
New Insights into the Bio-Chemical Changes in Wheat Induced by Cd and Drought: What Can We Learn on Cd Stress Using Neutron Imaging?
 , ,
, ,  , and
, and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. The Effects of Cd and Cd + Drought on Gas Exchange Parameters
2.2. The Levels of Cd in Wheat Tissues under Cd and Cd + Drought Stresses
2.3. Foliar Chlorophyll Content and Fluorescence under Cd and Cd + Drought Stresses
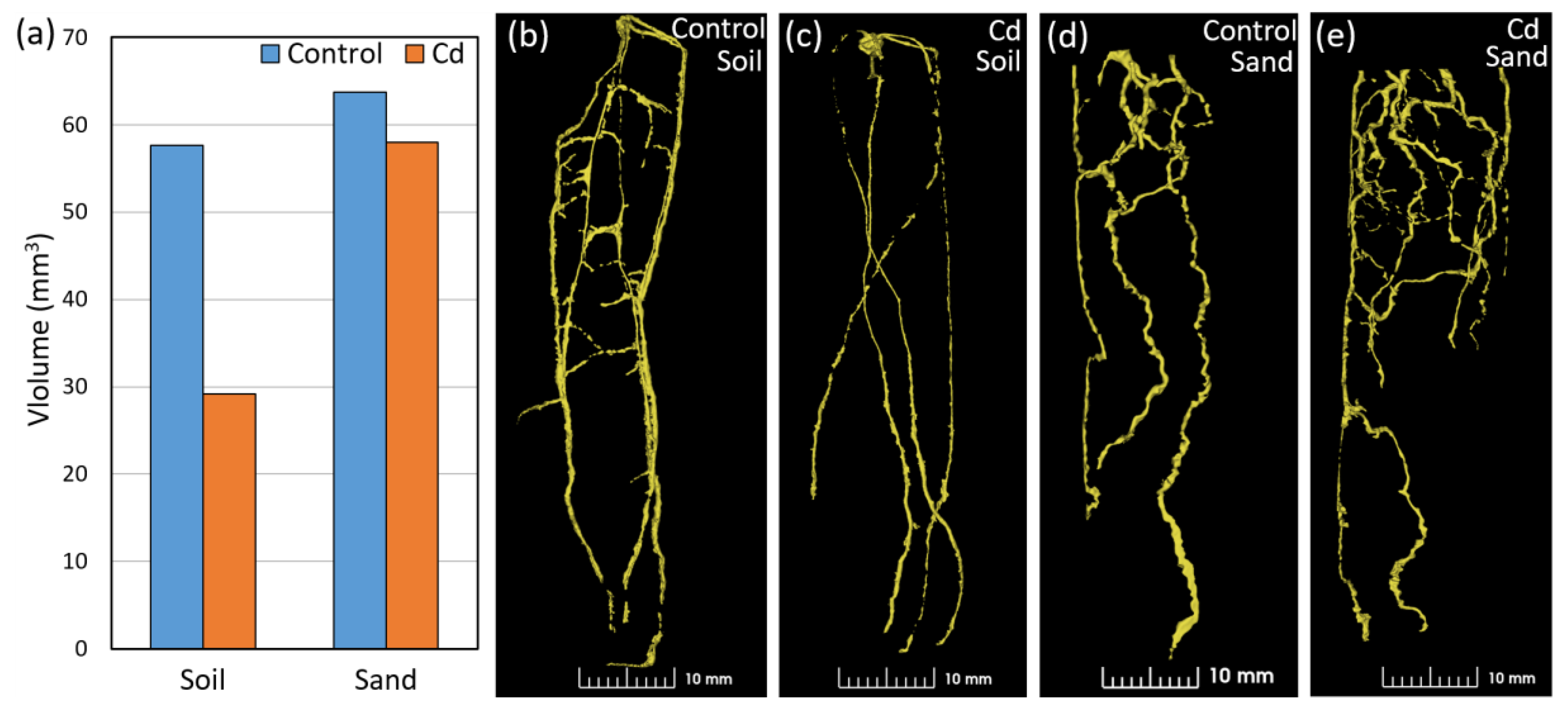
2.4. Effects of Growing Media on Root Volume and Architecture as Evaluated Using NCT
2.5. Root Volume and Architecture of Wheat under Cd Stress Evaluated Using NCT
2.6. Water Distribution in Different Growing Media as Monitored Using NCT
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Soil and Sand
4.3. Greenhouse Experiment for Physiological Traits
4.3.1. Growing Conditions and Treatments
4.3.2. Gas Exchange Measurements
4.3.3. Foliar Chlorophyll Content and Chlorophyll Fluorescence
4.3.4. Dry Weight and Cd Content
4.4. Root Architecture Assay Based on NCT
4.4.1. Growing Conditions and Treatments
4.4.2. NCT for Root Analysis
4.4.3. Image Reconstruction and Root Segmentation
4.5. Data Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lockwood, A.P.M. Effects of Pollutants on Aquatic Organisms; Cambridge University Press: Cambridge, UK, 1976; Volume 2. [Google Scholar]
- Taylor, G.; Pahlsson, A.; Benglsson, G.; Baath, E.; Tranvik, L. Heavy metal ecology and terrestrial plant, microorganisms and invertebrates—A Review. Water Air Soil Pollut 1989, 47, 189–215. [Google Scholar] [CrossRef]
- Backe, C.; Björn, H.; Holmqvist, J.; Andreasson, F. Kadmiumsituationen i Skåne. Exempel på kadmiumkällor och halter i den skånska miljön. Skåne Utveckl. 2003, 2003, 46. [Google Scholar]
- Das, P.; Samantaray, S.; Rout, G.R. Studies on cadmium toxicity in plants: A review. Environ. Pollut. 1997, 98, 29–36. [Google Scholar] [CrossRef]
- Di Toppi, L.S.; Gabbrielli, R. Response to cadmium in higher plants. Environ. Exp. Bot. 1999, 41, 105–130. [Google Scholar] [CrossRef]
- Nedjimi, B.; Daoud, Y. Cadmium accumulation in Atriplex halimus subsp. schweinfurthii and its influence on growth, proline, root hydraulic conductivity and nutrient uptake. Flora Morphol. Distrib. Funct. Ecol. Plants 2009, 204, 316–324. [Google Scholar] [CrossRef]
- Feng, R.; Wei, C.; Tu, S.; Ding, Y.; Song, Z. A dual role of se on Cd toxicity: Evidences from the uptake of Cd and some essential elements and the growth responses in paddy rice. Biol. Trace Element Res. 2013, 151, 113–121. [Google Scholar] [CrossRef]
- Walley, J.W.; Huerta, A.J. Exposure to environmentally relevant levels of cadmium primarily impacts transpiration in field-grown soybean. J. Plant Nutr. 2010, 33, 1519–1530. [Google Scholar] [CrossRef]
- Song, J.; Finnegan, P.M.; Liu, W.; Xiang, L.; Yong, J.W.H.; Xu, J.; Zhang, Q.; Wen, Y.; Qin, K.; Guo, J.; et al. Mechanisms underlying enhanced Cd translocation and tolerance in roots of Populus euramericana in response to nitrogen fertilization. Plant Sci. 2019, 287, 110206. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Abbas, T.; Zia-ur-Rehman, M.; Hannan, F.; Keller, C.; Al-Wabel, M.I.; Ok, Y.S. Cadmium minimization in wheat: A critical review. Ecotoxicol. Environ. Saf. 2016, 130, 43–53. [Google Scholar] [CrossRef]
