High Constitutive Overexpression of Glycosyl Hydrolase Family 17 Delays Floral Transition by Enhancing FLC Expression in Transgenic Arabidopsis



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. VvGHF17 Expression Is Negatively Related to the Length of Grape Inflorescence
2.2. VvGHF17 Induce Delays of Floral Transition
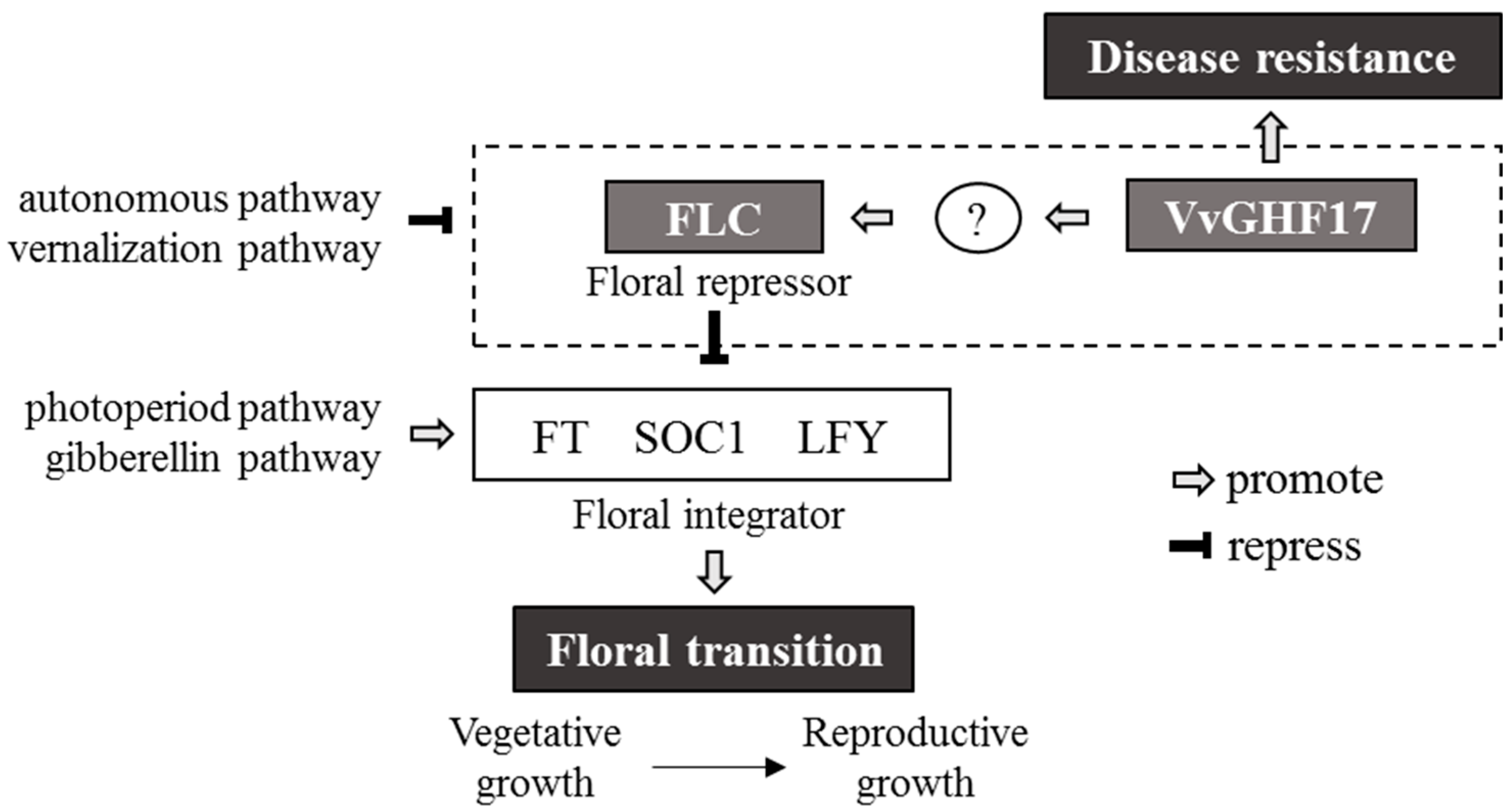
2.3. VvGHF17 Upregulates FLOWERING LOCUS C Expression
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Phenotypic Analysis in Arabidopsis Plants
4.3. Isolation of Total RNA
4.4. Real-Time RT-PCR
4.5. Simple Linear Regression Analysis
4.6. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Stintzi, A.; Heitz, T.; Prasad, V.; Wiedemann-Merdinoglu, S.; Kauffmann, S.; Geoffroy, P.; Legrand, M.; Fritig, B. Plant ‘pathogenesis-related’ proteins and their role in defense against pathogens. Biochimie 1993, 75, 687–706. [Google Scholar] [CrossRef]
- Van Loon, L.C.; Rep, M.; Pieterse, C.M.J. Significance of inducible defense-related proteins in infected plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.J. Fungal cell wall chitinases and glucanases. Microbiology 2004, 150, 2029–2035. [Google Scholar] [CrossRef] [PubMed]
- Simmons, C.R. The physiology and molecular biology of plant 1,3-β-d-glucanases and 1,3;1,4-β-d-glucanases. Crit. Rev. Plant Sci. 1994, 13, 325–387. [Google Scholar] [CrossRef]
- Fujimori, N.; Enoki, S.; Suzuki, A.; Naznin, H.A.; Shimizu, M.; Suzuki, S. Grape apoplasmic β-1,3-glucanase confers fungal disease resistance in Arabidopsis. Sci. Hortic. 2016, 200, 105–110. [Google Scholar] [CrossRef]
- Fulcher, R.G.; McCully, M.E.; Setterfield, G.; Sutherland, J. β-1,3-glucans may be associated with cell plate formation during cytokinesis. Can. J. Bot. 1976, 54, 539–542. [Google Scholar] [CrossRef]
- Masuda, Y.; Wada, S. Effect of beta-3-glucanase on elongation growth of oat coleoptile. Bot. Mag. 1967, 80, 100–102. [Google Scholar] [CrossRef]
- Akiyama, T.; Pillai, M.A.; Sentoku, N. Cloning, characterization and expression of OsGLN2, a rice endo-1,3-β-glucanase gene regulated developmentally in flowers and hormonally in germinating seeds. Planta 2004, 220, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Kauffmann, S.; Legrand, M.; Geoffroy, P.; Fritig, B. Biological function of ‘pathogenesis-related’ proteins: Four PR proteins of tobacco have 1,3-β-glucanase activity. EMBO J. 1987, 6, 3209–3212. [Google Scholar] [PubMed]
