 1583–1594. [Google Scholar] [CrossRef]
- Foster, S.A.; Walters, D.R. Polyamine concentrations and arginine decarboxylase activity in wheat exposed to osmotic stress. Physiol. Plant. 1991, 82, 185–190. [Google Scholar] [CrossRef]
- Hennion, F.; Frenot, Y.; Martin-Tanguy, J. High flexibility in growth and polyamine composition of the crucifer Pringlea antiscorbutica in relation to environmental conditions. Physiol. Plant. 2006, 127, 212–224. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Lavorel, S.; Garnier, E.; Diaz, S.; Buchmann, N.; Gurvich, D.E.; Reich, P.B.; ter Steege, H.; Morgan, H.D.; van der Heijden, M.G.A.; et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef]
- Smith, M.A.; Davies, P.J. Separation and quantitation of polyamines in plant tissue by high performance liquid chromatography of their dansyl derivatives. Plant Physiol. 1985, 78, 89–91. [Google Scholar] [CrossRef]
- Hayman, A.R.; Gray, D.O.; Evans, S.V. New high performance liquid chromatography system for the separation of biogenic amines as their DNS derivatives. J. Chromatogr. 1985, 325, 462–466. [Google Scholar] [CrossRef]
- Jubault, M.; Hamon, C.; Gravot, A.; Lariagon, C.; Delourme, R.; Bouchereau, A.; Manzanares-Dauleux, M.J. Differential regulation of root arginine catabolism and polyamine metabolism in clubroot-susceptible and partially resistant Arabidopsis genotypes. Plant Physiol. 2008, 146, 2008–2019. [Google Scholar] [CrossRef]
- R Core Team: R: A Language and Environment for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 18 July 2019).
- Terbraak, C.J.F. Canonical correspondence-analysis—A new eigenvector technique for multivariate direct gradient analysis. Ecology 1986, 67, 1167–1179. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology, Volume 24, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2012; p. 1006. [Google Scholar]
- Cabin, R.J.; Mitchell, R.J. To Bonferroni or not to Bonferroni: When and how are the questions. Bull. Ecol. Soc. Am. 2000, 81, 246–248. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amines | Population | |||
| p-Value | F Value | df Residuals | ||
| R. biternatus | across regions | 0.001 | 4.794 | 44 |
| Isthme Bas | 0.004 | 10.773 | 18 | |
| Australia | 0.010 | 6.565 | 24 | |
| R. pseudotrullifolius | across regions | <0.001 | 8.722 | 48 |
| Isthme Bas | >0.500 | 0.073 | 13 | |
| Australia | 0.005 | 6.716 | 24 | |
| R. moseleyi | across regions | 0.001 | 4.962 | 37 |
| Isthme Bas | 0.046 | 6.673 | 13 | |
| Australia | 0.018 | 4.818 | 27 | |
| Quercetins | Population | |||
| p-Value | F Value | df Residuals | ||
| R. biternatus | across regions | <0.001 | 17.883 | 44 |
| Isthme Bas | 0.088 | 4.697 | 18 | |
| Australia | <0.001 | 26.440 | 24 | |
| R. pseudotrullifolius | across regions | <0.001 | 19.441 | 46 |
| Isthme Bas | >0.500 | 0.202 | 22 | |
| Australia | <0.001 | 56.282 | 22 | |
