Combined Boron Toxicity and Salinity Stress—An Insight into Its Interaction in Plants




{kind=link}
{kind=link}
Abstract
1. Introduction
2. Combined Stress on Plants—A Major Problem of Today’s World
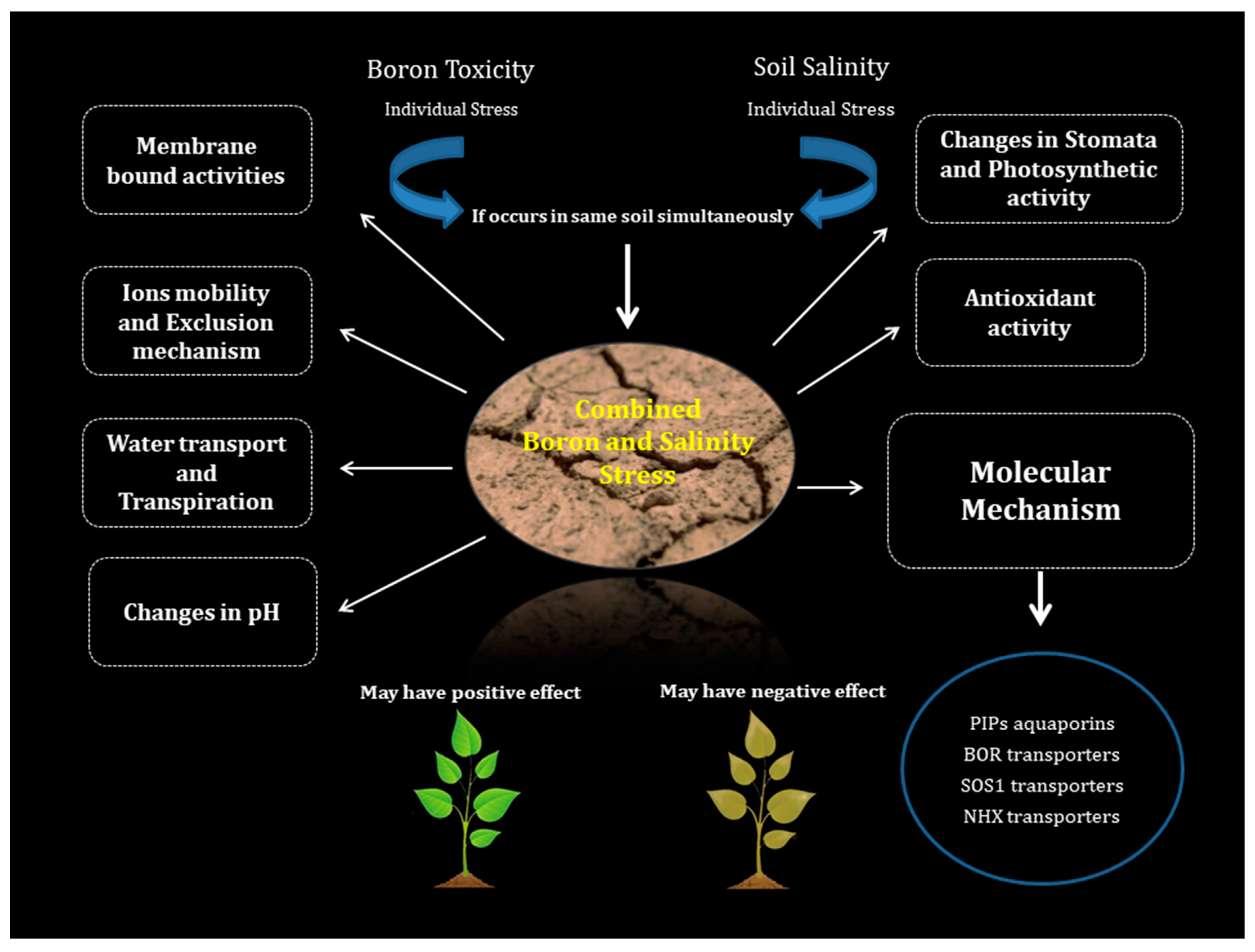
3. Prevalence of BorSal as a Combined Stress Condition
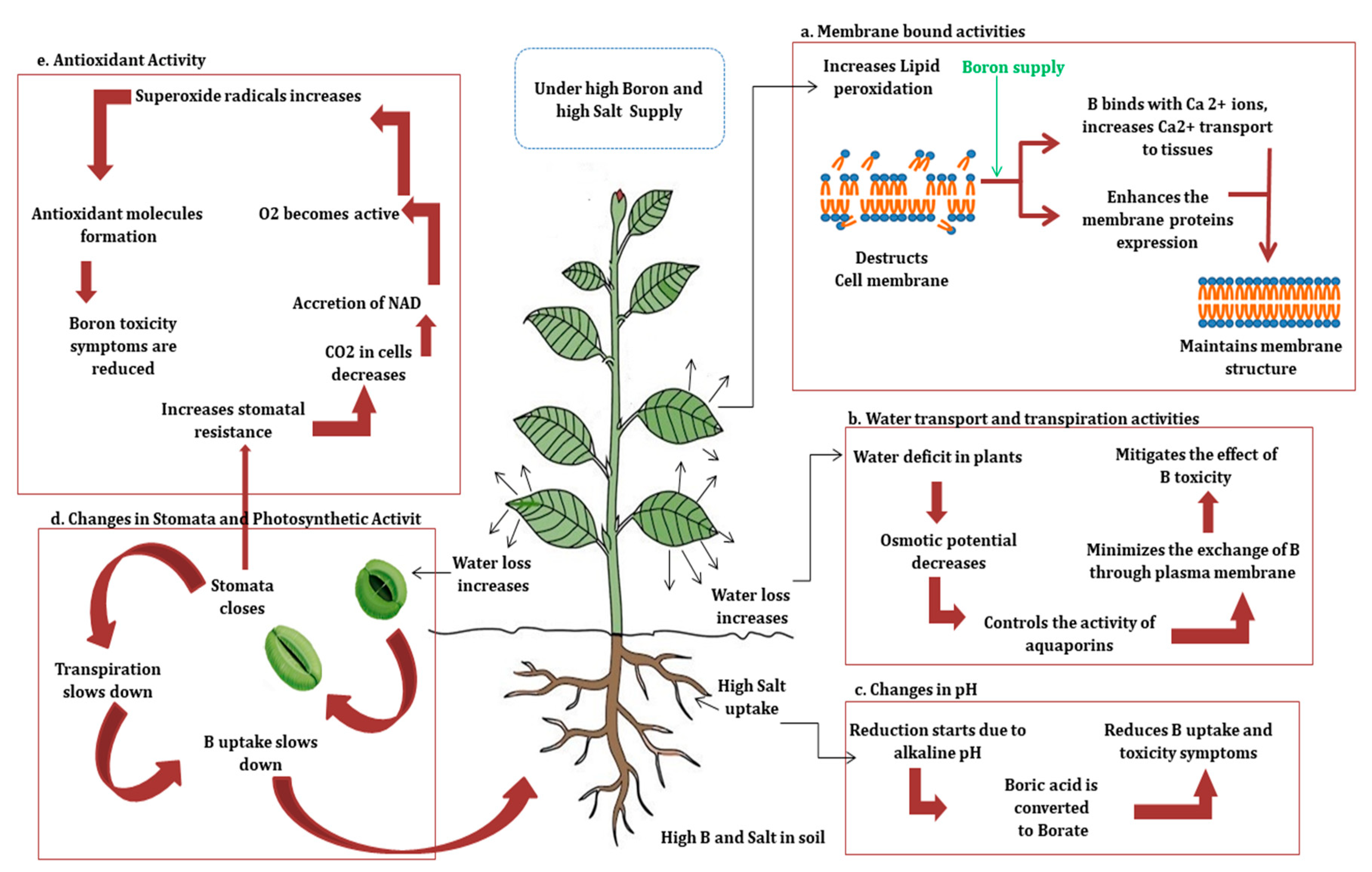
4. BorSal Interaction in Plants in Terms of Membrane Bound Activities
5. BorSal Interaction in Plants in Terms of Ion Mobility and Exclusion Mechanism
6. BorSal Interaction in Plants in Terms of Water Transport, Transpirational Movement, Photosynthetic Activity, and Changes in Stomatal Resistance
7. BorSal Interaction in Plants in Terms of Antioxidant Activity
8. BorSal Interaction in Plants in Terms of pH Changes
9. Molecular Mechanism Involved in B Stress as an Individual Stress Condition
10. Molecular Mechanism Involved in Salinity Stress as an Individual Stress Condition
11. Common Transporters Involved in BorSal Stress Condition
12. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zhu, J.K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef] [PubMed]
- Senthil-Kumar, M. Plant Tolerance to Individual and Concurrent Stresses; Springer: Boston, MA, USA, 2017. [Google Scholar]
- Kissoudis, C.; van de Wiel, C.; Visser, R.G.; van der Linden, G. Enhancing crop resilience to combined abiotic and biotic stress through the dissection of physiological and molecular crosstalk. Front. Plant Sci. 2014, 5, 207. [Google Scholar] [CrossRef] [PubMed]
- Reid, R.J. Boron Toxicity and Tolerance in Crop Plants. In Crop Improvement under Adverse Conditions; Springer: New York, NY, USA, 2013; pp. 333–346. [Google Scholar]
- Izadi, M.H.; Rabbani, J.; Emam, Y.; Pessarakli, M.; Tahmasebi, A. Effects of Salinity Stress on Physiological Performance of Various Wheat and Barley Cultivars. J. Plant Nutr. 2014, 37, 520–531. [Google Scholar] [CrossRef]
- Cartwright, B.; Zarcinas, B.; Mayfield, A. Toxic concentrations of boron in a red-Brown earth at Gladstone, South Australia. Soil Res. 1984, 22, 261–272. [Google Scholar] [CrossRef]
- Kalayci, M.; Alkan, A.; Cakmak, I.; Bayramoğlu, O.; Yilmaz, A.; Aydin, M.; Ozbek, V.; Ekiz, H.; Ozberisoy, F. Studies on differential response of wheat cultivars to boron toxicity. Euphytica 1998, 100, 123–129. [Google Scholar] [CrossRef]
- Nable, R.O.; Bañuelos, G.S.; Paull, J.G. Boron toxicity. Plant Soil 1997, 193, 181–198. [Google Scholar] [CrossRef]
- Takkar, P. Micronutrients-Forms, contents, distribution in profile, indices of availability and soil test methods. In Proceedings of the 12th International Congress of Soil Science, New Delhi, India, 8–16 February 1982. [Google Scholar]
- Gezgin, S.; Dursun, N.; Hamurcu, M.; Harmankaya, M.; Önder, M.; Sade, B.; Topal, A.; Soylu, S.; Akgün, N.; Yorgancilar, M.; et al. Boron Content of Cultivated Soils in Central-Southern Anatolia and its Relationship with Soil Properties and Irrigation Water Quality. In Boron in Plant and Animal Nutrition; Springer: Boston, MA, USA, 2002. [Google Scholar]
- Cartwright, B.; Rathjen, A.; Sparrow, D.; Paull, J.; Zarcinas, B. Boron tolerance in Australian varieties of wheat and barley. In Genetic Aspects of Plant Mineral Nutrition; Springer: Dordrecht, The Netherlands, 1987; pp. 139–151. [Google Scholar]
- Ravikovitch, S.; Margolin, M.; Navrot, J. Microelements in soils of Israel. Soil Sci. 1961, 92, 85–89. [Google Scholar] [CrossRef]
- Shorrocks, V. Boron toxicity in Hevea brasiliensis. Nature 1964, 204, 599–600. [Google Scholar] [CrossRef]
- Cáceres, V.L.; Gruttner, D.E.; Contreras, N.R. Water recycling in arid regions: Chilean case. Ambio 1992, 21, 138–144. [Google Scholar]
- Zyrin, N.; Zborishchuk, J. Boron in the ploughed layer of soils of the European part of the USSR. Pochvovedene 1975, 5, 44–51. [Google Scholar]
- Chesworth, W. Geochemistry of Micronutrients. In Micronutrients in Agriculture, Mortvedt, J.J., Ed.; Soil Science Society of America: Madison, WI, USA, 1991; pp. 1–30. [Google Scholar]
- Ghassemi, F.; Jakeman, A.J.; Nix, H.A. Salinisation of Land and Water Resources: Human Causes, Extent, Management and Case Studies; CAB international: Wallingford, UK, 1995. [Google Scholar]
- Smith, T.E.; Grattan, S.R.; Grieve, C.M.; Poss, J.A.; Läuchli, A.E.; Suarez, D.L. pH dependent salinity-Boron interactions impact yield, biomass, evapotranspiration and boron uptake in broccoli (Brassica oleracea L.). Plant Soil 2013, 370, 541–554. [Google Scholar] [CrossRef]
- Smith, T.E.; Grattan, S.R.; Grieve, C.M.; Poss, J.A.; Suarez, D.L. Salinity’s influence on boron toxicity in broccoli: II. Impacts on boron uptake, uptake mechanisms and tissue ion relations. Agric. Water Manag. 2010, 97, 783–791. [Google Scholar] [CrossRef]
- Grieve, C.; Poss, J.; Grattan, S.; Suarez, D.; Smith, T. The combined effects of salinity and excess boron on mineral ion relations in broccoli. Sci. Hortic. 2010, 125, 179–187. [Google Scholar] [CrossRef]
- Martinez-Ballesta Mdel, C.; Bastias, E.; Zhu, C.; Schaffner, A.R.; Gonzalez-Moro, B.; Gonzalez-Murua, C.; Carvajal, M. Boric acid and salinity effects on maize roots. Response of aquaporins ZmPIP1 and ZmPIP2, and plasma membrane H+-ATPase, in relation to water and nutrient uptake. Physiol. Plant 2008, 132, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Bastias, E.; Fernández-García, N.; Carvajal, M. Aquaporin functionality in roots of Zea mays in relation to the interactive effects of boron and salinity. Plant Biol. 2004, 6, 415–421. [Google Scholar] [CrossRef]
- Javid, M.; Ford, R.; Norton, R.; Nicolas, M. Sodium and boron exclusion in two Brassica juncea cultivars exposed to the combined treatments of salinity and boron at moderate alkalinity. Biologia 2014, 69, 1157–1163. [Google Scholar] [CrossRef]
- Wimmer, M.A.; Muhling, K.H.; Lauchli, A.; Brown, P.H.; Goldbach, H.E. The interaction between salinity and boron toxicity affects the subcellular distribution of ions and proteins in wheat leaves. Plant Cell Environ. 2003, 26, 1267–1274. [Google Scholar] [CrossRef]
- Bañuelos, G.S. Coping with naturally-high levels of soil salinity and boron in the westside of central california. Perspect. Sci. 2015, 3, 4–6. [Google Scholar] [CrossRef][Green Version]
- Naz, T.; Akhtar, J.; Iqbal, M.M.; ul Haq, M.A.; Saqib, M. Boron toxicity in salt-Affected soils and effects on plants. In Soil Science: Agricultural and Environmental Prospectives; Springer: Cham, Switzerland, 2016; pp. 259–286. [Google Scholar]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and biotic stress combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef]
