
Two New Nepenthes Species from the Philippines and an Emended Description of Nepenthes ramos

Abstract
:1. Introduction
2. Results
2.1. A New Pitcher Plant Species from Luzon
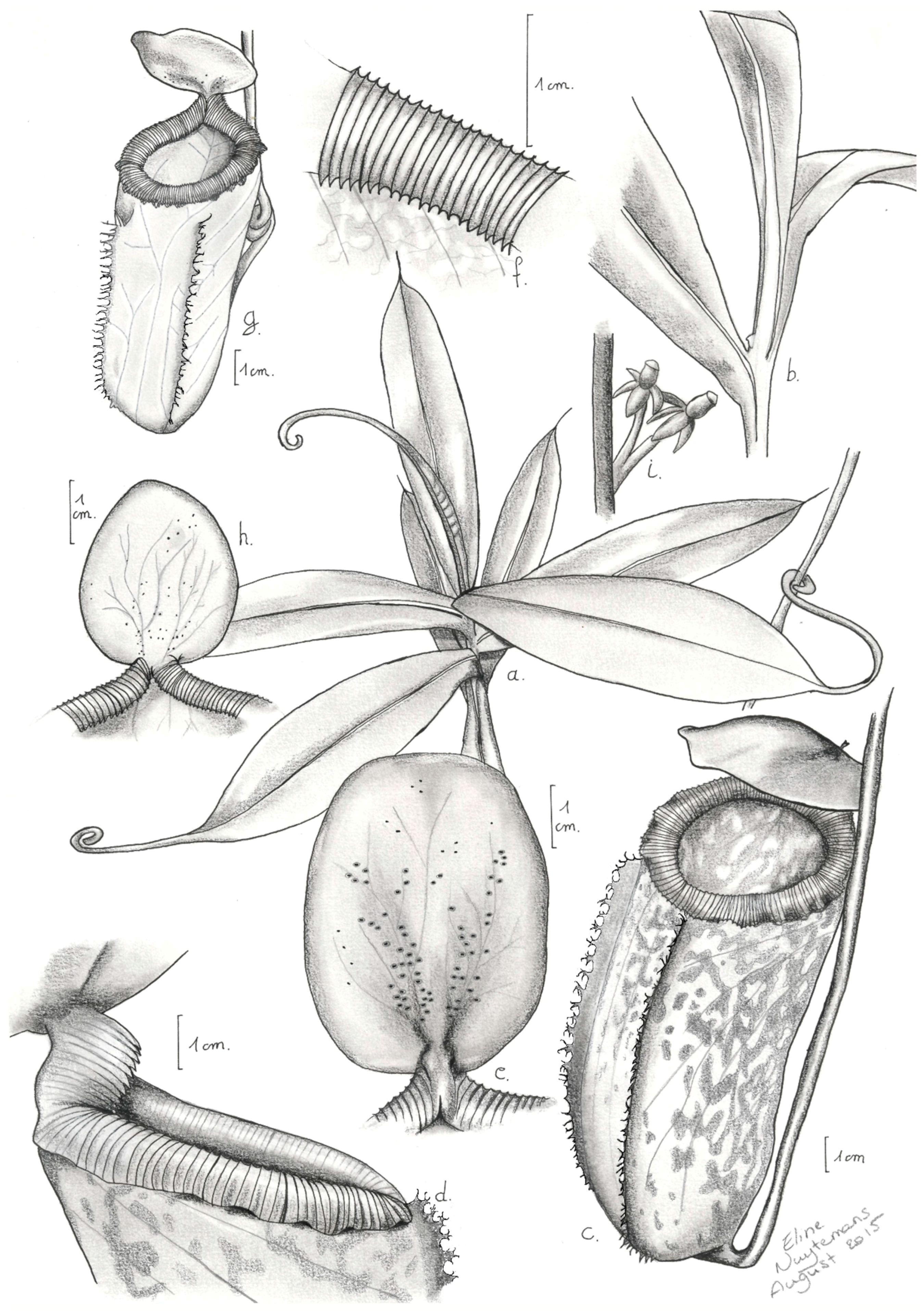
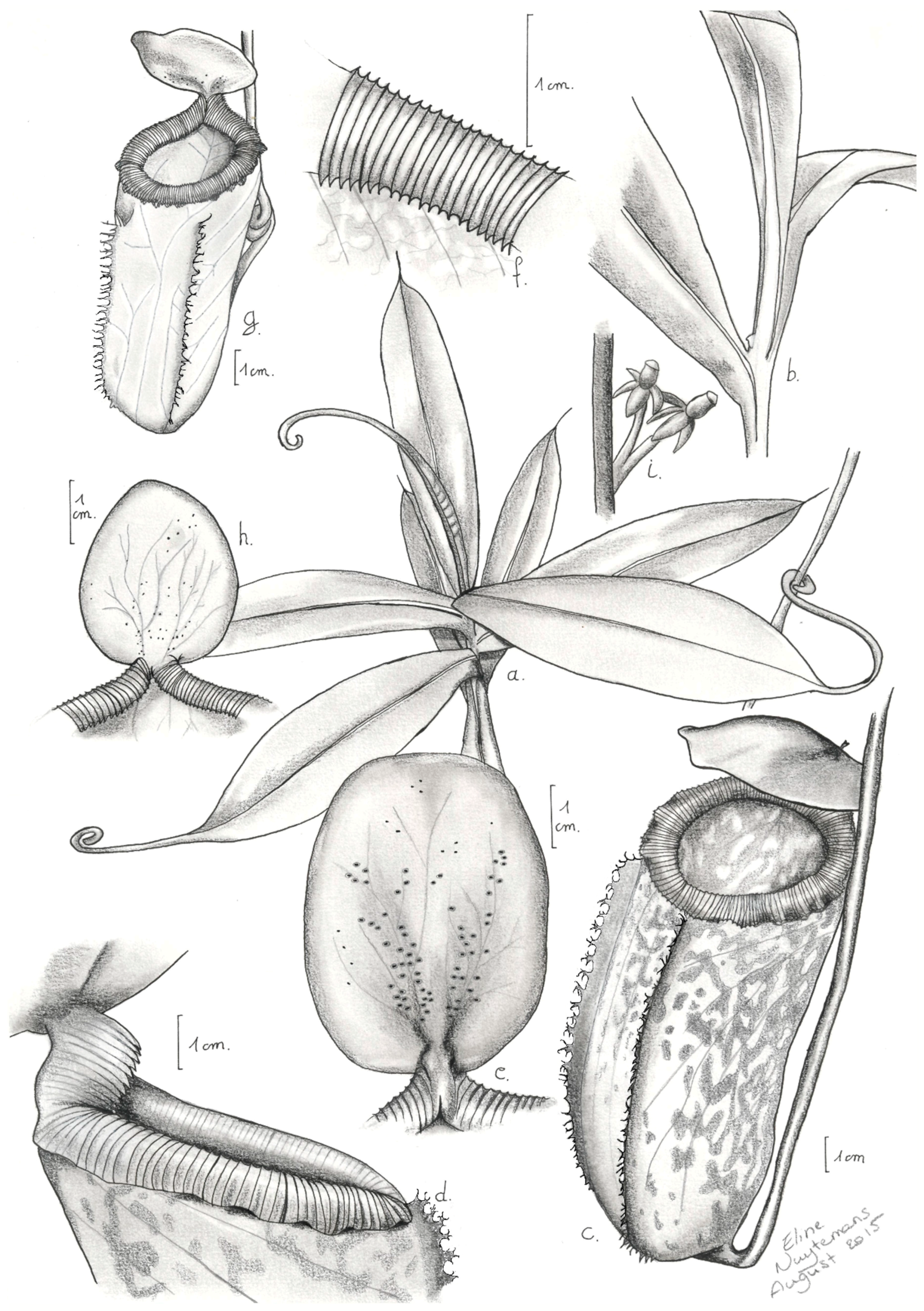
2.1.1. Taxonomic Description of Nepenthes aenigma
- Nepenthes aenigma Nuytemans, W. Suarez, Calaramo, sp. nov.
- Type:
- Diagnosis:
- Description:
- Etymology:
2.1.2. Distribution and Ecology
2.1.3. Conservation Notes
2.2. Another Endemic Species of Pitcher Plant from Mount Hamiguitan (Mindanao Island)
2.2.1. Taxonomic Description of Nepenthes justinae
- Nepenthes justinae Gronem., Wistuba, Mey, V.B.Amoroso, sp. nov.
- Type:
- Philippines, Mindanao Island, Mount Hamiguitan, 17.08.2004, V.B. Amoroso, holotype
- CMUH00003606 (CMUH).
