
Effects of Co-Inoculating Saccharomyces spp. with Bradyrhizobium japonicum on Atmospheric Nitrogen Fixation in Soybeans (Glycine max (L.))

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
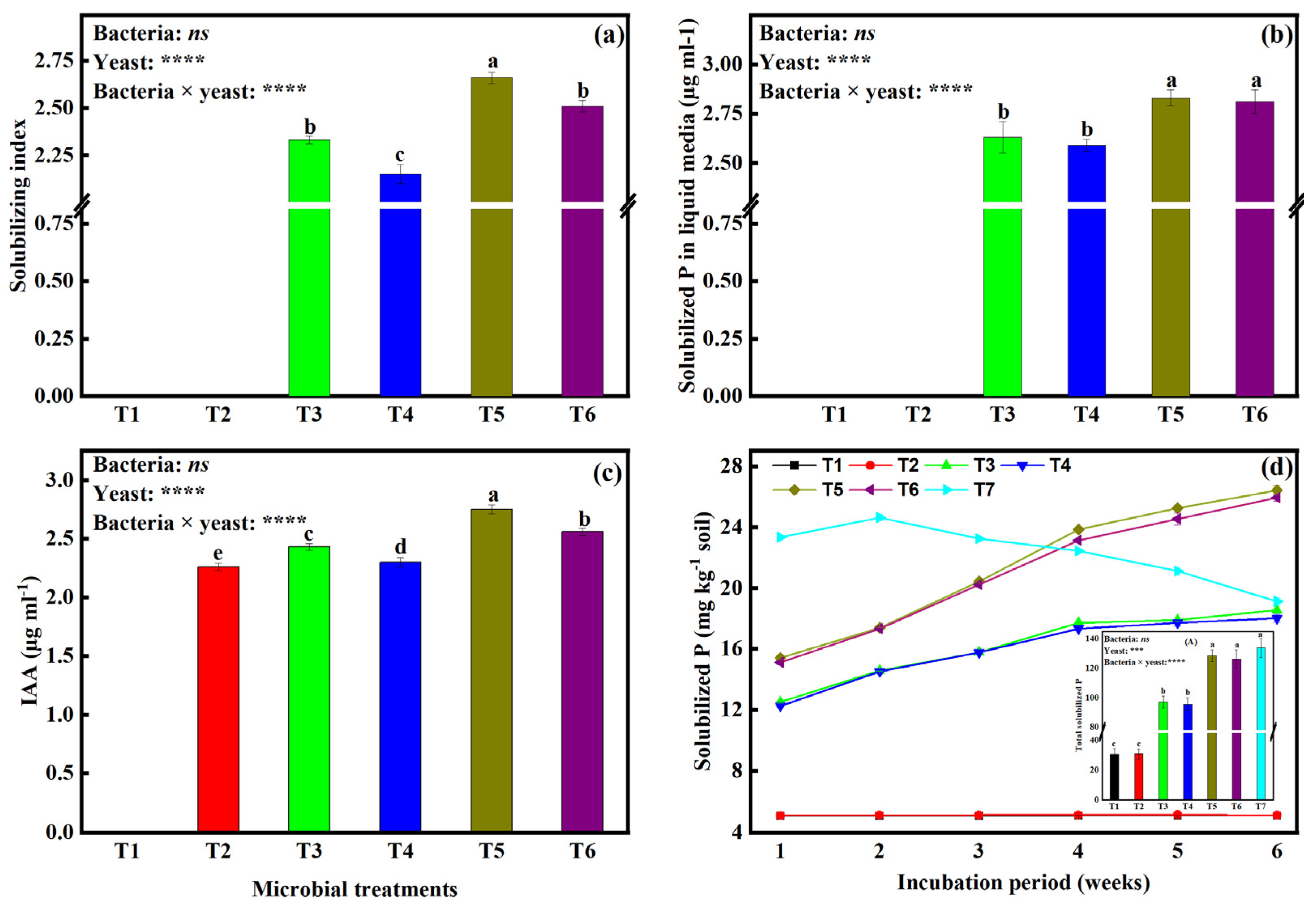
2.1. Inorganic Phosphate Solubilization and Solubilization Index by the Microbial Isolates
2.2. In Vitro Determination of IAA Production in Liquid Media by Different Microbial Isolates
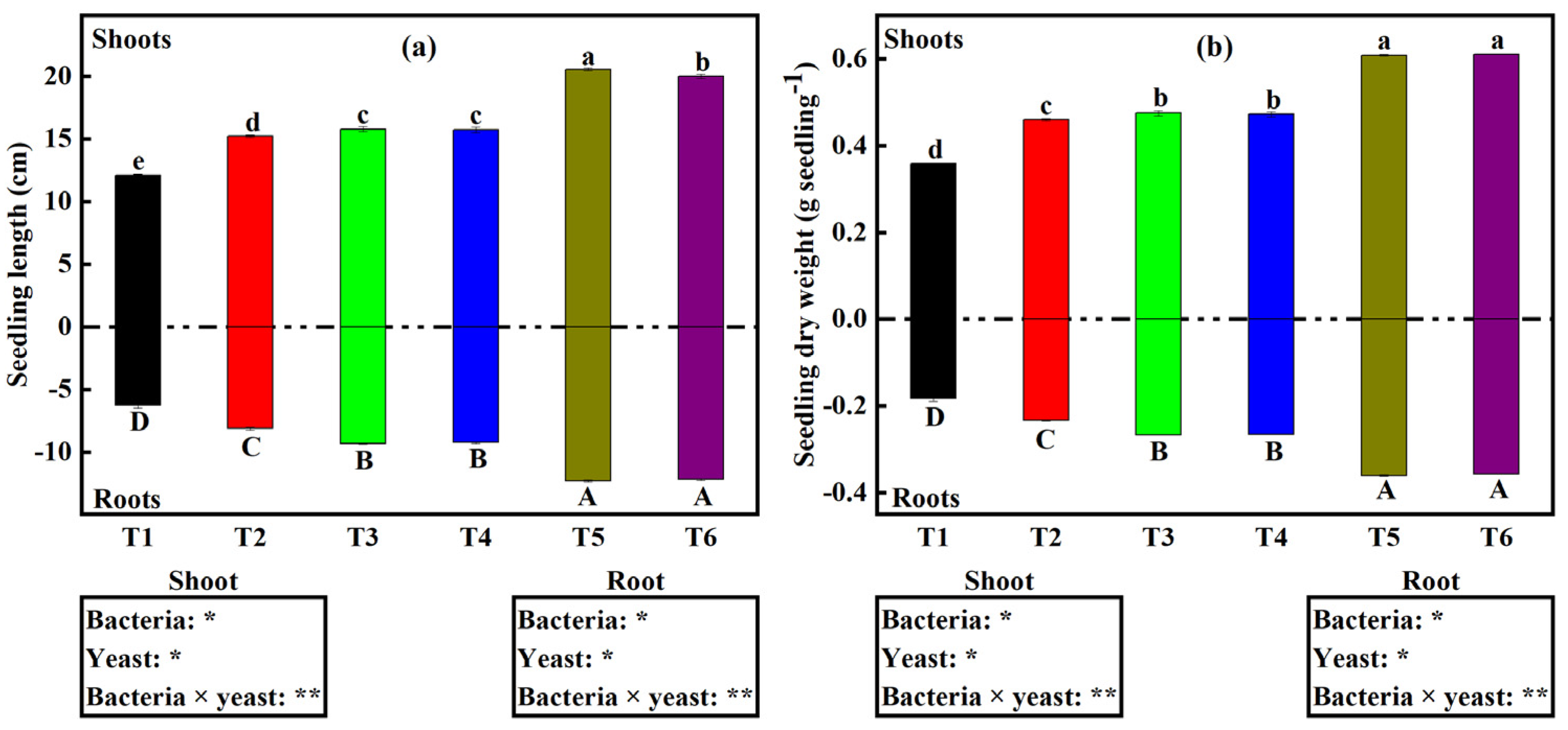
2.3. Soybean Seedling Growth and Development under Different Microbial Treatments
2.4. Soybean Root-Nodulation-Related Traits under Different Microbial Treatments
2.5. Atmospheric N2-Fixation in Soybeans under Different Microbial Treatments
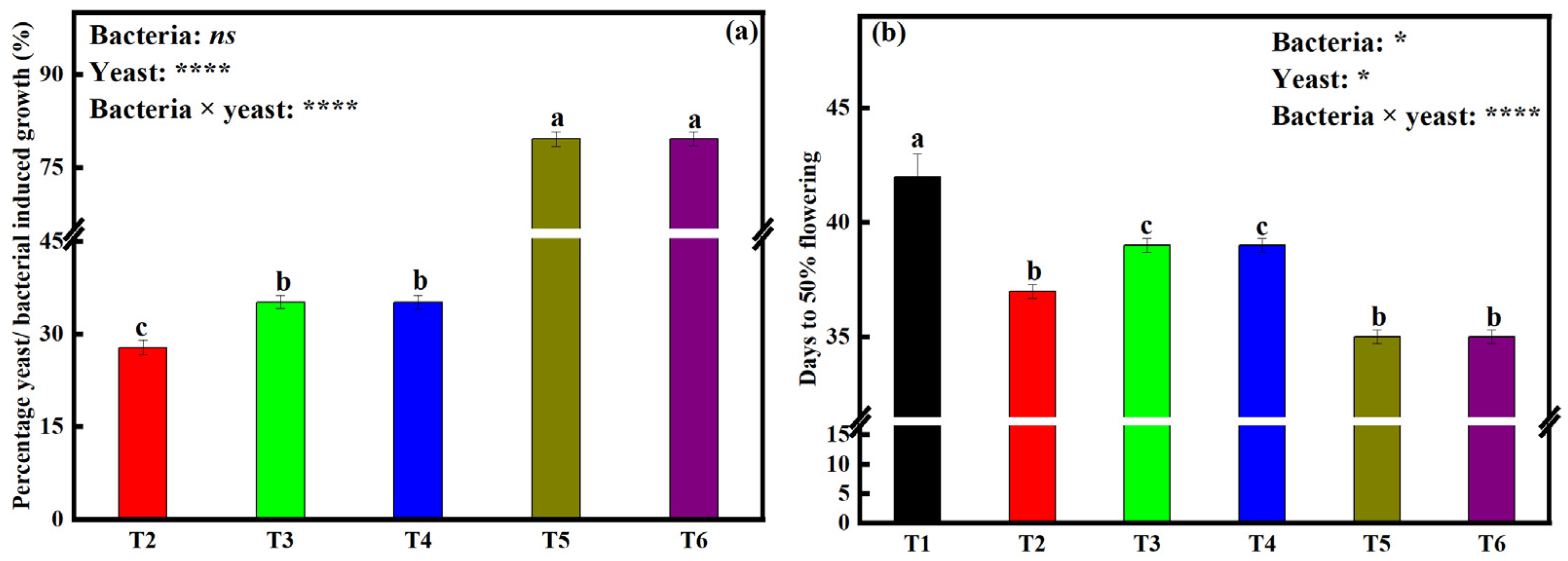
2.6. Effects of Bacterial and Yeast Inoculation on Time Taken to Reach 50% Flowering
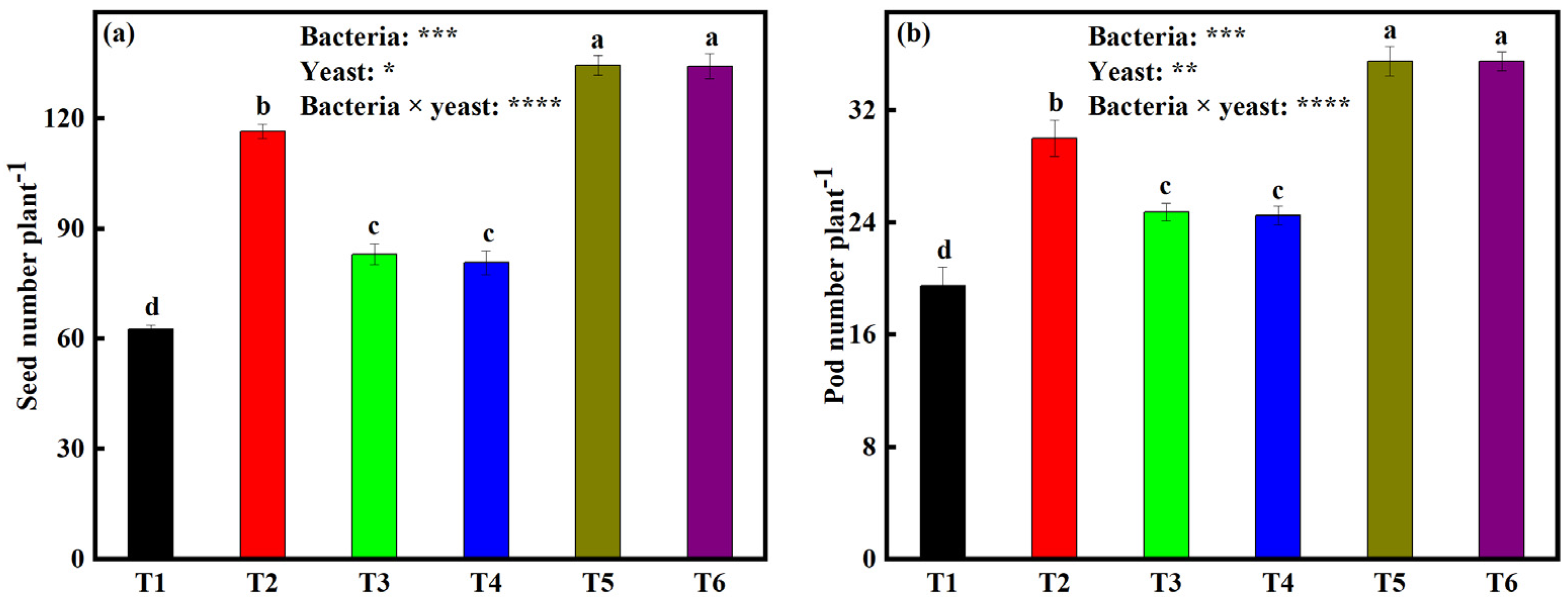
2.7. Effects of Bacterial and Yeast Inoculation on Seed Number per Plant (SNP) and Pod Number per Plant (PNP)
2.8. Straw and Seed N and P Concentrations under Different Microbial Treatments
2.9. Influence of Bacteria and Yeast Inoculation on Seed N and P Concentrations, and Rhizosphere Available P Concentration
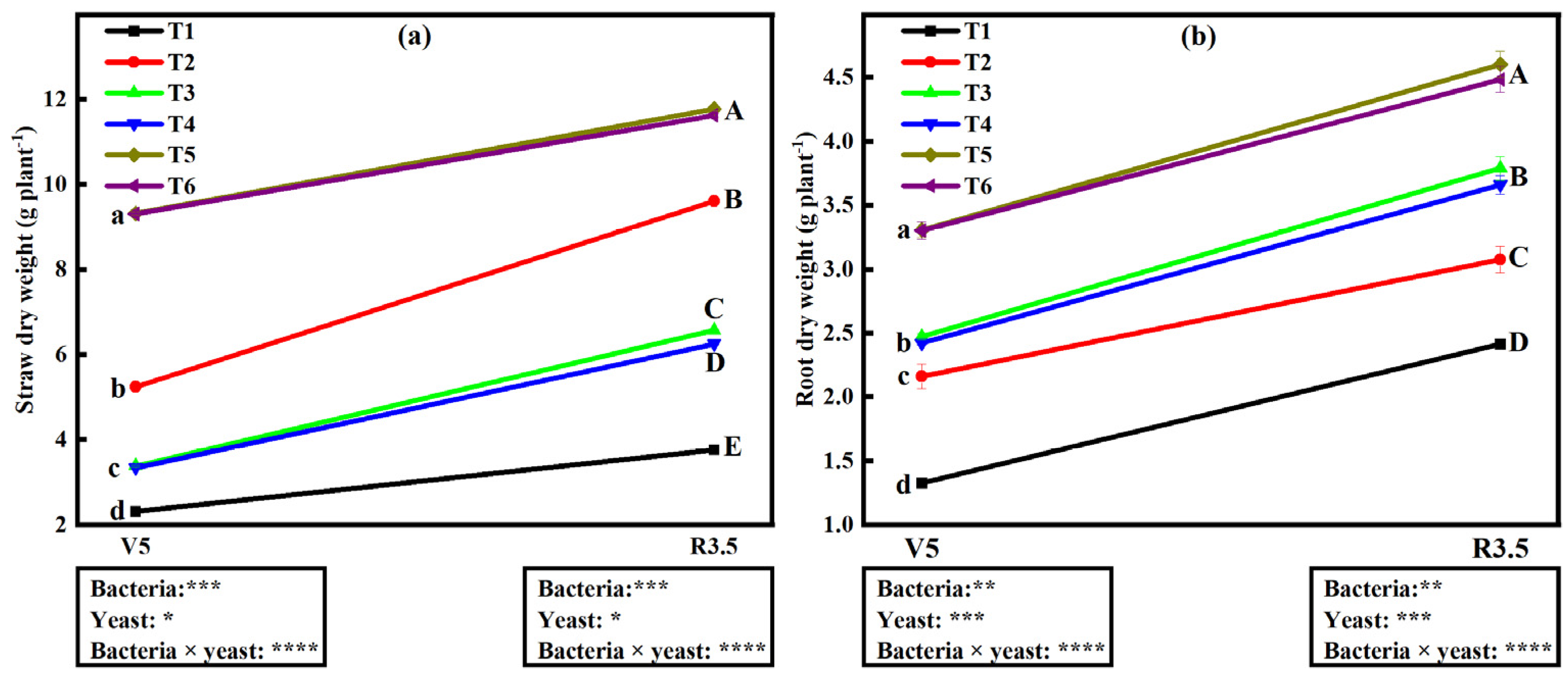
2.10. Influence of Bacterial and Yeast Inoculation on Straw Dry Weight (SDW)
2.11. Influence of Bacterial and Yeast Inoculation on Root Dry Weight (RDW)
2.12. Influence of Bacteria and Yeast Inoculation on Rhizosphere Soil Available N and P Concentration
3. Discussion
3.1. Co-Culturing of Bacteria and Yeast Enhances Inorganic P Solubilization and IAA Production by Yeast
3.2. Dual Inoculation of Bacteria and Yeast Enhances Soybean Seedling Establishment
3.3. Dual Inoculation of Bacteria and Yeast Enhances Nodule Number per Plant, Nodule Dry Weight, and N-Fixation in Soybean Plants
3.4. Bacteria and Yeast Dual Inoculation Shortens the Time Taken to Reach 50% Flowering by Soybean
3.5. Bacteria and Yeast Dual Inoculation on Soybeans Enhances Yield
3.5.1. Root and Shoot Dry Weight