- Greger, M.; Löfstedt, M. Comparison of uptake and distribution of cadmium in different cultivars of bread and durum wheat. Crop. Sci. 2004, 44, 501–507. [Google Scholar] [CrossRef]
- Jafarnejadi, A.R.; Homaee, M.; Sayyad, G.; Bybordi, M. Large scale spatial variability of accumulated cadmium in the wheat farm grains. Soil Sediment Contam. Int. J. 2011, 20, 98–113. [Google Scholar] [CrossRef]
- Abedi, T.; Mojiri, A. Cadmium uptake by wheat (Triticum aestivum L.): An overview. Plants 2020, 9, 500. [Google Scholar] [CrossRef]
- Hu, H.; Jin, Q.; Kavan, P. A study of heavy metal pollution in China: Current status, pollution-control policies and countermeasures. Sustainability 2014, 6, 5820–5838. [Google Scholar] [CrossRef]
- Wu, Y. General Standard for Contaminants and Toxins in Food and Feed; Codex Stan 193–1995; FAO/WHO: Geneva, Switzerland, 2014. [Google Scholar] [CrossRef]
- Grant, C.A.; Bailey, L.D. Nitrogen, phosphorus and zinc management effects on grain yield and cadmium concentration in two cultivars of durum wheat. Can. J. Plant Sci. 1998, 78, 63–70. [Google Scholar] [CrossRef]
- Gao, X.; Grant, C.A. Cadmium and zinc concentration in grain of durum wheat in relation to phosphorus fertilization, crop sequence and tillage management. Appl. Environ. Soil Sci. 2012, 2012, 817107. [Google Scholar] [CrossRef]
- Guo, G.; Lei, M.; Wang, Y.; Song, B.; Yang, J. Accumulation of As, Cd, and Pb in sixteen wheat cultivars grown in contaminated soils and associated health risk assessment. Int. J. Environ. Res. Public Health 2018, 15, 2601. [Google Scholar] [CrossRef]
- Eriksson, J.E.; Söderström, M. Cadmium in soil and winter wheat grain in southern Sweden: I. factors influencing Cd levels in soils and grain. Acta Agric. Scand. Sect. B Soil Plant Sci. 1996, 46, 240–248. [Google Scholar] [CrossRef]
- Hellstrand, S.; Landner, L. Cadmium in Fertilizers, Soil, Crops and Foods-the Swedish Situation; National Chemicals Inspectorate: Solna, Sweden, 1998. [Google Scholar]
- Khan, M.U.; Shahbaz, N.; Waheed, S.; Mahmood, A.; Shinwari, Z.K.; Malik, R.N. Comparative health risk surveillance of heavy metals via dietary foodstuff consumption in different land-use types of Pakistan. Hum. Ecol. Risk Assess. Int. J. 2015, 22, 168–186. [Google Scholar] [CrossRef]
- Simmons, R.; Pongsakul, P.; Saiyasitpanich, D.; Klinphoklap, S. Elevated levels of cadmium and zinc in paddy soils and elevated levels of cadmium in rice grain downstream of a zinc mineralized area in Thailand: Implications for public health. Environ. Geochem. Health 2005, 27, 501–511. [Google Scholar] [CrossRef]
- Erdem, H.; Tosun, Y.K.; Ozturk, M. Effect of cadmium-zinc interactions on growth and Cd-Zn concentration in durum and bread wheats. Fresenius Environ. Bull 2012, 21, 1046–1051. [Google Scholar]
