Deciphering the Genetic Architecture of Plant Height in Soybean Using Two RIL Populations Sharing a Common M8206 Parent



 ,
, 
Abstract
1. Introduction
2. Results
2.1. Evaluation of Phenotypic Variation for Plant Height
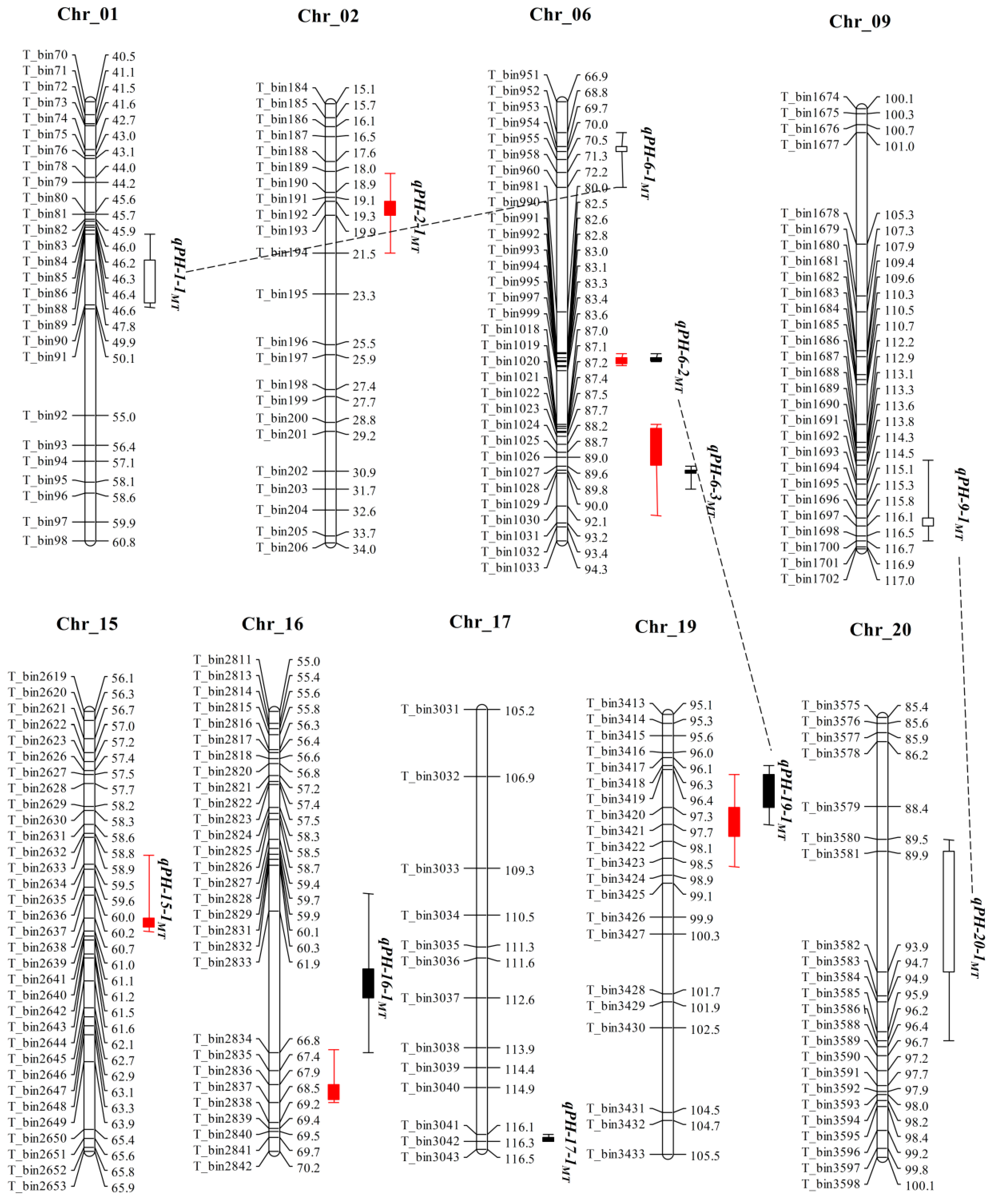
2.2. QTL Mapping for Plant Height by CIM Method in MT and ZM Populations
2.3. Additive Effect QTLs and QTL by Environment Interaction Analysis
2.4. Epistatic Interaction Effects
2.5. QTLs Mapping in Subpopulations of MT and ZM Populations
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Field Experiments
4.2. Phenotypic Data Analysis
4.3. QTL Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| PH | plant height |
| QTL | quantitative trait loci |
| RIL | recombinant inbred line |
| CIM | composite interval mapping |
| MCIM | mixed-model based composite interval mapping |
| MAS | marker-assisted selection |
| SNP | single nucleotide polymorphism |
| JP | Jiangpu experiment station |
| YC | Yancheng experiment station |
| SD | standard deviation |
| CV | coefficient of variation |
| MS | mean square |
| cM | centiMorgan |
| A | additive |
| AA | additive by additive interaction |
| AE | additive by environment interaction |
| AAE | epistatic by environment interaction |
References
- Wilcox, J.R. World Distribution and Trade of Soybean. In Soybeans: Improvement, Production, and Uses, 3rd ed.; American Society of Agronomy: Madison, WI, USA, 2004; pp. 1–14. [Google Scholar]
- Liu, J.; Dou, Y.; Batistella, M.; Challies, E.; Connor, T.; Friis, C.; Millington, J.D.A.; Parish, E.; Romulo, C.L.; Silva, R.F.B.; et al. Spillover systems in a telecoupled Anthropocene: Typology, methods, and governance for global sustainability. Curr. Opin. Environ. Sustain. 2018, 33, 58–69. [Google Scholar] [CrossRef]
- Liu, W.; Kim, M.Y.; Van, K.; Lee, Y.H.; Li, H.; Liu, X.; Lee, S.K. QTL identification of yield-related traits and their association with flowering and maturity in soybean. J. Crop Sci. Biotechnol. 2011, 14, 65–70. [Google Scholar] [CrossRef]