- Other material examined:
- CMUH00003607 (CMUH) (17.08.2004, V. B. Amoroso) (Paratype)
- ULM-22538, ULM-22539 (ULM) (18.2.2015, T. Gronemeyer, from a cultivated plant grown from seed).
- Diagnosis:
- Description:
- Etymology:
- Similarities to other species:
2.2.2. Distribution and Ecology
2.2.3. Conservation Notes
2.3. An Emended Description of Nepenthes ramos Based on Field Data
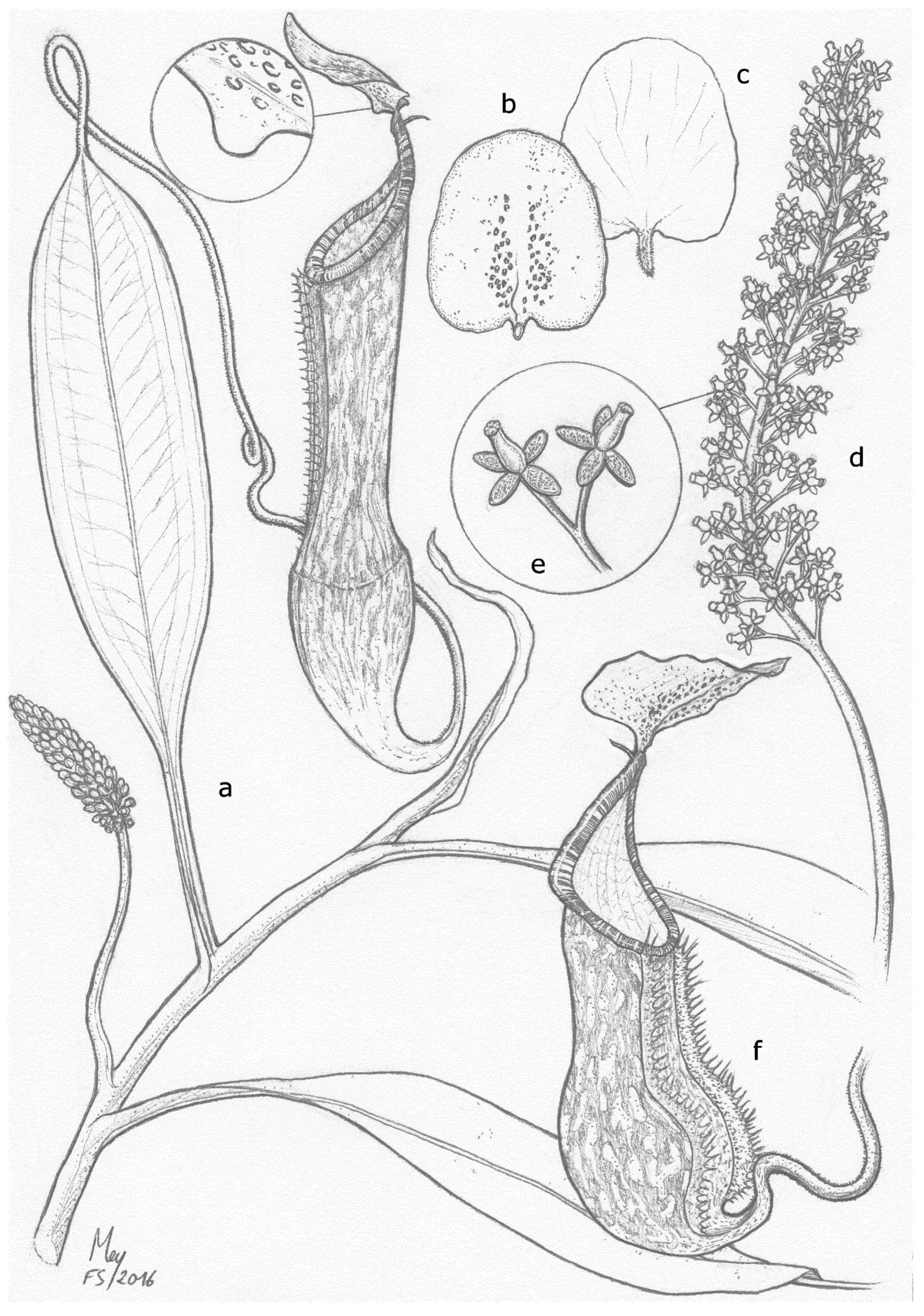
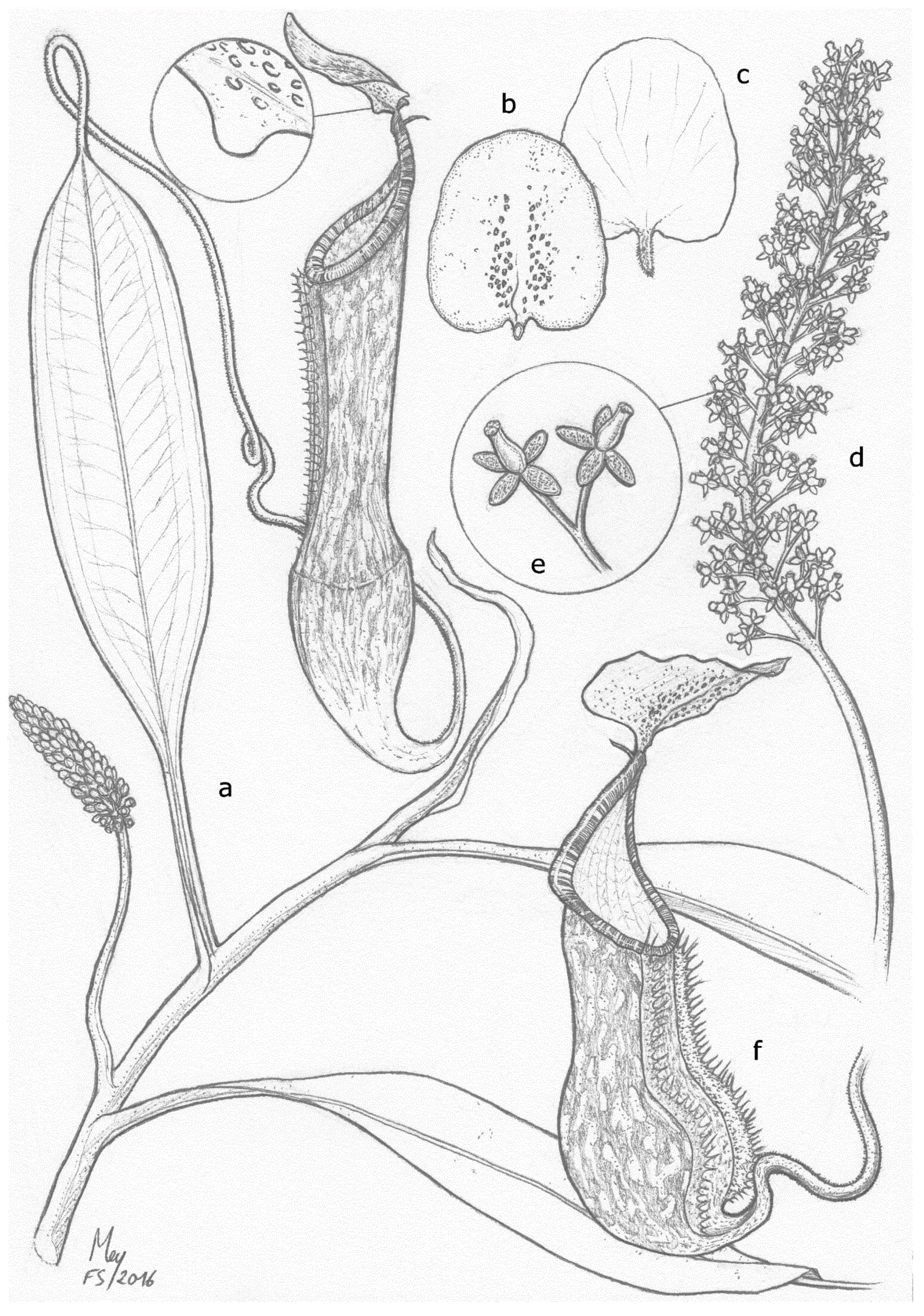
2.3.1. Emended Taxonomic Description of N. ramos and Synonymization of N. kurata
- Nepenthes ramos Jebb and Cheek
- Nepenthes kurata Jebb and Cheek, European Journal of Taxonomy (2013), 69: 6 syn. nov.
- Holotype: Philippines, Mindanao Island, April 1919, Ramos and Palacios s.n., K34500 (K)
- Material examined:
- Philippines, Camiguin Island, Mount Hibok-Hibok, 23.9.2014, T. Gronemeyer, CMUH00009782 (CMUH)
- Philippines, Mindanao Island, Misamis Occidental province, Mt. Malindang, North Peak, Brgy.