- Mahalingam, R. Consideration of combined stress: A crucial paradigm for improving multiple stress tolerance in plants. In Combined Stresses in Plants; Springer: Cham, Switzerland, 2015; pp. 1–25. [Google Scholar]
- Ramegowda, V.; Senthil-Kumar, M. The interactive effects of simultaneous biotic and abiotic stresses on plants: Mechanistic understanding from drought and pathogen combination. J. Plant Physiol. 2015, 176, 47–54. [Google Scholar] [CrossRef]
- Aprile, A.; Havlickova, L.; Panna, R.; Marè, C.; Borrelli, G.M.; Marone, D.; Perrotta, C.; Rampino, P.; De Bellis, L.; Curn, V. Different stress responsive strategies to drought and heat in two durum wheat cultivars with contrasting water use efficiency. BMC Genom. 2013, 14, 821. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Wang, X.; Leseberg, C.H.; Jia, J.; Mao, L. Biotic and abiotic stress responses through calcium-Dependent protein kinase (CDPK) signaling in wheat (Triticum aestivum L.). Plant Signal. Behav. 2008, 3, 654–656. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Koussevitzky, S.; Mittler, R.; Miller, G. ROS and redox signalling in the response of plants to abiotic stress. Plant Cell Environ. 2012, 35, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.K.; Iandolino, A.; da Silva, F.G.; Cook, D.R. Water deficit modulates the response of Vitis vinifera to the Pierce's disease pathogen Xylella fastidiosa. Mol. Plant Microbe Interact. 2013, 26, 643–657. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, N.J.; Urwin, P.E. The interaction of plant biotic and abiotic stresses: From genes to the field. J. Exp. Bot. 2012, 63, 3523–3543. [Google Scholar] [CrossRef] [PubMed]
- Rizhsky, L.; Liang, H.; Shuman, J.; Shulaev, V.; Davletova, S.; Mittler, R. When defense pathways collide. The response of Arabidopsis to a combination of drought and heat stress. Plant Physiol. 2004, 134, 1683–1696. [Google Scholar] [CrossRef] [PubMed]
- Paull, J.; Nable, R.; Rathjen, A. Physiological and genetic control of the tolerance of wheat to high concentrations of boron and implications for plant breeding. Plant Soil 1992, 146, 251–260. [Google Scholar] [CrossRef]
- Hayes, J.E.; Reid, R.J. Boron tolerance in barley is mediated by efflux of boron from the roots. Plant Physiol. 2004, 136, 3376–3382. [Google Scholar] [CrossRef] [PubMed]
- Hussain, H.A.; Men, S.; Hussain, S.; Chen, Y.; Ali, S.; Zhang, S.; Zhang, K.; Li, Y.; Xu, Q.; Liao, C.; et al. Interactive effects of drought and heat stresses on morpho-Physiological attributes, yield, nutrient uptake and oxidative status in maize hybrids. Sci. Rep. 2019, 9, 3890. [Google Scholar] [CrossRef]
- Lamaoui, M.; Jemo, M.; Datla, R.; Bekkaoui, F. Heat and drought stresses in crops and approaches for their mitigation. Front. Chem. 2018, 6, 26. [Google Scholar] [CrossRef]
- Priya, M.; Dhanker, O.P.; Siddique, K.H.M.; HanumanthaRao, B.; Nair, R.M.; Pandey, S.; Singh, S.; Varshney, R.K.; Prasad, P.V.V.; Nayyar, H. Drought and heat stress-Related proteins: An update about their functional relevance in imparting stress tolerance in agricultural crops. Theor. Appl. Genet. 2019, 132, 1607–1638. [Google Scholar] [CrossRef] [PubMed]
- Sehgal, A.; Sita, K.; Siddique, K.H.M.; Kumar, R.; Bhogireddy, S.; Varshney, R.K.; HanumanthaRao, B.; Nair, R.M.; Prasad, P.V.V.; Nayyar, H. Drought or/and heat-Stress effects on seed filling in food crops: Impacts on functional biochemistry, seed yields, and nutritional quality. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Keren, R.; Bingham, F.T. Boron in Water, Soils, and Plants. In Advances in Soil Science; Stewart, B.A., Ed.; Springer: New York, NY, USA, 1985. [Google Scholar]
- Holloway, R.; Alston, A. The effects of salt and boron on growth of wheat. Aust. J. Agric. Res. 1992, 43, 987–1001. [Google Scholar] [CrossRef]
- Grieve, C.M.; Grattan, S.R.; Maas, E.V. Plant salt tolerance. In Agricultural Salinity Assessment and Management, 2nd ed.; ASCE: Reston, VA, USA, 2012. [Google Scholar]
- Grieve, C.M.; Poss, J.A. Wheat response to interactive effects of boron and salinity. J. Plant Nutr. 2000, 23, 1217–1226. [Google Scholar] [CrossRef]
- Nuttall, J.G.; Armstrong, R.; Connor, D. Early growth of wheat is more sensitive to salinity than boron at levels encountered in alkaline soils of south-Eastern Australia. Aust. J. Exp. Agric. 2006, 46, 1507–1514. [Google Scholar] [CrossRef]
- Rengasamy, P. World salinization with emphasis on Australia. J. Exp. Bot. 2006, 57, 1017–1023. [Google Scholar] [CrossRef] [PubMed]
- Adcock, D.; McNeill, A.; McDonald, G.; Armstrong, R. Subsoil constraints to crop production on neutral and alkaline soils in south-eastern Australia: A review of current knowledge and management strategies. Aust. J. Exp. Agric. 2007, 47, 1245–1261. [Google Scholar] [CrossRef]
- Nuttall, J.; Armstrong, R. Impact of subsoil physicochemical constraints on crops grown in the Wimmera and Mallee is reduced during dry seasonal conditions. Soil Res. 2010, 48, 125–139. [Google Scholar] [CrossRef]
- Wimmer, M.A.; Goldbach, H.E. Boron-and-Salt interactions in wheat are affected by boron supply. J. Soil Sci. Plant Nutr. 2012, 175, 171–179. [Google Scholar] [CrossRef]
- Berger, K. Boron in soils and crops. In Advances in Agronomy; Elsevier: New York, NY, USA, 1949; Volume 1, pp. 321–351. [Google Scholar]
- Marschner, H. Mineral Nutrition of Higher Plants, 2nd ed.; Academic Press: New York, NY, USA, 1995. [Google Scholar]
- Alpaslan, M.; Gunes, A. Interactive effects of boron and salinity stress on the growth, membrane permeability and mineral composition of tomato and cucumber plants. Plant Soil 2001, 236, 123–128. [Google Scholar] [CrossRef]
- Bingham, F.; Strong, J.; Rhoades, J.; Keren, R. Effects of salinity and varying boron concentrations on boron uptake and growth of wheat. Plant Soil 1987, 97, 345–351. [Google Scholar] [CrossRef]
- Ehret, D.; Ho, L. The effects of salinity on dry matter partitioning and fruit growth in tomatoes grown in nutrient film culture. J. Hortic. Sci. 1986, 61, 361–367. [Google Scholar] [CrossRef]
- Arif, M.R.; Islam, M.T.; Robin, A.H.K. Salinity stress alters root morphology and root hair traits in Brassica Napus. Plants 2019, 8, 192. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, J.A. Salinity tolerance in plants: Trends and perspectives. Int. J. Mol. Sci. 2019, 20, 2408. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.H.; Lin, X.J.; Zhang, L.Y.; Wang, X.D.; Fan, G.C.; Chen, L.S. MicroRNA sequencing revealed citrus adaptation to long-Term boron toxicity through modulation of root development by mir319 and mir171. Int. J. Mol. Sci. 2019, 20, 1422. [Google Scholar] [CrossRef] [PubMed]
