Growth Media Induces Variation in Cell Wall Associated Gene Expression in Arabidopsis thaliana Pollen Tube

Abstract
:1. Introduction
2. Results and Discussion
2.1. Quantitative qPCR Analysis of Selected Genes
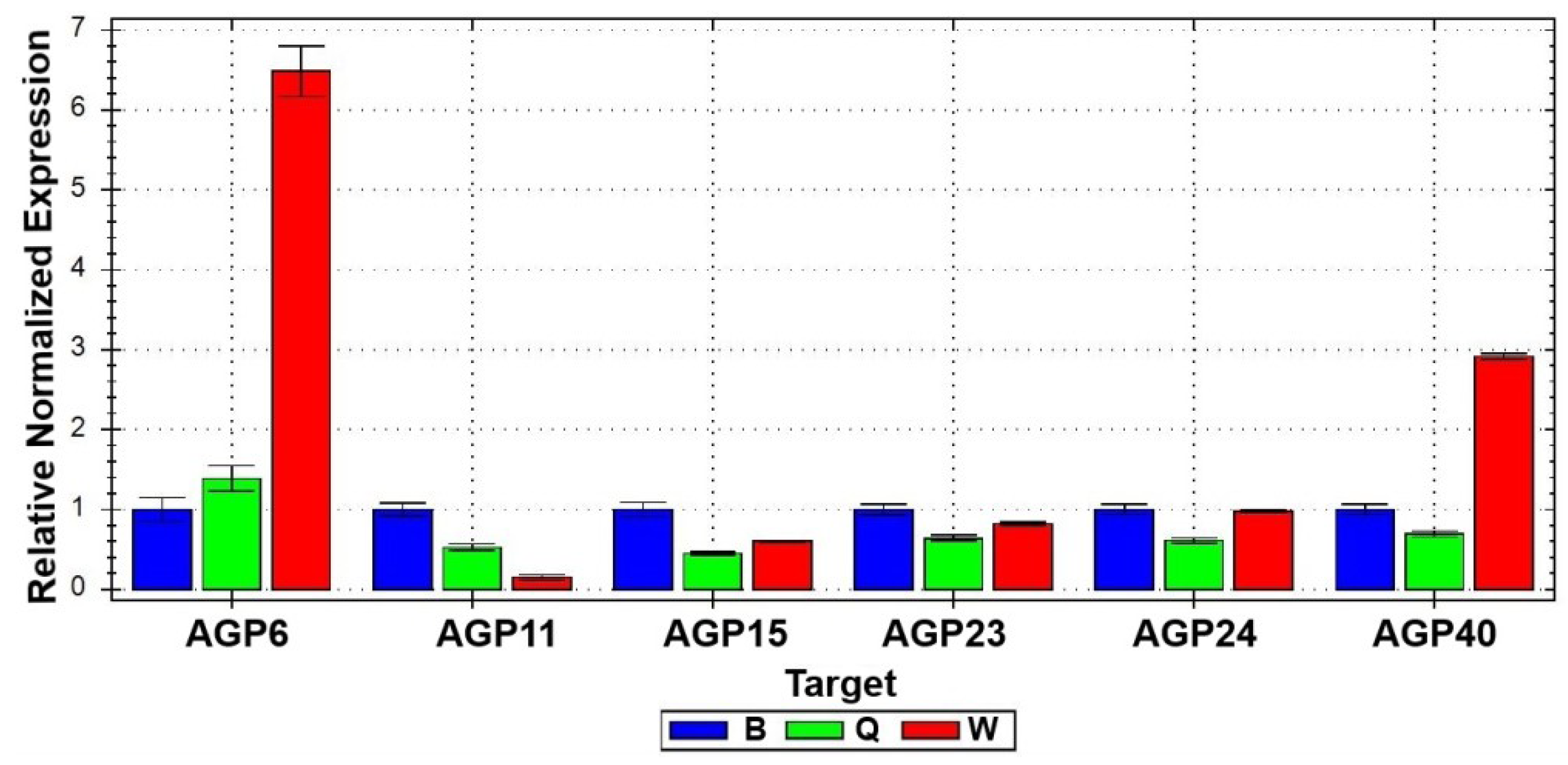
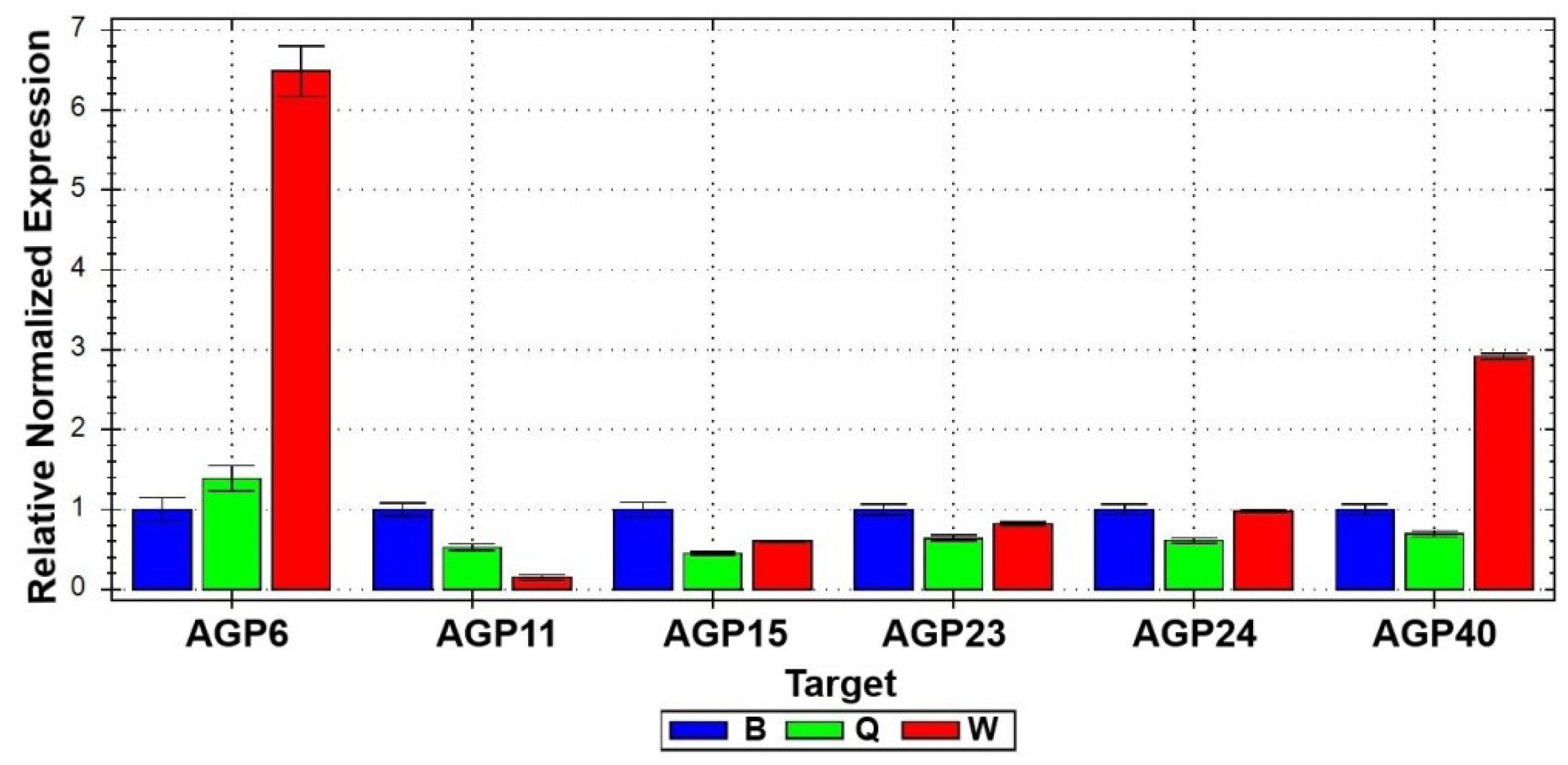
2.1.1. Arabinogalactan Protein Genes
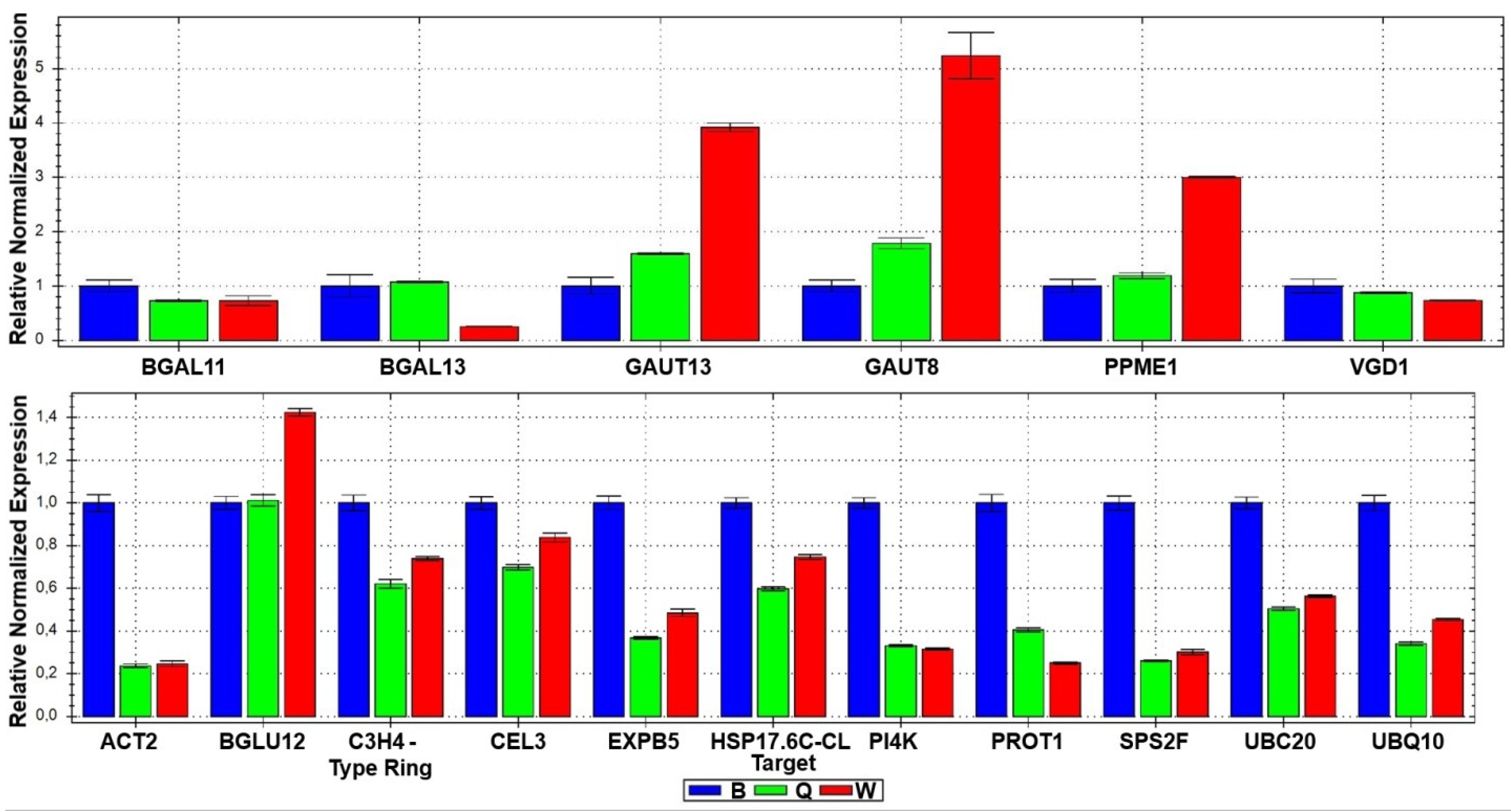
2.1.2. Cell Wall Related Genes

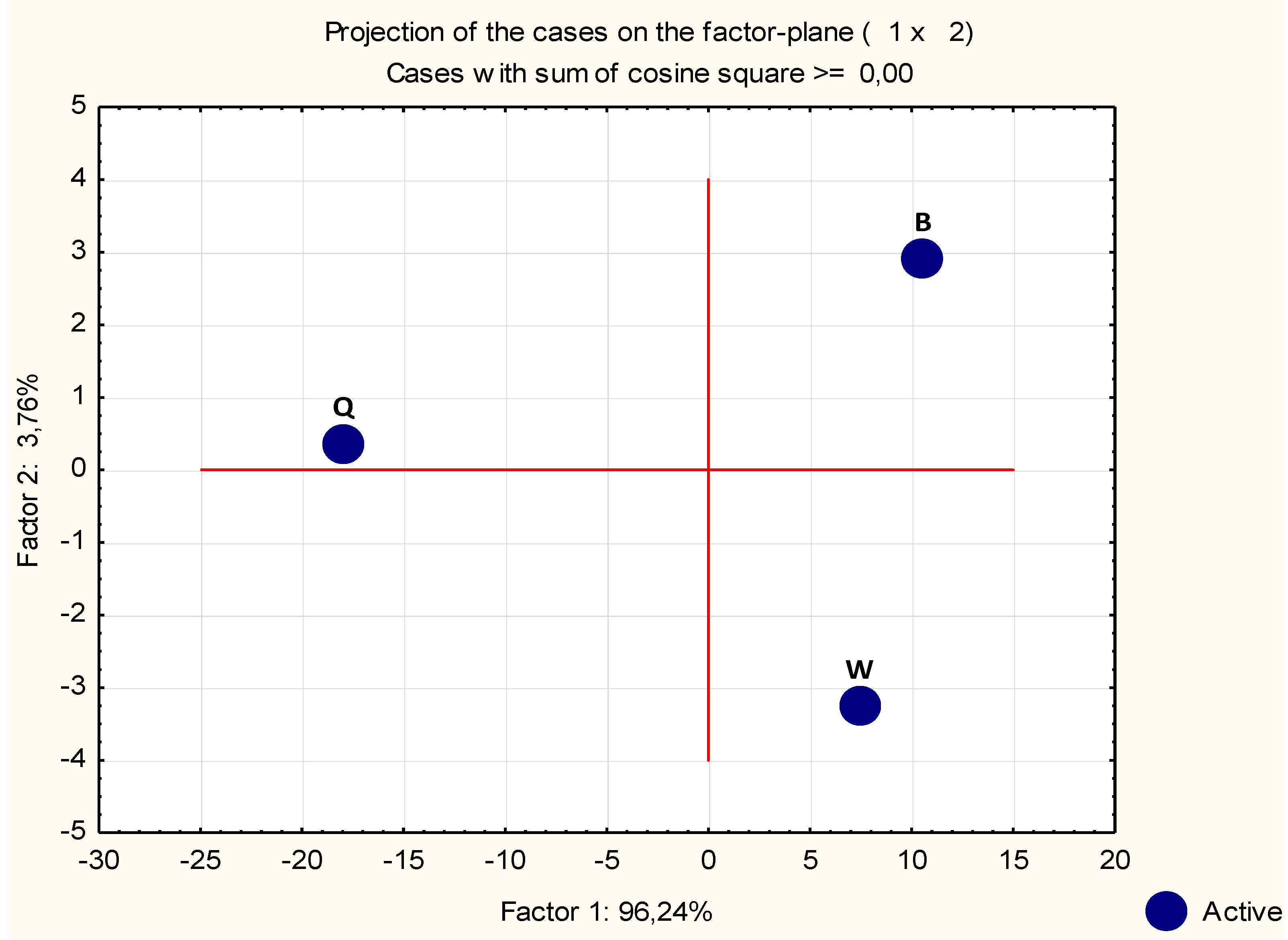
2.1.3. Statistical Analysis of Microarray Data

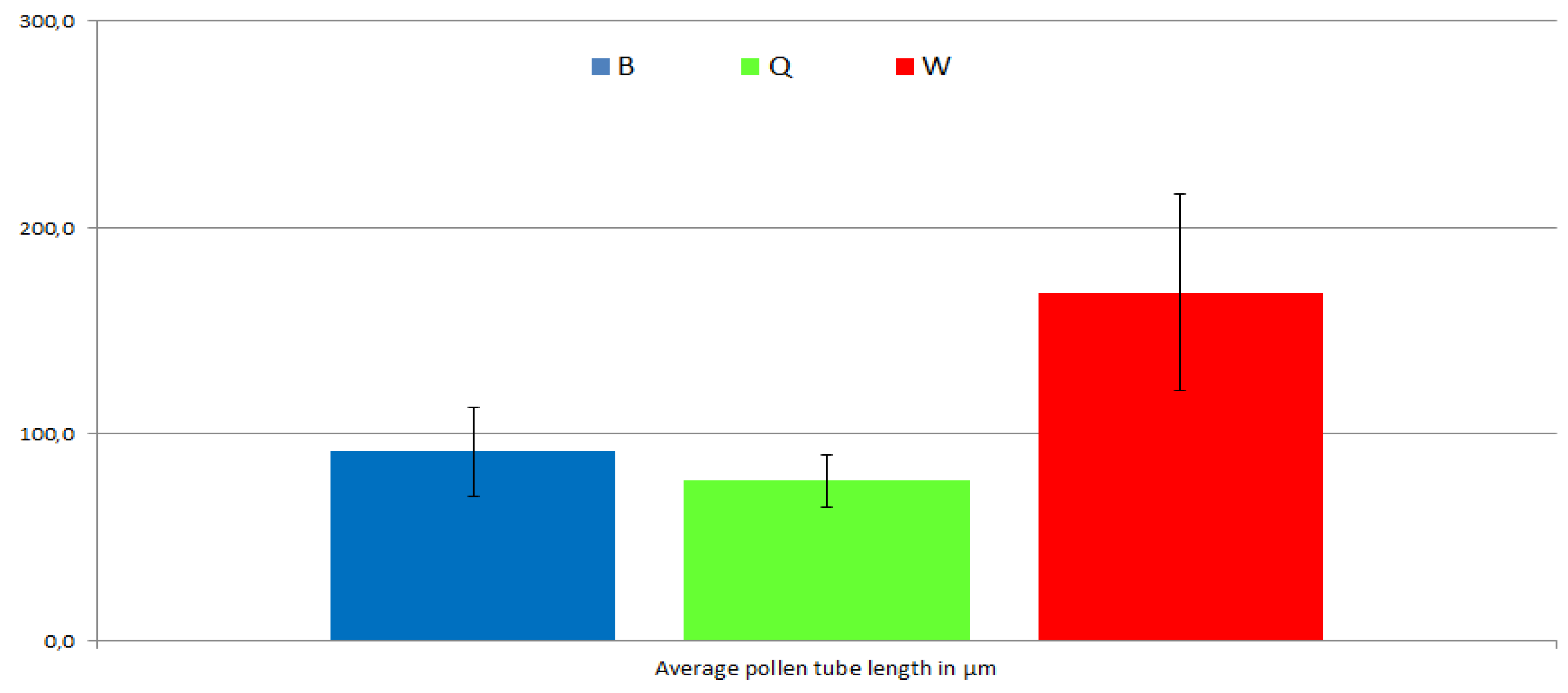
2.1.4. Pollen Tube Germination Rate and Length

3. Experimental Section
3.1. Plant Material and Growth Conditions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Medium W [8] | Medium Q [13] | Medium B [14] | |
|---|---|---|---|
| Sucrose | 19.8% | 18.0% | 10.0% |
| Lactalbumin hydrolysate | 0.05% | - | - |
| Myo-inositol | 10 mM | - | - |
| MES ¶ | 5 mM | - | - |
| CaCl2 | 5 mM | 2 mM | 5 mM |
| H3BO3 | 1.5 mM | 1.5 mM | 1.5 mM |
| KCl | 1 mM | - | 5 mM |
| MgSO4 | 0.8 mM | 1 mM | 1 mM |
| pH | 5.8 | 7.0 | 7.4 |
3.2. RNA Extraction and Real-Time PCR
3.3. Pollen Tube Microarray Data Analysis
3.4. Pollen Tube Germination Rates and Length Measurement
4. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Conflict of Interest
References
- Wang, Y.F.; Fan, L.M.; Zhang, W.Z.; Zhang, W.; Wu, W.H. Ca2+-permeable channels in the plasma membrane of Arabidopsis pollen are regulated by actin microfilaments. Plant Physiol. 2004, 136, 3892–3904. [Google Scholar] [CrossRef]
- Cheung, A.Y.; Wu, H.M. Structural and functional compartmentalization in pollen tubes. J. Exp. Bot. 2007, 58, 75–82. [Google Scholar] [CrossRef]
- Coimbra, S.; Costa, M.; Mendes, M.A.; Pereira, A.M.; Pinto, J.; Pereira, L.G. Early germination of Arabidopsis pollen in a double null mutant for the arabinogalactan protein genes AGP6 and AGP11. Sex. Plant Reprod. 2010, 23, 199–205. [Google Scholar] [CrossRef]
- Fan, L.M.; Wang, Y.F.; Wang, H.; Wu, W.H. In vitro Arabidopsis pollen germination and characterization of the inward potassium currents in Arabidopsis pollen grain protoplasts. J. Exp. Bot. 2001, 52, 1603–1614. [Google Scholar] [CrossRef]
- Honys, D.; Twell, D. Transcriptome analysis of haploid male gametophyte development in Arabidopsis. Genome Biol. 2004, 5, R85. [Google Scholar] [CrossRef] [Green Version]
