Variation in Aquaporin and Physiological Responses Among Pinus contorta Families Under Different Moisture Conditions

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
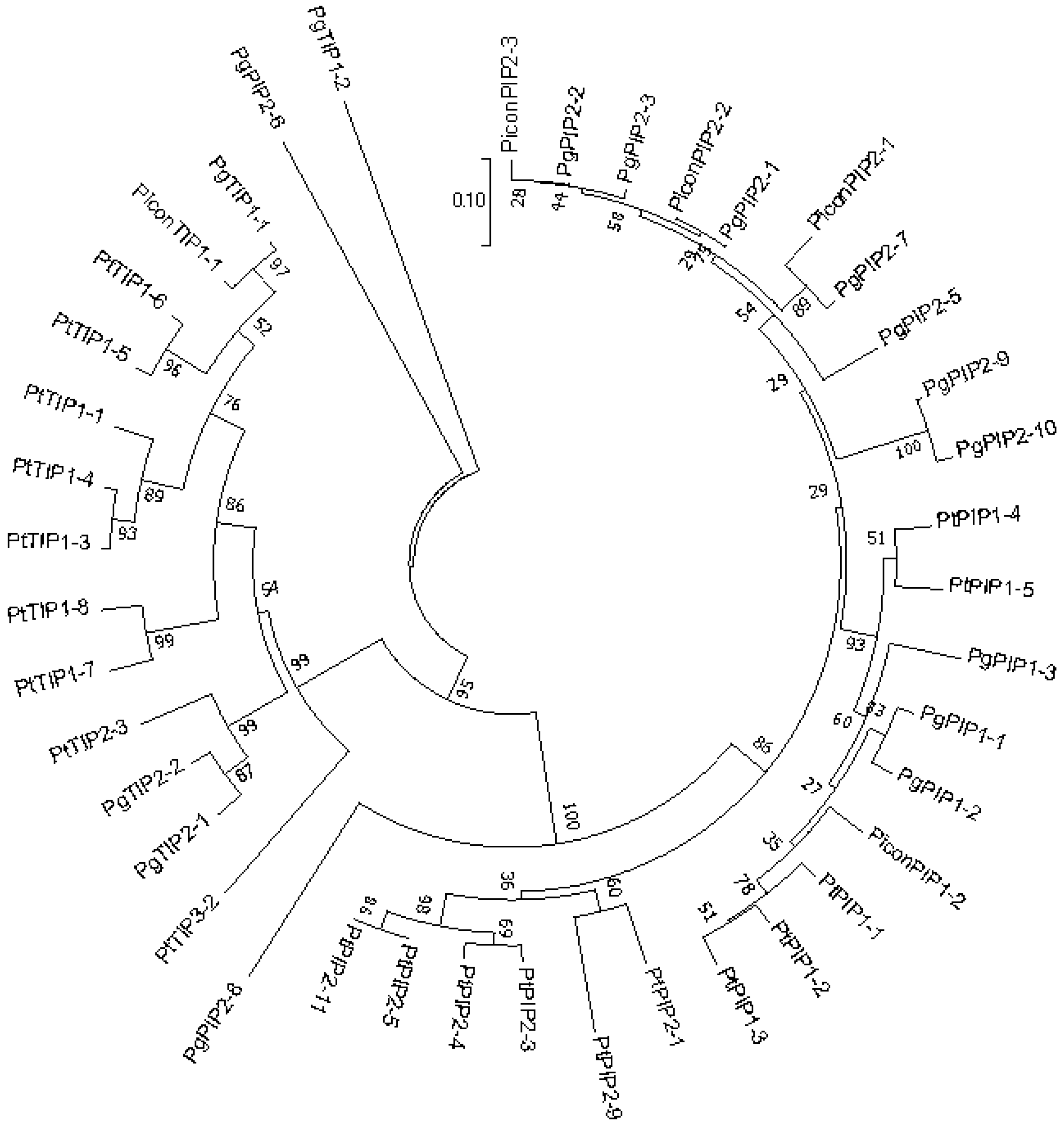
2.1. Cloning and Nomenclature
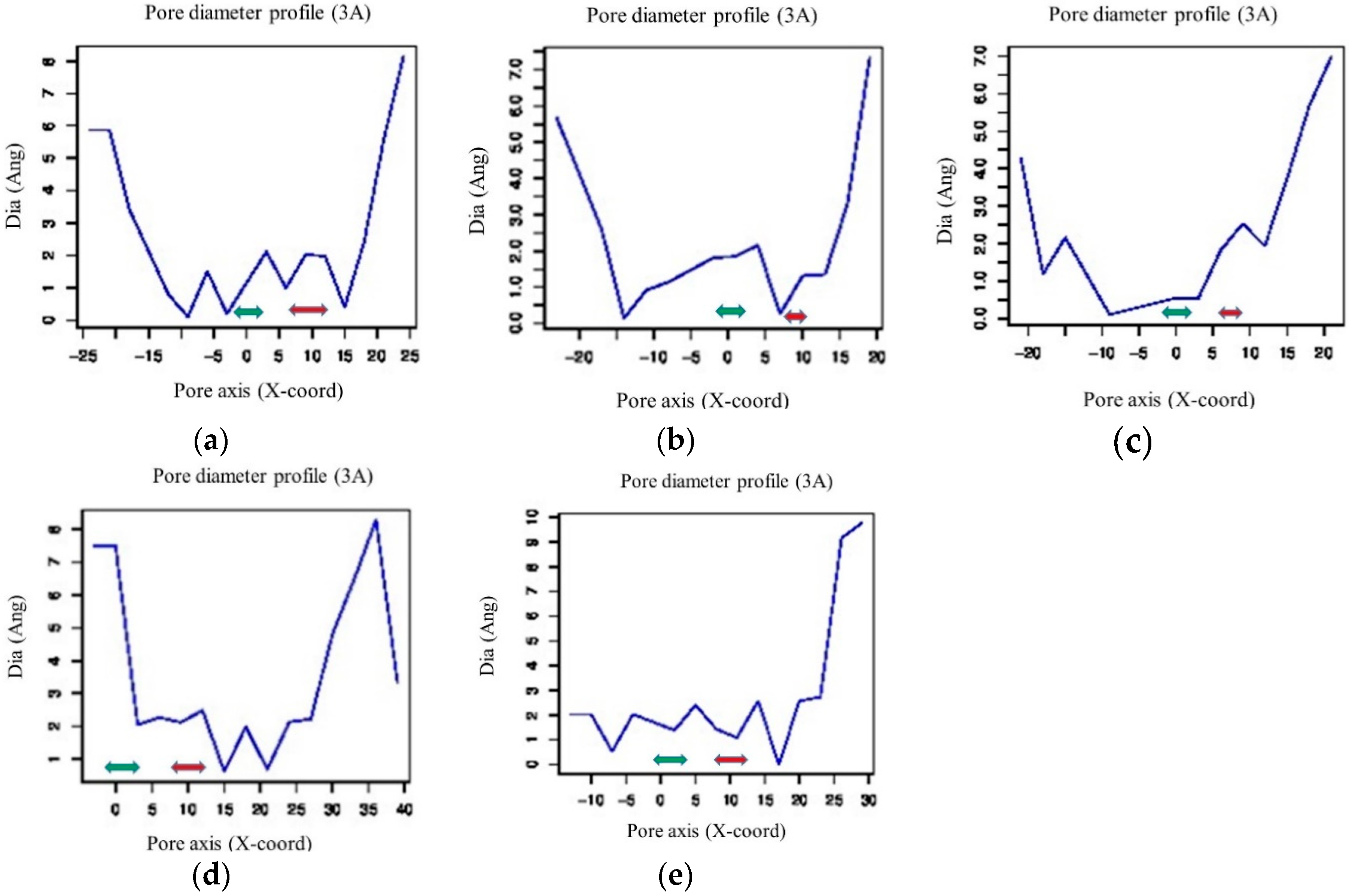
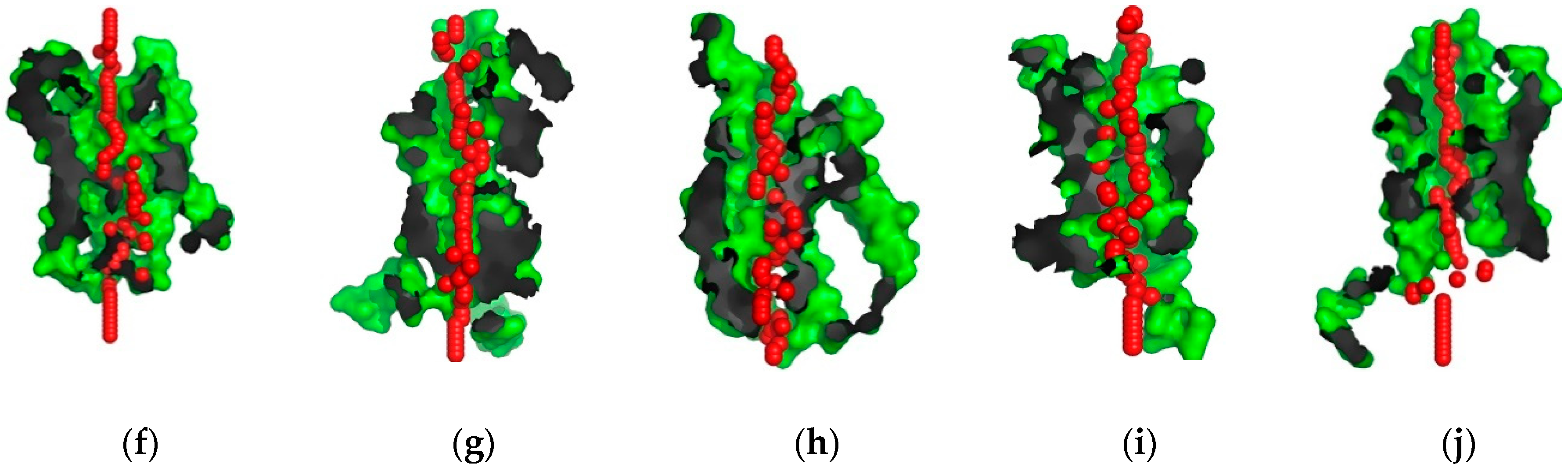
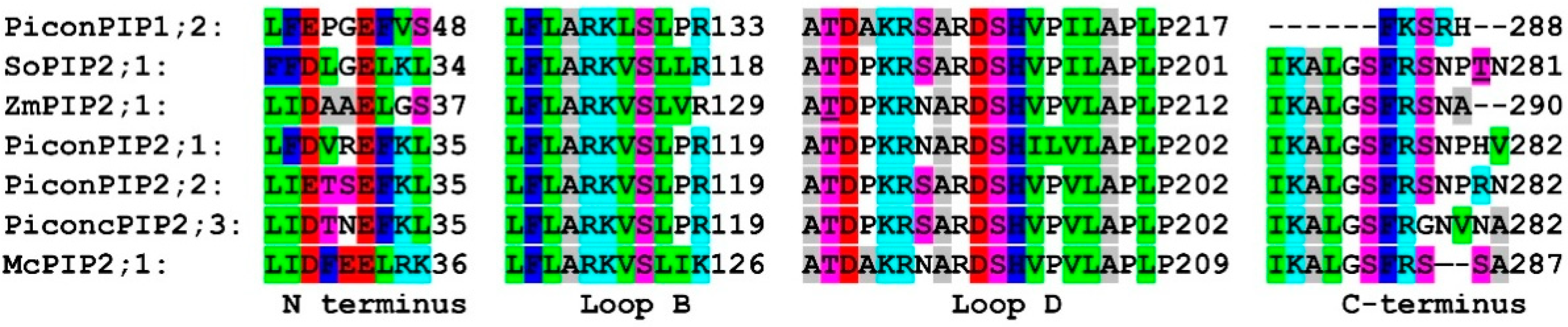
2.2. Structural Analysis
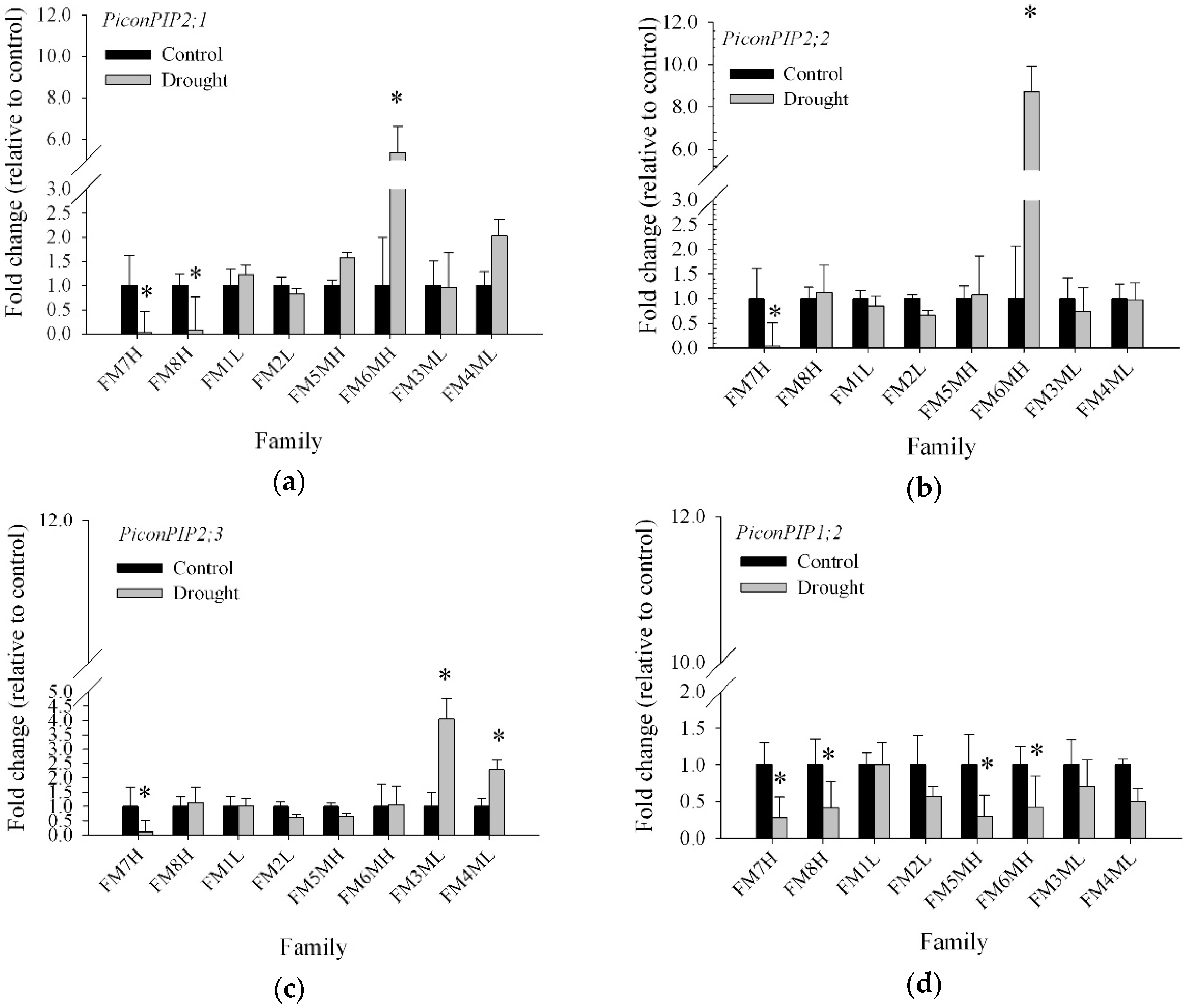
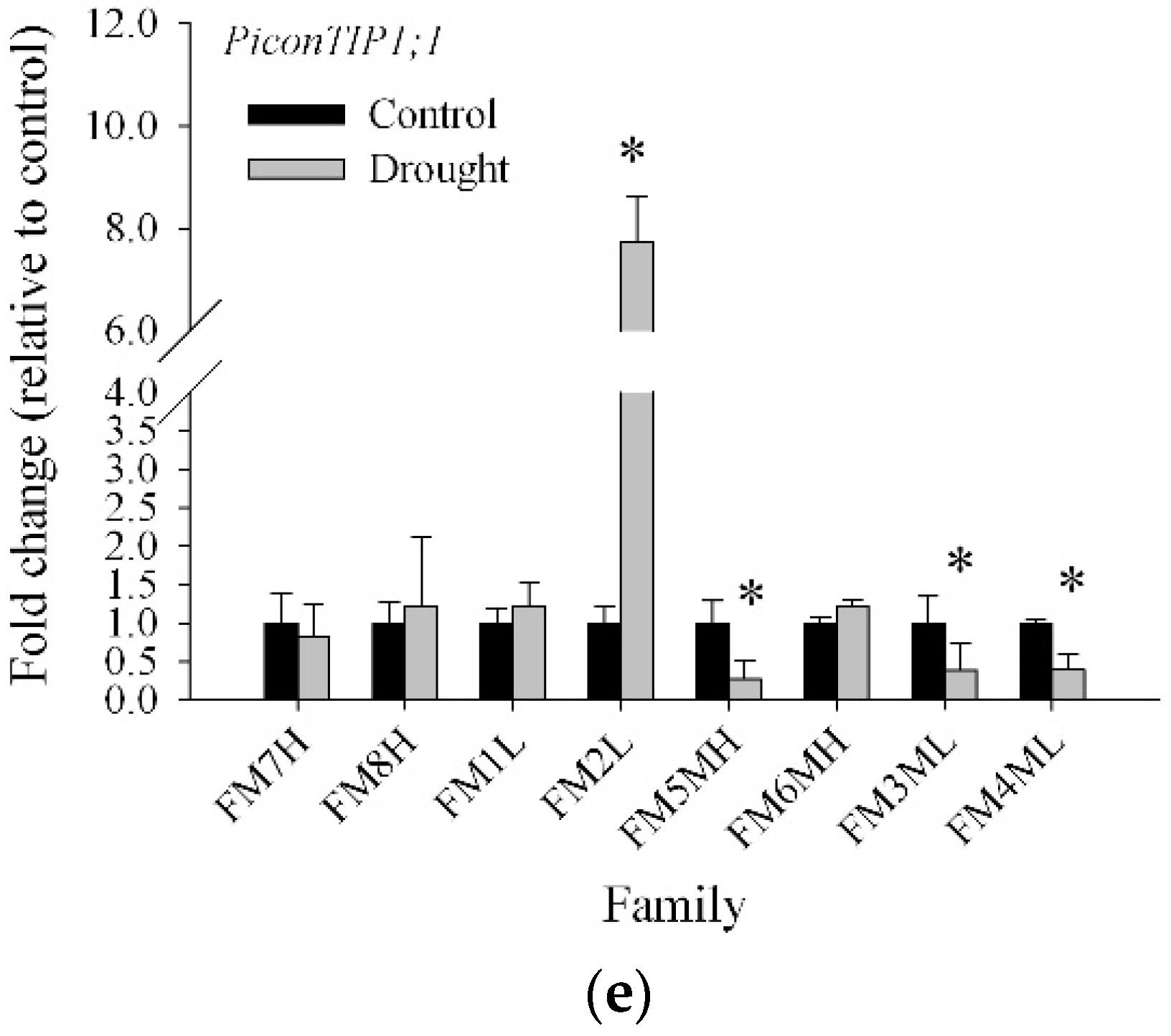
2.3. Transcript Abundance of Five Pinus Contorta Aquaporins
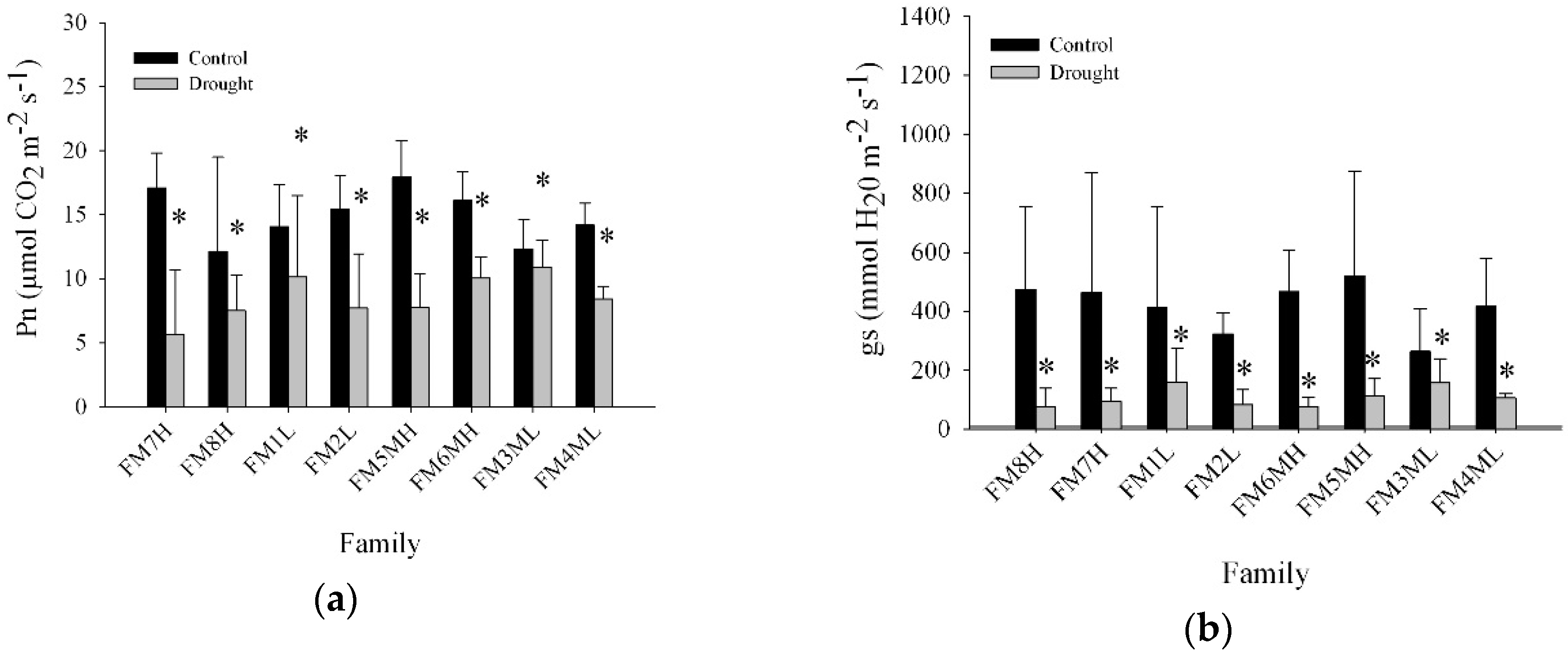
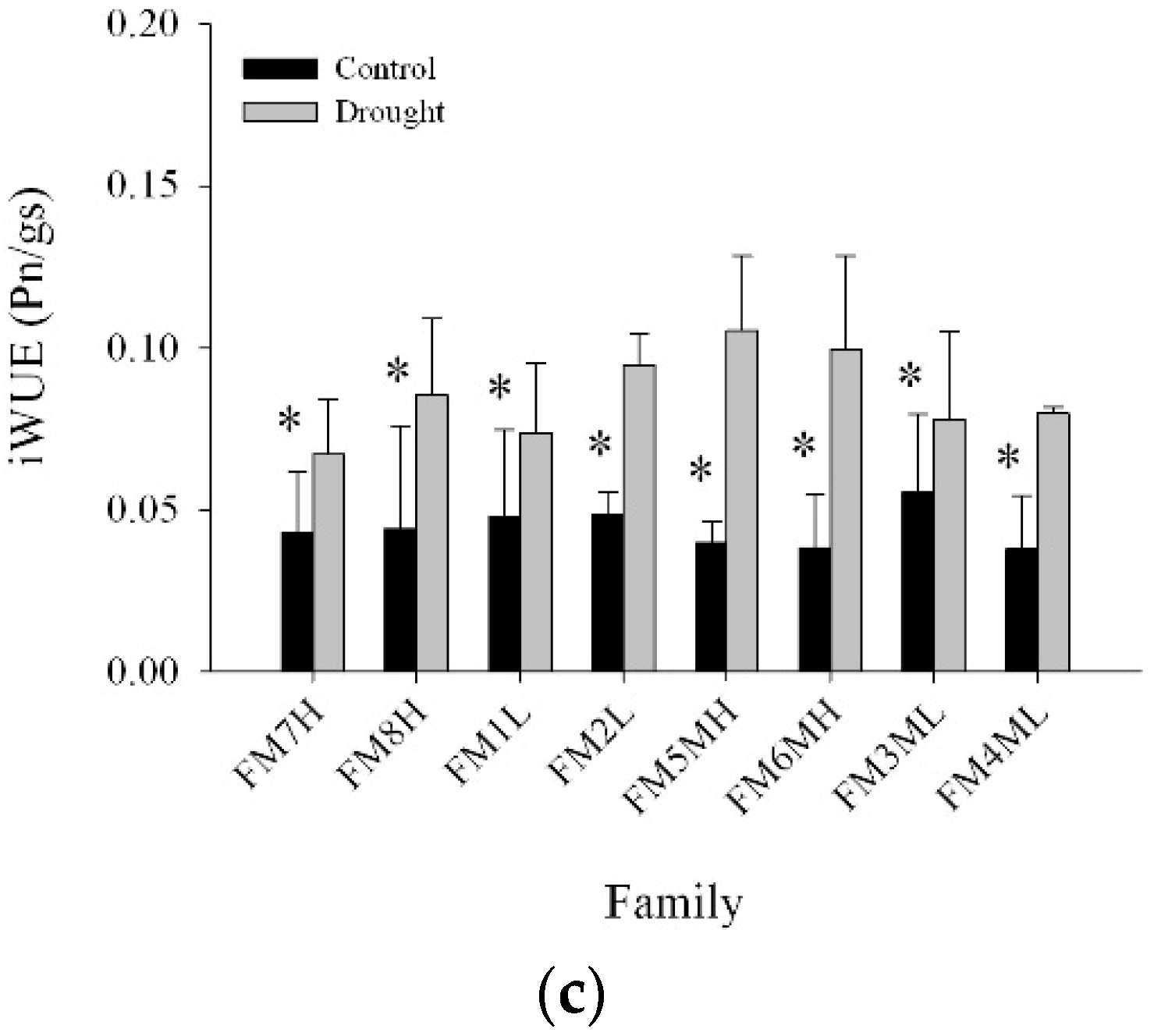
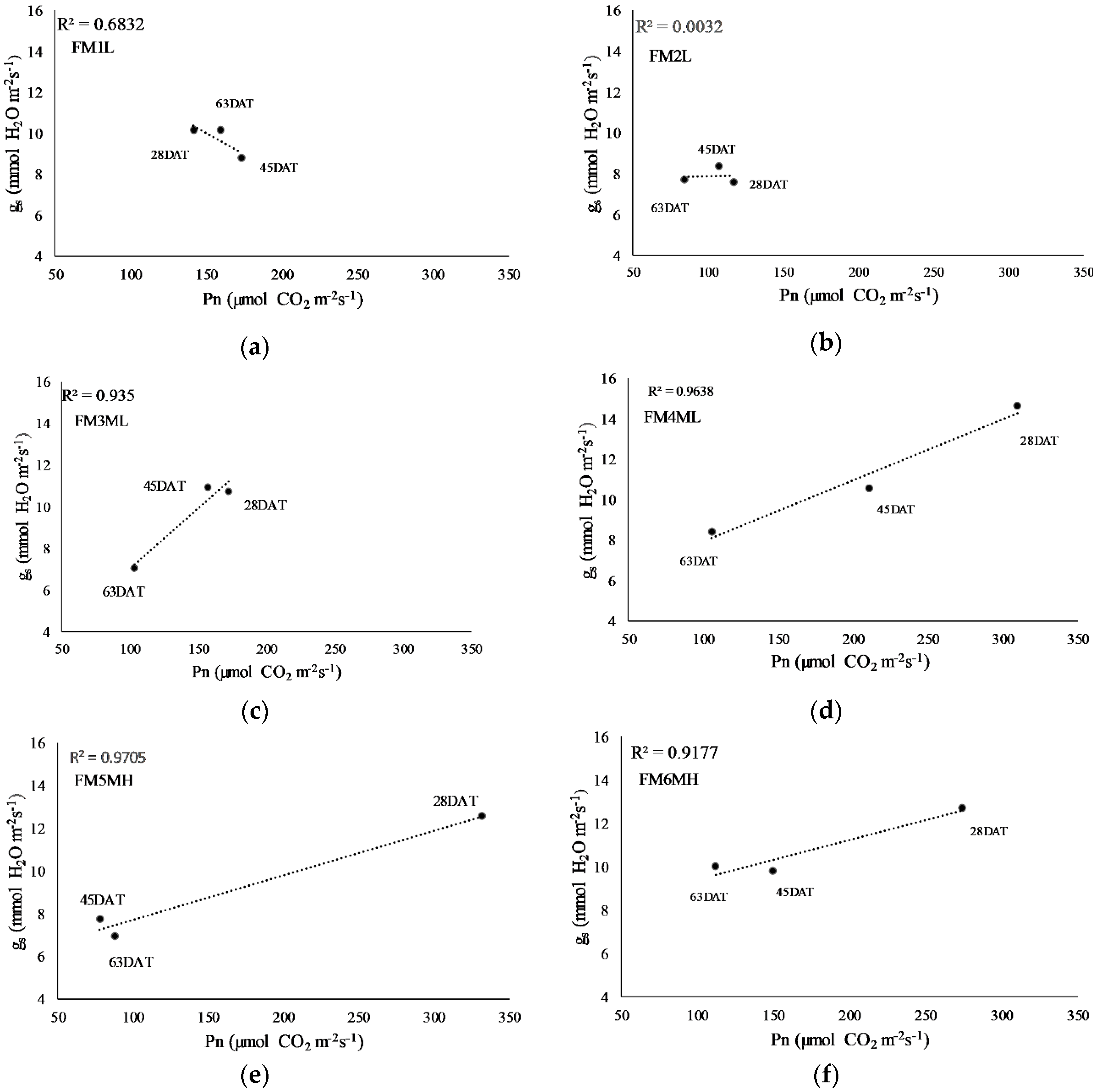
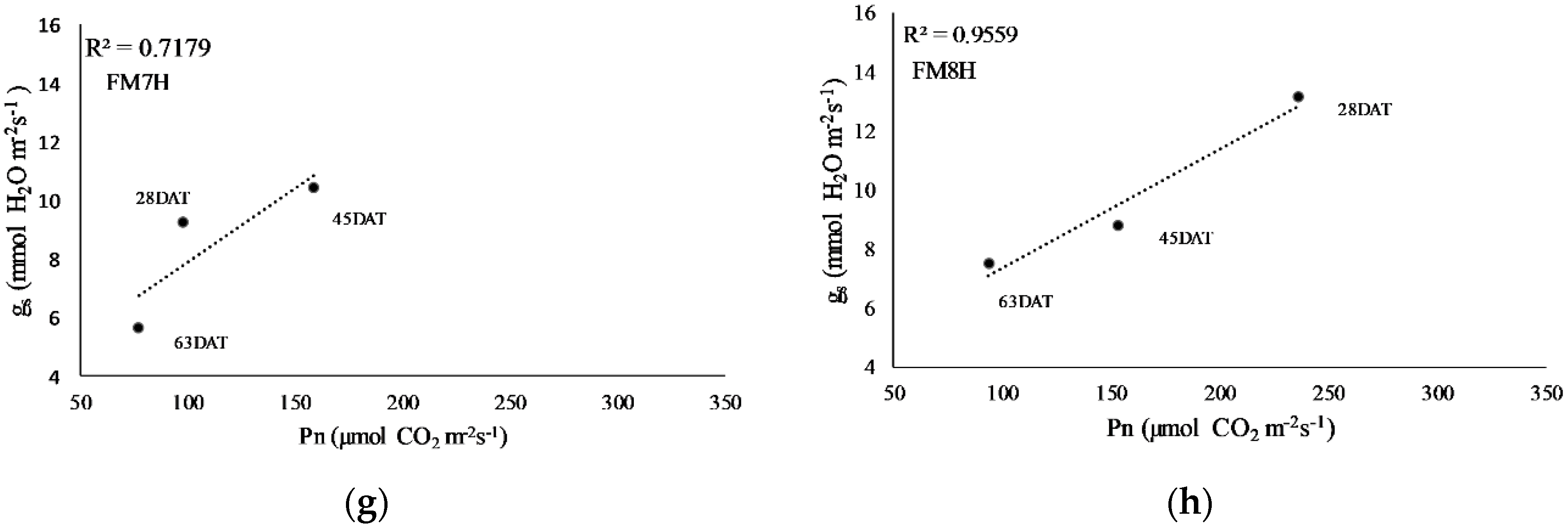
2.4. Net Photosynthesis, Intrinsic Water Use Efficiency, and Stomatal Conductance
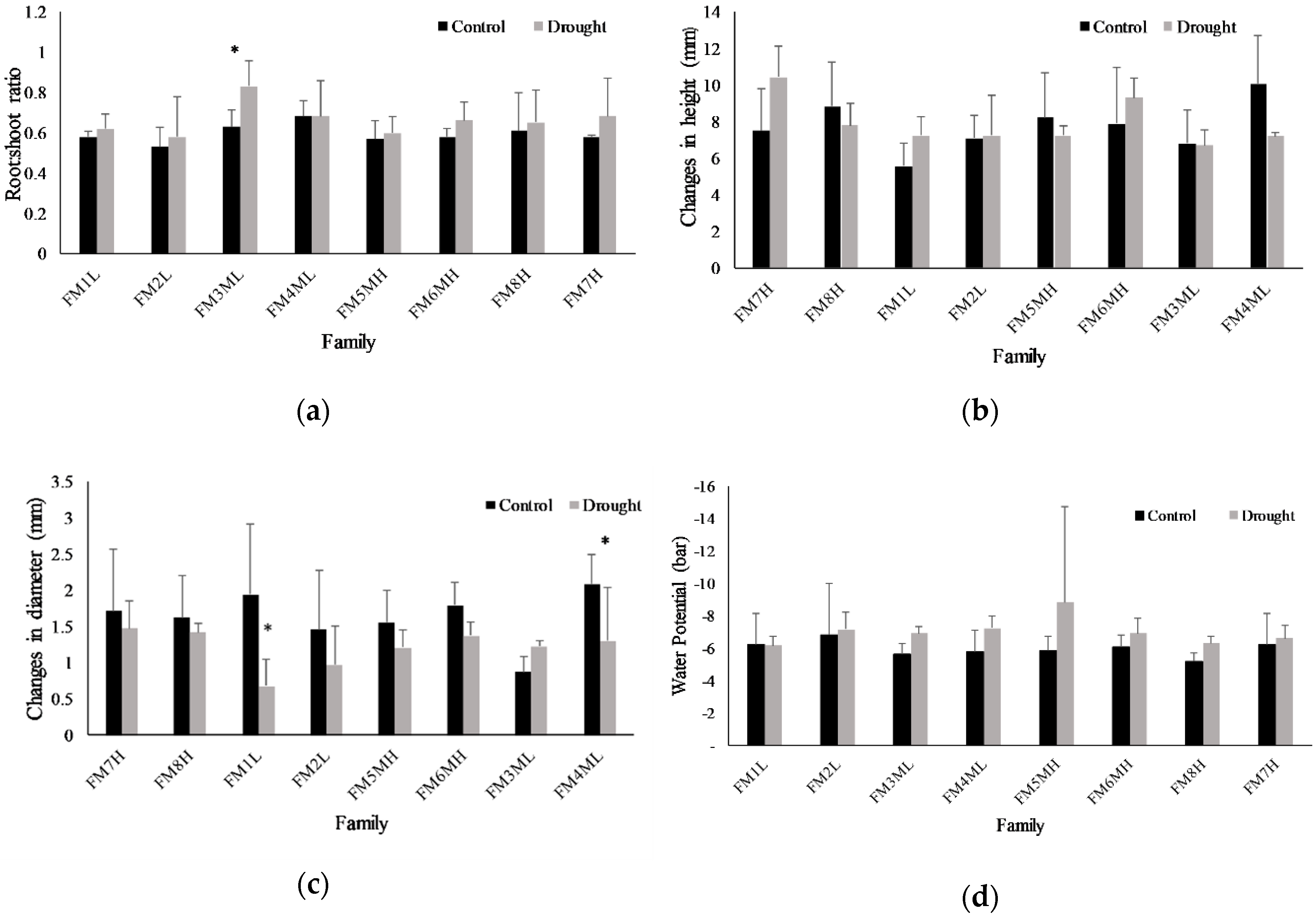
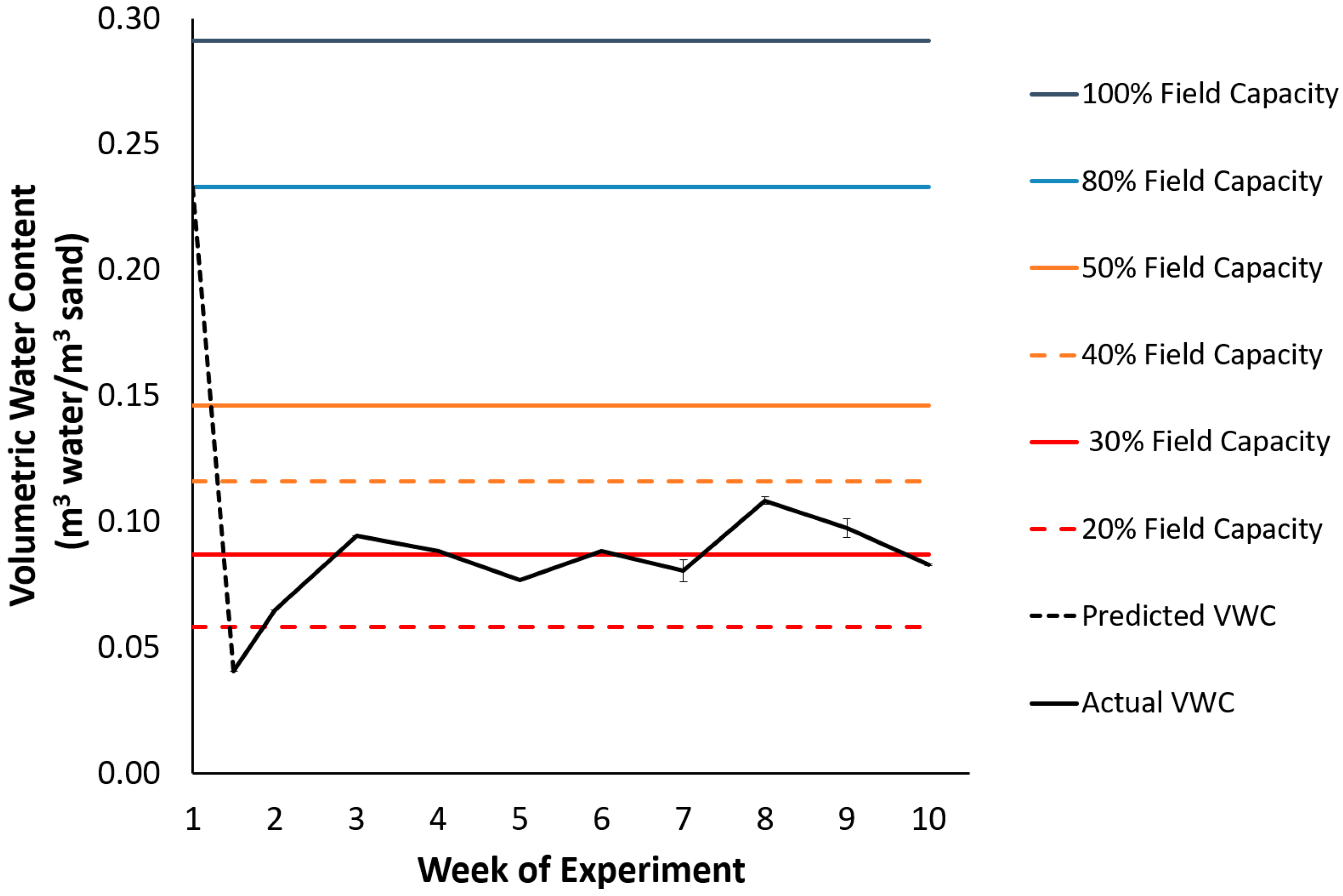
2.5. Growth Parameters and Water Content
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Drought Treatments and Experimental Design
4.3. Measurements
4.4. Database Sources
4.5. RNA Extraction, cDNA Synthesis, and RT-qPCR Analysis
4.6. Rapid Amplification of cDNA 3′ End and Cloning
4.7. Nomenclature of Pinus contorta Aquaporin Genes
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| PIP | plasma membrane intrinsic protein |
| TIP | tonoplast intrinsic protein |
| PiconAQPs | Pinus contorta aquaporins |
| DAT | days after treatment |
| Picon | Pinus contorta |
| AQPs | Aquaporins |
| ar/R | aromatic/arginine |
References
- Desprez-Loustau, M.; Marçais, B.; Nageleisen, L.; Piou, D.; Vannini, A. Interactive Effects of Drought and Pathogens in Forest Trees. Ann. For. Sci. 2006, 63, 597–612. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.T.; et al. A Global Overview of Drought and Heat-Induced Tree Mortality Reveals Emerging Climate Change Risks for Forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Zhao, M.; Running, S.W. Drought-Induced Reduction in Global Terrestrial Net Primary Production from 2000 through 2009. Science 2010, 329, 940–943. [Google Scholar] [CrossRef] [PubMed]
- Reichstein, M.; Bahn, M.; Ciais, P.; Frank, D.; Mahecha, M.D.; Seneviratne, S.I.; Zscheischler, J.; Beer, C.; Buchmann, N.; Frank, D.C.; et al. Climate Extremes and the Carbon Cycle. Nature 2013, 500, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Park Williams, A.; Allen, C.D.; Macalady, A.K.; Griffin, D.; Woodhouse, C.A.; Meko, D.M.; Swetnam, T.W.; Rauscher, S.A.; Seager, R.; Grissino-Mayer, H.D.; et al. Temperature as a Potent Driver of Regional Forest Drought Stress and Tree Mortality. Nat. Clim. Chang. 2013, 3, 292–297. [Google Scholar] [CrossRef]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On Underestimation of Global Vulnerability to Tree Mortality and Forest Die-Off from Hotter Drought in the Anthropocene. Ecosphere 2015, 6, 1–55. [Google Scholar] [CrossRef]
- Anderegg, W.R.; Schwalm, C.; Biondi, F.; Camarero, J.J.; Koch, G.; Litvak, M.; Ogle, K.; Shaw, J.D.; Shevliakova, E.; Williams, A.P.; et al. FOREST ECOLOGY. Pervasive Drought Legacies in Forest Ecosystems and their Implications for Carbon Cycle Models. Science 2015, 349, 528–532. [Google Scholar] [CrossRef]
- Netherer, S.; Matthews, B.; Katzensteiner, K.; Blackwell, E.; Henschke, P.; Hietz, P.; Pennerstorfer, J.; Rosner, S.; Kikuta, S.; Schume, H.; et al. Do Water-Limiting Conditions Predispose Norway Spruce to Bark Beetle Attack? New Phytol. 2015, 205, 1128–1141. [Google Scholar] [CrossRef]
