
Flooding Tolerance of Rice: Regulatory Pathways and Adaptive Mechanisms
by
 ,
,
Jing Wang
1,2,3,4,5,†,
Mingzhen Han
2,3,4,5,6,†,
Yongxiang Huang
1,†,
Junliang Zhao
2,3,4,5,
Chuanguang Liu
2,3,4,5 and
Yamei Ma
2,3,4,5,* 1
College of Coastal Agricultural Sciences, Guangdong Ocean University, Zhanjiang 524088, China
2
Rice Research Institute, Guangdong Academy of Agricultural Sciences, Guangzhou 510640, China
3
Key Laboratory of Genetics and Breeding of High Quality Rice in Southern China (Co-Construction by Ministry and Province), Ministry of Agriculture and Rural Affairs, Guangzhou 510640, China
4
Guangdong Key Laboratory of New Technology in Rice Breeding, Guangzhou 510640, China
5
Guangdong Rice Engineering Laboratory, Guangzhou 510640, China
6
College of Agriculture and Biology, Zhongkai University of Agriculture and Engineering, Guangzhou 510225, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Plants 2024, 13(9), 1178; https://doi.org/10.3390/plants13091178
Submission received: 1 March 2024
/
Revised: 20 April 2024
/
Accepted: 21 April 2024
/
Published: 23 April 2024
(This article belongs to the Special Issue Environmental and Genetic Factors in Field Crop Production and Improvement: Mechanisms and Regulation)
{kind=link}
{kind=link}
Abstract
:Rice is a major food crop for more than half of the world’s population, while its production is seriously threatened by flooding, a common environmental stress worldwide. Flooding leads to oxygen deficiency, which is a major problem for submerged plants. Over the past three decades, significant progress has been made in understanding rice adaptation and molecular regulatory mechanisms in response to flooding. At the seed germination and seedling establishment stages, the CIPK15-SnRK1A-MYBS1 signaling cascade plays a central role in determining rice submergence tolerance. However, from seedlings to mature plants for harvesting, SUB1A- and SK1/SK2-regulated pathways represent two principal and opposite regulatory mechanisms in rice. In addition, phytohormones, especially gibberellins, induce adaptive responses to flooding throughout the rice growth period. This review summarizes the significant adaptive traits observed in flooded rice varieties and updates the molecular genetics and mechanisms of submergence tolerance in rice.
1. Introduction
Flooding is one of the most destructive environmental stresses worldwide [1]. Rice, the primary staple food crop for more than half of the world’s population, is usually cultivated in paddy fields but is also affected by flooding in rain-fed and irrigated lowlands where plants face a high risk of submergence during their life cycle. Flooding can lead to two different stresses: (1) submergence stress, where the plant organ is completely under water, and (2) water-logging stress, where the plant leaf and stem are partially submerged under water [2]. Flooding is considered a compound stress, and its primary effect is restricted oxygen (O2) and carbon dioxide (CO2) availability in plant tissues, owing to the slow diffusion of gases in water [3,4]. The decline in molecular O2 results in the restriction of ATP synthesis and carbohydrate resources, which has major consequences for growth and survival [5].
Rice is the only crop that can successfully germinate and grow under submergence. When germinated underwater, the coleoptile elongates rapidly, whereas the growth of the roots and leaves are blocked. The rapid elongation of the hollow coleoptile acts as a snorkel, extending to the water surface, thereby obtaining more O2 to support seedling establishment. Thus, anaerobic germination (AG) and coleoptile elongation are two major adaptive traits during the early growth stage of rice upon submergence [6,7,8,9]. From seedlings to mature plants for harvesting, submergence seriously compromises the overall growth, development, and yield potential of rice [10]. Based on flooding regimes, rice can be roughly divided into two major ecotypes: rain-fed lowland rice that experiences shallow submergence, and deepwater rice that experiences medium to deep submergence during the monsoon season [11,12,13,14,15]. In these cases, two completely different strategies, quiescence and escape, are employed for optimal growth and survival. For example, certain landraces of lowland rice (e.g., FR13A) show flooding tolerance for up to two weeks by entering into an energy conservation mode, in which growth is temporarily arrested and energy sources are conserved for regrowth after the stress conditions subside. In contrast, deepwater rice exhibits the potential for rapid growth (especially internode elongation), consuming carbohydrate resources, so that the upper leaves rise above the water level to escape submergence [2,12,16].
From the middle of 1990 to now, the genetic basis of submergence tolerance in rice has been gradually discovered, and several major regulators such as Submergence-1 (SUB1) and SNORKEL1/2 (SK1/2) have been cloned and functionally characterized [17,18,19]. Recently, several studies have revealed the molecular regulatory mechanisms underlying rice submergence tolerance during the seed germination and seedling establishment stages. In this review, we highlight the significant adaptive traits observed in flooded rice varieties and update the molecular genetics and the mechanism of submergence tolerance in rice.
2. Tolerance of Rice to Submergence at Seed Germination and Seedling Establishment Stages and Underlying Molecular Regulatory Mechanisms
2.1. Metabolism and Growth Change in Response to Submergence
In traditional rice cultivation, seeds are usually germinated in the nursery, and then, well-established seedlings are transplanted into puddled fields for further growth. Recently, direct seeding, whereby rice seeds are sown directly into the soil, has become an increasingly popular cultivation method, owing to its low cost and convenience [20]. However, flooding is common under direct seeding, particularly in rain-fed areas and fields with uneven surfaces and heavy post-sowing rainfall. Flooding stress leads to unfavorable metabolic stress for plants owing to insufficient oxygen supply. Oxygen deprivation compromises the mitochondrial electron transport chain (mETC), leading to an oxidative burst and the release of sequestered calcium into the cytosol. This surge in oxidative stress activates the mitogen-activated protein kinase (MAPK) pathway, which in turn triggers multiple signaling mechanisms to circumvent the energy crisis [21,22]. Although rice exhibits a certain degree of tolerance to oxygen deficiency for AG and anaerobic seedling development (ASD), the AG/ASD capabilities of most rice varieties are poor.
Numerous studies show that rice mainly employs ‘metabolic adaptation’ and ‘escape’ strategies to ensure seed germination and seedling development under flooded anaerobic conditions [6,9,16]. Upon submergence, starch reserves within the endosperm are hydrolyzed to produce sugars for fermentation-mediated energy production to support seed germination and seedling growth. During this process, alpha amylase enzymes, principally the AMY1 and AMY2 subfamilies, enable the degradation of starch in rice grains [23]. In air, the rice coleoptile is initially white and turns green after one day. Along with starch degradation in the endosperm, energy supply enables primary root development [24]. When germinated underwater, coleoptiles can grow for several days and rapidly elongate to reach the water surface, allowing O2 to diffuse into the endosperm to hydrolyze nutrients and support vigorous seedling establishment [25]. However, flooded anaerobic conditions completely inhibit root growth, which is only reinitiated as coleoptiles approach the water surface. Several important regulators and signaling cascades of rice AG/ASD have been established. CIPK15-mediated energy signaling and phytohormone signaling, including gibberellins (GA), abscisic acid (ABA), and auxin, play essential roles in determining rice seed germination and seedling establishment during submergence (Figure 1).