- Meikle, P.J.; Bonig, I.; Hoogenraad, N.J.; Clarke, A.E.; Stone, B.A. The location of (1–3)-β-glucans in the walls of pollen tubes of Nicotiana alata using a (1–3)-β-glucan-specific monoclonal antibody. Planta 1991, 185, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ori, N.; Sessa, G.; Lotan, T.; Himmelhoch, S.; Fluhr, R. A major stylar matrix polypeptide (sp41) is a member of the pathogenesis-related proteins superclass. EMBO J. 1990, 9, 3429–3436. [Google Scholar] [PubMed]
- Deytieux, C.; Geny, L.; Lapailerie, D.; Claverol, S.; Bonneu, M.; Donèche, B. Proteome analysis of grape skins during ripening. J. Exp. Bot. 2007, 58, 1851–1862. [Google Scholar] [CrossRef] [PubMed]
- Hinton, D.M.; Pressey, R. Glucanases in fruits and vegetables. J. Am. Soc. Hortic. Sci. 1980, 9, 499–502. [Google Scholar]
- Michaels, S.D.; Amasino, R.M. FLOWERING LOCUS C encodes a novel MADS domain protein that acts as a repressor of flowering. Plant Cell 1999, 11, 949–956. [Google Scholar] [CrossRef] [PubMed]
- Koornneef, M.; Hanhart, C.J.; van der Veen, J.H. A genetic and physiological analysis of late flowering mutants in Arabidopsis thaliana. Mol. Gen. Genet. 1991, 229, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Parcy, F. Flowering: A time for integration. Int. J. Dev. Biol. 2005, 49, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Simpson, G.G.; Dean, C. Arabidopsis, the Rosetta stone of flowering time? Science 2002, 296, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Wen, J.; Zimmer, E.A. Expression patterns of AP1, FUL, FT and LEAFY orthologs in Vitaceae support the homology of tendrils and inflorescences throughout the grape family. J. Syst. Evol. 2015, 53, 469–476. [Google Scholar] [CrossRef]
- Wojtasik, W.; Kulma, A.; Dymińska, L.; Hanuza, J.; Żebrowski, J.; Szopa, J. Fibres from flax overproducing β-1,3-glucanase show increased accumulation of pectin and phenolics and thus higher antioxidant capacity. BMC Biotechnol. 2013, 13, 10. [Google Scholar] [CrossRef] [PubMed]



© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Enoki, S.; Fujimori, N.; Yamaguchi, C.; Hattori, T.; Suzuki, S. High Constitutive Overexpression of Glycosyl Hydrolase Family 17 Delays Floral Transition by Enhancing FLC Expression in Transgenic Arabidopsis. Plants 2017, 6, 31. https://doi.org/10.3390/plants6030031
Enoki S, Fujimori N, Yamaguchi C, Hattori T, Suzuki S. High Constitutive Overexpression of Glycosyl Hydrolase Family 17 Delays Floral Transition by Enhancing FLC Expression in Transgenic Arabidopsis. Plants. 2017; 6(3):31. https://doi.org/10.3390/plants6030031
Chicago/Turabian StyleEnoki, Shinichi, Nozomi Fujimori, Chiho Yamaguchi, Tomoki Hattori, and Shunji Suzuki. 2017. "High Constitutive Overexpression of Glycosyl Hydrolase Family 17 Delays Floral Transition by Enhancing FLC Expression in Transgenic Arabidopsis" Plants 6, no. 3: 31. https://doi.org/10.3390/plants6030031