- Brunner, I.; Herzog, C.; Dawes, M.A.; Arend, M.; Sperisen, C. How Tree Roots Respond to Drought. Front. Plant Sci. 2015, 6, 547. [Google Scholar] [CrossRef]
- Basu, S.; Ramegowda, V.; Kumar, A.; Pereira, A. Plant Adaptation to Drought Stress. F1000Res 2016, 5. [Google Scholar] [CrossRef]
- Pou, A.; Medrano, H.; Tomàs, M.; Martorell, S.; Ribas-Carbó, M.; Flexas, J. Anisohydric Behaviour in Grapevines Results in Better Performance Under Moderate Water Stress and Recovery than Isohydric Behaviour. Plant Soil 2012, 359, 335–349. [Google Scholar] [CrossRef]
- Nolf, M.; Creek, D.; Duursma, R.; Holtum, J.; Mayr, S.; Choat, B. Stem and Leaf Hydraulic Properties are Finely Coordinated in Three Tropical Rain Forest Tree Species. Plant Cell Environ. 2015, 38, 2652–2661. [Google Scholar] [CrossRef] [PubMed]
- Zargar, S.M.; Nagar, P.; Deshmukh, R.; Nazir, M.; Wani, A.A.; Masoodi, K.Z.; Agrawal, G.K.; Rakwal, R. Aquaporins as Potential Drought Tolerance Inducing Proteins: Towards Instigating Stress Tolerance. J. Proteom. 2017, 169, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Maurel, C.; Verdoucq, L.; Luu, D.T.; Santoni, V. Plant Aquaporins: Membrane Channels with Multiple Integrated Functions. Annu. Rev. Plant Biol. 2008, 59, 595–624. [Google Scholar] [CrossRef] [PubMed]
- Knepper, M.A. The Aquaporin Family of Molecular Water Channels. Proc. Natl. Acad. Sci. USA 1994, 91, 6255–6258. [Google Scholar] [CrossRef] [PubMed]
- Siefritz, F.; Tyree, M.T.; Lovisolo, C.; Schubert, A.; Kaldenhoff, R. PIP1 Plasma Membrane Aquaporins in Tobacco: From Cellular Effects to Function in Plants. Plant Cell 2002, 14, 869–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyerman, S.D.; Niemietz, C.M.; Bramley, H. Plant Aquaporins: Multifunctional Water and Solute Channels with Expanding Roles. Plant Cell Environ. 2002, 25, 173–194. [Google Scholar] [CrossRef]
- Maurel, C. Plant Aquaporins: Novel Functions and Regulation Properties. FEBS Lett. 2007, 581, 2227–2236. [Google Scholar] [CrossRef]
- Sade, N.; Gebretsadik, M.; Seligmann, R.; Schwartz, A.; Wallach, R.; Moshelion, M. The Role of Tobacco Aquaporin1 in Improving Water use Efficiency, Hydraulic Conductivity, and Yield Production Under Salt Stress. Plant Physiol. 2010, 152, 245–254. [Google Scholar] [CrossRef]
- Moshelion, M.; Halperin, O.; Wallach, R.; Oren, R.; Way, D.A. Role of Aquaporins in Determining Transpiration and Photosynthesis in Water-Stressed Plants: Crop Water-use Efficiency, Growth and Yield. Plant Cell Environ. 2015, 38, 1785–1793. [Google Scholar] [CrossRef]
- Chaumont, F.; Tyerman, S.D. Aquaporins: Highly Regulated Channels Controlling Plant Water Relations. Plant Physiol. 2014, 164, 1600–1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaumont, F.; Barrieu, F.; Wojcik, E.; Chrispeels, M.J.; Jung, R. Aquaporins Constitute a Large and Highly Divergent Protein Family in Maize. Plant Physiol. 2001, 125, 1206–1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luu, D.; Maurel, C. Aquaporins in a Challenging Environment: Molecular Gears for Adjusting Plant Water Status. Plant Cell Environ. 2005, 28, 85–96. [Google Scholar] [CrossRef]
- Lu, S.; Wang, J. Homoharringtonine and Omacetaxine for Myeloid Hematological Malignancies. J. Hematol. Oncol. 2014, 7, 2. [Google Scholar] [CrossRef] [PubMed]