| R. moseleyi | across regions | <0.001 | 78.570 | 36 |
| Isthme Bas | >0.500 | 1.075 | 13 | |
| Australia | <0.001 | 147.176 | 21 | |
| Amines | Population | |||
| p-Value | F-Value | df Residuals | ||
| R. biternatus | across regions | <0.001 | 5.850 | 44 |
| Isthme Bas | ||||
| Australia | <0.001 | 9.554 | 24 | |
| R. pseudotrullifolius | across regions | <0.001 | 11.256 | 48 |
| Isthme Bas | ||||
| Australia | <0.001 | 7.293 | 24 | |
| R. moseleyi | across regions | <0.001 | 11.159 | 48 |
| Isthme Bas | ||||
| Australia | <0.001 | 18.628 | 22 | |
| Quercetins | Population | |||
| p-Value | F Value | df Residuals | ||
| R. biternatus | across regions | <0.001 | 7.254 | 44 |
| Isthme Bas | ||||
| Australia | 0.001 | 16.269 | 24 | |
| R. pseudotrullifolius | across regions | <0.001 | 9.495 | 46 |
| Isthme Bas | ||||
| Australia | <0.001 | 16.336 | 22 | |
| R. moseleyi | across regions | <0.001 | 89.107 | 36 |
| Isthme Bas | ||||
| Australia | <0.001 | 126.020 | 21 | |
| Amines | Overall Environment | Water Saturation | pH | Conductivity | ||||||
| F Value | p-Value | F Value | p-Value | F Value | p-Value | F Value | p-Value | df Residuals | ||
| R. biternatus | across regions | 4.055 | <0.001 | 5.539 | <0.001 | 3.022 | 0.003 | 17.019 | <0.001 | 41 |
| Isthme Bas | 4.383 | 0.361 | 9.575 | 0.249 | 6.470 | 0.301 | 2.660 | 0.452 | 11 | |
| Australia | 5.777 | 0.001 | 15.488 | <0.001 | 2.754 | 0.036 | 9.360 | <0.001 | 13 | |
| R. pseudotrullifolius | across regions | 5.089 | <0.001 | 7.616 | <0.001 | 2.881 | 0.003 | 4.837 | <0.001 | 40 |
| Isthme Bas | 4.811 | 0.008 | 17.536 | <0.001 | 1.810 | 0.172 | 8.924 | <0.001 | 10 | |
| Australia | 4.221 | 0.006 | 17.765 | <0.001 | 2.866 | 0.031 | 4.279 | 0.006 | 13 | |
| R. moseleyi | across regions | 4.006 | <0.001 | 1.186 | 0.334 | 4.103 | <0.001 | 6.392 | <0.001 | 24 |
| Isthme Bas | ||||||||||
| Australia | 11.250 | <0.001 | 6.576 | 0.002 | 7.502 | <0.001 | 19.503 | <0.001 | 11 | |
| Quercetins | Overall Environment | |||||||||
| F Value | p-Value | F Value | p-Value | F Value | p-Value | F Value | p-Value | df Residuals | ||
| R. biternatus | across regions | 4.067 | <0.001 | 2.718 | 0.013 | 2.709 | 0.014 | 4.779 | <0.001 | 43 |
| Isthme Bas | 4.169 | 0.010 | 3.691 | 0.017 | 5.254 | 0.004 | 3.668 | 0.017 | 17 | |
| Australia | 14.812 | <0.001 | 105.210 | <0.001 | 7.995 | 0.002 | 10.625 | <0.001 | 17 | |
| R. pseudotrullifolius | across regions | 1.139 | 0.357 | 5.970 | <0.001 | 3.896 | 0.001 | 2.855 | 0.009 | 47 |
| Isthme Bas | 22.216 | <0.001 | 49.230 | <0.001 | 8.963 | <0.001 | 40.232 | <0.001 | 17 | |
| Australia | 3.017 | 0.022 | 1.714 | 0.158 | 7.465 | <0.001 | 7.088 | <0.001 | 21 | |
| R. moseleyi | across regions | 5.492 | <0.001 | 16.485 | <0.001 | 10.166 | <0.001 | 6.845 | <0.001 | 17 |
| Isthme Bas | 126.530 | <0.001 | 39.861 | <0.001 | 123.590 | <0.001 | 3.425 | 0.049 | 3 | |