3.5.2. Soybean Seed Yield
3.6. Dual Inoculation Enhances N and P Accumulation in Soybean Straw and Seed, and Rhizosphere Available P Concentrations
4. Materials and Methods
4.1. Materials Used and Culture Conditions
4.2. Experimental Design
4.3. Phosphate Solubilization under In Vitro Conditions
4.3.1. Liquid Culture Media
4.3.2. Soil Culture Media
4.3.3. Determination of the Solubilization Index
4.3.4. IAA Production by the Microbial Strains
4.3.5. Soil Characterization
4.3.6. Inoculum Preparation and Seed Treatment
4.4. Plant Growth Assessment
4.4.1. Root Elongation Measurements at Vc
4.4.2. Nodule Sampling and Determination of Percentage Nitrogen Fixed from the Atmosphere
(R3.5 = 0.94) vegetative and flowering stages,
(R3.5 = 0.94) plants in pod fill.
4.5. Plant Material Elemental Analysis
4.6. Pod Number per Plant
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Koester, R.P.; Skoneczka, J.A.; Cary, T.R.; Diers, B.W.; Ainsworth, E.A. Historical gains in soybean (Glycine max Merr.) seed yield are driven by linear increases in light interception, energy conversion, and partitioning efficiencies. J. Exp. Bot. 2014, 65, 3311–3321. [Google Scholar] [CrossRef]
- Balboa, G.R.; Sadras, V.O.; Ciampitti, I.A. Shifts in soybean yield, nutrient uptake, and nutrient stoichiometry: A historical synthesis-analysis. Crop. Sci. 2018, 58, 43–54. [Google Scholar] [CrossRef]
- Johan, P.D.; Ahmed, O.H.; Omar, L.; Hasbullah, N.A. Phosphorus transformation in soils following co-application of charcoal and wood ash. Agronomy 2021, 11, 2010. [Google Scholar] [CrossRef]
- Ikhajiagbe, B.; Anoliefo, G.O.; Olise, O.F.; Rackelmann, F.; Sommer, M.; Adekunle, I.J. Major phosphorus in soils is unavailable, yet critical for plant development. Not. Sci. Biol. 2020, 12, 500–535. [Google Scholar] [CrossRef]
- Chaudhary, M.I.; Fujita, K. Comparison of phosphorus deficiency effects on the growth parameters of mashbean, mungbean, and soybean. Soil Sci. Plant Nutr. 2012, 44, 19–30. [Google Scholar] [CrossRef]