- Maurel, C.; Boursiac, Y.; Luu, D.T.; Santoni, V.; Shahzad, Z.; Verdoucq, L. Aquaporins in Plants. Physiol. Rev. 2015, 95, 1321–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zwiazek, J.J.; Xu, H.; Tan, X.; Navarro-Rodenas, A.; Morte, A. Significance of Oxygen Transport through Aquaporins. Sci. Rep. 2017, 7, 40411. [Google Scholar] [CrossRef] [PubMed]
- Fujiyoshi, Y. Structure and Function of Water Channels. Tanpakushitsu Kakusan Koso 2005, 50, 1278–1283. [Google Scholar] [CrossRef]
- Gonen, T.; Walz, T. The Structure of Aquaporins. Q. Rev. Biophys. 2006, 39, 361–396. [Google Scholar] [CrossRef]
- Jung, J.S.; Preston, G.M.; Smith, B.L.; Guggino, W.B.; Agre, P. Molecular Structure of the Water Channel through Aquaporin CHIP. The Hourglass Model. J. Biol. Chem. 1994, 269, 14648–14654. [Google Scholar] [PubMed]
- Tan, X.; Xu, H.; Khan, S.; Equiza, M.A.; Lee, S.H.; Vaziriyeganeh, M.; Zwiazek, J.J. Plant Water Transport and Aquaporins in Oxygen-Deprived Environments. J. Plant Physiol. 2018, 227, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Tornroth-Horsefield, S.; Wang, Y.; Hedfalk, K.; Johanson, U.; Karlsson, M.; Tajkhorshid, E.; Neutze, R.; Kjellbom, P. Structural Mechanism of Plant Aquaporin Gating. Nature 2006, 439, 688–694. [Google Scholar] [CrossRef]
- Kreida, S.; Tornroth-Horsefield, S. Structural Insights into Aquaporin Selectivity and Regulation. Curr. Opin. Struct. Biol. 2015, 33, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, R.K.; Vivancos, J.; Ramakrishnan, G.; Guerin, V.; Carpentier, G.; Sonah, H.; Labbe, C.; Isenring, P.; Belzile, F.J.; Belanger, R.R. A Precise Spacing between the NPA Domains of Aquaporins is Essential for Silicon Permeability in Plants. Plant J. 2015, 83, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Maury, L.; Marguerat, S.; Bahler, J. Tuning Gene Expression to Changing Environments: From Rapid Responses to Evolutionary Adaptation. Nat. Rev. Genet. 2008, 9, 583–593. [Google Scholar] [CrossRef]
- Zupin, M.; Sedlar, A.; Kidric, M.; Meglic, V. Drought-Induced Expression of Aquaporin Genes in Leaves of Two Common Bean Cultivars Differing in Tolerance to Drought Stress. J. Plant Res. 2017, 130, 735–745. [Google Scholar] [CrossRef] [PubMed]
- Almeida-Rodriguez, A.M.; Hacke, U.G.; Laur, J. Influence of Evaporative Demand on Aquaporin Expression and Root Hydraulics of Hybrid Poplar. Plant Cell Environ. 2011, 34, 1318–1331. [Google Scholar] [CrossRef]
- Surbanovski, N.; Sargent, D.J.; Else, M.A.; Simpson, D.W.; Zhang, H.; Grant, O.M. Expression of Fragaria Vesca PIP Aquaporins in Response to Drought Stress: PIP Down-Regulation Correlates with the Decline in Substrate Moisture Content. PLoS ONE 2013, 8, e74945. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, M.D.; Silva, R.L.; Costa Ferreira Neto, J.R.; Guimaraes, A.C.; Veiga, D.T.; Chabregas, S.M.; Burnquist, W.L.; Kahl, G.; Benko-Iseppon, A.M.; Kido, E.A. Expression Analysis of Sugarcane Aquaporin Genes Under Water Deficit. J. Nucleic Acids 2013, 2013, 1–14. [Google Scholar] [CrossRef]
- Afzal, Z.; Howton, T.C.; Sun, Y.; Mukhtar, M.S. The Roles of Aquaporins in Plant Stress Responses. J. Dev. Biol. 2016, 4, 9. [Google Scholar] [CrossRef] [PubMed]