| Australia | 56.519 | <0.001 | 74.673 | <0.001 | 40.919 | <0.001 | 23.647 | <0.001 | 5 | |
| Amines | Phenotype | Plant height | Number of Leaves | Flower Size | Leaf Ratio | LDMC | ||||||||
| p Value | F | p Value | F | p Value | F | p Value | F | p Value | F | p Value | F | df Residuals | ||
| R. biternatus | across regions | 0.150 | 1.491 | 0.200 | 1.379 | 0.540 | 0.934 | 0.260 | 1.272 | 0.110 | 1.609 | 0.130 | 1.546 | 41 |
| Isthme Bas | 0.006 | 4.785 | 0.220 | 1.590 | 0.111 | 2.079 | 0.150 | 1.860 | 0.005 | 5.001 | 0.300 | 1.376 | 11 | |
| Australia | 0.350 | 1.243 | 0.570 | 1.041 | 0.700 | 0.763 | 0.230 | 1.508 | 0.560 | 0.455 | 0.340 | 1.262 | 13 | |
| R. pseudotrullifolius | across regions | 0.006 | 2.670 | 0.720 | 0.758 | 0.096 | 1.664 | 0.025 | 2.154 | 0.092 | 1.680 | 0.075 | 1.755 | 40 |
| Isthme Bas | 0.005 | 5.422 | 0.600 | 0.886 | 0.008 | 4.798 | 0.027 | 3.418 | 0.510 | 1.011 | 0.006 | 5.173 | 10 | |
| Australia | 0.012 | 3.621 | 0.760 | 0.692 | 0.005 | 4.413 | 0.050 | 2.515 | 0.009 | 3.871 | 0.006 | 4.240 | 13 | |
| R. moseleyi | across regions | 0.015 | 2.654 | 0.290 | 1.271 | 0.031 | 2.310 | 0.020 | 2.516 | 0.006 | 3.108 | 0.280 | 1.288 | 24 |
| Isthme Bas | ||||||||||||||
| Australia | 0.088 | 2.338 | 0.006 | 5.173 | 0.140 | 1.966 | 0.390 | 1.209 | 0.015 | 4.046 | 0.280 | 1.449 | 11 | |
| Quercetins | Phenotype | Plant height | Number of Leaves | Flower Size | Leaf Ratio | LDMC | ||||||||
| p Value | F | p Value | F | p Value | F | p Value | F | p Value | F | p Value | F | df Residuals | ||
| R. biternatus | across regions | 0.025 | 2.435 | 0.240 | 1.353 | 0.024 | 2.454 | 0.320 | 1.201 | 0.380 | 1.105 | 0.120 | 1.698 | 43 |
| Isthme Bas | 0.475 | 1.004 | 0.180 | 1.661 | 0.084 | 2.174 | 0.690 | 0.713 | 0.920 | 0.395 | 0.910 | 0.412 | 17 | |
| Australia | 0.350 | 1.217 | 0.480 | 0.997 | 0.540 | 0.910 | 0.680 | 0.726 | 0.130 | 1.878 | 0.120 | 1.932 | 17 | |
| R. pseudotrullifolius | across regions | 0.005 | 3.137 | 0.006 | 3.053 | 0.004 | 3.239 | 0.006 | 3.053 | 0.024 | 2.424 | 0.520 | 0.916 | 47 |
| Isthme Bas | 0.006 | 4.221 | 0.240 | 1.470 | 0.024 | 3.080 | 0.010 | 3.780 | 0.190 | 1.626 | 0.570 | 0.869 | 17 | |
| Australia | 0.006 | 3.822 | 0.120 | 1.854 | 0.072 | 2.166 | 0.260 | 1.383 | 0.035 | 2.619 | 0.057 | 2.311 | 21 | |
| R. moseleyi | across regions | 0.006 | 4.221 | 0.009 | 3.869 | 0.089 | 2.135 | 0.005 | 4.384 | 0.004 | 4.588 | 0.120 | 1.932 | 17 |
| Isthme Bas | 0.010 | 99.388 | 0.220 | 3.915 | 0.183 | 4.837 | 0.006 | 166.055 | 0.056 | 17.241 | 0.110 | 8.471 | 3 | |
| Australia | 0.820 | 0.503 | 0.460 | 1.209 | 0.342 | 1.606 | 0.780 | 0.567 | 0.250 | 2.081 | 0.006 | 19.212 | 5 | |
| Interaction Term between Regions and | ||||||||||||
| Amines | (a) Overall Environment | Soil Water Saturation | pH | Conductivity | ||||||||
| p-Value | F | R2 | p-Value | F | R2 | p-Value | F | R2 | p-Value | F | R2 | |