- Kalayu, G. Phosphate solubilizing microorganisms: Promising approach as biofertilizers. Int. J. Agron. Agric. Res. 2019, 2019, 7. [Google Scholar] [CrossRef]
- Chiluwal, A. Late-season nitrogen applications increase soybean yield and seed protein concentration. Front. Plant Sci. 2021, 12, 2030. [Google Scholar] [CrossRef]
- Onor, I.O.; Onor Junior, G.I.; Kambhampati, M.S. Ecophysiological effects of nitrogen on soybean [Glycine max (L.) Merr.]. Open J. Soil Sci. 2014, 4, 357–365. [Google Scholar] [CrossRef]
- Gumiere, T.; Rousseau, A.N.; da Costa, D.P.; da Costa, D.P.; Cotta, S.R.; Andreote, F.D.; Andreote, F.D.; Andreote, F.D. Phosphorus source driving the soil microbial interactions and improving sugarcane development. Sci. Rep. 2019, 9, 4400. [Google Scholar] [CrossRef]
- Senthilkumar, M.; Amaresan, N.; Sankaranarayanan, A. Isolation of mineral phosphate solubilizing microbes from rhizosphere soil. In Plant-Microbe Interactions; Humana: New York, NY, USA, 2021; pp. 55–57. [Google Scholar]
- Ameen, F.; AlYahya, S.A.; AlNadhari, S.; Alasmari, H.; Alhoshani, F.; Wainwright, M. Phosphate solubilizing bacteria and fungi in desert soils: Species, limitations and mechanisms. Arch. Agron. Soil Sci. 2019, 65, 1446–1459. [Google Scholar] [CrossRef]
- Sharma, S.B.; Sayyed, R.Z.; Trivedi, M.H.; Gobi, T.A. Phosphate solubilizing microbes: Sustainable approach for managing phosphorus deficiency in agricultural soils. Springerplus 2013, 2, 587. [Google Scholar] [CrossRef] [PubMed]
- Menezes-Blackburn, D.; Giles, C.; Darch, T.; George, T.S.; Blackwell, M.; Stutter, M.; Shand, C.; Lumsdon, D.; Cooper, P.; Wendler, R.; et al. Opportunities for mobilizing recalcitrant phosphorus from agricultural soils: A review. Plant Soil 2018, 427, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Kucey, R.M.N.; Janzen, H.H.; Leggett, M.E. Microbially mediated increases in plant-available phosphorus. Adv. Agron. 1989, 42, 199–228. [Google Scholar]