- Lake Duminagat, 28.12.2001, Guiller Opiso and Subanen, CMUH13153 (CMUH)
- Philippines, Mindanao, Surigao del Norte Province, Mt. Masay, 12.5.2015, T. Gronemeyer s.n.,
- ULM-22541 (ULM) (from a cultivated plant grown from seed)
- Material not examined:
- Philippines, Mindanao Island, April 1919, Ramos and Palacios, K34500 (K; type)
- Philippines, Mindanao Island, Mount Malindang, May 1906, Mearns and Hutchinson 4632 (K)
- At the time of writing, these specimens were not made accessible upon request.
2.3.2. Ecology and Distribution
2.3.3. Conservation Notes
3. Discussion
4. Material and Methods
- Mt. Hibok-Hibok, Camiguin Island (September 2007, August 2011 and September 2014) (TG)
- Mt. Masay, Surigao del Norte Province, Mindanao (September 2007 and July 2008) (TG)
- Pantaron Range, San Fernando area, Bukidnon Province, Mindanao (July 2008) (TG)
- Mt. Malindang, Misamis Occidental Province, Mindanao (December 2001) (VBA)
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Robinson, A.S.; Fleischmann, A.S.; Mcpherson, S.R.; Heinrich, V.B.; Gironella, E.P.; Peña, C.Q. A spectacular new species of Nepenthes L. (Nepenthaceae) pitcher plant from central Palawan, Philippines. Bot. J. Linn. Soc. 2009, 159, 195–202. [Google Scholar] [CrossRef]
- Gronemeyer, T.; Coritico, F.; Wistuba, A.; Marwinski, D.; Gieray, T.; Micheler, M.; Mey, F.S.; Amoroso, V. Four new species of Nepenthes L. (Nepenthaceae) from the central mountains of Mindanao, Philippines. Plants 2014, 3, 284–303. [Google Scholar] [CrossRef]
- Cheek, M.; Jebb, M. Typification and redelimitation of Nepenthes alata with notes on the N. alata group, and N. negros sp. nov. from the Philippines. Nordic J. Bot. 2013, 31, 616–622. [Google Scholar] [CrossRef]
- Cheek, M.; Jebb, M. Nepenthes alzapan (Nepenthaceae), a new species from Luzon, Philippines. Phytotaxa 2013, 100, 57–60. [Google Scholar] [CrossRef]
- Cheek, M.; Jebb, M. Nepenthes ultra (Nepenthaceae.), a new species from Luzon, Philippines. Blumea 2013, 58, 241–244. [Google Scholar] [CrossRef]
- Cheek, M.; Jebb, M. Identification and typification of Nepenthes blancoi, with N. abalata sp. nov. from the western Visayas, Philippines. Nordic J. Bot. 2013, 31, 151–156. [Google Scholar] [CrossRef]
- Cheek, M.; Tandang, D.M.; Pelser, P.B. Nepenthes barcelonae (Nepenthaceae), a new species from Luzon, Philippines. Phytotaxa 2015, 222, 145–150. [Google Scholar] [CrossRef]
- Cheek, M.; Jebb, M. Nepenthes ramos (Nepenthaceae), a new species from Mindanao, Philippines. Willdenowia 2013, 43, 107–111. [Google Scholar] [CrossRef]
- Cheek, M.; Jebb, M. The Nepenthes micramphora (Nepenthaceae) group, with two new species from Mindanao, Philippines. Phytotaxa 2013, 151, 25–34. [Google Scholar] [CrossRef]
- Cheek, M.; Jebb, M. Recircumscription of the Nepenthes alata group (Caryophyllales: Nepenthaceae), in the Philippines, with four new species. Eur. J. Taxon. 2013, 69, 1–23. [Google Scholar] [CrossRef]
- McPherson, S. New Nepenthes Vol. 1, 1st ed.; Redfern Natural History, GB: Poole, UK, 2012; pp. 457–459. [Google Scholar]
- McPherson, S. Pitcher Plants of the Old World, 1st ed.; Redfern Natural History, GB: Poole, UK, 2009. [Google Scholar]
- IUCN. The IUCN Red List of Threatened Species. Version 2013.2. Available online: http://www.iucnredlist.org (accessed on 21 November 2013).
- Kurata, S. Two new species of Nepenthes from Sumatra (Indonesia) and Mindanao (Philippines). J. Insectivorous Plant. Soc. 2001, 52, 30–34. [Google Scholar]
- Co’s Digital Flora of the Philippines. Available online: https://www.facebook.com/groups/philippineplants/ (accessed on 28 March 2013).