- Pina, C.; Pinto, F.; Feijó, J.A.; Becker, J.D. Gene family analysis of the Arabidopsis pollen transcriptome reveals biological implications for cell growth, division control, and gene expression regulation. Plant Physiol. 2005, 138, 744–756. [Google Scholar] [CrossRef]
- Becker, J.D.; Feijó, J.A. How many genes are needed to make a pollen tube? Lessons from transcriptomics. Ann. Bot. 2007, 100, 1117–1123. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, W.Z.; Song, L.F.; Zou, J.J.; Su, Z.; Wu, W.H. Transcriptome analyses show changes in gene expression to accompany pollen germination and tube growth in Arabidopsis. Plant Physiol. 2008, 148, 1201–1211. [Google Scholar] [CrossRef]
- Boavida, L.C.; McCormick, S. Temperature as a determinant factor for increased and reproducible in vitro pollen germination in Arabidopsis thaliana. Plant J. 2007, 52, 570–582. [Google Scholar] [CrossRef]
- Higashiyama, T.; Kuroiwa, H.; Kawano, S.; Kuroiwa, T. Guidance in vitro of the pollen tube to the naked embryo sac of torenia fournieri. Plant Cell 1998, 10, 2019–2032. [Google Scholar]
- Palanivelu, R.; Preuss, D. Distinct short-range ovule signals attract or repel Arabidopsis thaliana pollen tubes in vitro. BMC Plant Biol. 2006, 5, 6–7. [Google Scholar]