- de Borja Reis, A.F.; Rosso, L.M.; Purcell, L.C.; Naeve, S.; Casteel, S.N.; Kovács, P.; Archontoulis, S.; Davidson, D.; Ciampitti, I.A. Environmental factors associated with nitrogen fixation prediction in soybean. Front. Plant Sci. 2021, 12, 1013. [Google Scholar] [CrossRef] [PubMed]
- Campo, R.J.; Araujo, R.S.; Hungria, M. Molybdenum-enriched soybean seeds enhance N accumulation, seed yield, and seed protein content in Brazil. Field Crops Res. 2009, 110, 219–224. [Google Scholar] [CrossRef]
- Hungria, M.; Vargas, M.A.T. Environmental factors affecting N2 fixation in grain legumes in the tropics, with an emphasis on Brazil. Field Crops Res. 2000, 65, 151–164. [Google Scholar] [CrossRef]
- Zahran, H.H. Rhizobium-legume symbiosis and nitrogen fixation under severe conditions and in an arid climate. Microbiol. Mol. Biol. Rev. 1999, 63, 968–989. [Google Scholar] [CrossRef]
- Podar, D.; Maathuis, F.J.M. Primary nutrient sensors in plants. iScience 2022, 25, 104029. [Google Scholar] [CrossRef]
- Marinkovic, J.; Bjelic, D.; Tintor, B.; Miladinovic, J.; Dukic, V.; Dordevic, V. Effects of soybean co-inoculation with plant growth promoting rhizobacteria in field trial. Rom. Biotechnol. Lett. 2018, 23, 13401–13408. [Google Scholar]
- Vandana, H.M.; Sharma, S.; Pandey, R. Phosphorus nutrition: Plant growth in response to deficiency and excess. In Plant Nutrients and Abiotic Stress Tolerance; Springer: Singapore, 2018; pp. 171–190. [Google Scholar]
- Bender, F.R.; Nagamatsu, S.T.; Delamuta, J.R.M.; Ribeiro, R.A.; Nogueira, M.A.; Hungria, M. Genetic variation in symbiotic islands of natural variant strains of soybean Bradyrhizobium japonicum and Bradyrhizobium diazoefficiens differing in competitiveness and in the efficiency of nitrogen fixation. Microb. Genom. 2022, 8, 4. [Google Scholar] [CrossRef]
- Freire, J.R.J. Important limiting factors in soil for the Rhizobium-legume symbiosis. In Biological Nitrogen Fixation: Ecology, Technology, and Physiology; Alexander, M., Ed.; Plenum Publishing Corp.: New York, NY, USA, 1984; pp. 51–74. [Google Scholar]