2.2. CIPK15-Mediated Sugar-Sensing Signaling in AG/AGD Regulation
It is well established that the CIPK15-SnRK1A-MYBS1 signaling cascade plays a critical role in O2 deficiency tolerance during rice seed germination and seedling development stages [26,27]. Submergence can trigger Ca2+ flux, which in turn induces conformational changes in calcineurin B-like proteins (CBLs) to allow for their hydrophobic interactions with other proteins [28]. Previous studies have demonstrated that several CBL calcium sensors (CBL2-6) can interact with CBL protein-interacting protein kinases 14 and 15 (CIPK14 and CIPK15) [29]. Recently, Ye and his co-workers found that variations in the OsCBL10 promoter sequences contribute to this divergence in rice flooding tolerance. Flooding-tolerant rice cultivars containing the OsCBL10 T-type promoter (which exhibits a much lower response to flooding stress than the I-type promoter) have shown lower Ca2+ flow and increased CIPK15 protein accumulation, aAmy3 expression, and total aAmy activity in comparison to those in flooding-intolerant cultivars [30]. The interaction between CBLs and CIPKs activates sucrose non-fermenting 1 (Snf1)-related protein kinase 1 (SnRK1A), a global energy and stress sensor in rice, which results in the transcriptional activation and phosphorylation of MYB SUCROSE 1 (MYBS1) transcription factor. Subsequently, the expression of the starvation-induced α-amylase gene, αAmy3/RAmy3D, is activated by MYBS1 [27]. MYB SUCROSE 2 (MYBS2) competes with MYBS1 to bind and suppress the aAmy promoter during submergence [31].
Several regulators/substrates of SnRK1A have been reported to modulate rice submergence tolerance. Two hypoxia-inducible SnRK1A-INTERACTING NEGATIVE REGU-LATORs (SKIN1/2) can inhibit the expression of MYBS1 and aAmy3, leading to the suppression of starch-to-sugar hydrolysis, thus delaying germination and seedling growth under submergence [32]. TREHALOSE-6-PHOSPHATE PHOSPHATASE 7 (TPP7) is another important regulator of SnRK1A, which promotes AG/ARD [33]. TPP7 is the genetic determinant of a major quantitative trait locus (QTL), qAG-9-2, which catalyzes the conversion of T6P to trehalose. Reduced T6P levels release the inhibition of the CIPK15-SnRK1A-MYBS1 pathway and allow for increased starch mobilization in the form of readily fermentable sugars, thus enhancing coleoptile elongation and embryo germination [33]. Recently, a member of the FLC-like zinc finger (FLZ) protein family, FLZ18, was found to interact with SnRK1A and repress its transcriptional activation to αAmy3. Consistent with this, FLZ18-overexpressing rice plants are more sensitive to submergence than wild-type Nipponbare plants during the seed germination stage [34]. EBP89, a member of the AP2/ERF subfamily, was recently identified as a substrate of SnRK1A that can repress the expression of genes involved in the ROS-scavenging system, for example ascorbate peroxidase 1 (APX1), thus negatively regulating AG/ASD [35].
2.3. Hormone Signaling in AG/AGD Regulation
Plant hormones are central regulators of plant growth, development, and responses to changing environments. Recent studies have revealed the critical roles of GA, ABA, and auxin in rice AG/ASD regulation. GA is a well-known hormone that can promote seed germination and seedling growth, whereas ABA represses these two processes. The balance between GA and ABA signaling determines the final seed germination performance. Recently, the 14-3-3 protein, GF14h, was shown to confer tolerance to rice AG/AGD. GF14h can interact with HOX3 and enhance its transcriptional repression activity toward downstream PYL5, an ABA receptor gene, to attenuate ABA signaling. GF14h can also interact with OsVP1, a transcription activator of ABA signaling, thus precisely controlling ABA response under submergence conditions. Interestingly, GF14h also promotes GA biosynthesis, possibly through the upregulation of GA20ox1 expression, along with other unknown transcription factors. Therefore, GF14h can act as a signal switch to balance ABA signaling and GA biosynthesis, thereby boosting the seeding rate of anaerobic-sensitive varieties under flooded direct-seeded conditions [36]. After germination, rapid elongation of coleoptiles is important for conferring tolerance to submergence. Most recently, He et al. reported that the natural variation in rice coleoptile length subjected to submergence is determined by UGT75A, a glucosyl-transferase-encoding gene that decreases free ABA and jasmone acid (JA) levels by promoting the glycosylation of these two phytohormones under submergence conditions [37]. In addition, they found that UGT75A accelerates coleoptile elongation by inhibiting the interactions between JASMONATE ZIMDOMAIN (JAZ) and ABSCISIC ACID-INSENSITIVE (ABI) proteins. This study further highlights the negative effects of ABA signaling on AG/AGD in rice.
Rapid coleoptile elongation during submergence is mainly attributed to cell elongation. Auxin plays an essential role in this process [38]. MiR393 can shear the mRNA of TRANSPORT INHIBI-TOR RESPONSE 1 (TIR1)/AUXIN SIGNALING F-BOX 2 (AFB2) auxin receptors, which regulate AUXIN RESPONSE FACTOR (ARF) expression [39]. Submergence downregulates the expression of miR393, thus activating the auxin signaling pathway and stimulating coleoptile elongation in rice [40]. Interestingly, a recent report has shown that a certain threshold level of endogenous auxin is required for rice germination and early seedling growth during submergence [41]. Lee and his co-workers showed that a submergence-repressed miR167 is involved in the metabolic regulation of IAA through the miR167a-ARF-GH3 pathway [41]. MiR167-directed ARF mRNA cleavage is accompanied by the downregulation of glycoside hydrolase 3 (GH3) [42]. GH3-8 encodes an IAA-amido synthetase that prevents free IAA accumulation and maintains auxin homeostasis [43]. Reduced miR167 levels or increased GH3-8 levels reduce the endogenous levels of free IAA, thus promoting rice AG/AGD [41]. Recently, OsPIN2, a gene encoding an auxin efflux transporter, was found to negatively regulate hypoxia tolerance by affecting auxin transport and ROS accumulation [44]. These findings indicate that plant hormones are important regulators of AG/AGD in rice.
3. The Tolerance of Rice to Submergence at the Vegetative Stage and the Underlying Molecular Regulatory Mechanisms
3.1. Morphologic Adaptation to Submergence at the Vegetative Stage
In natural fields, from seedlings to mature plants for harvesting, rice is frequently subjected to submergence stress, which seriously compromises its growth, development, and yield. As a wetland plant species, rice possesses several adaptive traits, such as aerenchyma and adventitious roots, to cope with submergence stress.
3.1.1. Aerenchyma Formation
The major adaptive feature of rice in response to submergence is the formation of aerenchyma, which is well-developed in the leaves, sheaths, roots, and internodes [2,10]. Ethylene triggers the formation of aerenchyma in maize and rice [5]. This aerenchyma constitutes gas spaces and becomes interconnected, which allows oxygen to transit from the above-water parts of the plant to the submerged roots. Generally, aerenchyma formation begins at the apical parts of the roots and gradually increases toward the basal parts of the roots [45]. There are two main types of aerenchyma: schizogenous and lysigenous. Lysigenous aerenchyma has been well studied, which can be constitutively formed in roots even under well-drained soil conditions, and its formation can be further induced under waterlogging conditions [46]. In addition, aerenchyma formation is less extensive in non-wetland plant species than in wetland plant species. Consistent with this, non-wetland plants are less tolerant to waterlogging than wetland plants such as rice [45,47]. Hence, the capacity to produce enough aerenchyma is important for increasing the tolerance to waterlogging in dryland crops [48,49].