- Naeem, A.; Saifullah; Rehman, M.Z.-U.; Akhtar, T.; Ok, Y.S.; Rengel, Z. Genetic variation in cadmium accumulation and tolerance among wheat cultivars at the seedling stage. Commun. Soil Sci. Plant Anal. 2016, 47, 554–562. [Google Scholar] [CrossRef]
- Fischer, R.A.; Maurer, R. Drought resistance in spring wheat cultivars. I. Grain yield responses. Aust. J. Agric. Res. 1978, 29, 897–912. [Google Scholar] [CrossRef]
- Giunta, F.; Motzo, R.; Deidda, M. Effect of drought on yield and yield components of durum wheat and triticale in a Mediterranean environment. Field Crop. Res. 1993, 33, 399–409. [Google Scholar] [CrossRef]
- Senapati, N.; Stratonovitch, P.; Paul, M.J.; Semenov, M. Drought tolerance during reproductive development is important for increasing wheat yield potential under climate change in Europe. J. Exp. Bot. 2019, 70, 2549–2560. [Google Scholar] [CrossRef]
- Gregorová, Z.; Kováčik, J.; Klejdus, B.; Maglovski, M.; Kuna, R.; Hauptvogel, P.; Matušíková, I. Drought-induced responses of physiology, metabolites, and PR proteins in Triticum aestivum. J. Agric. Food Chem. 2015, 63, 8125–8133. [Google Scholar] [CrossRef]
- Lan, Y.; Chawade, A.; Kuktaite, R.; Johansson, E. Climate Change Impact on Wheat Performance—Effects on vigour, plant traits and yield from early and late drought stress in diverse lines. Int. J. Mol. Sci. 2022, 23, 3333. [Google Scholar] [CrossRef]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef]
- Ehdaie, B.; Layne, A.P.; Waines, J.G. Root system plasticity to drought influences grain yield in bread wheat. Euphytica 2012, 186, 219–232. [Google Scholar] [CrossRef]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and biotic stress combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef]
- Bauddh, K.; Singh, R.P. Growth, tolerance efficiency and phytoremediation potential of Ricinus communis (L.) and Brassica juncea (L.) in salinity and drought affected cadmium contaminated soil. Ecotoxicol. Environ. Saf. 2012, 85, 13–22. [Google Scholar] [CrossRef]
- Thomas, H. Drought Resistance in Plants; Routledge: London, UK, 1997; pp. 1–42. [Google Scholar]
- Shi, G.; Xia, S.; Ye, J.; Huang, Y.; Liu, C.; Zhang, Z. PEG-simulated drought stress decreases cadmium accumulation in castor bean by altering root morphology. Environ. Exp. Bot. 2015, 111, 127–134. [Google Scholar] [CrossRef]
- Pierret, A.; Moran, C.J.; Doussan, C. Conventional detection methodology is limiting our ability to understand the roles and functions of fine roots. New Phytol. 2005, 166, 967–980. [Google Scholar] [CrossRef]
- Crestana, S.; Cesareo, R.; Mascarenhas, S. Using A computed tomography miniscanner in soil science. Soil Sci. 1986, 142, 56. [Google Scholar] [CrossRef]
- Ahmed, S.; Klassen, T.N.; Keyes, S.; Daly, M.; Jones, D.L.; Mavrogordato, M.; Sinclair, I.; Roose, T. Imaging the interaction of roots and phosphate fertiliser granules using 4D X-ray tomography. Plant Soil 2015, 401, 125–134. [Google Scholar] [CrossRef]
- Pflugfelder, D.; Metzner, R.; van Dusschoten, D.; Reichel, R.; Jahnke, S.; Koller, R. Non-invasive imaging of plant roots in different soils using magnetic resonance imaging (MRI). Plant Methods 2017, 13, 1–9. [Google Scholar] [CrossRef]