- Song, B.; Hao, X.; Wang, X.; Yang, S.; Dong, Y.; Ding, Y.; Wang, Q.; Wang, X.; Zhou, J. Boron stress inhibits beet (Beta vulgaris l.) growth through influencing endogenous hormones and oxidative stress response. Soil Sci. Plant Nutr. 2019, 65, 346–352. [Google Scholar] [CrossRef]
- Ben-Gal, A.; Shani, U. Yield, transpiration and growth of tomatoes under combined excess boron and salinity stress. Plant Soil 2002, 247, 211–221. [Google Scholar] [CrossRef]
- Grattan, S.R.; Shannon, M.C.; Grieve, C.M.; Poss, J.A.; Suarez, D.; Leland, F. Interactive effects of salinity and boron on the performance and water use of eucalyptus. In International Symposium on Irrigation of Horticultural Crops; ISHS Acta Horticulturae: Crete, Greece, 1996; pp. 607–614. [Google Scholar]
- Mikkelsen, R.; Haghnia, G.; Page, A.; Bingham, F. The influence of selenium, salinity, and boron on alfalfa tissue composition and yield. J. Environ. Qual. 1988, 17, 85–88. [Google Scholar] [CrossRef]
- Edelstein, M.; Ben-Hur, M.; Cohen, R.; Burger, Y.; Ravina, I. Boron and salinity effects on grafted and non-grafted melon plants. Plant Soil 2005, 269, 273–284. [Google Scholar] [CrossRef]
- Shani, U.; Hanks, R. Model of integrated effects of boron, inert salt, and water flow on crop yield. Agronomy 1993, 85, 713–717. [Google Scholar] [CrossRef]
- Wimmer, M.A.; Bassil, E.S.; Brown, P.H.; Läuchli, A. Boron response in wheat is genotype-Dependent and related to boron uptake, translocation, allocation, plant phenological development and growth rate. Funct. Plant Biol. 2005, 32, 507–515. [Google Scholar] [CrossRef]
- Eraslan, F.; Inal, A.; Gunes, A.; Alpaslan, M. Impact of exogenous salicylic acid on the growth, antioxidant activity and physiology of carrot plants subjected to combined salinity and boron toxicity. Sci. Hort. 2007, 113, 120–128. [Google Scholar] [CrossRef]
- Ismail, A. Response of maize and sorghum to excess boron and salinity. Biol. Plant. 2003, 47, 313–316. [Google Scholar] [CrossRef]
- El-Motaium, R.; Hu, H.; Brown, P.H. The relative tolerance of six Prunus rootstocks to boron and salinity. J. Am. Soc. Hortic. Sci. 1994, 119, 1169–1175. [Google Scholar] [CrossRef]
- Poss, J.A.; Grattan, S.R.; Grieve, C.M.; Shannon, M.C. Characterization of leaf boron injury in salt-Stressed Eucalyptus by image analysis. Plant Soil 1998, 206, 237–245. [Google Scholar] [CrossRef]
- Apostol, K.G.; Zwiazek, J.J.; MacKinnon, M.D. NaCl and Na2SO4 alter responses of jack pine (Pinus banksiana) seedlings to boron. Plant Soil 2002, 240, 321–329. [Google Scholar] [CrossRef]
- Grattan, S.; Grieve, C.; Poss, J.; Suarez, D.; Läuchli, A. Continued Investigation into the Interactions of Saline Drainage Water on Crop Tolerance to Boron. 2004–2005 Technical Progress Report: UC Salinity/Drainage Research Program; DANR, University California: San Diego, CA, USA, 2005. [Google Scholar]
- Rogers, M.; Grieve, C.; Shannon, M. The response of lucerne (Medicago sativa L.) to sodium sulphate and chloride salinity. Plant Soil 1998, 202, 271–280. [Google Scholar] [CrossRef]
- Grattan, S.; Grieve, C.; Poss, J.; Suarez, D.; Läuchli, A.; Smith, T. Continued Investigation into the Interactions of Saline Drainage Water on Crop Tolerance to Boron. 2005–2006 Technical Progress Report: UC Salinity/Drainage Research Program; DANR, University California: San Diego, CA, USA, 2006. [Google Scholar]
- Yermiyahu, U.; Ben-Gal, A.; Sarig, P.; Zipilevitch, E. Boron toxicity in grapevine (Vitis vinifera L.) in conjunction with salinity and rootstock effects. J. Hortic. Sci. Biotechnol. 2007, 82, 547–554. [Google Scholar] [CrossRef]
- Mittler, R. Abiotic stress, the field environment and stress combination. Trends Plant Sci. 2006, 11, 15–19. [Google Scholar] [CrossRef]
- Cakmak, I.; Kurz, H.; Marschner, H. Short-Term effects of boron, germanium and high light intensity on membrane permeability in boron deficient leaves of sunflower. Physiol. Plant. 1995, 95, 11–18. [Google Scholar] [CrossRef]
- Brown, P.; Bellaloui, N.; Wimmer, M.; Bassil, E.; Ruiz, J.; Hu, H.; Pfeffer, H.; Dannel, F.; Römheld, V. Boron in plant biology. Plant Biol. 2002, 4, 205–223. [Google Scholar] [CrossRef]
- Bonilla, I.; El-Hamdaoui, A.; Bolaños, L. Boron and calcium increase Pisum sativum seed germination and seedling development under salt stress. Plant Soil 2004, 267, 97–107. [Google Scholar] [CrossRef]
- Dannel, F.; Pfeffer, H.; Römheld, V. Distribution within the plant or compartmentation does not contribute substantially to the detoxification of excess boron in sunflower (Helianthus annus). Funct. Plant Biol. 1999, 26, 95–99. [Google Scholar] [CrossRef]
- Pfeffer, H.; Dannel, F.; Römheld, V. Are there connections between phenol metabolism, ascorbate metabolism and membrane integrity in leaves of boron-Deficient sunflower plants? Physiol. Plant. 1998, 104, 479–485. [Google Scholar] [CrossRef]
- Reid, R.J.; Hayes, J.E.; Post, A.; Stangoulis, J.C.R.; Graham, R.D. A critical analysis of the causes of boron toxicity in plants. Plant Cell Environ. 2004, 27, 1405–1414. [Google Scholar] [CrossRef]
- Pilar, C.; Antonio, O.; Antonio, C. Effects of saline stress and calcium on lipid composition in bean roots. Phytochemistry 1993, 32, 1131–1136. [Google Scholar] [CrossRef]
- Shalata, A.; Tal, M. The effect of salt stress on lipid peroxidation and antioxidants in the leaf of the cultivated tomato and its wild salt-Tolerant relative Lycopersicon pennellii. Physiol. Plant. 1998, 104, 169–174. [Google Scholar] [CrossRef]
- Wimmer, M.A.; Muehling, K.H.; Läuchli, A.; Brown, P.H.; Goldbach, H.E. Interaction of salinity and boron toxicity in wheat (Triticum aestivum L.). In Plant Nutrition; Springer: Dordrecht, The Netherlands, 2001; pp. 426–427. [Google Scholar]
- Reid, R.; Fitzpatrick, K. Influence of leaf tolerance mechanisms and rain on boron toxicity in barley and wheat. Plant Physiol. 2009, 151, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Munns, R. Genes and salt tolerance: Bringing them together. New Phytol. 2005, 167, 645–663. [Google Scholar] [CrossRef] [PubMed]