- Taylor, L.P.; Hepler, P.K. Pollen germination and tube growth. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1997, 48, 461–491. [Google Scholar] [CrossRef]
- Qin, Y.; Leydon, A.R.; Manziello, A.; Pandey, R.; Mount, D.; Denic, S.; Vasic, B.; Johnson, M.A.; Palanivelu, R. Penetration of the stigma and style elicits a novel transcriptome in pollen tubes, pointing to genes critical for growth in a pistil. PLoS Genet. 2009, 5, e1000621. [Google Scholar] [CrossRef]
- Costa, M.; Nobre, S.; Becker, J.; Masiero, S.; Amorim, M.I.; Pereira, L.G.; Coimbra, S. On hand, putative ligands for arabinogalactan proteins in Arabidopsis pollen development. BMC Plant Biol. 2013, 13, 7. [Google Scholar] [CrossRef]
- Daher, F.B.; Geitmann, A. Actin is involved in pollen tube tropism through redefining the spatial targeting of secretory vesicles. Traffic 2011, 12, 1537–1551. [Google Scholar] [CrossRef]
- Burton, R.A.; Gidley, M.J.; Fincher, G.B. Heterogeneity in the chemistry, structure and function of plant cell walls. Nat. Chem. Biol. 2010, 6, 724–732. [Google Scholar] [CrossRef]