- Pawlowicz, I.; Rapacz, M.; Perlikowski, D.; Gondek, K.; Kosmala, A. Abiotic Stresses Influence the Transcript Abundance of PIP and TIP Aquaporins in Festuca Species. J. Appl. Genet. 2017, 58, 421–435. [Google Scholar] [CrossRef]
- Cramer, G.R.; Ergul, A.; Grimplet, J.; Tillett, R.L.; Tattersall, E.A.; Bohlman, M.C.; Vincent, D.; Sonderegger, J.; Evans, J.; Osborne, C.; et al. Water and Salinity Stress in Grapevines: Early and Late Changes in Transcript and Metabolite Profiles. Funct. Integr. Genom. 2007, 7, 111–134. [Google Scholar] [CrossRef] [PubMed]
- Lienard, D.; Durambur, G.; Kiefer-Meyer, M.C.; Nogue, F.; Menu-Bouaouiche, L.; Charlot, F.; Gomord, V.; Lassalles, J.P. Water Transport by Aquaporins in the Extant Plant Physcomitrella Patens. Plant Physiol. 2008, 146, 1207–1218. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.B.; Sankararamakrishnan, R. Genome-Wide Analysis of Major Intrinsic Proteins in the Tree Plant Populus trichocarpa: Characterization of XIP Subfamily of Aquaporins from Evolutionary Perspective. BMC Plant Biol. 2009, 9, 134. [Google Scholar] [CrossRef] [PubMed]
- Laur, J.; Hacke, U.G. Exploring Picea Glauca Aquaporins in the Context of Needle Water Uptake and Xylem Refilling. New Phytol. 2014, 203, 388–400. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The Neighbor-Joining Method: A New Method for Reconstructing Phylogenetic Trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Yachdav, G.; Kloppmann, E.; Kajan, L.; Hecht, M.; Goldberg, T.; Hamp, T.; Honigschmid, P.; Schafferhans, A.; Roos, M.; Bernhofer, M.; et al. PredictProtein—An Open Resource for Online Prediction of Protein Structural and Functional Features. Nucleic Acids Res. 2014, 42, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini-Calace, M.; Maiwald, T.; Thornton, J.M. PoreWalker: A Novel Tool for the Identification and Characterization of Channels in Transmembrane Proteins from their Three-Dimensional Structure. PLoS Comput. Biol. 2009, 5, e1000440. [Google Scholar] [CrossRef]
- Amezcua-Romero, J.C.; Pantoja, O.; Vera-Estrella, R. Ser123 is Essential for the Water Channel Activity of McPIP2;1 from Mesembryanthemum crystallinum. J. Biol. Chem. 2010, 285, 16739–16747. [Google Scholar] [CrossRef]
- Sui, H.; Han, B.G.; Lee, J.K.; Walian, P.; Jap, B.K. Structural Basis of Water-Specific Transport through the AQP1 Water Channel. Nature 2027, 414, 872–878. [Google Scholar] [CrossRef]
- Gonen, T.; Sliz, P.; Kistler, J.; Cheng, Y.; Walz, T. Aquaporin-0 Membrane Junctions Reveal the Structure of a Closed Water Pore. Nature 2004, 429, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Harries, W.E.; Akhavan, D.; Miercke, L.J.; Khademi, S.; Stroud, R.M. The Channel Architecture of Aquaporin 0 at a 2.2-A Resolution. Proc. Natl. Acad. Sci. USA 2004, 101, 14045–14050. [Google Scholar] [CrossRef] [PubMed]
- Fu, D.; Libson, A.; Miercke, L.J.; Weitzman, C.; Nollert, P.; Krucinski, J.; Stroud, R.M. Structure of a Glycerol-Conducting Channel and the Basis for its Selectivity. Science 2000, 290, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Savage, D.F.; Egea, P.F.; Robles-Colmenares, Y.; O’Connell, J.D.; Stroud, R.M. Architecture and Selectivity in Aquaporins: 2.5 a X-ray Structure of Aquaporin Z. PLoS Biol. 2003, 1, e72. [Google Scholar] [CrossRef] [PubMed]