- Bačić, G.; Ratković, S. NMR studies of radial exchange and distribution of water in maize roots: The relevance of modelling of exchange kinetics. J. Exp. Bot. 1987, 38, 1284–1297. [Google Scholar] [CrossRef]
- Brown, D.P.; Pratum, T.K.; Bledsoe, C.; Ford, E.D.; Cothern, J.S.; Perry, D. Noninvasive studies of conifer roots: Nuclear magnetic resonance (NMR) imaging of Douglas-fir seedlings. Can. J. For. Res. 1991, 21, 1559–1566. [Google Scholar] [CrossRef]
- Willatt, S.T.; Struss, R.G.; Taylor, H.M. In situ Root Studies Using Neutron Radiography. Agron. J. 1978, 70, 581–586. [Google Scholar] [CrossRef]
- Furukawa, J.; Nakanishi, T.; Matsubayashi, M. Neutron radiography of a root growing in soil with vanadium. Nucl. Instrum. Methods Phys. Res. Sect. A Accel. Spectrometers Detect. Assoc. Equip. 1999, 424, 116–121. [Google Scholar] [CrossRef]
- Tötzke, C.; Kardjilov, N.; Manke, I.; Oswald, S.E. Capturing 3D water flow in rooted soil by ultra-fast neutron tomography. Sci. Rep. 2017, 7, 6192. [Google Scholar] [CrossRef]
- Zarebanadkouki, M.; Carminati, A.; Kaestner, A.; Mannes, D.; Morgano, M.; Peetermans, S.; Lehmann, E.; Trtik, P. On-the-fly neutron tomography of water transport into lupine roots. Phys. Procedia 2015, 69, 292–298. [Google Scholar] [CrossRef]
- Moradi, A.B.; Carminati, A.; Vetterlein, D.; Vontobel, P.; Lehmann, E.; Weller, U.; Hopmans, J.W.; Vogel, H.-J.; Oswald, S.E. Three-dimensional visualization and quantification of water content in the rhizosphere. New Phytol. 2011, 192, 653–663. [Google Scholar] [CrossRef]
- Mawodza, T.; Burca, G.; Casson, S.; Menon, M. Wheat root system architecture and soil moisture distribution in an aggregated soil using neutron computed tomography. Geoderma 2020, 359, 113988. [Google Scholar] [CrossRef]
- Zhang, G.; Fukami, M.; Sekimoto, H. Influence of cadmium on mineral concentrations and yield components in wheat genotypes differing in Cd tolerance at seedling stage. Field Crop. Res. 2002, 77, 93–98. [Google Scholar] [CrossRef]
- Rahman, S.U.; Xuebin, Q.; Yasin, G.; Cheng, H.; Mehmood, F.; Zain, M.; Shehzad, M.; Ahmad, M.I.; Riaz, L.; Rahim, A.; et al. Role of silicon on root morphological characters of wheat (Triticum aestivum L.) plants grown under Cd-contaminated nutrient solution. Acta Physiol. Plant. 2021, 43, 1–13. [Google Scholar] [CrossRef]
- Berkelaar, E.; Hale, B. The relationship between root morphology and cadmium accumulation in seedlings of two durum wheat cultivars. Can. J. Bot. 2000, 78, 381–387. [Google Scholar] [CrossRef]
- Khan, N.; Ahmad, I.; Singh, S.; Nazar, R. Variation in growth, photosynthesis and yield of five wheat cultivars exposed to cadmium stress. World J. Agric. Sci. 2006, 2, 223–226. [Google Scholar]
- Abbas, T.; Rizwan, M.; Ali, S.; Adrees, M.; Mahmood, A.; Zia-Ur-Rehman, M.; Ibrahim, M.; Arshad, M.; Qayyum, M.F. Biochar application increased the growth and yield and reduced cadmium in drought stressed wheat grown in an aged contaminated soil. Ecotoxicol. Environ. Saf. 2018, 148, 825–833. [Google Scholar] [CrossRef]