- Herridge, D.F.; Peoples, M.B. Timing of xylem sampling for ureide analysis of nitrogen fixation. Plant Soil 2001, 238, 57–67. [Google Scholar] [CrossRef]
- de Lima Palermo, G.C.; Coutouné, N.; Bueno, J.G.R.; Maciel, L.F.; dos Santos, L.V. Exploring metal ion metabolisms to improve xylose fermentation in Saccharomyces cerevisiae. Microb. Biotechnol. 2021, 14, 2101. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Fernández, M.; Cordero-Bueso, G.; Ruiz-Muñoz, M.; Cantoral, J.M. Culturable yeasts as biofertilizers and biopesticides for a sustainable agriculture: A comprehensive review. Plants 2021, 10, 822. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Chi, R.; Pan, X.; Liu, F.; He, J. Rock phosphate solubilization by four yeast strains. Ann. Microbiol. 2013, 63, 173–178. [Google Scholar] [CrossRef]
- Amprayn, K.O.; Rose, M.T.; Kecskés, M.; Pereg, L.; Nguyen, H.T.; Kennedy, I.R. Plant growth promoting characteristics of soil yeast (Candida tropicalis HY) and its effectiveness for promoting rice growth. Appl. Soil Ecol. 2019, 61, 295–299. [Google Scholar] [CrossRef]
- Kim, Y.C.; Leveau, J.; McSpadden Gardener, B.B.; Pierson, E.A.; Pierson, L.S.; Ryu, C.-M. The multifactorial basis for plant health promotion by plant-associated bacteria. Appl. Environ. Microbiol. 2011, 77, 1548–1555. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, L.; Zhang, Z.; Yang, A.; Liu, D. Effect of phosphorus supply levels on nodule nitrogen fixation and nitrogen accumulation in soybean (Glycine max L.). Agronomy 2022, 12, 2802. [Google Scholar] [CrossRef]
- Li, H.; Wang, X.; Liang, Q.; Lyu, X.; Li, S.; Gong, Z.; Dong, S.; Yan, C.; Ma, C. Regulation of phosphorus supply on nodulation and nitrogen fixation in soybean plants with dual-root systems. Agronomy 2021, 11, 2354. [Google Scholar] [CrossRef]
- Yao, Y.; Yuan, H.; Wu, G.; Yan, J.; Zhao, D.; Chen, S.; Kang, Q.; Ma, C.; Gong, Z. Nitrogen fixation capacity and metabolite responses to phosphorus in soybean nodules. Symbiosis 2022, 1, 21–35. [Google Scholar] [CrossRef]