- Apse, M.P.; Aharon, G.S.; Snedden, W.A.; Blumwald, E. Salt tolerance conferred by overexpression of a vacuolar Na+/H+ antiport in Arabidopsis. Science 1999, 285, 1256–1258. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, A.; Chiba, K.; Maeda, M.; Nakamura, A.; Maeshima, M.; Tanaka, Y. Effect of salt and osmotic stresses on the expression of genes for the vacuolar H+-pyrophosphatase, H+-ATPase subunit A, and Na+/H+ antiporter from barley. J. Exp. Bot. 2004, 55, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Ferrol, N.; Belver, A.; Roldan, M.; Rodriguez-Rosales, M.P.; Donaire, J.P. Effects of Boron on Proton Transport and Membrane Properties of Sunflower (Helianthus annuus L.) Cell Microsomes. Plant Physiol. 1993, 103, 763–769. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Salus, S.; Schumaker, K. Salt salinity and the activities of the H+-ATPases in cotton seedlings. Crop Sci. 1997, 37, 190–197. [Google Scholar] [CrossRef]
- Fukuda, A.; Yazaki, Y.; Ishikawa, T.; Koike, S.; Tanaka, Y. Na+/H+ antiporter in tonoplast vesicles from rice roots. Plant Cell Physiol. 1998, 39, 196–201. [Google Scholar] [CrossRef]
- Chaumont, F.; Tyerman, S.D. Aquaporins: Highly regulated channels controlling plant water relations. Plant Physiol. 2014, 164, 1600–1618. [Google Scholar] [CrossRef] [PubMed]
- Maurel, C.; Boursiac, Y.; Luu, D.T.; Santoni, V.; Shahzad, Z.; Verdoucq, L. Aquaporins in plants. Physiol. Rev. 2015, 95, 1321–1358. [Google Scholar] [CrossRef] [PubMed]
- Moshelion, M.; Halperin, O.; Wallach, R.; Oren, R.; Way, D.A. Role of aquaporins in determining transpiration and photosynthesis in water-stressed plants: Crop water-use efficiency, growth and yield. Plant Cell Environ. 2015, 38, 1785–1793. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Plant salt tolerance. Trends Plant Sci. 2001, 6, 66–71. [Google Scholar] [CrossRef]
- Karabal, E.; Yücel, M.; Öktem, H.A. Antioxidant responses of tolerant and sensitive barley cultivars to boron toxicity. Plant Sci. 2003, 164, 925–933. [Google Scholar] [CrossRef]
- Eraslan, F.; Inal, A.; Savasturk, O.; Gunes, A. Changes in antioxidative system and membrane damage of lettuce in response to salinity and boron toxicity. Sci. Hortic. 2007, 114, 5–10. [Google Scholar] [CrossRef]
- del Carmen Martinez-Ballesta, M.; Bastias, E.; Carvajal, M. Combined effect of boron and salinity on water transport: The role of aquaporins. Plant Signal. Behav. 2008, 3, 844–845. [Google Scholar] [CrossRef] [PubMed]
- Macho-Rivero, M.A.; Herrera-Rodriguez, M.B.; Brejcha, R.; Schaffner, A.R.; Tanaka, N.; Fujiwara, T.; Gonzalez-Fontes, A.; Camacho-Cristobal, J.J. Boron Toxicity Reduces Water Transport from Root to Shoot in Arabidopsis Plants. Evidence for a Reduced Transpiration Rate and Expression of Major PIP Aquaporin Genes. Plant Cell Physiol. 2018, 59, 836–844. [Google Scholar] [CrossRef] [PubMed]
- Cramer, G.R. Sodium-calcium interactions under salinity stress. In Salinity: Environment-Plants-Molecules; Springer: Dordrecht, The Netherlands, 2002; pp. 205–227. [Google Scholar]
- Lauchli, A.; Schubert, S. The role of calcium in the regulation of membrane and cellular growth processes under salt stress. In Environmental Stress in Plants; Springer: Berlin, Heidelberg, 1989; pp. 131–138. [Google Scholar]
- Blumwald, E.; Aharon, G.S.; Apse, M.P. Sodium transport in plant cells. Biochim. Biophys. Acta Biomembr. 2000, 1465, 140–151. [Google Scholar] [CrossRef]
- Mohamed, A.K.; Qayyum, M.F.; Shahzad, A.N.; Gul, M.; Wakeel, A. Interactive Effect of Boron and Salinity on Growth, Physiological and Biochemical Attributes of Wheat (Triticum aestivum). Int. J. Agric. Biol. 2016, 18, 238–244. [Google Scholar] [CrossRef]
- Läuchli, A.; Grattan, S.R. Plant Growth and Development Under Salinity Stress. In Advances in Molecular Breeding Toward Drought and Salt Tolerant Crops; Springer: Dordrecht, The Netherlands, 2007; pp. 1–32. [Google Scholar]
- Ganmore-Neumann, R.; Davidov, S. Uptake and distribution of calcium in rose plantlets as affected by calcium and boron concentration in culture solution. Plant Soil 1993, 155, 151–154. [Google Scholar] [CrossRef]
- Carpena, R.O.; Esteban, E.; Sarro, M.J.; Peñalosa, J.; Gárate, A.N.; Lucena, J.J.; Zornoza, P. Boron and calcium distribution in nitrogen-Fixing pea plants. Plant Sci. 2000, 151, 163–170. [Google Scholar] [CrossRef]
- Supanjani, L.K.; Lee, K. Hot pepper response to interactive effects of salinity and boron. Plant Soil Environ. 2006, 52, 227–233. [Google Scholar] [CrossRef]
- Al-Hakimi, A.; Hamada, A. Counteraction of salinity stress on wheat plants by grain soaking in ascorbic acid, thiamin or sodium salicylate. Biol. Plant. 2001, 44, 253–261. [Google Scholar] [CrossRef]
- Munns, R.; James, R.A.; Lauchli, A. Approaches to increasing the salt tolerance of wheat and other cereals. J. Exp. Bot. 2006, 57, 1025–1043. [Google Scholar] [CrossRef]
- Wu, H.; Shabala, L.; Barry, K.; Zhou, M.; Shabala, S. Ability of leaf mesophyll to retain potassium correlates with salinity tolerance in wheat and barley. Physiol. Plant. 2013, 149, 515–527. [Google Scholar] [CrossRef]
- Møller, I.S.; Tester, M. Salinity tolerance of Arabidopsis: A good model for cereals? Trends Plant Sci. 2007, 12, 534–540. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, W.H. Potassium transport and signaling in higher plants. Annu. Rev. Plant Biol. 2013, 64, 451–476. [Google Scholar] [CrossRef] [PubMed]
- Macho-Rivero, M.A.; Camacho-Cristobal, J.J.; Herrera-Rodriguez, M.B.; Muller, M.; Munne-Bosch, S.; Gonzalez-Fontes, A. Abscisic acid and transpiration rate are involved in the response to boron toxicity in Arabidopsis plants. Physiol. Plant. 2017, 160, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Miwa, K.; Fujiwara, T. Boron transport in plants: Co-ordinated regulation of transporters. Ann. Bot. 2010, 105, 1103–1108. [Google Scholar] [CrossRef] [PubMed]
- Takano, J.; Miwa, K.; Fujiwara, T. Boron transport mechanisms: Collaboration of channels and transporters. Trends Plant Sci. 2008, 13, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Sutton, T.; Baumann, U.; Hayes, J.; Collins, N.C.; Shi, B.J.; Schnurbusch, T.; Hay, A.; Mayo, G.; Pallotta, M.; Tester, M.; et al. Boron-Toxicity tolerance in barley arising from efflux transporter amplification. Science 2007, 318, 1446–1449. [Google Scholar] [CrossRef]
- Brown, P.H.; Hu, H. Phloem mobility of boron is species dependent: Evidence for phloem mobility in sorbitol-Rich species. Ann. Bot. 1996, 77, 497–506. [Google Scholar] [CrossRef]
- Brown, P.H.; Shelp, B.J. Boron mobility in plants. Plant Soil 1997, 193, 85–101. [Google Scholar] [CrossRef]
- Stangoulis, J.; Tate, M.; Graham, R.; Bucknall, M.; Palmer, L.; Boughton, B.; Reid, R. The mechanism of boron mobility in wheat and canola phloem. Plant Physiol. 2010, 153, 876–881. [Google Scholar] [CrossRef]