- Lü, H.Y.; Li, H.; Fan, R.; Li, H.; Yin, J.; Zhang, J.; Zhang, D. Genome-wide association study of dynamic developmental plant height in soybean. Can. J. Plant Sci. 2016, 97, 308–315. [Google Scholar] [CrossRef]
- Lee, S.H.; Bailey, M.A.; Mian, M.A.R.; Shipe, E.R.; Ashley, D.A.; Parrott, W.A.; Hussey, R.S.; Boerma, H.R. Identification of quantitative trait loci for plant height, lodging, and maturity in a soybean population segregating for growth habit. Theor. Appl. Genet. 1996, 92, 516–523. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Graef, G.L.; Procopiuk, A.M.; Diers, B.W. Identification of putative QTL that underlie yield in interspecific soybean backcross populations. Theor. Appl. Genet. 2004, 108, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Allen, F.L. Usefulness of plant genome mapping to plant breeding. In Plant Genome Analysis; CRC Press: Boca Raton, FL, USA, 1994; pp. 11–18. [Google Scholar]
- Chapman, A.; Pantalone, V.; Ustun, A.; Allen, F.; Landau-Ellis, D.; Trigiano, R.; Gresshoff, P. Quantitative trait loci for agronomic and seed quality traits in an F2 and F4:6 soybean population. Euphytica 2003, 129, 387–393. [Google Scholar] [CrossRef]
- Mansur, L.M.; Orf, J.H.; Chase, K.; Jarvik, T.; Cregan, P.B.; Lark, K.G. Genetic mapping of agronomic traits using recombinant inbred lines of soybean. Crop Sci. 1996, 36, 1327–1336. [Google Scholar] [CrossRef]
- Orf, J.H.; Chase, K.; Jarvik, T.; Mansur, L.M.; Cregan, P.B.; Adler, F.R.; Lark, K.G. Genetics of soybean agronomic traits: I. Comparison of three related recombinant inbred populations. Crop Sci. 1999, 39, 1642–1651. [Google Scholar] [CrossRef]
- Liu, Y.L.; Li, Y.H.; Reif, J.C.; Mette, M.F.; Liu, Z.X.; Liu, B.; Zhang, S.S.; Yan, L.; Chang, R.Z.; Qiu, L.J. Identification of quantitative trait loci underlying plant height and seed weight in soybean. Plant Genome 2013, 6, 841–856. [Google Scholar] [CrossRef]
- Yin, Z.; Qi, H.; Chen, Q.; Zhang, Z.; Jiang, H.; Zhu, R.; Hu, Z.; Wu, X.; Li, C.; Zhang, Y.; et al. Soybean plant height QTL mapping and meta-analysis for mining candidate genes. Plant Breed. 2017, 136, 688–698. [Google Scholar] [CrossRef]
- Soybase. Available online: http://soybase.org/ (accessed on 7 July 2019).