- Ahmad, M.; Adil, Z.; Hussain, A.; Mumtaz, M.Z.; Nafees, M.; Ahmad, I.; Jamil, M. Potential of phosphate solubilizing Bacillus strains for improving growth and nutrient uptake in mungbean and maize crops. Pak. J. Agric. Sci. 2019, 56, 283–289. [Google Scholar]
- Shome, S.; Barman, A.; Solaiman, Z.M. Rhizobium and phosphate solubilizing bacteria influence the soil nutrient availability, growth, yield, and quality of soybean. Agriculture 2022, 12, 1136. [Google Scholar] [CrossRef]
- Mahmoud, M.H. Impact of inoculation with arbuscular mycorrhizal, phosphate solubilizing bacteria and soil yeast on growth, yield and phosphorus content of onion plants. Int. J. Soil Sci. 2015, 10, 93–99. [Google Scholar]
- Kumawat, K.C.; Sharma, P.; Nagpal, S.; Gupta, R.K.; Sirari, A.; Nair, R.M.; Bindumadhava, H.; Singh, S. Dual microbial inoculation, a game changer? Bacterial biostimulants with multifunctional growth promoting traits to mitigate salinity stress in spring mungbean. Front. Microbiol. 2021, 11, 3491. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Chen, F.; Zhao, S.; Yang, C.; Meng, Y.; Shuai, H.; Luo, X.; Dai, Y.; Yin, H.; Du, J.; et al. DA-6 promotes germination and seedling establishment from aged soybean seeds by mediating fatty acid metabolism and glycometabolism. J. Exp. Bot. 2019, 70, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Mirabal, L.; Alonso, K.D.; Eduardo, O. Spores of the mycorrhizal fungus Glomus mosseae host yeasts that solubilize phosphate and accumulate polyphosphates. Mycorrhiza 2008, 18, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Latif, H.; Mohamed, H.M. Molecular genetic identification of yeast strains isolated from Egyptian soils for solubilization of inorganic phosphates and growth promotion of corn plants. J. Microbiol. Biotechnol. 2011, 21, 55–61. [Google Scholar]
- Aloni, R.; Aloni, E.; Langhans, M.; Ullrich, C.I. Role of cytokinin and auxin in shaping root architecture: Regulating vascular differentiation, lateral root initiation, root apical dominance and root gravitropism. Ann. Bot. 2006, 97, 883–893. [Google Scholar] [CrossRef]