3.1.2. Adventitious Roots Formation
One of the most prominent outcomes of flooding is the low oxygen availability, which severely inhibits root growth. Limited oxygen availability impedes cell division and elongation, thus inhibiting the elongation of rice roots [50]. Under submerged conditions, rice root systems tend to become shallower and more compact, accompanied by an increase in adventitious roots (ARs) [10]. Adventitious roots reduce the distance between oxygen and the nutrient supply from the shoot. Pre-existing and newly formed ARs emerge upon submergence to replace the primary root system, which becomes easily dysfunctional when flooded under water. This adaptation enables plants to acquire oxygen and nutrients efficiently under flooded conditions. Recently, Lin et al. found that rice cultivar T65 produces only one type of AR, whereas NIL-12 (near-isogenic lines) enables the production of two types of ARs following partial submergence. Further morphological and anatomical traits suggest that AR2 (type 2 ARs, emergence several days later at the nodal region above type 1 ARs) is better adapted to flooding than AR1 (type 1 ARs, emergence from nodes early during submergence) [51].
3.1.3. Shoot Elongation
Under flooded conditions, shoot elongation in rice is observed to be a response to flooding. Depending on the depth and duration of submergence, rice cultivars employ two different approaches to modulate shoot morphology. One of the approaches is the quiescence strategy, in which shoot elongation is arrested so that carbohydrates can be preserved for regrowth after floodwater recedes [4,6,52]. A comprehensive study performed on a group of 903 cultivars from the International Rice Research Institute provided strong evidence supporting the negative correlation between shoot elongation and submergence tolerance [53]. This deliberate reduction in shoot elongation allows these genotypes to effectively allocate resources and increase their probability of survival during the recovery stage [54]. Another approach is the escape strategy, which is characteristic of upland cultivars, in which the shoot/internode elongates rapidly to potentially grow above water [4,6,52]. Deepwater rice cultivars frequently use this approach to optimize survival under submerged conditions during the monsoon season [2].
3.2. Genetic Mechanism of Submergence Tolerance
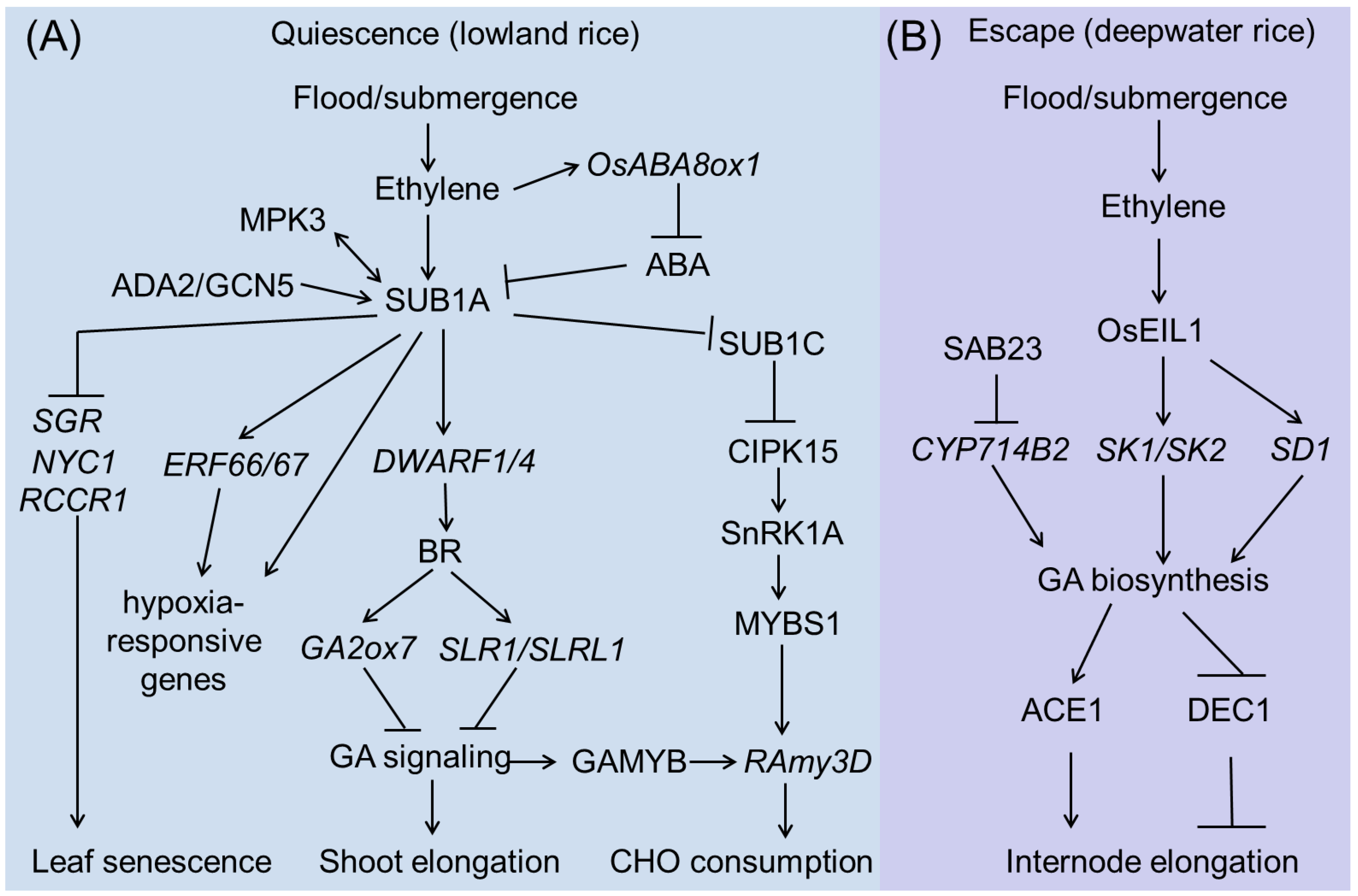
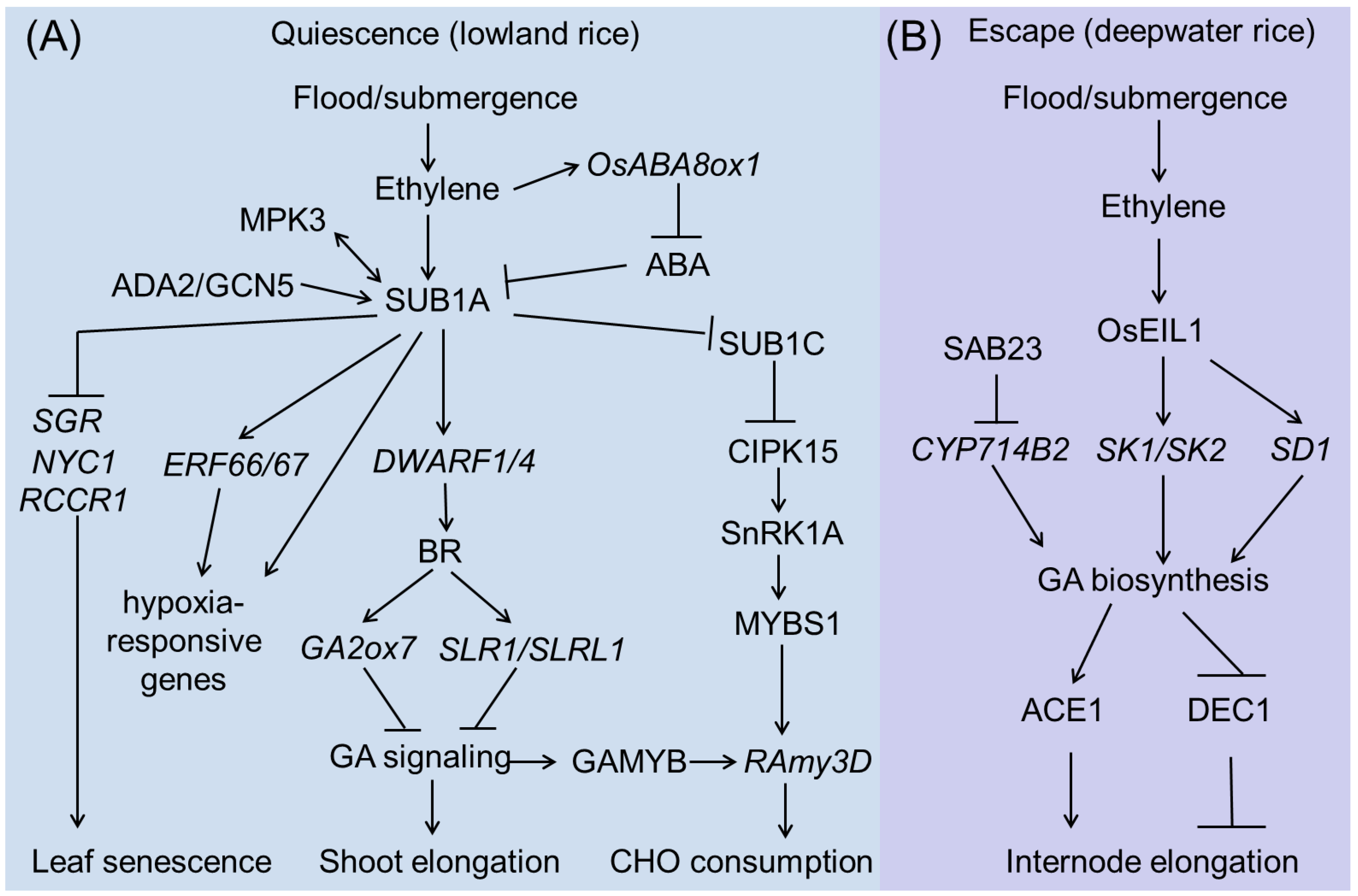
The genetic basis of rice submergence tolerance at the vegetative stage is becoming increasingly clear. An overview picture is shown in Figure 2. ‘Quiescence’ and ‘Escape’ are two major and opposite strategies employed to counteract the effects of submergence by lowland rice and deepwater rice, respectively (Figure 2).
3.2.1. SUB1A Pathway
Genetic mapping studies have associated the submergence-tolerant phenotype with a single QTL on chromosome 9 in FR13A, named SUB1. This QTL accounts for 69% of the phenotypic variation in rice flooding tolerance [55]. Further studies have shown that the SUB1 locus contains three genes, SUB1A, SUB1B, and SUB1C, which encode proteins belonging to the Group VII ethylene-responsive factor (ERFVII) [17,19,56]. Most rice varieties contain SUB1B and SUB1C genes, whereas SUB1A only exists in flooding-tolerant varieties [15,57]. Upon submergence, SUB1A and SUB1C expression is rapidly induced; however, the expression of SUB1B is only slightly responsive [17]. Populations carrying SUB1A have two alleles, SUB1A-1 and SUB1A-2. The only difference between the two alleles is the presence of a proline residue at position 186 in SUB1A-2, instead of a serine residue in SUB1A-1 [15,17,19,58]. SUB1A-1 is specifically identified in submergence-tolerant cultivars, such as FR13A, and it can be strongly induced upon submergence to exert a determinant role in submergence stress responses, whereas the SUB1A-2 allele, which is only found in submergence-sensitive cultivars, such as IR29, is poorly induced during submergence [15,58].