| R. biternatus | 0.059 | 3.723 | 0.213 | 0.157 | 2.07 | 0.116 | 0.291 | 1.138 | 0.007 | 0.608 | 0.267 | 0.52 |
| R. pseudotrullifolius | 0.349 | 0.892 | 0.306 | 0.023 | 5.467 | 0.336 | 0.952 | 0.004 | 0.309 | 0.116 | 2.55 | 0.32 |
| R. moseleyi | 0.111 | 2.647 | 0.527 | <0.001 | 23.706 | 0.317 | 0.068 | 3.522 | 0.375 | 0.051 | 4.048 | 0.55 |
| Quercetins | Overall Environment | Soil Water Saturation | pH | Conductivity | ||||||||
| p-Value | F | R2 | p-Value | F | R2 | p-Value | F | R2 | p-Value | F | R2 | |
| R. biternatus | 0.949 | 0.004 | 0.21 | 0.191 | 1.755 | 0.196 | 0.104 | 2.747 | 0.356 | 0.073 | 3.351 | 0.18 |
| R. pseudotrullifolius | 0.005 | 8.719 | 0.104 | 0.005 | 8.719 | 0.011 | <0.001 | 14.71 | 0.204 | 0.312 | 1.044 | 0.17 |
| R. moseleyi | 0.593 | 0.291 | 0.006 | 0.502 | 0.459 | 0.314 | 0.574 | 0.321 | 0.062 | 0.001 | 11.71 | 0.31 |
| Interaction Term between Regions and | ||||||||||||
| Amines | (b) Phenotype | Plant height | Number of Leaves | |||||||||
| p-Value | F | R2 | p-Value | F | R2 | p-Value | F | R2 | ||||
| R. biternatus | 0.91 | 0 | 0.042 | 0.89 | 0.774 | 0.025 | 0.094 | 4.709 | 0.066 | |||
| R. pseudotrullifolius | 0.99 | 0.029 | 0.26 | 0.31 | 0.203 | 0.27 | 0.001 | 10.82 | 0.39 | |||
| R. moseleyi | 0.046 | 9.388 | 0.35 | 0.042 | 5.76 | 0.32 | 0.41 | 0.048 | 0.26 | |||
| Quercetins | Phenotype | Plant height | Number of Leaves | |||||||||
| p-value | F | R2 | p-value | F | R2 | p-value | F | R2 | ||||
| R. biternatus | 0.63 | 0.005 | 0.008 | 0.76 | 0.757 | 0.022 | 0.61 | 0.152 | 0.17 | |||
| R. pseudotrullifolius | 0.77 | 0.084 | 0.025 | 0.87 | 0.026 | 0.1 | 0.022 | 7.952 | 0.13 | |||
| R. moseleyi | 0.17 | 3.349 | 0.18 | 0.08 | 0.078 | 0.11 | 0.57 | 2.624 | 0.1 | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Labarrere, B.; Prinzing, A.; Dorey, T.; Chesneau, E.; Hennion, F. Variations of Secondary Metabolites among Natural Populations of Sub-Antarctic Ranunculus Species Suggest Functional Redundancy and Versatility. Plants 2019, 8, 234. https://doi.org/10.3390/plants8070234
Labarrere B, Prinzing A, Dorey T, Chesneau E, Hennion F. Variations of Secondary Metabolites among Natural Populations of Sub-Antarctic Ranunculus Species Suggest Functional Redundancy and Versatility. Plants. 2019; 8(7):234. https://doi.org/10.3390/plants8070234
Chicago/Turabian StyleLabarrere, Bastien, Andreas Prinzing, Thomas Dorey, Emeline Chesneau, and Françoise Hennion. 2019. "Variations of Secondary Metabolites among Natural Populations of Sub-Antarctic Ranunculus Species Suggest Functional Redundancy and Versatility" Plants 8, no. 7: 234. https://doi.org/10.3390/plants8070234
APA StyleLabarrere, B., Prinzing, A., Dorey, T., Chesneau, E., & Hennion, F. (2019). Variations of Secondary Metabolites among Natural Populations of Sub-Antarctic Ranunculus Species Suggest Functional Redundancy and Versatility. Plants, 8(7), 234. https://doi.org/10.3390/plants8070234