- Schmutz, J.; Cannon, S.B.; Schlueter, J.; Ma, J.X.; Mitros, T.; Nelson, W.; Hyten, D.L.; Song, Q.J.; Thelen, J.J.; Cheng, J.L.; et al. Genome sequence of the palaeopolyploid soybean. Nature 2010, 465, 120. [Google Scholar] [CrossRef]
- Hyten, D.L.; Song, Q.; Choi, I.Y.; Yoon, M.S.; Specht, J.E.; Matukumalli, L.K.; Nelson, R.L.; Shoemaker, R.C.; Young, N.D.; Cregan, P.B. High-throughput genotyping with the goldengate assay in the complex genome of soybean. Theor. Appl. Genet. 2008, 116, 945–952. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Fan, S.; Yu, F.; Chen, Y.; Zhang, S.; Han, F.; Yan, S.; Wang, L.; Sun, J. High-resolution mapping of QTL for fatty acid composition in soybean using specific-locus amplified fragment sequencing. Theor. Appl. Genet. 2017, 130, 1467–1479. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Li, H.; Wang, J.; Zhang, H.; Hu, Z.; Chu, S.; Lv, H.; Yu, D. High-density genetic mapping identifies new major loci for tolerance to low-phosphorus stress in soybean. Front. Plant. Sci. 2016, 7, 372. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Crouch, J.H. Marker-assisted selection in plant breeding: From publications to practice. Crop Sci. 2008, 48, 391–407. [Google Scholar] [CrossRef]
- Yang, J.; Zhu, J. Methods for predicting superior genotypes under multiple environments based on QTL effects. Theor. Appl. Genet. 2005, 110, 1268–1274. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhu, J.; Li, Z.; Paterson, A.H. Mapping QTLs with epistatic effects and QTL× environment interactions by mixed linear model approaches. Theor. Appl. Genet. 1999, 99, 1255–1264. [Google Scholar] [CrossRef]
- Lai, Y.; Cheng, J.; He, Y.; Yang, B.; Wang, Z.; Zhan, H. Identification of QTLs with additive, epistatic, and QTL× seed maturity interaction effects for seed vigor in rice. Plant. Mol. Biol. Rep. 2016, 34, 160–171. [Google Scholar] [CrossRef]
- Cao, Y.; Li, S.; He, X.; Chang, F.; Kong, J.; Gai, J.; Zhao, T. Mapping QTLs for plant height and flowering time in a Chinese summer planting soybean RIL population. Euphytica 2017, 213, 39. [Google Scholar] [CrossRef]
- Pei, R.; Zhang, J.; Tian, L.; Zhang, S.; Han, F.; Yan, S.; Wang, L.; Li, B.; Sun, J. Identification of novel QTL associated with soybean isoflavone content. Crop J. 2018, 6, 244–252. [Google Scholar] [CrossRef]
- Timmerman-Vaughan, G.M.; Moya, L.; Frew, T.J.; Murray, S.R.; Crowhurst, R. Ascochyta blight disease of pea (Pisum sativum L.): Defence-related candidate genes associated with QTL regions and identification of epistatic QTL. Theor. Appl. Genet. 2016, 129, 879–896. [Google Scholar] [CrossRef] [PubMed]
- Panthee, D.R.; Pantalone, V.R.; Saxton, A.M.; West, D.R.; Sams, C.E. Quantitative trait loci for agronomic traits in soybean. Plant Breed. 2007, 126, 51–57. [Google Scholar] [CrossRef]
- Lee, S.; Jun, T.H.; Michel, A.P.; Mian, M.A.R. SNP markers linked to QTL conditioning plant height, lodging, and maturity in soybean. Euphytica 2015, 203, 521–532. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, W.; Guo, N.; Zhang, Y.; Bu, Y.; Zhao, J.; Xing, H. Combining QTL-seq and linkage mapping to fine map a wild soybean allele characteristic of greater plant height. BMC Genom. 2018, 19, 226. [Google Scholar] [CrossRef] [PubMed]
- Bernard, R.L. Two genes affecting stem termination in soybeans. Crop Sci. 1972, 12, 235–239. [Google Scholar] [CrossRef]