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nepenthes ventricosa | Nepenthes burkei | Nepenthes aenigma | |
|---|---|---|---|
| Growing habit and habitat | Terrestrial, epiphytic or lithophytic. In bright to shady conditions. | Terrestrial or epiphytic. In bright to shady conditions. | Terrestrial in deep shade. |
| Lamina | Long and narrow, length-breadth ratio ±6.4:1 | Long and narrow, length-breadth ratio ±6:1 | Shorter and broader, length-breadth ratio ±3.5:1 |
| Lower pitchers | Absent on mature plants | Absent on mature plants | Absent on mature plants |
| Comparison intermediate and upper pitchers | Monomorphic | Monomorphic | Strongly dimorphic |
| Pitcher shape | Strongly narrowed in the middle, forming a distinct waist. | Slightly narrowed in the middle or cylindrical. | Cylindrical with widely infundibular base in upper pitcher |
| Pitcher opening | Horizontal to slightly oblique | Strongly oblique | Intermediate between N. ventricosa and N. burkei |
| Wings | Reduced to ridges or not apparent | Reduced to ridges or not apparent | Prominent on all pitchers |
| Peristome | Up to 2 cm wide with teeth up to 1 mm long | Up to 3 cm wide with teeth up to 2 mm long | Less than 1 cm wide with teeth less than 1 mm long |
| Inflorescence | 1-flowered partial peduncles | 1-flowered partial peduncles | 2-flowered partial peduncles |
| Nepenthes justinae | Nepenthes mindanaoensis | |
|---|---|---|
| Lower pitchers | Bulbous in the lower ⅔, slightly infundibulate in the upper ⅓. | Obovate or subglobose in the lower ¼, otherwise cylindrical |
| Upper pitchers | Strongly dimorphic | Overall shape similar than the lower pitchers |
| Pitcher opening | Strongly oblique (lower pitcher) | Slightly oblique |
| Nectar glands | Monomorphic | Dimorphic |
| Appendage | Present in upper pitchers | Absent |
| Nepenthes ramos emend. | Nepenthes kurata [10] | Nepenthes mindanaoensis [14] | |
|---|---|---|---|
| Upper pitchers | Mostly equally wide at base and apex, sometimes slightly dilated towards the apex. Constricted in the middle to various extent. | Slightly dilated towards the apex. Constricted in the middle. | Obovate or subglobose in the lower ¼, otherwise cylindrical. |
| Fringed wings | Present in the upper ⅔ to ¼ of the trap or reduced to ribs. | Present in the upper ⅓ to ¼ of the trap. | Present on the whole length of the pitcher. |
| Lid | Smaller, as large as or larger than the pitcher opening. Ovate to orbicular, sometimes domed. | Smaller than the pitcher opening. Ovate. | About as large as the pitcher opening. Orbicular. |
| Nectar glands | Dimorphic | Dimorphic | Dimorphic |
| Appendage | Present (2 mm) or reduced to keel. | Modestly developed, 1–2 mm | Absent |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gronemeyer, T.; Suarez, W.; Nuytemans, H.; Calaramo, M.; Wistuba, A.; Mey, F.S.; Amoroso, V.B. Two New Nepenthes Species from the Philippines and an Emended Description of Nepenthes ramos. Plants 2016, 5, 23. https://doi.org/10.3390/plants5020023
Gronemeyer T, Suarez W, Nuytemans H, Calaramo M, Wistuba A, Mey FS, Amoroso VB. Two New Nepenthes Species from the Philippines and an Emended Description of Nepenthes ramos. Plants. 2016; 5(2):23. https://doi.org/10.3390/plants5020023
Chicago/Turabian StyleGronemeyer, Thomas, Wally Suarez, Herman Nuytemans, Michael Calaramo, Andreas Wistuba, François S. Mey, and Victor B. Amoroso. 2016. "Two New Nepenthes Species from the Philippines and an Emended Description of Nepenthes ramos" Plants 5, no. 2: 23. https://doi.org/10.3390/plants5020023
APA StyleGronemeyer, T., Suarez, W., Nuytemans, H., Calaramo, M., Wistuba, A., Mey, F. S., & Amoroso, V. B. (2016). Two New Nepenthes Species from the Philippines and an Emended Description of Nepenthes ramos. Plants, 5(2), 23. https://doi.org/10.3390/plants5020023