Under flooding conditions, ethylene inhibits internode growth in submergence-tolerant rice cultivars by inducing SUB1A expression [19]. As a transcription factor, SUB1A-1 can regulate a spectrum of genes involved in anaerobic metabolism, reactive oxygen species regulation, and leaf senescence during and after submergence by directly interacting with the GCC box in their promoters. During submergence, mitogen-activated protein kinase3 (MPK3) can specifically interact with and phosphorylate SUB1A-1 to activate it. Interestingly, activated SUB1A-1 can promote the expression of MPK3 by binding to the GCC box in its promoter, thus forming a positive feedback loop during submergence [59]. SUB1A-1 also activates the transcription of other Group VII ERF transcription factors, such as ERF66 and ERF67, and enhances submergence tolerance by regulating various hypoxia-responsive genes [60]. Recently, one report showed that SUB1A-1 can interact with ADA2b (ALTERATION/DEFICIENCY IN AC-TIVATION 2) of the ADA2b-GCN5 (GENERAL CONTROL NON-REPRESSIBLE 5) acetyltransferase complex and recruits this complex to modify the chromatin structure of the ERF66/ERF67 promoter region and activate gene expression, which in turn enhances rice submergence tolerance [58]. In addition, the expression of DWARF1 (DWF1) and DWARF4 (DWF4), two key genes involved in the biosynthesis of brassinosteroids (BRs), is significantly higher in M202-Sub1 (tolerant variety) than in M202 (intolerant variety) during submergence, suggesting that SUB1A-1 can activate BR signaling. Consistent with the gene expression data, the SUB1A genotype has higher BR levels after submergence than those of the intolerant genotype. BRs in turn activate the transcription of the GA catabolic gene GA2 oxidase7 (GA2ox7) or the protein accumulation of slender rice-1 (SLR1) and SLR1 like-1 (SLRL1), which negatively regulate GA responsiveness [61,62]. Apart from GA and BR signaling, the increase in ethylene levels also promotes the expression of ABA 8′-hydroxylases (ABA8ox), which stimulates the catabolism of bioactive ABA to unstable ABA. However, the presence or absence of SUB1A does not affect the decline in ABA levels in response to submergence, suggesting that growth inhibition in these submergence-tolerant plants is mainly brought about through the regulation of GA synthesis and signaling [63]. SUB1A overexpression also delays leaf yellowing and enhances recovery from dark stress. Physiological analysis revealed that SUB1A postpones dark-induced senescence by maintaining chlorophyll and carbohydrate reserves in the photosynthetic tissue. In line with this, RT-PCR results showed that SUB1A can downregulate the expression of senescence-associated genes (SAGs), including stay-green (SGR), red chlorophyll catabolite reductase1 (RCCR1), and non-yellow coloring1 (NYC1), when submerged [64].
Interestingly, the above shown SUB1A-mediated regulatory mechanisms are not usually active for anoxic seed germination [14], whereas the CIPK15 pathway is essential for this growth stage. But both SUB1A and CIPK15 play important roles in carbohydrate consumption in adult plants upon submergence. SUB1A can inhibit SUB1C, a positive regulator of the CIPK15-SnRK1A-MYBS1-RAmy3D pathway, resulting in limited growth under submerged conditions [65].