- Liu, B.H.; Watanabe, S.; Uchiyama, T.; Kong, F.J.; Kanazawa, A.; Xia, Z.J.; Nagamatsu, A.; Arai, M.; Yamada, T.; Kitamura, K.; et al. The soybean stem growth habit gene Dt1 is an ortholog of Arabidopsis TERMINAL FLOWER1. Plant Physiol. 2010, 153, 198–210. [Google Scholar] [CrossRef] [PubMed]
- Ping, J.; Liu, Y.; Sun, L.; Zhao, M.; Li, Y.; She, M.; Sui, Y.; Lin, F.; Liu, X.; Tang, Z.; et al. Dt2 is a gain-of-function MADS-domain factor gene that specifies semideterminacy in soybean. Plant Cell 2014, 26, 2831–2842. [Google Scholar] [CrossRef] [PubMed]
- Zou, G.; Zhai, G.; Feng, Q.; Yan, S.; Wang, A.; Zhao, Q.; Shao, J.; Zhang, Z.; Zou, J.; Han, B. Identification of QTLs for eight agronomically important traits using an ultra-high-density map based on SNPs generated from high-throughput sequencing in sorghum under contrasting photoperiods. J. Exp. Bot. 2012, 15, 5451–5462. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Gonzalez, J.J.; Vuong, T.D.; Zhong, R.; Yu, O.; Lee, J.D.; Shannon, G.; Ellersieck, M.; Nguyen, H.T.; Sleper, D.A. Major locus and other novel additive and epistatic loci involved in modulation of isoflavone concentration in soybean seeds. Theor. Appl. Genet. 2011, 123, 1375–1385. [Google Scholar] [CrossRef] [PubMed]
- Shoemaker, R.C.; Specht, J.E. Integration of the soybean molecular and classical genetic linkage groups. Crop Sci. 1995, 35, 436–446. [Google Scholar] [CrossRef]
- Brummer, E.C.; Graef, G.L.; Orf, J.; Wilcox, J.R.; Shoemaker, R.C. Mapping QTL for seed protein and oil content in eight soybean populations. Crop Sci. 1997, 37, 370–378. [Google Scholar] [CrossRef]
- Wang, F.; Guan, C.Y. Molecular mapping and identification of quantitative trait loci for yield components in rapeseed (Brasscianapus L.). Yi Chuan=Hereditas 2010, 32, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Miao, H.; Gu, X.; Zhang, S.; Zhang, Z.; Huang, S.; Wang, Y.; Zhang, Z. Mapping QTLs for fruit-associated traits in Cucumis sativus L. Sci. Agric. Sin. 2011, 44, 5031–5040. [Google Scholar]
- Miao, H.; Gu, X.F.; Zhang, S.P.; Zhang, Z.H.; Huang, S.W.; Wang, Y.; Fang, Z.Y. Mapping QTLs for seedling-associated traits in cucumber. Acta Hortic. Sin. 2012, 39, 879–887. [Google Scholar]
- Wurschum, T.; Maurer, H.P.; Schulz, B.; Mohring, J.; Reif, J.C. Genome-wide association mapping reveals epistasis and genetic interaction networks in sugar beet. Theor. Appl. Genet. 2011, 123, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Yesudas, C.; Bashir, R.; Geisler, M.B.; Lightfoot, D. Identification of germplasm with stacked QTL underlying seed traits in an inbred soybean population from cultivars Essex and Forrest. Mol. Breed. 2013, 31, 693–703. [Google Scholar] [CrossRef]
- Hyten, D.L.; Pantalone, V.R.; Sams, C.E.; Saxton, A.M.; Landau-Ellis, D.; Stefaniak, T.R.; Schmidt, M.E. Seed quality QTL in a prominent soybean population. Theor. Appl. Genet. 2004, 109, 552–561. [Google Scholar] [CrossRef]
- Carlborg, O.; Haley, C.S. Epistasis: Too often neglected in complex trait studies? Nat. Rev. Genet. 2004, 5, 618–625. [Google Scholar] [CrossRef] [PubMed]
- Tajuddin, T.; Watanabe, S.; Yamanaka, N.; Harada, K. Analysis of quantitative trait loci for protein and lipid contents in soybean seeds using recombinant inbred lines. Breed. Sci. 2003, 53, 133–140. [Google Scholar] [CrossRef]
- Gai, J.; Wang, Y.; Wu, X.; Chen, S. A comparative study on segregation analysis and QTL mapping of quantitative traits in plants—with a case in soybean. Front. Agric. China 2007, 1, 1–7. [Google Scholar] [CrossRef]