- Ismael, M.A.; Elyamine, A.M.; Moussa, M.G.; Cai, M.; Zhao, X.; Hu, C. Cadmium in plants: Uptake, toxicity, and its interactions with selenium fertilizers. Metallomics 2019, 11, 255–277. [Google Scholar] [CrossRef]
- Yan, B.; Nguyen, C.; Pokrovsky, O.; Candaudap, F.; Coriou, C.; Bussière, S.; Robert, T.; Cornu, J. Cadmium allocation to grains in durum wheat exposed to low Cd concentrations in hydroponics. Ecotoxicol. Environ. Saf. 2019, 184, 109592. [Google Scholar] [CrossRef]
- Karaba, A.; Dixit, S.; Greco, R.; Aharoni, A.; Trijatmiko, K.R.; Marsch-Martinez, N.; Krishnan, A.; Nataraja, K.N.; Udayakumar, M.; Pereira, A. Improvement of water use efficiency in rice by expression of HARDY, an Arabidopsis drought and salt tolerance gene. Proc. Natl. Acad. Sci. USA 2007, 104, 15270–15275. [Google Scholar] [CrossRef]
- Mega, R.; Abe, F.; Kim, J.-S.; Tsuboi, Y.; Tanaka, K.; Kobayashi, H.; Sakata, Y.; Hanada, K.; Tsujimoto, H.; Kikuchi, J.; et al. Tuning water-use efficiency and drought tolerance in wheat using abscisic acid receptors. Nat. Plants 2019, 5, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Blum, A. Effective use of water (EUW) and not water-use efficiency (WUE) is the target of crop yield improvement under drought stress. Field Crop. Res. 2009, 112, 119–123. [Google Scholar] [CrossRef]
- Sinclair, T.R.; Devi, J.; Shekoofa, A.; Choudhary, S.; Sadok, W.; Vadez, V.; Riar, M.; Rufty, T. Limited-transpiration response to high vapor pressure deficit in crop species. Plant Sci. 2017, 260, 109–118. [Google Scholar] [CrossRef]
- Chaves, M.M.; Marôco, J.P.; Pereira, J.S. Understanding plant responses to drought—From genes to the whole plant. Funct. Plant Biol. 2003, 30, 239–264. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Manivannan, P.; Wahid, A.; Farooq, M.; Al-Juburi, H.J.; Somasundaram, R.; Panneerselvam, R. Drought stress in plants: A review on morphological characteristics and pigments composition. Int. J. Agric. Biol. 2009, 11, 100–105. [Google Scholar]
- Baize, D.; Saby, N.; Deslais, W.; Bispo, A.; Feix, I. Analyses totales et pseudo-totales d’éléments en traces dans les sols. Principaux résultats et enseignements d’une collecte nationale. Étude Gest. Sols 2006, 13, 181–200. [Google Scholar]
- Zhao, X.; Luo, L.; Cao, Y.; Liu, Y.; Li, Y.; Wu, W.; Lan, Y.; Jiang, Y.; Gao, S.; Zhang, Z.; et al. Genome-wide association analysis and QTL mapping reveal the genetic control of cadmium accumulation in maize leaf. BMC Genom. 2018, 19, 91. [Google Scholar] [CrossRef]
- Yaylalı-Abanuz, G. Heavy metal contamination of surface soil around Gebze industrial area, Turkey. Microchem. J. 2011, 99, 82–92. [Google Scholar] [CrossRef]
- Stolt, P.; Asp, H.; Hultin, S. Genetic variation in wheat cadmium accumulation on soils with different cadmium concentrations. J. Agron. Crop. Sci. 2006, 192, 201–208. [Google Scholar] [CrossRef]
- Riaz, M.; Kamran, M.; Fang, Y.; Wang, Q.; Cao, H.; Yang, G.; Deng, L.; Wang, Y.; Zhou, Y.; Anastopoulos, I.; et al. Arbuscular mycorrhizal fungi-induced mitigation of heavy metal phytotoxicity in metal contaminated soils: A critical review. J. Hazard. Mater. 2020, 402, 123919. [Google Scholar] [CrossRef] [PubMed]