- Masood, S.; Saleh, L.; Witzel, K.; Plieth, C.; Mühling, K.H. Determination of oxidative stress in wheat leaves as influenced by boron toxicity and NaCl stress. Plant Physiol. Biochol. 2012, 56, 56–61. [Google Scholar] [CrossRef]
- Gunes, A.; Soylemezoglu, G.; Inal, A.; Bagci, E.; Coban, S.; Sahin, O. Antioxidant and stomatal responses of grapevine (Vitis vinifera L.) to boron toxicity. Sci. Hortic. 2006, 110, 279–284. [Google Scholar] [CrossRef]
- Rozema, J.; De Bruin, J.; Broekman, R. Effect of boron on the growth and mineral economy of some halophytes and non-Halophytes. New Phytol. 1992, 121, 249–256. [Google Scholar] [CrossRef]
- Hamurcu, M.; Hakki, E.E.; Sert, T.D.; Özdemir, C.; Minareci, E.; Avsaroglu, Z.Z.; Gezgin, S.; Kayis, S.A.; Bell, R.W. Extremely high boron tolerance in Puccinellia distans (Jacq.) Parl. related to root boron exclusion and a well-regulated antioxidant system. Z. Naturforsch. C 2016, 71, 273–285. [Google Scholar] [CrossRef]
- Cervilla, L.M.; Blasco, B.; Rios, J.J.; Romero, L.; Ruiz, J.M. Oxidative stress and antioxidants in tomato (Solanum lycopersicum) plants subjected to boron toxicity. Ann. Bot. 2007, 100, 747–756. [Google Scholar] [CrossRef]
- Jain, M.; Mathur, G.; Koul, S.; Sarin, N. Ameliorative effects of proline on salt stress-Induced lipid peroxidation in cell lines of groundnut (Arachis hypogaea L.). Plant Cell Rep. 2001, 20, 463–468. [Google Scholar] [CrossRef]
- Panda, S.; Upadhyay, R. Salt stress injury induces oxidative alterations and antioxidative defence in the roots of Lemna minor. Biol. Plant. 2004, 48, 249–253. [Google Scholar] [CrossRef]
- Reddy, A.R.; Chaitanya, K.V.; Vivekanandan, M. Drought-Induced responses of photosynthesis and antioxidant metabolism in higher plants. J. Plant Physiol. 2004, 161, 1189–1202. [Google Scholar] [CrossRef]
- Sairam, R.K.; Srivastava, G.C.; Agarwal, S.; Meena, R.C. Differences in antioxidant activity in response to salinity stress in tolerant and susceptible wheat genotypes. Biol. Plant. 2005, 49, 85. [Google Scholar] [CrossRef]
- Sairam, R.K.; Rao, K.V.; Srivastava, G.C. Differential response of wheat genotypes to long term salinity stress in relation to oxidative stress, antioxidant activity and osmolyte concentration. Plant Sci. 2002, 163, 1037–1046. [Google Scholar] [CrossRef]
- Cervilla, L.M.; Blasco, B.; Rios, J.J.; Rosales, M.A.; Sánchez-Rodríguez, E.; Rubio-Wilhelmi, M.M.; Romero, L.; Ruiz, J.M. Parameters Symptomatic for Boron Toxicity in Leaves of Tomato Plants. J. Bot. 2012, 2012, 726206. [Google Scholar] [CrossRef]
- Molassiotis, A.; Sotiropoulos, T.; Tanou, G.; Diamantidis, G.; Therios, I. Boron-Induced oxidative damage and antioxidant and nucleolytic responses in shoot tips culture of the apple rootstock EM 9 (Malus domestica Borkh). Environ. Exp. Bot. 2006, 56, 54–62. [Google Scholar] [CrossRef]
- Samet, H.; Çıkılı, Y. Response of purslane (Portulaca oleracea L.) to excess boron and salinity: Physiological approach. Russ. J. Plant Physiol. 2019, 66, 316–325. [Google Scholar] [CrossRef]
- Kaya, C.; Akram, N.A.; Ashraf, M. Kinetin and indole acetic acid promote antioxidant defense system and reduce oxidative stress in maize (Zea mays L.) plants grown at boron toxicity. J. Plant Growth Regul. 2018, 37, 1258–1266. [Google Scholar] [CrossRef]
- Goldberg, S.; Lesch, S.M.; Suarez, D.L. Predicting boron adsorption by soils using soil chemical parameters in the constant capacitance model. Soil Sci. Soc. Am. J. 2000, 64, 1356–1363. [Google Scholar] [CrossRef]
- Läuchli, A.; Epstein, E. Plant responses to saline and sodic conditions. Agric. Salin. Assess. Manag. 1990, 71, 113–137. [Google Scholar]
- Francois, L. Effect of excess boron on broccoli, cauliflower, and radish. J Am. Soc. Hortic. Sci. 1986, 111, 494–498. [Google Scholar]
- Läuchli, A.; Grattan, S.R. Soil pH extremes. In Plant Stress Physiology; Shabala, S., Ed.; CAB International: Wallingford, UK, 2012. [Google Scholar]
- Sternberg, P.; Ulery, A.; Villa-C, M. Salinity and boron effects on growth and yield of tepary and kidney beans. HortScience 2001, 36, 1269–1272. [Google Scholar] [CrossRef]
- Yadav, H.; Yadav, O.; Dhankar, O.; Oswal, M. Effect of chloride salinity and boron on germination, growth and mineral-Composition of chickpea (Cicer-Arietinum L). Ann Arid Zone 1989, 28, 63–67. [Google Scholar]
- Goldberg, S.; Grieve, C.M. Boron adsorption by maize cell walls. Plant Soil 2003, 251, 137–142. [Google Scholar] [CrossRef]
- Assaha, D.V.M.; Ueda, A.; Saneoka, H.; Al-Yahyai, R.; Yaish, M.W. The Role of Na+ and K+ Transporters in Salt Stress Adaptation in Glycophytes. Front. Physiol. 2017, 8, 509. [Google Scholar] [CrossRef]
- Volkov, V. Salinity tolerance in plants. Quantitative approach to ion transport starting from halophytes and stepping to genetic and protein engineering for manipulating ion fluxes. Front. Plant Sci. 2015, 6, 873. [Google Scholar] [CrossRef]
- Atmodjo, M.A.; Hao, Z.; Mohnen, D. Evolving views of pectin biosynthesis. Annu. Rev. Plant Biol. 2013, 64, 747–779. [Google Scholar] [CrossRef]
- O'Neill, M.A.; Ishii, T.; Albersheim, P.; Darvill, A.G. Rhamnogalacturonan II: Structure and function of a borate cross-Linked cell wall pectic polysaccharide. Annu. Rev. Plant Biol. 2004, 55, 109–139. [Google Scholar] [CrossRef]
- Voxeur, A.; Fry, S.C. Glycosylinositol phosphorylceramides from Rosa cell cultures are boron-Bridged in the plasma membrane and form complexes with rhamnogalacturonan II. Plant J. 2014, 79, 139–149. [Google Scholar] [CrossRef]
- Funakawa, H.; Miwa, K. Synthesis of borate cross-Linked rhamnogalacturonan II. Front. Plant Sci. 2015, 6, 223. [Google Scholar] [CrossRef]
- Stangoulis, J.C.; Reid, R.J.; Brown, P.H.; Graham, R.D. Kinetic analysis of boron transport in Chara. Planta 2001, 213, 142–146. [Google Scholar] [CrossRef]
- Schnurbusch, T.; Hayes, J.; Sutton, T. Boron toxicity tolerance in wheat and barley: Australian perspectives. Breed. Sci. 2010, 60, 297–304. [Google Scholar] [CrossRef]
- Yoshinari, A.; Takano, J. Insights into the Mechanisms Underlying Boron Homeostasis in Plants. Front. Plant Sci. 2017, 8, 1951. [Google Scholar] [CrossRef]
- Kapilan, R.; Vaziri, M.; Zwiazek, J.J. Regulation of aquaporins in plants under stress. Biol. Res. 2018, 51, 4. [Google Scholar] [CrossRef]
- Takano, J.; Wada, M.; Ludewig, U.; Schaaf, G.; von Wiren, N.; Fujiwara, T. The Arabidopsis major intrinsic protein NIP5;1 is essential for efficient boron uptake and plant development under boron limitation. Plant Cell 2006, 18, 1498–1509. [Google Scholar] [CrossRef]
- Bienert, M.D.; Bienert, G.P. Plant aquaporins and metalloids. In Plant Aquaporins; Springer: Boston, MA, USA, 2017; pp. 297–332. [Google Scholar]