- Kramer-Walter, K.R.; Bellingham, P.J.; Millar, T.R.; Smissen, R.D.; Richardson, S.J.; Laughlin, D.C. Root traits are multidimensional: Specific root length is independent from root tissue density and the plant economic spectrum. J. Ecol. 2016, 104, 1299–1310. [Google Scholar] [CrossRef]
- Ghosh, P.K.; Kumar De, T.; Maiti, T.K. Production and metabolism of indole acetic acid in root nodules and symbiont (Rhizobium undicola) isolated from root nodule of aquatic medicinal legume Neptunia oleracea Lour. J. Bot. 2015, 2015, 575067. [Google Scholar]
- Desbrosses, G.J.; Stougaard, J. Root nodulation: A paradigm for how plant-microbe symbiosis influences host developmental pathways. Cell Host Microbe 2011, 10, 348–358. [Google Scholar] [CrossRef]
- Defez, R.; Andreozzi, A.; Romano, S.; Pocsfalvi, G.; Fiume, I.; Esposito, R.; Angelini, C.; Bianco, C. Bacterial IAA-delivery into Medicago root nodules triggers a balanced stimulation of C and N metabolism leading to a biomass increase. Microorganisms 2019, 7, 403. [Google Scholar] [CrossRef] [PubMed]
- Sudadi, S.; Suryono, S. Exogenous application of tryptophan and indole acetic acid (IAA) to induce root nodule formation and increase soybean yield in acid, neutral and alkaline soil. Agrivita 2015, 37, 37–44. [Google Scholar] [CrossRef]
- Torres, D.; Benavidez, I.; Donadio, F.; Mongiardini, E.; Rosas, S.; Spaepen, S.; Vanderleyden, J.; Pěnčík, A.; Novák, O.; Strnad, M.; et al. New insights into auxin metabolism in Bradyrhizobium japonicum. Res. Microbiol. 2018, 169, 313–323. [Google Scholar] [CrossRef]
- Pan, B.; Vessey, J.K.; Smith, D.L. Response of field-grown soybean to co-inoculation with the plant growth promoting rhizobacteria Serratia proteamaculans or Serratia liquefaciens, and Bradyrhizobium japonicum pre-incubated with genistein. Eur. J. Agron. 2002, 17, 143–153. [Google Scholar] [CrossRef]