- Wali, M.; Fourati, E.; Hmaeid, N.; Ghabriche, R.; Poschenrieder, C.; Abdelly, C.; Ghnaya, T. NaCl alleviates Cd toxicity by changing its chemical forms of accumulation in the halophyte Sesuvium portulacastrum. Environ. Sci. Pollut. Res. 2015, 22, 10769–10777. [Google Scholar] [CrossRef]
- Liu, K.; Lv, J.; He, W.; Zhang, H.; Cao, Y.; Dai, Y. Major factors influencing cadmium uptake from the soil into wheat plants. Ecotoxicol. Environ. Saf. 2015, 113, 207–213. [Google Scholar] [CrossRef]
- Peijnenburg, W.J.; Zablotskaja, M.; Vijver, M.G. Monitoring metals in terrestrial environments within a bioavailability framework and a focus on soil extraction. Ecotoxicol. Environ. Saf. 2007, 67, 163–179. [Google Scholar] [CrossRef] [PubMed]
- Fairbrother, A.; Wenstel, R.; Sappington, K.; Wood, W. Framework for metals risk assessment. Ecotoxicol. Environ. Saf. 2007, 68, 145–227. [Google Scholar] [CrossRef]
- Hussain, B.; Ashraf, M.N.; Rahman, S.U.; Abbas, A.; Li, J.; Farooq, M. Cadmium stress in paddy fields: Effects of soil conditions and remediation strategies. Sci. Total. Environ. 2020, 754, 142188. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Jiang, Y.; Tan, Y.; Meng, X.; Sun, H.; Wang, N. Co-transport of graphene oxide and heavy metal ions in surface-modified porous media. Chemosphere 2018, 218, 1–13. [Google Scholar] [CrossRef]
- Uddin, M.K. A review on the adsorption of heavy metals by clay minerals, with special focus on the past decade. Chem. Eng. J. 2017, 308, 438–462. [Google Scholar] [CrossRef]
- Sarwar, N.; Saifullah; Malhi, S.S.; Zia, M.H.; Naeem, A.; Bibi, S.; Farid, G. Role of mineral nutrition in minimizing cadmium accumulation by plants. J. Sci. Food Agric. 2010, 90, 925–937. [Google Scholar] [CrossRef]
- Sebastian, A.; Prasad, M.N.V. Cadmium minimization in rice. A review. Agron. Sustain. Dev. 2013, 34, 155–173. [Google Scholar] [CrossRef]
- Guo, X.; Zhang, S.; Shan, X.; Luo, L.; Pei, Z.; Zhu, Y.; Liu, T.; Xie, Y.; Gault, A. Characterization of Pb, Cu, and Cd adsorption on particulate organic matter in soil. Environ. Toxicol. Chem. 2006, 25, 2366–2373. [Google Scholar] [CrossRef]
- Weng, L.; Temminghoff, E.J.M.; Lofts, S.; Tipping, E.; Van Riemsdijk, W.H. Complexation with dissolved organic matter and solubility control of heavy metals in a sandy soil. Environ. Sci. Technol. 2002, 36, 4804–4810. [Google Scholar] [CrossRef]
- Zeng, F.; Ali, S.; Zhang, H.; Ouyang, Y.; Qiu, B.; Wu, F.; Zhang, G. The influence of pH and organic matter content in paddy soil on heavy metal availability and their uptake by rice plants. Environ. Pollut. 2011, 159, 84–91. [Google Scholar] [CrossRef]
- Shaghaleh, H.; Azhar, M.; Zia-ur-Rehman, M.; Hamoud, Y.A.; Hamad, A.A.A.; Usman, M.; Rizwan, M.; Yong, J.W.H.; Alharby, H.F.; Al-Ghamdi, A.J.; et al. Effects of agro based organic amendments on growth and cadmium uptake in wheat and rice crops irrigated with raw city effluents: Three years field study. Environ. Pollut. 2024, 344, 123365. [Google Scholar] [CrossRef]