- Coimbra, S.; Almeida, J.; Junqueira, V.; Costa, M.; Pereira, L.G. Arabinogalactan proteins as molecular markers in Arabidopsis thaliana sexual reproduction. J. Exp. Bot. 2007, 58, 4027–4035. [Google Scholar] [CrossRef]
- Coimbra, S.; Costa, M.; Jones, B.; Mendes, M.A.; Pereira, L.G. Pollen grain development is compromised in Arabidopsis agp6 agp11 null mutants. J. Exp. Bot. 2009, 60, 3133–3142. [Google Scholar] [CrossRef]
- Del Campillo, E.; Abdel-Aziz, A.; Crawford, D.; Patterson, S.E. Root cap specific expression of an endo-beta-1,4-D-glucanase (cellulase): A new marker to study root development in Arabidopsis. Plant Mol. Biol. 2004, 56, 309–323. [Google Scholar] [CrossRef]
- Valdivia, E.R.; Stephenson, A.G.; Durachko, D.M.; Cosgrove, D. Class B beta-expansins are needed for pollen separation and stigma penetration. Sex. Plant Reprod. 2009, 22, 141–152. [Google Scholar] [CrossRef]
- Suzuki, T.; Masaoka, K.; Nishi, M.; Nakamura, K.; Ishiguro, S. Identification of kaonashi mutants showing abnormal pollen exine structure in Arabidopsis thaliana. Plant Cell Physiol. 2008, 49, 1465–1477. [Google Scholar] [CrossRef]