- Fischer, G.; Kosinska-Eriksson, U.; Aponte-Santamaria, C.; Palmgren, M.; Geijer, C.; Hedfalk, K.; Hohmann, S.; de Groot, B.L.; Neutze, R.; Lindkvist-Petersson, K. Crystal Structure of a Yeast Aquaporin at 1.15 Angstrom Reveals a Novel Gating Mechanism. PLoS Biol. 2009, 7, e1000130. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.K.; Kozono, D.; Remis, J.; Kitagawa, Y.; Agre, P.; Stroud, R.M. Structural Basis for Conductance by the Archaeal Aquaporin AqpM at 1.68 A. Proc. Natl. Acad. Sci. USA 2005, 102, 18932–18937. [Google Scholar] [CrossRef] [PubMed]
- Kirscht, A.; Kaptan, S.S.; Bienert, G.P.; Chaumont, F.; Nissen, P.; de Groot, B.L.; Kjellbom, P.; Gourdon, P.; Johanson, U. Crystal Structure of an Ammonia-Permeable Aquaporin. PLoS Biol. 2016, 14, e1002411. [Google Scholar] [CrossRef]
- Chothia, C.; Lesk, A.M. The Relation between the Divergence of Sequence and Structure in Proteins. EMBO J. 1986, 5, 823–826. [Google Scholar] [CrossRef]
- Pou, A.; Medrano, H.; Flexas, J.; Tyerman, S.D. A Putative Role for TIP and PIP Aquaporins in Dynamics of Leaf Hydraulic and Stomatal Conductances in Grapevine Under Water Stress and Re-Watering. Plant Cell Environ. 2013, 36, 828–843. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Z.; Chai, T.; Wen, Z.; Zhang, H. Indian Mustard Aquaporin Improves Drought and Heavy-Metal Resistance in Tobacco. Mol. Biotechnol. 2008, 40, 280–292. [Google Scholar] [CrossRef]
- Cui, X.H.; Hao, F.S.; Chen, H.; Chen, J.; Wang, X.C. Expression of the Vicia Faba VfPIP1 Gene in Arabidopsis thaliana Plants Improves their Drought Resistance. J. Plant Res. 2008, 121, 207–214. [Google Scholar] [CrossRef]
- Deshmukh, R.K.; Sonah, H.; Belanger, R.R. Plant Aquaporins: Genome-Wide Identification, Transcriptomics, Proteomics, and Advanced Analytical Tools. Front. Plant Sci. 2016, 7, 1896. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Ballesta Mdel, C.; Carvajal, M. New Challenges in Plant Aquaporin Biotechnology. Plant Sci. 2014, 217–218, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Wang, C.; Liu, R.; Han, Q.; Vandeleur, R.K.; Du, J.; Tyerman, S.; Shou, H. Constitutive Overexpression of Soybean Plasma Membrane Intrinsic Protein GmPIP1;6 Confers Salt Tolerance. BMC Plant Biol. 2014, 14, 181. [Google Scholar] [CrossRef] [PubMed]
- Horsefield, R.; Norden, K.; Fellert, M.; Backmark, A.; Tornroth-Horsefield, S.; Terwisscha van Scheltinga, A.C.; Kvassman, J.; Kjellbom, P.; Johanson, U.; Neutze, R. High-Resolution X-ray Structure of Human Aquaporin 5. Proc. Natl. Acad. Sci. USA 2008, 105, 13327–13332. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.N.; Cerda, J. Evolution and Functional Diversity of Aquaporins. Biol. Bull. 2015, 229, 6–23. [Google Scholar] [CrossRef] [PubMed]
- Shivaraj, S.M.; Deshmukh, R.K.; Rai, R.; Belanger, R.; Agrawal, P.K.; Dash, P.K. Genome-Wide Identification, Characterization, and Expression Profile of Aquaporin Gene Family in Flax (Linum usitatissimum). Sci. Rep. 2017, 7, 46137. [Google Scholar] [CrossRef]
- Wallace, I.S.; Roberts, D.M. Homology Modeling of Representative Subfamilies of Arabidopsis Major Intrinsic Proteins. Classification Based on the Aromatic/Arginine Selectivity Filter. Plant Physiol. 2004, 135, 1059–1068. [Google Scholar] [CrossRef] [Green Version]
- Beitz, E.; Wu, B.; Holm, L.M.; Schultz, J.E.; Zeuthen, T. Point Mutations in the Aromatic/Arginine Region in Aquaporin 1 Allow Passage of Urea, Glycerol, Ammonia, and Protons. Proc. Natl. Acad. Sci. USA 2006, 103, 269–274. [Google Scholar] [CrossRef]