- McCauley, A.; Jones, C.; Jacobsen, J. Soil pH and organic matter. Nutr. Manag. Modul. 2009, 8, 1–12. [Google Scholar]
- Almås, A.R.; Singh, B. Plant uptake of cadmium-109 and zinc-65 at different temperature and organic matter levels. J. Environ. Qual. 2001, 30, 869–877. [Google Scholar] [CrossRef]
- Sharma, R.K.; Archana, G. Cadmium minimization in food crops by cadmium resistant plant growth promoting rhizobacteria. Appl. Soil Ecol. 2016, 107, 66–78. [Google Scholar] [CrossRef]
- Liu, C.; Yu, R.; Shi, G. Effects of drought on the accumulation and redistribution of cadmium in peanuts at different developmental stages. Arch. Agron. Soil Sci. 2016, 63, 1049–1057. [Google Scholar] [CrossRef]
- Xia, S.; Wang, X.; Su, G.; Shi, G. Effects of drought on cadmium accumulation in peanuts grown in a contaminated calcareous soil. Environ. Sci. Pollut. Res. 2015, 22, 18707–18717. [Google Scholar] [CrossRef] [PubMed]
- Tracy, S.R.; Black, C.R.; Roberts, J.A.; Sturrock, C.; Mairhofer, S.; Craigon, J.; Mooney, S.J. Quantifying the impact of soil compaction on root system architecture in tomato (Solanum lycopersicum) by X-ray micro-computed tomography. Ann. Bot. 2012, 110, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Popova, L.; van Dusschoten, D.; Nagel, K.A.; Fiorani, F.; Mazzolai, B. Plant root tortuosity: An indicator of root path formation in soil with different composition and density. Ann. Bot. 2016, 118, 685–698. [Google Scholar] [CrossRef]
- Atkinson, J.A.; Hawkesford, M.J.; Whalley, W.R.; Zhou, H.; Mooney, S.J. Soil strength influences wheat root interactions with soil macropores. Plant Cell Environ. 2019, 43, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Materechera, S.A.; Dexter, A.R.; Alston, A.M. Penetration of very strong soils by seedling roots of different plant species. Plant Soil 1991, 135, 31–41. [Google Scholar] [CrossRef]
- Watt, M.; McCully, M.E.; Kirkegaard, J.A. Soil strength and rate of root elongation alter the accumulation of Pseudomonas spp. and other bacteria in the rhizosphere of wheat. Funct. Plant Biol. 2003, 30, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Clark, L.; Whalley, W.; Barraclough, P. How do roots penetrate strong soil? In Roots: The Dynamic Interface between Plants and the Earth; Springer: Berlin/Heidelberg, Germany, 2003; pp. 93–104. [Google Scholar]
- Shafi, M.; Zhang, G.; Bakht, J.; Khan, M.A.; Islam, U.; Khan, M.D.; Raziuddin, G. Effect of cadmium and salinity stresses on root morphology of wheat. Pak. J. Bot. 2010, 42, 2747–2754. [Google Scholar]
- Mawodza, T.; Menon, M.; Brooks, H.; Magdysyuk, O.V.; Burca, G.; Casson, S. Preferential wheat (Triticum aestivum L. cv. Fielder) root growth in different sized aggregates. Soil Tillage Res. 2021, 212, 105054. [Google Scholar] [CrossRef]
- Burca, G.; Kockelmann, W.; James, J.; Fitzpatrick, M. Modelling of an imaging beamline at the ISIS pulsed neutron source. J. Instrum. 2013, 8, P10001. [Google Scholar] [CrossRef]
- Hussain, A.; Larsson, H.; Kuktaite, R.; Johansson, E. Concentration of some heavy metals in organically grown primitive, old and modern wheat genotypes: Implications for human health. J. Environ. Sci. Health Part B 2012, 47, 751–758. [Google Scholar] [CrossRef]