- Vicré, M.; Santaella, C.; Blanchet, S.; Gateau, A.; Driouich, A. Root border-like cells of Arabidopsis. Microscopical characterization and role in the interaction with Rhizobacteria. Plant Phys. 2005, 138, 998–1008. [Google Scholar] [CrossRef]
- Sterling, J.D.; Atmodjo, M.A.; Inwood, S.E.; Kolli, V.S.; Quigley, H.F.; Hahn, M.G.; Mohnen, D. Functional identification of an Arabidopsis pectin biosynthetic homogalacturonan galacturonosyltransferase. Proc. Natl. Acad. Sci. USA 2006, 103, 5236–5241. [Google Scholar]
- Caffall, K.H.; Pattathil, S.; Phillips, S.E.; Hahn, M.G.; Mohnen, D. Arabidopsis thaliana T-DNA mutants implicate GAUT genes in the biosynthesis of pectin and xylan in cell walls and seed testa. Mol. Plant 2009, 2, 1000. [Google Scholar] [CrossRef]
- Eckart, N.A. VANGUARD1-At the forefront of pollen tube growth. Plant Cell 2005, 17, 327–329. [Google Scholar] [CrossRef]
- Cosgrove, D.J. Creeping walls, softening fruit, and penetrating pollen tubes: The growing roles of expansions. Proc. Natl. Acad. Sci. USA 1997, 94, 5504–5505. [Google Scholar] [CrossRef]
- Tabouchi, A.; Li, L.-C.; Cosgrove, D.J. Matrix solubilization and cell wall weakening by β-expansin (group-1 allergen) from maize pollen. Plant J. 2011, 68, 546–559. [Google Scholar] [CrossRef]