- Roberts, D.M.; Routray, P. The nodulin 26 intrinsic protein subfamily. In Plant Aquaporins; Springer: Boston, MA, USA, 2017; pp. 267–296. [Google Scholar]
- Takano, J.; Tanaka, M.; Toyoda, A.; Miwa, K.; Kasai, K.; Fuji, K.; Onouchi, H.; Naito, S.; Fujiwara, T. Polar localization and degradation of Arabidopsis boron transporters through distinct trafficking pathways. Proc. Natl. Acad. Sci. USA 2010, 107, 5220–5225. [Google Scholar] [CrossRef]
- Tanaka, M.; Takano, J.; Chiba, Y.; Lombardo, F.; Ogasawara, Y.; Onouchi, H.; Naito, S.; Fujiwara, T. Boron-dependent degradation of NIP5;1 mRNA for acclimation to excess boron conditions in Arabidopsis. Plant Cell 2011, 23, 3547–3559. [Google Scholar] [CrossRef]
- Tanaka, M.; Wallace, I.S.; Takano, J.; Roberts, D.M.; Fujiwara, T. NIP6; 1 is a boric acid channel for preferential transport of boron to growing shoot tissues in Arabidopsis. Plant Cell 2008, 20, 2860–2875. [Google Scholar] [CrossRef]
- Li, T.; Choi, W.G.; Wallace, I.S.; Baudry, J.; Roberts, D.M. Arabidopsis thaliana NIP7; 1: An anther-Specific boric acid transporter of the aquaporin superfamily regulated by an unusual tyrosine in helix 2 of the transport pore. Biochemistry 2011, 50, 6633–6641. [Google Scholar] [CrossRef]
- Durbak, A.R.; Phillips, K.A.; Pike, S.; O’Neill, M.A.; Mares, J.; Gallavotti, A.; Malcomber, S.T.; Gassmann, W.; McSteen, P. Transport of boron by the tassel-Less1 aquaporin is critical for vegetative and reproductive development in maize. Plant Cell 2014, 26, 2978–2995. [Google Scholar] [CrossRef]
- Hanaoka, H.; Uraguchi, S.; Takano, J.; Tanaka, M.; Fujiwara, T. O s NIP 3; 1, a rice boric acid channel, regulates boron distribution and is essential for growth under boron-Deficient conditions. Plant J. 2014, 78, 890–902. [Google Scholar] [CrossRef]
- Lv, Q.; Wang, L.; Wang, J.Z.; Li, P.; Chen, Y.L.; Du, J.; He, Y.K.; Bao, F. Shb1/hy1 alleviates excess boron stress by increasing BOR4 expression level and maintaining boron homeostasis in Arabidopsis roots. Front. Plant Sci. 2017, 8, 790. [Google Scholar] [CrossRef]
- Miwa, K.; Takano, J.; Omori, H.; Seki, M.; Shinozaki, K.; Fujiwara, T. Plants tolerant of high boron levels. Science 2007, 318, 1417. [Google Scholar] [CrossRef]
- Miwa, K.; Aibara, I.; Fujiwara, T. Arabidopsis thaliana bor4 is upregulated under high boron conditions and confers tolerance to high boron. Soil Sci. Plant Nutr. 2014, 60, 349–355. [Google Scholar] [CrossRef]
- Aibara, I.; Hirai, T.; Kasai, K.; Takano, J.; Onouchi, H.; Naito, S.; Fujiwara, T.; Miwa, K. Boron-Dependent translational suppression of the borate exporter BOR1 contributes to the avoidance of boron toxicity. Plant Physiol. 2018, 177, 759–774. [Google Scholar] [CrossRef]
- Takano, J.; Noguchi, K.; Yasumori, M.; Kobayashi, M.; Gajdos, Z.; Miwa, K.; Hayashi, H.; Yoneyama, T.; Fujiwara, T. Arabidopsis boron transporter for xylem loading. Nature 2002, 420, 337. [Google Scholar] [CrossRef]
- Takano, J.; Toyoda, A.; Miwa, K.; Fujiwara, T. Polar trafficking and boron-Dependent endocytosis of borate transporters. In Plant and Cell Physiology; Oxford University Press: Oxford, UK, 2007; p. 228. [Google Scholar]
- Reid, R. Identification of Boron Transporter Genes Likely to be Responsible for Tolerance to Boron Toxicity in Wheat and Barley. Plant Cell Physiol. 2007, 48, 1673–1678. [Google Scholar] [CrossRef]
- Leaungthitikanchana, S.; Fujibe, T.; Tanaka, M.; Wang, S.; Sotta, N.; Takano, J.; Fujiwara, T. Differential expression of three BOR1 genes corresponding to different genomes in response to boron conditions in hexaploid wheat (Triticum aestivum L.). Plant Cell Physiol. 2013, 54, 1056–1063. [Google Scholar] [CrossRef]
- Miwa, K.; Fujiwara, T. Role of overexpressed BOR4, a boron exporter, in tolerance to high level of boron in shoots. Soil Sci. Plant Nutr. 2011, 57, 558–565. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Hanaoka, H.; Kobayashi, M.; Miyoshi, K.; Miwa, K.; Fujiwara, T. Cell-Type specificity of the expression of OsBOR1, a rice efflux boron transporter gene, is regulated in response to boron availability for efficient boron uptake and xylem loading. Plant Cell 2007, 19, 2624–2635. [Google Scholar] [CrossRef]
- Rajendran, K.; Tester, M.; Roy, S.J. Quantifying the three main components of salinity tolerance in cereals. Plant Cell Environ. 2009, 32, 237–249. [Google Scholar] [CrossRef]
- Roy, S.J.; Negrão, S.; Tester, M. Salt resistant crop plants. Curr. Opin. Biotechnol. 2014, 26, 115–124. [Google Scholar] [CrossRef]
- Isayenkov, S.V.; Maathuis, F.J.M. Plant salinity stress: Many unanswered questions remain. Front. Plant Sci. 2019, 10, 80. [Google Scholar] [CrossRef]
- Apse, M.P.; Blumwald, E. Na+ transport in plants. FEBS Lett. 2007, 581, 2247–2254. [Google Scholar] [CrossRef]
- Hauser, F.; Horie, T. A conserved primary salt tolerance mechanism mediated by HKT transporters: A mechanism for sodium exclusion and maintenance of high K+/Na+ ratio in leaves during salinity stress. Plant Cell Environ. 2010, 33, 552–565. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Blumwald, E. Developing salt-Tolerant crop plants: Challenges and opportunities. Trends Plant Sci. 2005, 10, 615–620. [Google Scholar] [CrossRef]
- Agarwal, P.K.; Shukla, P.S.; Gupta, K.; Jha, B. Bioengineering for salinity tolerance in plants: State of the art. Mol. Biotechnol. 2013, 54, 102–123. [Google Scholar] [CrossRef]
- Wang, Q.; Guan, C.; Wang, P.; Lv, M.L.; Ma, Q.; Wu, G.Q.; Bao, A.K.; Zhang, J.L.; Wang, S.M. AtHKT1; 1 and AtHAK5 mediate low-Affinity Na+ uptake in Arabidopsis thaliana under mild salt stress. Plant Growth Regul. 2015, 75, 615–623. [Google Scholar] [CrossRef]
- Takahashi, R.; Nishio, T.; Ichizen, N.; Takano, T. High-Affinity K+ transporter PhaHAK5 is expressed only in salt-sensitive reed plants and shows Na+ permeability under NaCl stress. Plant Cell Rep. 2007, 26, 1673–1679. [Google Scholar] [CrossRef]
- Roy, S.; Chakraborty, U. Role of sodium ion transporters and osmotic adjustments in stress alleviation of Cynodon dactylon under NaCl treatment: A parallel investigation with rice. Protoplasma 2018, 255, 175–191. [Google Scholar] [CrossRef]
- De Boer, A.; Volkov, V. Logistics of water and salt transport through the plant: Structure and functioning of the xylem. Plant Cell Environ. 2003, 26, 87–101. [Google Scholar] [CrossRef]
- Keisham, M.; Mukherjee, S.; Bhatla, S.C. Mechanisms of Sodium Transport in Plants-Progresses and Challenges. Int. J. Mol. Sci. 2018, 19, 647. [Google Scholar] [CrossRef]