- Mansur, L.M.; Lark, K.G.; Kross, H.; Oliveira, A. Interval mapping of quantitative trait loci for reproductive, morphological, and seed traits of soybean (Glycine max L.). Theor. Appl. Genet. 1993, 86, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Specht, J.E.; Chase, K.; Macrander, M.; Graefa, G.L.; Chungd, J.; Markwella, J.P.; Germanne, M.; Orff, J.H.; Larkb, K.G. Soybean response to water. Crop Sci. 2001, 41, 493–509. [Google Scholar] [CrossRef]
- Kim, K.S.; Diers, B.W.; Hyten, D.L.; Rouf, M.M.A.; Shannon, J.G.; Nelson, R.L. Identification of positive yield QTL alleles from exotic soybean germplasm in two backcross populations. Theor. Appl. Genet. 2012, 125, 1353–1369. [Google Scholar] [CrossRef] [PubMed]
- Cober, E.R.; Morrison, M.J. Regulation of seed yield and agronomic characters by photoperiod sensitivity and growth habit genes in soybean. Theor. Appl. Genet. 2010, 120, 1005–1012. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.; Watanabe, S.; Yamada, T.; Tsubokura, Y.; Nakashima, H.; Zhai, H.; Anai, T.; Sato, S.; Yamazaki, T.; Lü, S. Positional cloning and characterization reveal the molecular basis for soybean maturity locus E1 that regulates photoperiodic flowering. Proc. Natl. Acad. Sci. USA 2012, 109, E2155–E2164. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.P.; Song, Q.J.; Cregan, P.B.; Nelson, R.L.; Wang, X.Z.; Wu, J.X.; Jiang, G.L. Genome-wide association study for flowering time, maturity dates and plant height in early maturing soybean (Glycine max) germplasm. BMC Genom. 2015, 16, 1. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Xia, Z.; Hideshima, R.; Tsubokura, Y.; Sato, S.; Yamanaka, N.; Takahashi, R.; Anai, T.; Tabata, S.; Kitamura, K.; et al. A map-based cloning strategy employing a residual heterozygous line reveals that the GIGANTEA gene is involved in soybean maturity and flowering. Genetics 2011, 188, 395–407. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Takeshima, R.; Zhu, J.; Xu, M.; Sato, M.; Watanabe, S.; Kanazawa, A.; Liu, B.H.; Kong, F.J.; Yamada, T.; et al. A recessive allele for delayed flowering at the soybean maturity locus E9 is a leaky allele of FT2a, a FLOWERING LOCUS T ortholog. BMC Plant Biol. 2016, 16, 20. [Google Scholar] [CrossRef]
- Nyquist, W.E.; Baker, R.J. Estimation of heritability and prediction of selection response in plant-populations. Crit. Rev. Plant Sci. 1991, 10, 235–322. [Google Scholar] [CrossRef]
- Li, S.; Cao, Y.; He, J.; Zhao, T.; Gai, J. Detecting the QTL-allele system conferring flowering date in a nested association mapping population of soybean using a novel procedure. Theor. Appl. Genet. 2017, 130, 2297–2314. [Google Scholar] [CrossRef]
- Van-Ooijen, J.W. JoinMap® 4, Software for the Calculation of Genetic Linkage Maps in Experimental Populations; Kyazma BV: Wageningen, The Netherlands, 2006; Volume 33. [Google Scholar]
- Kosambi, D.D. The estimation of map distances from recombination values. In D.D. Kosambi, Selected Works in Mathematics and Statistics; Springer: New Delhi, India, 2016; pp. 125–130. [Google Scholar]
- Wang, S.; Basten, C.; Zeng, Z. Windows QTL Cartographer 2.5; Department of Statistics, North Carolina State University: Raleigh, NC, USA, 2007. [Google Scholar]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Hu, C.C.; Hu, H.; Yu, R.D.; Xia, Z.; Ye, X.Z.; Zhu, J. QTLNetwork: Mapping and visualizing genetic architecture of complex traits in experimental populations. Bioinformatics 2008, 24, 721–723. [Google Scholar] [CrossRef] [PubMed]