3.2.2. SK1/SK2 Pathway
In contrast to the quiescent strategy, deepwater rice adapts to submergence by rapidly elongating its internodes, thereby maintaining its leaves above the water surface. Through map-based cloning, another major submergence tolerance QTL of qTIL12 in deepwater rice was cloned, which contains two ERF transcription factors genes: SNORKEL1 (SK1) and SNORKEL2 (SK2) [18]. SK1/SK2 is not expressed under aerobic conditions; however, as early as 3 h of submergence treatment leads to their rapid accumulation. The expression of SK1 and SK2 is significantly induced by ethylene treatments, and overexpression of these genes promotes internode elongation in normal paddy rice, suggesting that SK1 and SK2 regulate internode elongation in response to ethylene [18]. SK1/SK2 cultivars can accumulate higher levels of GAs that promote internode elongation [18]. EIL1, a rice homolog of Arabidopsis EIN3, positively regulates ethylene response in rice [66]. A recent study showed that EIL1 can target the promoters of SK1 and SK2 to activate their expression.
SEMIDWARF1 (SD1), encoding a GA biosynthesis enzyme, was identified as the qTIL1 causative gene [67]. Two amino acid differences in SD1 exist between the deepwater rice variety C9285 and normal paddy rice, resulting in higher enzymatic activities in deepwater rice than in normal paddy rice. EIL1 can also directly bind to and activate SD1 expression, thus promoting GA accumulation, particularly GA4, which promotes internode elongation. Recently, a gene in the SUB1A-binding protein family, SAB23, encoding a plant homeodomain (PHD)-type transcription factor, was identified as a novel negative regulator of submergence tolerance in rice. SAB23 binds to the promoter of CYTOCHROME P450 714B2 (CYP714B2), which encodes a GA13-oxidase that catalyzes the conversion of GA53 to GA19, thus promoting the accumulation of bioactive GAs [68]. These findings clearly indicate that the activation of GA signaling is a key step in conferring submergence tolerance to deepwater rice.
Recently, an antagonistic regulatory mechanism of the GA-regulated stem growth in deepwater rice has been explored. ACCELERATOR OF IN-TERNODE ELONGATION 1 (ACE1), which encodes a protein of unknown function, confers cells of the intercalary meristematic region with the competence for cell division, leading to internode elongation in the presence of GA. In contrast, DECEL-ERATOR OF INTERNODE ELONGATION 1 (DEC1), which encodes a zinc finger transcription factor, suppresses internode elongation in response to GA [69]. More importantly, an analysis of genetic diversity suggests that mutations in ACE1 and DEC1 have historically contributed to the selection of shorter plants in domesticated populations of rice to increase their resistance to lodging, and of taller plants in wild species of rice to adapt to grow in deep water. Based on these findings, the regulation or modification of the GA pathway will be an efficient strategy for the molecular breeding of submergence-tolerant rice cultivars.
4. Conclusions and Perspectives
During evolution, rice has developed several different strategies to cope with flooding/submergence stress, which employ different genes or proteins to induce various morphological and physiological changes to survive and grow. During the seed germination and seedling establishment stages, CIPK15-mediated sugar signaling and the crosstalk between GA, ABA, JA, and auxin signaling commonly determine the tolerance of rice to submergence. At the vegetative growth stage, SUB1A functions as the central component of the submergence response that regulates multiple processes, such as entering a quiescent stage to preserve carbohydrate resources and helping the submerged plants acclimate during de-submergence. In contrast, SK1/SK2 promotes GA accumulation and triggers rapid shoot/internode elongation using carbohydrate reserves to escape flooding. Despite these exciting findings, the entire landscape of submergence tolerance in rice is not entirely depicted, and further exploration in the field is necessary to completely understand the mechanisms. In addition, the identification of genes that regulate submergence tolerance at both the seeding establishment and vegetative growth stages will be quite useful. In future, CRISPR-CAS9-mediated gene editing technology and molecular marker-assisted crossing breeding will be used to develop valuable commercialized varieties that harbor better tolerance to flooding.
Author Contributions
J.W., M.H., Y.H., J.Z. and C.L. wrote the draft of this manuscript. Y.M. reviewed and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Special Fund for Scientific Innovation Strategy-Construction of High-Level Academy of Agriculture Science (R2021PY-QF001), Seed industry revitalization project of the special fund for rural revitalization strategy in Guangdong Province (2022NPY00005), and the Guangdong Key Laboratory of New Technology in Rice Breeding (2023B1212060042).
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Perata, P.; Voesenek, L.A. Submergence tolerance in rice requires Sub1A, an ethylene-response-factor-like gene. Plant Sci. 2007, 12, 43–46. [Google Scholar] [CrossRef] [PubMed]
- Nishiuchi, S.; Yamauchi, T.; Takahashi, H.; Kotula, L.; Nakazono, M.J.R. Mechanisms for coping with submergence and waterlogging in rice. Rice 2012, 5, 2. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, J.; Greenway, H. Mechanisms of anoxia tolerance in plants. I. Growth, survival and anaerobic catabolism. Funct. Plant Biol. 2003, 30, 1–47. [Google Scholar] [CrossRef] [PubMed]
- Bailey-Serres, J.; Lee, S.C.; Brinton, E. Waterproofing crops: Effective flooding survival strategies. Plant Physiol. 2012, 160, 1698–1709. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, U.; Hussain, H.A.; Hussain, S.; Ashraf, U.; Khaliq, A.; Hussain, S. Submergence stress in rice: Physiological disorders, tolerance mechanisms, and management. In Plant Abiotic Stress. Tolerance: Agronomic, Molecular and Biotechnological Approaches; Springer: Cham, Switzerland, 2019; pp. 173–189. [Google Scholar]
- Lee, K.; Chen, P.W.; Yu, S. Metabolic adaptation to sugar/O2 deficiency for anaerobic germination and seedling growth in rice. Plant Cell Environ. 2014, 37, 2234–2244. [Google Scholar] [CrossRef] [PubMed]
- Pucciariello, C. Molecular mechanisms supporting rice germination and coleoptile elongation under low oxygen. Plants 2020, 9, 1037. [Google Scholar] [CrossRef]
- Haque, M.A.; Rafii, M.Y.; Yusoff, M.M.; Ali, N.S.; Yusuff, O.; Arolu, F.; Anisuzzaman, M. Flooding tolerance in Rice: Adaptive mechanism and marker-assisted selection breeding approaches. Mol. Biol. Rep. 2023, 50, 2795–2812. [Google Scholar] [CrossRef]
- Yu, S.M.; Lee, H.T.; Lo, S.F.; Ho, T.H.D. How does rice cope with too little oxygen during its early life? New Phytol. 2021, 229, 36–41. [Google Scholar] [CrossRef]
- Phukan, U.J.; Jindal, S.; Laldinsangi, C.; Singh, P.K.; Longchar, B. A microscopic scenario on recovery mechanisms under waterlogging and submergence stress in rice. Planta 2024, 259, 9. [Google Scholar] [CrossRef]