- Johansson, I.; Karlsson, M.; Shukla, V.K.; Chrispeels, M.J.; Larsson, C.; Kjellbom, P. Water Transport Activity of the Plasma Membrane Aquaporin PM28A is Regulated by Phosphorylation. Plant Cell 1998, 10, 451–459. [Google Scholar] [CrossRef]
- Shibasaka, M.; Sasano, S.; Utsugi, S.; Katsuhara, M. Functional Characterization of a Novel Plasma Membrane Intrinsic Protein2 in Barley. Plant Signal. Behav. 2012, 7, 1648–1652. [Google Scholar] [CrossRef]
- Hove, R.M.; Bhave, M. Plant Aquaporins with Non-Aqua Functions: Deciphering the Signature Sequences. Plant Mol. Biol. 2011, 75, 413–430. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.; Sun, W. Arabidopsis Seed-Specific Vacuolar Aquaporins are Involved in Maintaining Seed Longevity Under the Control of ABSCISIC ACID INSENSITIVE 3. J. Exp. Bot. 2015, 66, 4781–4794. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, L.; Liu, M.; Yuan, X.; Yang, Z.; Huang, B. Physiological Effects of Aquaporin in Regulating Drought Tolerance through Overexpressing of Festuca Arundinacea Aquaporin Gene FaPIP2;1. J. Am. Soc. Hortic. Sci. 2015, 140, 404–412. [Google Scholar]
- Uehlein, N.; Otto, B.; Eilingsfeld, A.; Itel, F.; Meier, W.; Kaldenhoff, R. Gas-Tight Triblock-Copolymer Membranes are Converted to CO2 Permeable by Insertion of Plant Aquaporins. Sci. Rep. 2012, 2, 538. [Google Scholar] [CrossRef]
- Heckwolf, M.; Pater, D.; Hanson, D.T.; Kaldenhoff, R. The Arabidopsis thaliana Aquaporin AtPIP1;2 is a Physiologically Relevant CO(2) Transport Facilitator. Plant J. 2011, 67, 795–804. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Fischer, M.; Colot, V.; Bossdorf, O. Epigenetic Variation Creates Potential for Evolution of Plant Phenotypic Plasticity. New Phytol. 2013, 197, 314–322. [Google Scholar] [CrossRef]
- He, Y.; Li, Z. Epigenetic Environmental Memories in Plants: Establishment, Maintenance, and Reprogramming. Trends Genet. 2018, 34, 856–866. [Google Scholar] [CrossRef]
- Holmes, G.D.; Hall, N.E.; Gendall, A.R.; Boon, P.I.; James, E.A. Using Transcriptomics to Identify Differential Gene Expression in Response to Salinity among Australian Phragmites Australis Clones. Front. Plant. Sci. 2016, 7, 432. [Google Scholar] [CrossRef]
- Payne, S.H. The Utility of Protein and mRNA Correlation. Trends Biochem. Sci. 2015, 40, 1–3. [Google Scholar] [CrossRef]
- Vogel, C.; Marcotte, E.M. Insights into the Regulation of Protein Abundance from Proteomic and Transcriptomic Analyses. Nat. Rev. Genet. 2012, 13, 227–232. [Google Scholar] [CrossRef]
- Nagaraj, N.; Wisniewski, J.R.; Geiger, T.; Cox, J.; Kircher, M.; Kelso, J.; Paabo, S.; Mann, M. Deep Proteome and Transcriptome Mapping of a Human Cancer Cell Line. Mol. Syst. Biol. 2011, 7, 548. [Google Scholar] [CrossRef] [PubMed]
- Maier, T.; Guell, M.; Serrano, L. Correlation of mRNA and Protein in Complex Biological Samples. FEBS Lett. 2009, 583, 3966–3973. [Google Scholar] [CrossRef] [PubMed]
- Carvajal, M.; Cooke, D.T.; Clarkson, D.T. Response of Wheat Plants to Nutrient Deprivation may Involve the Regulation of Water Uptake. Planta 1996, 199, 372–381. [Google Scholar] [CrossRef]
- Martinez-Ballesta, M.D.C.; Carvajal, M. Mutual Interactions between Aquaporins and Membrane Components. Front. Plant Sci. 2016, 7, 1322. [Google Scholar] [CrossRef] [PubMed]