- Jolliffe, I.T. Principal Component Analysis, 2nd ed.; Springer-Verlag: Berlim, Germany, 1986. [Google Scholar]
- Raychaudhuri, S.; Stuart, J.M.; Altman, R.B. Principal Components Analysis to summarize microarray experiments: Application to sporulation time series. Pac. Symp. Biocomput. 2000, 2000, 455–466. [Google Scholar]
- Smyth, D.R.; Bowman, J.L.; Meyerowitz, E.M. Early flower development in Arabidopsis. Plant Cell 1990, 2, 755–767. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Da Costa, M.L.; Pereira, L.G.; Coimbra, S. Growth Media Induces Variation in Cell Wall Associated Gene Expression in Arabidopsis thaliana Pollen Tube. Plants 2013, 2, 429-440. https://doi.org/10.3390/plants2030429
Da Costa ML, Pereira LG, Coimbra S. Growth Media Induces Variation in Cell Wall Associated Gene Expression in Arabidopsis thaliana Pollen Tube. Plants. 2013; 2(3):429-440. https://doi.org/10.3390/plants2030429
Chicago/Turabian StyleDa Costa, Mário Luís, Luís Gustavo Pereira, and Sílvia Coimbra. 2013. "Growth Media Induces Variation in Cell Wall Associated Gene Expression in Arabidopsis thaliana Pollen Tube" Plants 2, no. 3: 429-440. https://doi.org/10.3390/plants2030429