- McCouch, S.; Cho, Y.; Yano, M.; Paul, E.; Blinstrub, M.; Morishima, H.; Kinoshita, T. Report on QTL nomenclature. Rice Genet. Newsl. 1997, 14, 11–13. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Population | Environment | Parents | RILs | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| M8206 | Tongshan | Zhengyang | Mean | SD | Range | Skewness | Kurtosis | CV (%) | h2 (%) | ||
| MT | 2012JP | 62.1 ± 3.1 | 73.3 ± 8.3 | - | 67.6 | 18.0 | 19.5–126.8 | 0.4 | 0.4 | 26.6 | 92.3 |
| 2014JP | 67.8 ± 7.4 | 75.4 ± 11.1 | - | 89.0 | 26.2 | 17.8–163.1 | 0.1 | 0.2 | 29.5 | ||
| 2014YC | 58.2 ± 8.4 | 75.0 ± 5.0 | - | 76.3 | 15.6 | 22.3–112.3 | −0.9 | 2.0 | 20.4 | ||
| ZM | 2012JP | 62.1 ± 3.1 | - | 76.2 ± 11.1 | 80.9 | 19.0 | 34.0–130.3 | 0.1 | −0.4 | 23.5 | 92.5 |
| 2014JP | 67.8 ± 7.4 | - | 75.7 ± 6.7 | 93.4 | 21.3 | 43.0–167.1 | 0.7 | 1.0 | 22.8 | ||
| 2014YC | 58.2 ± 8.4 | - | 77.7 ± 13.7 | 71.7 | 11.5 | 42.3–101.3 | 0.4 | 0.2 | 16.0 | ||
| Population | QTL a | Chr. | Position (cM) | Marker Interval | LOD b | Confidence interval (cM) c | Physical Region (Mb) d | A e | R2 (%) f | Env. | ReportedQTL/Gene g |
|---|---|---|---|---|---|---|---|---|---|---|---|
| MT | qPH-2-1MT (QPH-2) | 2 | 19.3 | T_bin192–T_bin193 | 3.8 | 18.1–21.5 | 4.5–5.2 | −2.7 | 3.0 | 2014YC | Plant height 13-1 |
| qPH-6-2MT | 6 | 82.8 | T_bin992–T_bin993 | 28.6 | 82.6–83.0 | 19.1–21.9 | −9.8 | 29.3 | 2014YC | E1 | |
| 83.0 | T_bin994–T_bin995 | 26.9 | 82.7–83.2 | −13.9 | 19.4 | 2014JP | |||||
| 83.0 | T_bin994–T_bin995 | 14.1 | 82.7–83.3 | −7.8 | 13.7 | 2012JP | |||||
| qPH-6-3MT | 6 | 87.1 | T_bin1020–T_bin1021 | 26.4 | 87.0–87.4 | 42.9–45.4 | −12.7 | 19.2 | 2014JP | Plant height 3-2 | |
| (QPH-6) | 89.8 | T_bin1028–T_bin1029 | 13.3 | 89.6–92.7 | −7.0 | 13.0 | 2012JP | ||||
| qPH-15-1MT | 15 | 60.7 | T_bin2638–T_bin2639 | 3.2 | 59.3–61.0 | 14.4–15.4 | 3.6 | 1.8 | 2014JP | Novel | |
| qPH-16-1MT | 16 | 67.9 | T_bin2836–T_bin2837 | 3.4 | 66.7–68.5 | 30.3–30.8 | −2.5 | 2.7 | 2014JP | E9 | |
| qPH-19-1MT | 19 | 97.7 | T_bin3421–T_bin3422 | 23.8 | 97.0–98.7 | 44.9–45.5 | 7.6 | 22.2 | 2014YC | Dt1 | |
| (QPH-19) | 97.3 | T_bin3420–T_bin3421 | 55.7 | 96.5–97.8 | 19.4 | 50.7 | 2014JP | ||||
| 97.3 | T_bin3420–T_bin3421 | 31.2 | 96.5–97.9 | 11.0 | 33.9 | 2012JP | |||||
| ZM | qPH-5-1ZM | 5 | 51.4 | Z_bin554–Z_bin555 | 2.9 | 48.9–53.4 | 32.1–32.9 | −2.9 | 6.1 | 2014JP | Plant height 26-1 |
| qPH-6-2ZM | 6 | 117.7 | Z_bin718–Z_bin719 | 6.5 | 117.3–118.7 | 37.8–43.9 | −7.1 | 11.3 | 2012JP | Plant height 3-2 | |
| (QPH-6) | 118.6 | Z_bin719–Z_bin720 | 7.5 | 117.3–122.8 | −8.6 | 12.7 | 2014JP | ||||
| qPH-7-1ZM | 7 | 100.2 | Z_bin879–Z_bin880 | 3.0 | 98.6–101.0 | 38.0–38.6 | 3.0 | 6.5 | 2014YC | Plant height 37-5 | |
| qPH-9-1ZM | 9 | 110.6 | Z_bin1195–Z_bin1196 | 4.6 | 109.7–111.5 | 40.7–42.4 | −5.3 | 7.6 | 2012JP | Plant height 33-5 | |
| 108.6 | Z_bin1192–Z_bin1193 | 2.8 | 106.6–111.9 | −4.3 | 3.9 | 2014JP | |||||
| qPH-10-1ZM | 10 | 104.2 | Z_bin1330–Z_bin1331 | 3.4 | 102.7–106.8 | 43.6–44.4 | 4.8 | 5.5 | 2012JP | E2 | |
| 104.2 | Z_bin1330–Z_bin1331 | 4.5 | 101.7–105.1 | 6.1 | 7.2 | 2014JP | |||||
| qPH-17-1ZM (QPH-17) | 17 | 134.1 | Z_bin2179–Z_bin2180 | 2.7 | 126.0–135.1 | 40.3–41.9 | −4.1 | 4.2 | 2012JP | Novel | |
| 134.1 | Z_bin2179–Z_bin2180 | 5.1 | 132.7–135.1 | −6.4 | 8.2 | 2014JP | |||||