- Colmer, T.D. Aerenchyma and an inducible barrier to radial oxygen loss facilitate root aeration in upland, paddy and deep-water rice (Oryza sativa L.). Ann. Bot. 2002, 91, 301–309. [Google Scholar] [CrossRef]
- Nagai, K.; Ashikari, M. Molecular mechanism of internode elongation in rice. Breed. Sci. 2023, 73, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.B.; Ram, P.C. Physiological and molecular basis of susceptibility and tolerance of rice plants to complete submergence. Ann. Bot. 2003, 91, 227–241. [Google Scholar] [CrossRef] [PubMed]
- Magneschi, L.; Perata, P. Rice germination and seedling growth in the absence of oxygen. Ann. Bot. 2009, 103, 181–196. [Google Scholar] [CrossRef]
- Bailey-Serres, J.; Fukao, T.; Ronald, P.; Ismail, A.; Heuer, S.; Mackill, D. Submergence tolerant rice: SUB1’s journey from landrace to modern cultivar. Rice 2010, 3, 138–147. [Google Scholar] [CrossRef]
- Kumar, A.; Nayak, A.; Hanjagi, P.; Kumari, K.; Vijayakumar, S.; Mohanty, S.; Tripathi, R.; Panneerselvam, P. Submergence stress in rice: Adaptive mechanisms, coping strategies and future research needs. Environ. Exp. Bot. 2021, 186, 104448. [Google Scholar] [CrossRef]
- Fukao, T.; Xu, K.; Ronald, P.C.; Bailey-Serres, J. A variable cluster of ethylene response factor–like genes regulates metabolic and developmental acclimation responses to submergence in rice. Plant Cell 2006, 18, 2021–2034. [Google Scholar] [CrossRef]
- Hattori, Y.; Nagai, K.; Furukawa, S.; Song, X.-J.; Kawano, R.; Sakakibara, H.; Wu, J.; Matsumoto, T.; Yoshimura, A.; Kitano, H. The ethylene response factors SNORKEL1 and SNORKEL2 allow rice to adapt to deep water. Nature 2009, 460, 1026–1030. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Xu, X.; Fukao, T.; Canlas, P.; Maghirang-Rodriguez, R.; Heuer, S.; Ismail, A.M.; Bailey-Serres, J.; Ronald, P.C.; Mackill, D.J. Sub1A is an ethylene-response-factor-like gene that confers submergence tolerance to rice. Nature 2006, 442, 705–708. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Ladha, J.K. Direct seeding of rice: Recent developments and future research needs. Adv. Agron. 2011, 111, 297–413. [Google Scholar]
- Taiz, L.; Zeiger, E.; Møller, I.M.; Murphy, A. Plant Physiology and Development; Sinauer Associates, Inc.: Sunderland, MA, USA, 2015. [Google Scholar]
- Zahra, N.; Hafeez, M.B.; Shaukat, K.; Wahid, A.; Hussain, S.; Naseer, R.; Raza, A.; Iqbal, S.; Farooq, M. Hypoxia and Anoxia Stress: Plant responses and tolerance mechanisms. J. Agron. Crop Sci. 2021, 207, 249–284. [Google Scholar] [CrossRef]
- Hwang, Y.-S.; Thomas, B.; Rodriguez, R. Differential expression of rice α-amylase genes during seedling development under anoxia. Plant Mol. Biol. 1999, 40, 911–920. [Google Scholar] [CrossRef]
- Narsai, R.; Secco, D.; Schultz, M.D.; Ecker, J.R.; Lister, R.; Whelan, J. Dynamic and rapid changes in the transcriptome and epigenome during germination and in developing rice (Oryza sativa) coleoptiles under anoxia and re-oxygenation. Plant J. 2017, 89, 805–824. [Google Scholar] [CrossRef] [PubMed]
- Nghi, K.N.; Tagliani, A.; Mariotti, L.; Weits, D.A.; Perata, P.; Pucciariello, C. Auxin is required for the long coleoptile trait in japonica rice under submergence. New Phytol. 2021, 229, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.-A.; Lin, C.-C.; Lee, K.-W.; Chen, J.-L.; Huang, L.-F.; Ho, S.-L.; Liu, H.-J.; Hsing, Y.-I.; Yu, S.-M. The SnRK1A protein kinase plays a key role in sugar signaling during germination and seedling growth of rice. Plant Cell 2007, 19, 2484–2499. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-W.; Chen, P.-W.; Lu, C.-A.; Chen, S.; Ho, T.-H.D.; Yu, S.-M. Coordinated responses to oxygen and sugar deficiency allow rice seedlings to tolerate flooding. Sci. Signal. 2009, 2, ra61. [Google Scholar] [CrossRef]
- Sánchez-Barrena, M.J.; Martínez-Ripoll, M.; Zhu, J.-K.; Albert, A. The structure of the Arabidopsis thaliana SOS3: Molecular mechanism of sensing calcium for salt stress response. J. Mol. Biol. 2005, 345, 1253–1264. [Google Scholar] [CrossRef]
- Kurusu, T.; Hamada, J.; Nokajima, H.; Kitagawa, Y.; Kiyoduka, M.; Takahashi, A.; Hanamata, S.; Ohno, R.; Hayashi, T.; Okada, K. Regulation of microbe-associated molecular pattern-induced hypersensitive cell death, phytoalexin production, and defense gene expression by calcineurin B-like protein-interacting protein kinases, OsCIPK14/15, in rice cultured cells. Plant Physiol. 2010, 153, 678–692. [Google Scholar] [CrossRef] [PubMed]
- Ye, N.H.; Wang, F.Z.; Shi, L.; Chen, M.X.; Cao, Y.Y.; Zhu, F.Y.; Wu, Y.Z.; Xie, L.J.; Liu, T.Y.; Su, Z.Z. Natural variation in the promoter of rice calcineurin B-like protein10 (OsCBL10) affects flooding tolerance during seed germination among rice subspecies. Plant J. 2018, 94, 612–625. [Google Scholar] [CrossRef]
- Chen, Y.-S.; David Ho, T.-H.; Liu, L.; Lee, D.H.; Lee, C.-H.; Chen, Y.-R.; Lin, S.-Y.; Lu, C.-A.; Yu, S.-M. Sugar starvation-regulated MYBS2 and 14-3-3 protein interactions enhance plant growth, stress tolerance, and grain weight in rice. Proc. Natl. Acad. Sci. USA 2019, 116, 21925–21935. [Google Scholar] [CrossRef]
- Lin, C.-R.; Lee, K.-W.; Chen, C.-Y.; Hong, Y.-F.; Chen, J.-L.; Lu, C.-A.; Chen, K.-T.; Ho, T.-H.D.; Yu, S.-M. SnRK1A-interacting negative regulators modulate the nutrient starvation signaling sensor SnRK1 in source-sink communication in cereal seedlings under abiotic stress. Plant Cell 2014, 26, 808–827. [Google Scholar] [CrossRef]
- Kretzschmar, T.; Pelayo, M.; Trijatmiko, K.; Gabunada, L.; Alam, R.; Jimenez, R.; Mendioro, M.; Slamet-Loedin, I.; Sreenivasulu, N.; Bailey-Serres, J. A trehalose-6-phosphate phosphatase enhances anaerobic germination tolerance in rice. Nat Plants 2015, 1, 15124. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Zhao, J.; Fu, H.; Yang, T.; Dong, J.; Yang, W.; Chen, L.; Zhou, L.; Wang, J.; Liu, B. Genome-wide identification, expression and functional analysis reveal the involvement of FCS-like zinc finger gene family in submergence response in rice. Rice 2021, 14, 76. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, J.; Chen, S.; Ma, X.; Wei, H.; Chen, C.; Gao, N.; Zou, Y.; Kong, D.; Li, T.; et al. An APETALA2/ethylene responsive factor, OsEBP89 knockout enhances adaptation to direct-seeding on wet land and tolerance to drought stress in rice. Mol. Genet. Genom. 2020, 295, 941–956. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Zhang, G.; Cui, Z.; Kong, X.; Yu, X.; Gui, R.; Han, Y.; Li, Z.; Lang, H.; Hua, Y. Regain flood adaptation in rice through a 14-3-3 protein OsGF14h. Nat. Commun. 2022, 13, 5664. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Sun, S.; Zhao, J.; Huang, Z.; Peng, L.; Huang, C.; Tang, Z.; Huang, Q.; Wang, Z. UDP-glucosyltransferase OsUGT75A promotes submergence tolerance during rice seed germination. Nat. Commun. 2023, 14, 2296. [Google Scholar] [CrossRef] [PubMed]
- Kawai, M.; Uchimiya, H. Coleoptile senescence in rice (Oryza sativa L.). Ann. Bot. 2000, 86, 405–414. [Google Scholar] [CrossRef]