- Slama, I.; Messedi, D.; Ghnaya, T.; Savoure, A.; Abdelly, C. Effects of Water Deficit on Growth and Proline Metabolism in Sesuvium Portulacastrum. Environ. Exp. Bot. 2006, 56, 231–238. [Google Scholar] [CrossRef]
- Liu, F.; Stützel, H. Biomass Partitioning, Specific Leaf Area, and Water use Efficiency of Vegetable Amaranth (Amaranthus spp.) in Response to Drought Stress. Sci. Hortic. 2004, 102, 15–27. [Google Scholar] [CrossRef]
- Osorio, J.; Osorio, M.L.; Chaves, M.M.; Pereira, J.S. Water Deficits are More Important in Delaying Growth than in Changing Patterns of Carbon Allocation in Eucalyptus Globulus. Tree Physiol. 1998, 18, 363–373. [Google Scholar] [CrossRef]
- Martinez-Ferri, E.; Balaguer, L.; Valladares, F.; Chico, J.M.; Manrique, E. Energy Dissipation in Drought-Avoiding and Drought-Tolerant Tree Species at Midday during the Mediterranean Summer. Tree Physiol. 2000, 20, 131–138. [Google Scholar] [CrossRef]
- De la Mata, R.; Merlo, E.; Zas, R. Among-Population Variation and Plasticity to Drought of Atlantic, Mediterranean, and Interprovenance Hybrid Populations of Maritime Pine. Tree Genet. Genomes 2014, 10, 1191–1203. [Google Scholar] [CrossRef]
- Nybom, H. Comparison of Different Nuclear DNA Markers for Estimating Intraspecific Genetic Diversity in Plants. Mol. Ecol. 2004, 13, 1143–1155. [Google Scholar] [CrossRef] [PubMed]
- Alberto, F.J.; Aitken, S.N.; Alia, R.; Gonzalez-Martinez, S.C.; Hanninen, H.; Kremer, A.; Lefevre, F.; Lenormand, T.; Yeaman, S.; Whetten, R.; et al. Potential for Evolutionary Responses to Climate Change—Evidence from Tree Populations. Glob. Chang. Biol. 2013, 19, 1645–1661. [Google Scholar] [CrossRef] [PubMed]
- Petit, R.J.; Hampe, A. Some Evolutionary Consequences of being a Tree. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 187–214. [Google Scholar] [CrossRef]
- Walsh, B.; Blows, M.W. Abundant Genetic Variation Plus Strong Selection = Multivariate Genetic Constraints: A Geometric View of Adaptation. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 41–59. [Google Scholar] [CrossRef]
- Johnson, T.; Barton, N. Theoretical Models of Selection and Mutation on Quantitative Traits. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2005, 360, 1411–1425. [Google Scholar] [CrossRef]
- Mangel, M.; Stamps, J. Trade-Offs between Growth and Mortality and the Maintenance of Individual Variation in Growth. Evol. Ecol. Res. 2001, 3, 583–593. [Google Scholar]
- Kingsolver, J.G.; Diamond, S.E.; Seiter, S.A.; Higgins, J.K. Direct and Indirect Phenotypic Selection on Developmental Trajectories in Manduca Sexta. Funct. Ecol. 2012, 26, 598–607. [Google Scholar] [CrossRef]
- Siepielski, A.M.; DiBattista, J.D.; Carlson, S.M. It’s about Time: The Temporal Dynamics of Phenotypic Selection in the Wild. Ecol. Lett. 2009, 12, 1261–1276. [Google Scholar] [CrossRef]
- Bell, G. Fluctuating Selection: The Perpetual Renewal of Adaptation in Variable Environments. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 87–97. [Google Scholar] [CrossRef]
- Kolb, T.E.; Fettig, C.; Ayres, M.P.; Bentz, B.J.; Hicke, J.A.; Mathiasen, R.; Stewart, J.E.; Weed, A.S. Observed and Anticipated Impacts of Drought on Forest Insects and Diseases in the United States. For. Ecol. Manag. 2016, 380, 321–334. [Google Scholar] [CrossRef]