| 127.7 | Z_bin2175–Z_bin2176 | 3.8 | 126.9–128.6 | −3.5 | 9.0 | 2014YC | |||||
| qPH-18-1ZM | 18 | 88.3 | Z_bin2283–Z_bin2284 | 4.0 | 85.1–91.0 | 53.5–54.6 | 3.6 | 9.3 | 2014YC | Dt2 | |
| qPH-19-1ZM | 19 | 108.8 | Z_bin2431–Z_bin2432 | 15.0 | 108.4–109.5 | 44.6–45.9 | 11.4 | 30.9 | 2012JP | Dt1 | |
| (QPH-19) | 108.8 | Z_bin2431–Z_bin2432 | 10.8 | 108.2–110.1 | 10.2 | 19.7 | 2014JP |
| Population | QTL | Chr. | Marker Interval | Position (cM) | Confidence Interval (cM) | A a | R2 (A) (%) b | AE c | R2 (AE) (%) d | Physical Interval (Mb) | ReportedQTL/Gene e |
|---|---|---|---|---|---|---|---|---|---|---|---|
| MT | qPH-6-2MT | 6 | T_bin992–T_bin993 | 82.8 | 82.6–83.1 | −7.5 ** | 13.5 | −2.5 ** (AE2)/3.1 ** (AE3) | 1.9 | 19.1–21.3 | E1 |
| qPH-6-3MT (QPH-6) | 6 | T_bin1028–T_bin1029 | 89.8 | 89.6–91.0 | −3.7 ** | 3.2 | 44.1–45.0 | Plant height 20-3 | |||
| qPH-16-1MT | 16 | T_bin2833–T_bin2834 | 63.9 | 61.3–66.8 | −3.0 ** | 2.1 | 29.5–30.3 | E9 | |||
| qPH-17-1MT(QPH-17) | 17 | T_bin3042–T_bin3043 | 116.3 | 116.1–116.3 | −2.7 ** | 1.7 | 40.8–41.8 | Novel | |||
| qPH-19-1MT (QPH-19) | 19 | T_bin3419–T_bin3420 | 96.4 | 96.3–97.7 | 12.9 ** | 39.3 | −3.4 ** (AE1)/6.3 ** (AE2)/−2.7 ** (AE3) | 6.9 | 44.7–45.3 | Dt1 | |
| ZM | qPH-5-1ZM | 5 | Z_bin554–Z_bin555 | 53.4 | 49.9–55.6 | −2.6 ** | 2.1 | 32.1–33.3 | Plant height 26-1 | ||
| qPH-6-2ZM | 6 | Z_bin723–Z_bin724 | 123.1 | 122.0–124.4 | −4.8 ** | 7.2 | −1.8 * (AE2) | 0.9 | 43.8–44.23 | Plant height 20-3 | |
| qPH-10-1ZM | 10 | Z_bin1334–Z_bin1335 | 106.4 | 103.3–106.8 | 4.9 ** | 7.6 | −1.8 * (AE3) | 1.0 | 43.8–44.4 | E2 | |
| qPH-17-1ZM(QPH-17) | 17 | Z_bin2179–Z_bin2180 | 134.1 | 132.7–135.5 | −4.9 ** | 7.6 | −1.7 * (AE2) | 0.7 | 41.0–41.9 | Novel | |
| qPH-18-1ZM | 18 | Z_bin2283–Z_bin2284 | 87.3 | 86.7–90.3 | 4.2 ** | 5.5 | 53.5–54.5 | Dt2 | |||
| qPH-19-1ZM (QPH-19) | 19 | Z_bin2434–Z_bin2435 | 111.5 | 109.8–111.6 | 6.3 ** | 12.5 | −4.3 ** (AE1)/2.0 * (AE2)/2.4 * (AE3) | 4.5 | 44.8–45.9 | Dt1 |
| Population | Pair | QTL | Chr. | Position (cM) | Marker Interval | Confidence Interval (cM) | Physical Interval (Mb) | AA a | R2 (AA) (%) b | AAE c | R2 (AAE) (%) d |
|---|---|---|---|---|---|---|---|---|---|---|---|
| MT | 1 | qPH-6-2MT | 6 | 82.8 | T_bin992–T_bin993 | 82.6–83.1 | 19.1–21.3 | 1.1 * | 0.3 | 2.7 ** (AE1)/ | 1.6 |
| qPH-19-1MT | 19 | 96.4 | T_bin3419–T_bin3420 | 96.3–97.7 | 44.7–45.3 | −2.4 ** (AE3) | |||||
| 2 | qPH-1-1MT | 1 | 49.8 | T_bin89–T_bin90 | 46.6–49.9 | 42.5–46.0 | −2.6 ** | 1.6 | |||
| qPH-6-1MT | 6 | 69.7 | T_bin953–T_bin954 | 68.8–72.2 | 14.9–15.6 | ||||||
| 3 | qPH-9-1MT | 9 | 115.8 | T_bin1696–T_bin1697 | 113.6–116.7 | 45.3–46.2 | −2.6 ** | 1.6 | |||
| qPH-20-1MT | 20 | 91.9 | T_bin3581–T_bin3582 | 89.5–96.2 | 42.0–43.9 | ||||||
| ZM | 1 | qPH-5-1ZM | 5 | 53.4 | Z_bin554–Z_bin555 | 49.9–55.6 | 32.1–33.3 | −2.3 ** | 1.6 | ||
| qPH-17-1ZM | 17 | 134.1 | Z_bin2179–Z_bin2180 | 132.7–135.5 | 41.0–41.9 | ||||||
| 2 | qPH-1-1ZM | 1 | 28.3 | Z_bin22–Z_bin23 | 24.1–31.6 | 3.2–4.4 | −2.2 ** | 1.5 | |||
| qPH-6-1ZM | 6 | 108.5 | Z_bin701–Z_bin702 | 106.0–111.2 | 16.2–18.0 | ||||||
| 3 | qPH-2-2ZM | 2 | 49.5 | Z_bin165–Z_bin166 | 48.6–50.2 | 6.8–8.2 | −3.3 ** | 3.4 | −2.2 * (AE3) | 1.1 | |
| qPH-4-1ZM | 4 | 61.7 | Z_bin433–Z_bin434 | 60.3–65.6 | 8.7–11.0 |
| Sub-Populations a | QTL | Chr. | Position (cM) b | Flanking Marker | LOD | Physical Interval (Mb) | Additive Effect | R2 (%) | Env. | Reported QTL/Gene c |
|---|---|---|---|---|---|---|---|---|---|---|