- Si-Ammour, A.; Windels, D.; Arn-Bouldoires, E.; Kutter, C.; Ailhas, J.; Meins, F., Jr.; Vazquez, F. miR393 and secondary siRNAs regulate expression of the TIR1/AFB2 auxin receptor clade and auxin-related development of Arabidopsis leaves. Plant Physiol. 2011, 157, 683–691. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Han, N.; Xie, Y.; Fang, K.; Yang, Y.; Zhu, M.; Wang, J.; Bian, H. The miR393a/target module regulates seed germination and seedling establishment under submergence in rice (Oryza sativa L.). Plant Cell Environ. 2016, 39, 2288–2302. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.W.; Chen, J.J.; Wu, C.S.; Chang, H.C.; Chen, H.Y.; Kuo, H.H.; Lee, Y.S.; Chang, Y.L.; Chang, H.C.; Shiue, S.; et al. Auxin plays a role in the adaptation of rice to anaerobic germination and seedling establishment. Plant Cell Environ. 2023, 46, 1157–1175. [Google Scholar] [CrossRef]
- Yang, J.H.; Han, S.J.; Yoon, E.K.; Lee, W.S. Evidence of an auxin signal pathway, microRNA167-ARF8-GH3, and its response to exogenous auxin in cultured rice cells. Nucleic Acids Res. 2006, 34, 1892–1899. [Google Scholar] [CrossRef]
- Ludwig-Müller, J. Auxin conjugates: Their role for plant development and in the evolution of land plants. J. Exp. Bot. 2011, 62, 1757–1773. [Google Scholar] [CrossRef] [PubMed]
- Hao, B.; Zhang, R.; Zhang, C.; Wen, N.; Xia, Y.; Zhao, Y.; Li, Q.; Qiao, L.; Li, W.J.P. Characterization of OsPIN2 Mutants Reveal Novel Roles for Reactive Oxygen Species in Modulating Not Only Root Gravitropism but Also Hypoxia Tolerance in Rice Seedlings. Plants 2024, 13, 476. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, W. Aeration in higher plants. In Advances in Botanical Research; Elsevier: Amsterdam, The Netherlands, 1980; Volume 7, pp. 225–332. [Google Scholar]
- Shiono, K.; Ogawa, S.; Yamazaki, S.; Isoda, H.; Fujimura, T.; Nakazono, M.; Colmer, T.D. Contrasting dynamics of radial O2-loss barrier induction and aerenchyma formation in rice roots of two lengths. Ann. Bot. 2011, 107, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Colmer, T.D.; Voesenek, L. Flooding tolerance: Suites of plant traits in variable environments. Funct. Plant Biol. 2009, 36, 665–681. [Google Scholar] [CrossRef] [PubMed]
- Malik, A.; Colmer, T.; Lambers, H.; Schortemeyer, M. Aerenchyma formation and radial O2 loss along adventitious roots of wheat with only the apical root portion exposed to O2 deficiency. Plant Cell Environ. 2003, 26, 1713–1722. [Google Scholar] [CrossRef]
- Setter, T.; Waters, I. Review of prospects for germplasm improvement for waterlogging tolerance in wheat, barley and oats. Plant Soil. 2003, 253, 1–34. [Google Scholar] [CrossRef]
- Lin, C.; Zhang, Z.; Shen, X.; Liu, D.; Pedersen, O. Flooding-adaptive root and shoot traits in rice. Funct. Plant Biol. 2024, 51, FP23226. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Ogorek, L.L.P.; Liu, D.; Pedersen, O.; Sauter, M. A quantitative trait locus conferring flood tolerance to deepwater rice regulates the formation of two distinct types of aquatic adventitious roots. New Phytol. 2023, 238, 1403–1419. [Google Scholar] [CrossRef] [PubMed]
- Hattori, Y.; Nagai, K.; Ashikari, M. Rice growth adapting to deepwater. Curr. Opin. Plant Biol. 2011, 14, 100–105. [Google Scholar] [CrossRef]
- Setter, T.L.; Laureles, E.V. The beneficial effect of reduced elongation growth on submergence tolerance of rice. J. Exp. Bot. 1996, 47, 1551–1559. [Google Scholar] [CrossRef]
- Bui, L.T.; Ella, E.S.; Dionisio-Sese, M.L.; Ismail, A.M. Morpho-physiological changes in roots of rice seedling upon submergence. Rice Sci. 2019, 26, 167–177. [Google Scholar] [CrossRef]
- Xu, K.; Mackill, D.J. A major locus for submergence tolerance mapped on rice chromosome 9. Mol. Breed. 1996, 2, 219–224. [Google Scholar] [CrossRef]
- Jung, K.-H.; Seo, Y.-S.; Walia, H.; Cao, P.; Fukao, T.; Canlas, P.E.; Amonpant, F.; Bailey-Serres, J.; Ronald, P.C. The submergence tolerance regulator Sub1A mediates stress-responsive expression of AP2/ERF transcription factors. Plant Physiol. 2010, 152, 1674–1692. [Google Scholar] [CrossRef] [PubMed]
- Perata, P. The rice SUB1A gene: Making adaptation to submergence and post-submergence possible. Plant Cell Environ. 2018, 41, 717–720. [Google Scholar] [CrossRef]
- Lin, C.-C.; Lee, W.-J.; Zeng, C.-Y.; Chou, M.-Y.; Lin, T.-J.; Lin, C.-S.; Ho, M.-C.; Shih, M.-C. SUB1A-1 anchors a regulatory cascade for epigenetic and transcriptional controls of submergence tolerance in rice. Pnas Nexus. 2023, 2, pgad229. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Sinha, A.K. A positive feedback loop governed by SUB1A1 interaction with MITOGEN-ACTIVATED PROTEIN KINASE3 imparts submergence tolerance in rice. Plant Cell 2016, 28, 1127–1143. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-C.; Chao, Y.-T.; Chen, W.-C.; Ho, H.-Y.; Chou, M.-Y.; Li, Y.-R.; Wu, Y.-L.; Yang, H.-A.; Hsieh, H.; Lin, C.-S. Regulatory cascade involving transcriptional and N-end rule pathways in rice under submergence. Proc. Natl. Acad. Sci. USA 2019, 116, 3300–3309. [Google Scholar] [CrossRef]
- Schmitz, A.J.; Folsom, J.J.; Jikamaru, Y.; Ronald, P.; Walia, H. SUB1A-mediated submergence tolerance response in rice involves differential regulation of the brassinosteroid pathway. New Phytol. 2013, 198, 1060–1070. [Google Scholar] [CrossRef]
- Fukao, T.; Bailey-Serres, J. Submergence tolerance conferred by Sub1A is mediated by SLR1 and SLRL1 restriction of gibberellin responses in rice. Proc. Natl. Acad. Sci. USA 2008, 105, 16814–16819. [Google Scholar] [CrossRef]
- Saika, H.; Okamoto, M.; Miyoshi, K.; Kushiro, T.; Shinoda, S.; Jikumaru, Y.; Fujimoto, M.; Arikawa, T.; Takahashi, H.; Ando, M.J.P.; et al. Ethylene promotes submergence-induced expression of OsABA8ox1, a gene that encodes ABA 8′-hydroxylase in rice. Plant Cell Physiol. 2007, 48, 287–298. [Google Scholar] [CrossRef]
- Fukao, T.; Yeung, E.; Bailey-Serres, J. The submergence tolerance gene SUB1A delays leaf senescence under prolonged darkness through hormonal regulation in rice. Plant Physiol. 2012, 160, 1795–1807. [Google Scholar] [CrossRef] [PubMed]
- Kudahettige, N.; Pucciariello, C.; Parlanti, S.; Alpi, A.; Perata, P. Regulatory interplay of the Sub1A and CIPK15 pathways in the regulation of α-amylase production in flooded rice plants. Plant Biol. 2011, 13, 611–619. [Google Scholar] [CrossRef] [PubMed]
- Mao, C.; Wang, S.; Jia, Q.; Wu, P. OsEIL1, a rice homolog of the Arabidopsis EIN3 regulates the ethylene response as a positive component. Plant Mol. Biol. 2006, 61, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Kuroha, T.; Nagai, K.; Gamuyao, R.; Wang, D.R.; Furuta, T.; Nakamori, M.; Kitaoka, T.; Adachi, K.; Minami, A.; Mori, Y. Ethylene-gibberellin signaling underlies adaptation of rice to periodic flooding. Science 2018, 361, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Duan, A.; Liu, R.; Liu, C.; Wu, F.; Su, H.; Zhou, S.; Huang, M.; Tian, X.; Jia, H.; Liu, Y.J. Mutation of the gene encoding the PHD-type transcription factor SAB23 confers submergence tolerance in rice. J. Exp. Bot. 2024, 75, 168–179. [Google Scholar] [CrossRef]
- Nagai, K.; Mori, Y.; Ishikawa, S.; Furuta, T.; Gamuyao, R.; Niimi, Y.; Hobo, T.; Fukuda, M.; Kojima, M.; Takebayashi, Y. Antagonistic regulation of the gibberellic acid response during stem growth in rice. Nature 2020, 584, 109–114. [Google Scholar] [CrossRef]
Figure 1.