- Liu, Q.; Liu, R.; Ma, Y.; Song, J. Physiological and molecular evidence for Na+ and Cl− exclusion in the roots of two Suaeda salsa populations. Aquat. Bot. 2018, 146, 1–7. [Google Scholar] [CrossRef]
- Davenport, R.; Munoz-Mayor, A.; Jha, D.; Essah, P.; Rus, A.; Tester, M. The Na+ transporter AtHKT1 controls xylem retrieval of Na+ from the xylem in Arabidopsis. Plant Cell Environ. 2007, 30, 497–507. [Google Scholar] [CrossRef]
- Rus, A.; Yokoi, S.; Sharkhuu, A.; Reddy, M.; Lee, B.H.; Matsumoto, T.K.; Koiwa, H.; Zhu, J.K.; Bressan, R.A.; Hasegawa, P.M. AtHKT1 is a salt tolerance determinant that controls Na+ entry into plant roots. Proc. Natl. Acad. Sci. USA 2001, 98, 14150–14155. [Google Scholar] [CrossRef]
- Shi, H.; Ishitani, M.; Kim, C.; Zhu, J.K. The Arabidopsis thaliana salt tolerance gene SOS1 encodes a putative Na+/H+ antiporter. Proc. Natl. Acad. Sci. USA 2000, 97, 6896–6901. [Google Scholar] [CrossRef]
- Shi, H.; Quintero, F.J.; Pardo, J.M.; Zhu, J.K. The putative plasma membrane Na(+)/H(+) antiporter SOS1 controls long-distance Na(+) transport in plants. Plant Cell 2002, 14, 465–477. [Google Scholar] [CrossRef]
- Ma, Q.; Li, Y.X.; Yuan, H.J.; Hu, J.; Wei, L.; Bao, A.K.; Zhang, J.L.; Wang, S.M. ZxSOS1 is essential for long-distance transport and spatial distribution of Na+ and K+ in the xerophyte Zygophyllum xanthoxylum. Plant Soil 2014, 374, 661–676. [Google Scholar] [CrossRef]
- Guo, Q.; Wang, P.; Ma, Q.; Zhang, J.L.; Bao, A.K.; Wang, S.M. Selective transport capacity for K+ over Na+ is linked to the expression levels of PtSOS1 in halophyte Puccinellia tenuiflora. Funct. Plant Biol. 2012, 39, 1047–1057. [Google Scholar] [CrossRef]
- Yuan, H.J.; Ma, Q.; Wu, G.Q.; Wang, P.; Hu, J.; Wang, S.M. ZxNHX controls Na+ and K+ homeostasis at the whole-Plant level in Zygophyllum xanthoxylum through feedback regulation of the expression of genes involved in their transport. Ann. Bot. 2014, 115, 495–507. [Google Scholar] [CrossRef]
- Qiu, Q.S.; Guo, Y.; Dietrich, M.A.; Schumaker, K.S.; Zhu, J.K. Regulation of SOS1, a plasma membrane Na+/H+ exchanger in Arabidopsis thaliana, by SOS2 and SOS3. Proc. Natl. Acad. Sci. USA. 2002, 99, 8436–8441. [Google Scholar] [CrossRef]
- Song, J.; Wang, B. Using euhalophytes to understand salt tolerance and to develop saline agriculture: Suaeda salsa as a promising model. Ann. Bot. 2015, 115, 541–553. [Google Scholar] [CrossRef]
- Lacan, D.; Durand, M. Na+-K+ exchange at the xylem/symplast boundary (its significance in the salt sensitivity of soybean). Plant Physiol. 1996, 110, 705–711. [Google Scholar] [CrossRef]
- Shi, H.; Lee, B.H.; Wu, S.J.; Zhu, J.K. Overexpression of a plasma membrane Na+/H+ antiporter gene improves salt tolerance in Arabidopsis thaliana. Nat. Biotechnol. 2003, 21, 81–85. [Google Scholar] [CrossRef]
- Wu, Y.; Ding, N.; Zhao, X.; Zhao, M.; Chang, Z.; Liu, J.; Zhang, L. Molecular characterization of PeSOS1: The putative Na+/H+ antiporter of Populus euphratica. Plant Mol. Biol. 2007, 65, 1. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Jiang, X.; Zhan, K.; Cheng, X.; Chen, X.; Pardo, J.M.; Cui, D. Functional characterization of a wheat plasma membrane Na+/H+ antiporter in yeast. Arch. Biochem. Biophys. 2008, 473, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Atienza, J.; Jiang, X.; Garciadeblas, B.; Mendoza, I.; Zhu, J.K.; Pardo, J.M.; Quintero, F.J. Conservation of the salt overly sensitive pathway in rice. Plant Physiol. 2007, 143, 1001–1012. [Google Scholar] [CrossRef] [PubMed]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genom. 2014, 2014, 701596. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ishitani, M.; Halfter, U.; Kim, C.S.; Zhu, J.K. The Arabidopsis thaliana SOS2 gene encodes a protein kinase that is required for salt tolerance. Proc. Natl. Acad. Sci. USA 2000, 97, 3730–3734. [Google Scholar] [CrossRef] [PubMed]
- Ishitani, M.; Liu, J.; Halfter, U.; Kim, C.S.; Shi, W.; Zhu, J.K. SOS3 function in plant salt tolerance requires n-myristoylation and calcium binding. Plant Cell 2000, 12, 1667–1678. [Google Scholar] [CrossRef]
- Guo, Y.; Qiu, Q.S.; Quintero, F.J.; Pardo, J.M.; Ohta, M.; Zhang, C.; Schumaker, K.S.; Zhu, J.K. Transgenic evaluation of activated mutant alleles of SOS2 reveals a critical requirement for its kinase activity and c-terminal regulatory domain for salt tolerance in Arabidopsis thaliana. Plant Cell 2004, 16, 435–449. [Google Scholar] [CrossRef]
- Guo, Y.; Halfter, U.; Ishitani, M.; Zhu, J.K. Molecular characterization of functional domains in the protein kinase SOS2 that is required for plant salt tolerance. Plant Cell 2001, 13, 1383–1400. [Google Scholar] [CrossRef]
- Kim, W.Y.; Ali, Z.; Park, H.J.; Park, S.J.; Cha, J.Y.; Perez-Hormaeche, J.; Quintero, F.J.; Shin, G.; Kim, M.R.; Qiang, Z.; et al. Release of SOS2 kinase from sequestration with Gigantea determines salt tolerance in Arabidopsis. Nat. Commun. 2013, 4, 1352. [Google Scholar] [CrossRef]
- Quintero, F.J.; Ohta, M.; Shi, H.; Zhu, J.K.; Pardo, J.M. Reconstitution in yeast of the Arabidopsis SOS signaling pathway for Na+ homeostasis. Proc. Natl. Acad. Sci. USA 2002, 99, 9061–9066. [Google Scholar] [CrossRef]
- Horie, T.; Hauser, F.; Schroeder, J.I. HKT transporter-Mediated salinity resistance mechanisms in Arabidopsis and monocot crop plants. Trends Plant Sci. 2009, 14, 660–668. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.H.; Gao, J.P.; Li, L.G.; Cai, X.L.; Huang, W.; Chao, D.Y.; Zhu, M.Z.; Wang, Z.Y.; Luan, S.; Lin, H.X. A rice quantitative trait locus for salt tolerance encodes a sodium transporter. Nat. Genet. 2005, 37, 1141–1146. [Google Scholar] [CrossRef] [PubMed]
- Schachtman, D.P.; Schroeder, J.I. Structure and transport mechanism of a high-Affinity potassium uptake transporter from higher plants. Nature 1994, 370, 655–658. [Google Scholar] [CrossRef] [PubMed]
- Horie, T.; Motoda, J.; Kubo, M.; Yang, H.; Yoda, K.; Horie, R.; Chan, W.Y.; Leung, H.Y.; Hattori, K.; Konomi, M. Enhanced salt tolerance mediated by AtHKT1 transporter-Induced Na+ unloading from xylem vessels to xylem parenchyma cells. Plant J. 2005, 44, 928–938. [Google Scholar]