- Xu, H.; Kemppainen, M.; El Kayal, W.; Lee, S.H.; Pardo, A.G.; Cooke, J.E.; Zwiazek, J.J. Overexpression of Laccaria Bicolor Aquaporin JQ585595 Alters Root Water Transport Properties in Ectomycorrhizal White Spruce (Picea glauca) Seedlings. New Phytol. 2015, 205, 757–770. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.I.F.; Roitto, M.; Lehto, T.; Zwiazek, J.J.; Calvo-Planco, M.; Repo, T. Waterlogging Under Simulated Late-Winter Conditions had Little Impact on the Physiology and Growth of Norway Spruce Seedlings. Ann. For. Sci. 2013, 70, 781–790. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, Y. Protein Structure and Function Prediction using I-TASSER. Curr. Protoc. Bioinform. 2015, 52, 15. [Google Scholar]
- Deokar, A.A.; Tar’an, B. Genome-Wide Analysis of the Aquaporin Gene Family in Chickpea. Front. Plant Sci. 2016, 7, 1802. [Google Scholar] [CrossRef] [PubMed]
- Merlaen, B.; De Keyser, E.; Van Labeke, M.C. Identification and Substrate Prediction of New Fragaria X Ananassa Aquaporins and Expression in Different Tissues and during Strawberry Fruit Development. Hortic. Res. 2018, 5, 20. [Google Scholar] [CrossRef]
- Jordon-Thaden, I.E.; Chanderbali, A.S.; Gitzendanner, M.A.; Soltis, D.E. Modified CTAB and TRIzol Protocols Improve RNA Extraction from Chemically Complex Embryophyta. Appl. Plant Sci. 2015, 3. [Google Scholar] [CrossRef] [PubMed]
- Scotto-Lavino, E.; Du, G.; Frohman, M.A. 3’ End cDNA Amplification using Classic RACE. Nat. Protoc. 2006, 1, 2742–2745. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aquaporin | ar/R Selectivity Filter | Froger’s Positions (P1–P5) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| H2 | H5 | Loop E1 | Loop E2 | P1 | P2 | P3 | P4 | P5 | |
| PiconPIP2;1 | F | H | T | R | Q | S | A | F | W |
| PiconPIP2;2 | F | H | T | R | Q | S | A | F | W |
| PiconPIP2;3 | F | H | T | R | Q | S | A | F | W |
| PiconPIP1;2 | F | H | T | R | Q | S | A | F | W |
| PiconTIP1;1 | H | I | A | R | T | A | A | Y | W |
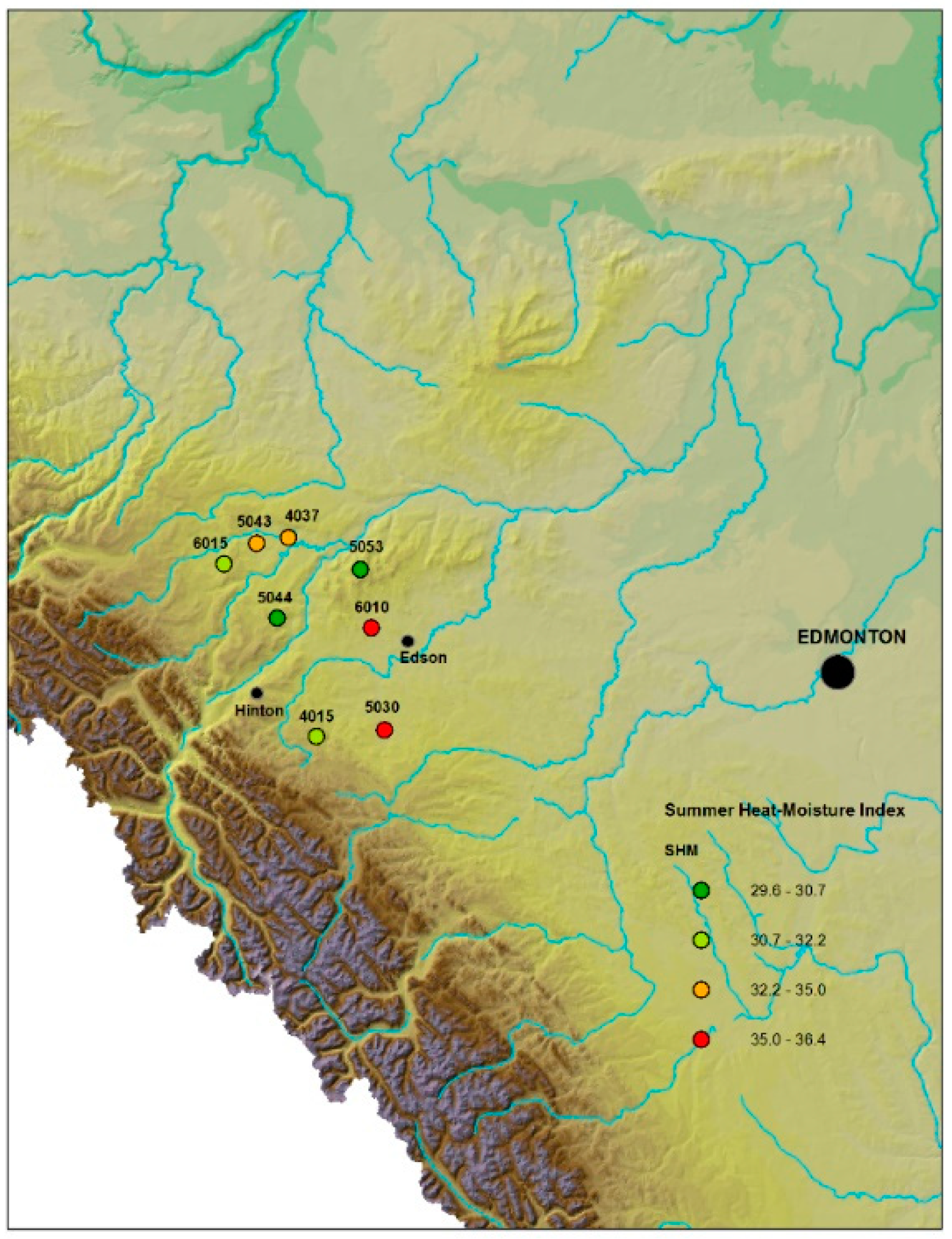
| UI | Latitude °N | Longitude °W | Elevation (m) | SH:M | Drought Severity | Assigned Name |
|---|---|---|---|---|---|---|
| 5044 | 53.690000 | −117.364 | 1317 | 30.7 | Low | FM1L |
| 5053 | 53.901944 | −116.794 | 1330 | 29.6 | Low | FM2L |
| 4015 | 53.208333 | −117.07 | 1313 | 32.1 | Low-mid | FM3ML |
| 6015 | 53.907500 | −117.749 | 1196 | 32.2 | Low-mid | FM4ML |
| 6010 | 53.661667 | −116.709 | 1000 | 35.4 | High | FM7H |
| 5030 | 53.241389 | −116.603 | 1036 | 36.4 | High | FM8H |
| 4037 | 54.025278 | −117.305 | 1026 | 35.0 | Mid-high | FM5MH |
| 5043 | 53.996389 | −117.525 | 1047 | 34.2 | Mid-high | FM6MH |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, S.; Thomas, B.R.; De la Mata, R.; Randall, M.J.; Zhang, W.; Zwiazek, J.J. Variation in Aquaporin and Physiological Responses Among Pinus contorta Families Under Different Moisture Conditions. Plants 2019, 8, 13. https://doi.org/10.3390/plants8010013
Khan S, Thomas BR, De la Mata R, Randall MJ, Zhang W, Zwiazek JJ. Variation in Aquaporin and Physiological Responses Among Pinus contorta Families Under Different Moisture Conditions. Plants. 2019; 8(1):13. https://doi.org/10.3390/plants8010013
Chicago/Turabian StyleKhan, Shanjida, Barb R. Thomas, Raul De la Mata, Morgan J. Randall, Wenqing Zhang, and Janusz J. Zwiazek. 2019. "Variation in Aquaporin and Physiological Responses Among Pinus contorta Families Under Different Moisture Conditions" Plants 8, no. 1: 13. https://doi.org/10.3390/plants8010013