CIPK15-mediated (A) and phytohormone-mediated (B) signaling pathways in regulating rice submergence tolerance during seed germination and seedling establishment stages.
Figure 1.
CIPK15-mediated (A) and phytohormone-mediated (B) signaling pathways in regulating rice submergence tolerance during seed germination and seedling establishment stages.
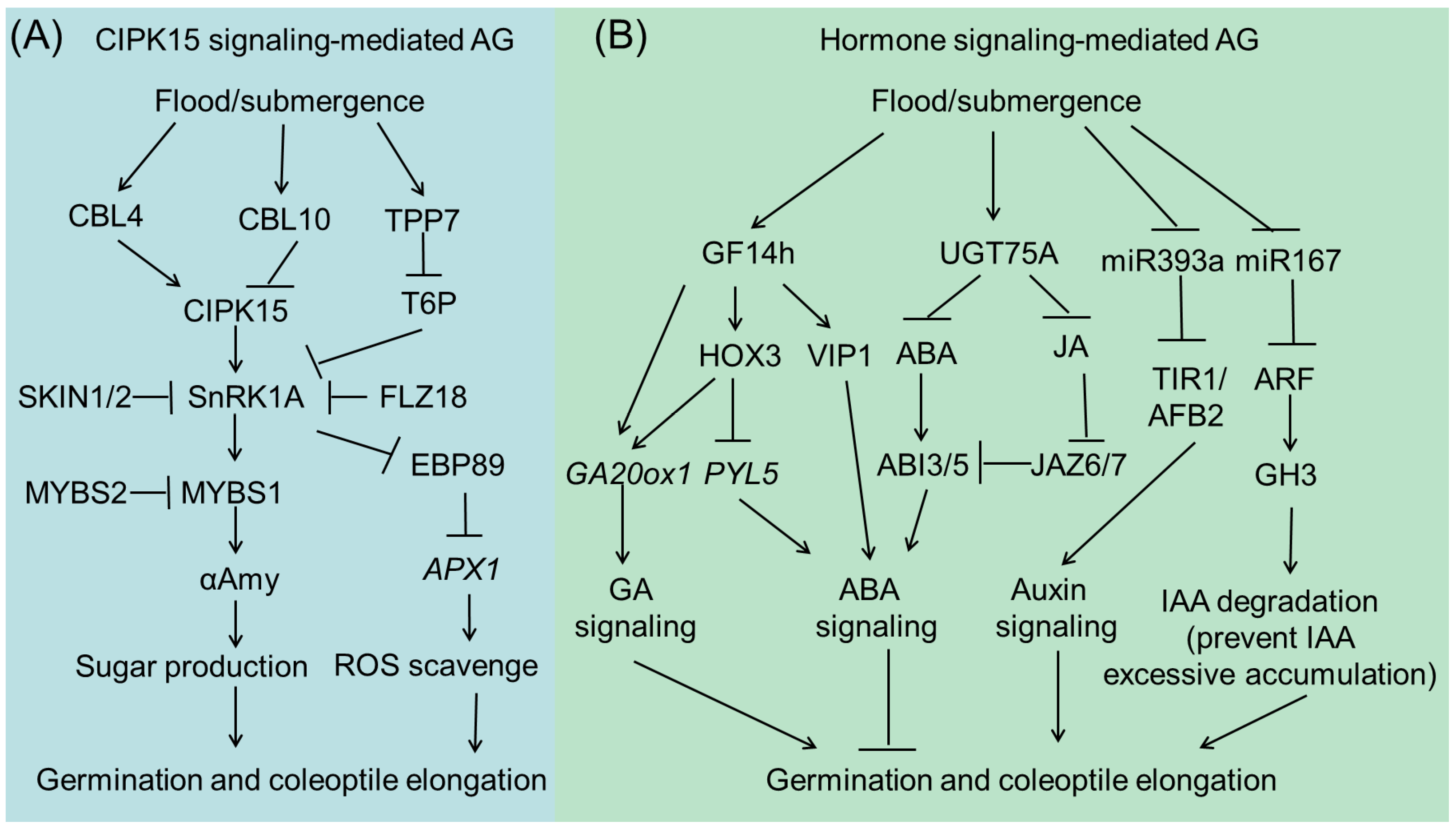
Figure 2.
SUB1A-regulated (A) and SKl/SK2-regulated (B) submergence tolerance during rice vegetative stage.
Figure 2.
SUB1A-regulated (A) and SKl/SK2-regulated (B) submergence tolerance during rice vegetative stage.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, J.; Han, M.; Huang, Y.; Zhao, J.; Liu, C.; Ma, Y. Flooding Tolerance of Rice: Regulatory Pathways and Adaptive Mechanisms. Plants 2024, 13, 1178. https://doi.org/10.3390/plants13091178
AMA Style
Wang J, Han M, Huang Y, Zhao J, Liu C, Ma Y. Flooding Tolerance of Rice: Regulatory Pathways and Adaptive Mechanisms. Plants. 2024; 13(9):1178. https://doi.org/10.3390/plants13091178
Chicago/Turabian StyleWang, Jing, Mingzhen Han, Yongxiang Huang, Junliang Zhao, Chuanguang Liu, and Yamei Ma. 2024. "Flooding Tolerance of Rice: Regulatory Pathways and Adaptive Mechanisms" Plants 13, no. 9: 1178. https://doi.org/10.3390/plants13091178
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.