| MT-D | qPH-6-2MT | 6 | 82.8 | T_bin992-T_bin993 | 22.2 | 18.8–21.9 | −15.4 | 49.6 | 2014YC | E1 |
| 82.8 | T_bin992-T_bin993 | 15.8 | −14.3 | 35.7 | 2014JP | |||||
| 83.1 | T_bin994-T_bin995 | 6.5 | −6.9 | 16.7 | 2012JP | |||||
| qPH-16-1MT | 16 | 67.9 | T_bin2836-T_bin2837 | 3.1 | 29.0–31.0 | −3.5 | 7.2 | 2012JP | E9 | |
| 68.5 | T_bin2837-T_bin2838 | 4.6 | −4.2 | 6.2 | 2014YC | |||||
| 69.5 | T_bin2840-T_bin2841 | 4.2 | −5.0 | 7.4 | 2014JP | |||||
| MT-I | qPH-6-2MT | 6 | 83.1 | T_bin994-T_bin995 | 13.7 | 19.3–21.9 | −6.4 | 21.4 | 2014YC | E1 |
| 83.0 | T_bin993-T_bin994 | 15.5 | −12.5 | 28.8 | 2014JP | |||||
| qPH-6-3MT | 6 | 86.9 | T_bin1016-T_bin1018 | 13.6 | 41.3–43.8 | −6.2 | 21.3 | 2014YC | Plant height 20-3 | |
| (QPH-6) | 87.5 | T_bin1022-T_bin1023 | 15.8 | −12.1 | 29.2 | 2014JP | ||||
| 87.5 | T_bin1022-T_bin1023 | 9.4 | −7.4 | 18.3 | 2012JP | |||||
| qPH-15-2MT | 15 | 74.3 | T_bin2691-T_bin2692 | 5.4 | 43.5–47.0 | 3.6 | 7.8 | 2014YC | Plant height26-10 | |
| qPH-17-1MT (QPH-17) | 17 | 114.9 | T_bin3040-T_bin3041 | 3.4 | 40.3–41.8 | −4.2 | 5.9 | 2012JP | Novel | |
| ZM-I | qPH-2-1ZM (QPH-2) | 2 | 30.0 | Z_bin145-Z_bin146 | 8.3 | 4.6–5.2 | −5.7 | 23.5 | 2014YC | Plant height13-1 |
| qPH-4-2ZM | 4 | 74.1 | Z_bin451-Z_bin452 | 3.8 | 37.0–40.7 | −5.4 | 11.0 | 2012JP | Novel | |
| qPH-6-2ZM | 6 | 125.3 | Z_bin726-Z_bin727 | 10.7 | 37.8–45.2 | −12.8 | 34.1 | 2014JP | Plant height 20-3 | |
| (QPH-6) | 121.1 | Z_bin721-Z_bin722 | 7.3 | −5.6 | 22.2 | 2014YC | ||||
| 118.6 | Z_bin725-Z_bin726 | 5.0 | −6.4 | 14.8 | 2012JP | |||||
| ZM-D | qPH-10-1ZM | 10 | 95.3 | Z_bin1320-Z_bin1321 | 8.7 | 42.2–43.8 | 10.7 | 35.2 | 2012JP | E2 |
| 98.6 | Z_bin1325-Z_bin1326 | 4.2 | 6.4 | 13.5 | 2014JP | |||||
| qPH-13-1ZM | 13 | 42.4 | Z_bin1598-Z_bin1599 | 3.2 | 20.3–21.4 | −6.4 | 13.8 | 2014JP | Plant height37-8 | |
| 42.4 | Z_bin1598-Z_bin1599 | 4.0 | −6.7 | 14.1 | 2012JP | |||||
| qPH-17-1ZM | 17 | 134.1 | Z_bin2179-Z_bin2180 | 6.5 | 40.2–41.9 | −9.4 | 24.0 | 2014JP | Novel | |
| (QPH-17) | 128.1 | Z_bin2176-Z_bin2177 | 8.2 | −6.5 | 30.9 | 2014YC | ||||
| 134.1 | Z_bin2179-Z_bin2180 | 3.2 | −6.0 | 9.4 | 2012JP | |||||
| qPH-20-1ZM | 20 | 51.7 | Z_bin2531-Z_bin2532 | 3.8 | 35.5–37.2 | 4.1 | 11.5 | 2014YC | Plant height16-1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, Y.; Li, S.; Chen, G.; Wang, Y.; Bhat, J.A.; Karikari, B.; Kong, J.; Gai, J.; Zhao, T. Deciphering the Genetic Architecture of Plant Height in Soybean Using Two RIL Populations Sharing a Common M8206 Parent. Plants 2019, 8, 373. https://doi.org/10.3390/plants8100373
Cao Y, Li S, Chen G, Wang Y, Bhat JA, Karikari B, Kong J, Gai J, Zhao T. Deciphering the Genetic Architecture of Plant Height in Soybean Using Two RIL Populations Sharing a Common M8206 Parent. Plants. 2019; 8(10):373. https://doi.org/10.3390/plants8100373
Chicago/Turabian StyleCao, Yongce, Shuguang Li, Guoliang Chen, Yanfeng Wang, Javaid Akhter Bhat, Benjamin Karikari, Jiejie Kong, Junyi Gai, and Tuanjie Zhao. 2019. "Deciphering the Genetic Architecture of Plant Height in Soybean Using Two RIL Populations Sharing a Common M8206 Parent" Plants 8, no. 10: 373. https://doi.org/10.3390/plants8100373
APA StyleCao, Y., Li, S., Chen, G., Wang, Y., Bhat, J. A., Karikari, B., Kong, J., Gai, J., & Zhao, T. (2019). Deciphering the Genetic Architecture of Plant Height in Soybean Using Two RIL Populations Sharing a Common M8206 Parent. Plants, 8(10), 373. https://doi.org/10.3390/plants8100373