- Munns, R.; James, R.A.; Xu, B.; Athman, A.; Conn, S.J.; Jordans, C.; Byrt, C.S.; Hare, R.A.; Tyerman, S.D.; Tester, M. Wheat grain yield on saline soils is improved by an ancestral Na+ transporter gene. Nat. Biotechol. 2012, 30, 360. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.D.; Wang, P.; Bao, Z.; Ma, Q.; Duan, L.J.; Bao, A.K.; Zhang, J.L.; Wang, S.M. SOS1, HKT1;5, and NHX1 Synergistically Modulate Na+ Homeostasis in the Halophytic Grass Puccinellia tenuiflora. Front. Plant Sci. 2017, 8, 576. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.M.; Zhang, J.L.; Liu, X.S.; Li, Z.; Wu, G.Q.; Cai, J.Y.; Flowers, T.J.; Wang, S.M. Puccinellia tenuiflora maintains a low Na+ level under salinity by limiting unidirectional Na+ influx resulting in a high selectivity for K+ over Na+. Plant Cell Environ. 2009, 32, 486–496. [Google Scholar] [CrossRef] [PubMed]
- Deinlein, U.; Stephan, A.B.; Horie, T.; Luo, W.; Xu, G.; Schroeder, J.I. Plant salt-Tolerance mechanisms. Trends Plant Sci. 2014, 19, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Garciadeblas, B.; Senn, M.E.; Banuelos, M.A.; Rodriguez-Navarro, A. Sodium transport and HKT transporters: The rice model. Plant J. 2003, 34, 788–801. [Google Scholar] [CrossRef]
- Uozumi, N.; Kim, E.J.; Rubio, F.; Yamaguchi, T.; Muto, S.; Tsuboi, A.; Bakker, E.P.; Nakamura, T.; Schroeder, J.I. The Arabidopsis HKT1 gene homolog mediates inward Na(+) currents in Xenopus laevis oocytes and Na(+) uptake in Saccharomyces cerevisiae. Plant Physiol. 2000, 122, 1249–1259. [Google Scholar] [CrossRef]
- Oomen, R.J.; Benito, B.; Sentenac, H.; Rodriguez-Navarro, A.; Talon, M.; Very, A.A.; Domingo, C. HKT2;2/1, a K(+)-Permeable transporter identified in a salt-tolerant rice cultivar through surveys of natural genetic polymorphism. Plant J. 2012, 71, 750–762. [Google Scholar] [CrossRef] [PubMed]
- Horie, T.; Costa, A.; Kim, T.H.; Han, M.J.; Horie, R.; Leung, H.Y.; Miyao, A.; Hirochika, H.; An, G.; Schroeder, J.I. Rice OsHKT2;1 transporter mediates large na+ influx component into K+-Starved roots for growth. EMBO J. 2007, 26, 3003–3014. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Yan, J.; Shen, G.; Fu, L.; Holaday, A.S.; Auld, D.; Blumwald, E.; Zhang, H. Expression of an Arabidopsis vacuolar sodium/proton antiporter gene in cotton improves photosynthetic performance under salt conditions and increases fiber yield in the field. Plant Cell Physiol. 2005, 46, 1848–1854. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.Y.; Yang, A.F.; Zhang, K.W.; Zhang, J.R. Production and analysis of transgenic maize with improved salt tolerance by the introduction of AtNHX1 gene. Acta Bot. Sin. 2004, 46, 854–861. [Google Scholar]
- Xue, Z.Y.; Zhi, D.Y.; Xue, G.P.; Zhang, H.; Zhao, Y.X.; Xia, G.M. Enhanced salt tolerance of transgenic wheat (Triticum aestivum L.) expressing a vacuolar Na+/H+ antiporter gene with improved grain yields in saline soils in the field and a reduced level of leaf Na+. Plant Sci. 2004, 167, 849–859. [Google Scholar] [CrossRef]
- Aharon, G.S.; Apse, M.P.; Duan, S.; Hua, X.; Blumwald, E. Characterization of a family of vacuolar Na+/H+ antiporters in Arabidopsis thaliana. Plant Soil 2003, 253, 245–256. [Google Scholar] [CrossRef]
- Rodriguez-Rosales, M.P.; Galvez, F.J.; Huertas, R.; Aranda, M.N.; Baghour, M.; Cagnac, O.; Venema, K. Plant NHX cation/proton antiporters. Plant Signal. Behav. 2009, 4, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.X.; Hodson, J.N.; Williams, J.P.; Blumwald, E. Engineering salt-tolerant brassica plants: Characterization of yield and seed oil quality in transgenic plants with increased vacuolar sodium accumulation. Proc. Natl. Acad. Sci. USA 2001, 98, 12832–12836. [Google Scholar] [CrossRef]
- Ohta, M.; Hayashi, Y.; Nakashima, A.; Hamada, A.; Tanaka, A.; Nakamura, T.; Hayakawa, T. Introduction of a Na+/H+ antiporter gene from Atriplex gmelini confers salt tolerance to rice. FEBS Lett. 2002, 532, 279–282. [Google Scholar] [CrossRef]
- Silva, P.; Geros, H. Regulation by salt of vacuolar H+-Atpase and H+-Pyrophosphatase activities and Na+/H+ exchange. Plant Signal. Behav. 2009, 4, 718–726. [Google Scholar] [CrossRef]
- Baiges, I.; Schäffner, A.R.; Affenzeller, M.J.; Mas, A. Plant aquaporins. Physiol. Plant. 2002, 115, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Hove, R.M.; Ziemann, M.; Bhave, M. Identification and expression analysis of the barley (Hordeum vulgare L.) aquaporin gene family. PLoS ONE 2015, 10, e0128025. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, D.; Cornacchione, M.V.; Ferreira, J.F.; Suarez, D.L. Variable salinity responses of 12 alfalfa genotypes and comparative expression analyses of salt-response genes. Sci. Rep. 2017, 7, 42958. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Wang, T.Z.; Zhang, W.H. Sodium extrusion associated with enhanced expression of SOS1 underlies different salt tolerance between Medicago falcata and Medicago truncatula seedlings. Environ. Exp. Bot. 2015, 110, 46–55. [Google Scholar] [CrossRef]
- Sathee, L.; Sairam, R.K.; Chinnusamy, V.; Jha, S.K. Differential transcript abundance of salt overly sensitive (SOS) pathway genes is a determinant of salinity stress tolerance of wheat. Acta Physiol. Plant. 2015, 37, 169. [Google Scholar] [CrossRef]
- Huanca-Mamani, W.; Ortiz, M.V.; Cárdenas-Ninasivincha, S.; Acosta-Garcia, G.; Bastías, E. Gene expression analysis in response to combined salt and boron (B) stresses in a tolerant maize landrace. Plant Omics 2018, 11, 80–88. [Google Scholar] [CrossRef]
- Jin, H.; Dong, D.; Yang, Q.; Zhu, D. Salt-responsive transcriptome profiling of Suaeda glauca via RNA sequencing. PLoS ONE 2016, 11, e0150504. [Google Scholar] [CrossRef] [PubMed]
- Barragán, V.; Leidi, E.O.; Andrés, Z.; Rubio, L.; De Luca, A.; Fernández, J.A.; Cubero, B.; Pardo, J.M. Ion exchangers NHX1 and NHX2 mediate active potassium uptake into vacuoles to regulate cell turgor and stomatal function in Arabidopsis. Plant Cell 2012, 24, 1127–1142. [Google Scholar] [CrossRef] [PubMed]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pandey, A.; Khan, M.K.; Hakki, E.E.; Gezgin, S.; Hamurcu, M. Combined Boron Toxicity and Salinity Stress—An Insight into Its Interaction in Plants. Plants 2019, 8, 364. https://doi.org/10.3390/plants8100364
Pandey A, Khan MK, Hakki EE, Gezgin S, Hamurcu M. Combined Boron Toxicity and Salinity Stress—An Insight into Its Interaction in Plants. Plants. 2019; 8(10):364. https://doi.org/10.3390/plants8100364
Chicago/Turabian StylePandey, Anamika, Mohd Kamran Khan, Erdogan Esref Hakki, Sait Gezgin, and Mehmet Hamurcu. 2019. "Combined Boron Toxicity and Salinity Stress—An Insight into Its Interaction in Plants" Plants 8, no. 10: 364. https://doi.org/10.3390/plants8100364
APA StylePandey, A., Khan, M. K., Hakki, E. E., Gezgin, S., & Hamurcu, M. (2019). Combined Boron Toxicity and Salinity Stress—An Insight into Its Interaction in Plants. Plants, 8(10), 364. https://doi.org/10.3390/plants8100364