- Kornfält, K.; Andersson, M.; Daniel, E.; Persson, M. Kadmium i marken i sydöstra Skåne. In SGU Rapporter Och Meddelanden; Sveriges Geologiska Undersökning: Uppsala, Sweden, 1996. [Google Scholar]
- Yong, J.W.H.; Letham, D.S.; Wong, S.C.; Farquhar, G.D. Effects of root restriction on growth and associated cytokinin levels in cotton (Gossypium hirsutum). Funct. Plant Biol. 2010, 37, 974–984. [Google Scholar] [CrossRef]
- Field, C.; Merino, J.; Mooney, H.A. Compromises between water-use efficiency and nitrogen-use efficiency in five species of California evergreens. Oecologia 1983, 60, 384–389. [Google Scholar] [CrossRef]
- Yong, J.W.H.; Ng, Y.F.; Tan, S.N.; Chew, A.Y.L. Effect of fertilizer application on photosynthesis and oil yield of Jatropha curcas L. Photosynthetica 2010, 48, 208–218. [Google Scholar] [CrossRef]
- Wang, F.; Gao, J.; Yong, J.W.H.; Wang, Q.; Ma, J.; He, X. Higher atmospheric CO2 levels favor C3 plants over C4 plants in utilizing ammonium as a nitrogen Source. Front. Plant Sci. 2020, 11, 537443. [Google Scholar] [CrossRef]
- Burca, G.; Nagella, S.; Clark, T.; Tasev, D.; Rahman, I.; Garwood, R.; Spencer, A.; Turner, M.; Kelleher, J. Exploring the potential of neutron imaging for life sciences on IMAT. J. Microsc. 2018, 272, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Mawodza, T. Plant-Soil Interactions: The Impact of Plant Water Use Efficiency on Root Architecture and Soil Structure. PhD. Thesis, University of Sheffield, Sheffield, UK, 2019. [Google Scholar]
- Dierick, M.; Masschaele, B.; Van Hoorebeke, L. Octopus, a fast and user-friendly tomographic reconstruction package developed in LabView®. Meas. Sci. Technol. 2004, 15, 1366. [Google Scholar] [CrossRef]
- Vlassenbroeck, J.; Masschaele, B.; Cnudde, V.; Dierick, M.; Pieters, K.; Van Hoorebeke, L.; Jacobs, P. Octopus 8: A high performance tomographic reconstruction package for X-ray tube and synchrotron micro-CT. In Advances in X-ray Tomography for Geomaterials; Wiley: Hoboken, NJ, USA, 2006; pp. 167–173. [Google Scholar] [CrossRef]
- Team, R. RStudio: Integrated Development for R; RStudio. Inc.: Boston, MA, USA, 2015; p. 700. [Google Scholar]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lan, Y.; Burca, G.; Yong, J.W.H.; Johansson, E.; Kuktaite, R. New Insights into the Bio-Chemical Changes in Wheat Induced by Cd and Drought: What Can We Learn on Cd Stress Using Neutron Imaging? Plants 2024, 13, 554. https://doi.org/10.3390/plants13040554
Lan Y, Burca G, Yong JWH, Johansson E, Kuktaite R. New Insights into the Bio-Chemical Changes in Wheat Induced by Cd and Drought: What Can We Learn on Cd Stress Using Neutron Imaging? Plants. 2024; 13(4):554. https://doi.org/10.3390/plants13040554
Chicago/Turabian StyleLan, Yuzhou, Genoveva Burca, Jean Wan Hong Yong, Eva Johansson, and Ramune Kuktaite. 2024. "New Insights into the Bio-Chemical Changes in Wheat Induced by Cd and Drought: What Can We Learn on Cd Stress Using Neutron Imaging?" Plants 13, no. 4: 554. https://doi.org/10.3390/plants13040554