- Santachiara, G.; Salvagiotti, F.; Rotundo, J.L. Nutritional and environmental effects on biological nitrogen fixation in soybean: A meta-analysis. Field Crops Res. 2019, 240, 106–115. [Google Scholar] [CrossRef]
- Taliman, N.A.; Dong, Q.; Echigo, K.; Raboy, V.; Saneoka, H. Effect of phosphorus fertilization on the growth, photosynthesis, nitrogen fixation, mineral accumulation, seed yield, and seed quality of a soybean low-phytate line. Plants 2019, 8, 119. [Google Scholar] [CrossRef] [Green Version]
- Abugalieva, S.; Didorenko, S.; Anuarbek, S.; Volkova, L.; Gerasimova, Y.; Sidorik, I.; Turuspekov, Y. Assessment of soybean flowering and seed maturation time in different latitude regions of Kazakhstan. PLoS ONE 2016, 11, e0166894. [Google Scholar] [CrossRef]
- Khan, B.; Hussain, A.; Elahi, A.; Adnan, M.; Amin, M.M.; Toor, M.D.; Aziz, A.; Sohail, M.K.; Wahab, A.; Ahmad, R. Effect of phosphorus on growth, yield and quality of soybean (Glycine max L.); A review. Int. J. Appl. Res. 2020, 6, 540–545. [Google Scholar]
- Tewari, G.P. Effects of nitrogen, phosphorus and potassium on flowering and yield of soybeans in Nigeria. Exp. Agric. 1965, 1, 185–188. [Google Scholar] [CrossRef]
- Morsy, E.M.; El-Batanony, N.; Massoud, O.N. Improvement of soybean growth and productivity by inoculation with two yeast species in new reclaimed sandy soil amended with humic acid. Afr. J. Microbiol. Res. 2014, 8, 3794–3803. [Google Scholar]
- Patterson, T.G.; LaRue, T.A. N2 fixation (C2H2) and ureide content of soybeans: Ureides as an index of fixation. Crop. Sci. 2010, 23, 825. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Chen, H.W.; Chou, J.Y. Variation in indole-3-acetic acid production by wild Saccharomyces cerevisiae and S. paradoxus strains from diverse ecological sources and its effect on growth. PLoS ONE 2016, 11, e0160524. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.F.; Fang, W.T.; Shin, L.Y.; Wei, J.Y.; Fu, S.F.; Chou, J.Y. Indole-3-acetic acid-producing yeasts in the phyllosphere of the carnivorous plant Drosera indica L. PLoS ONE 2014, 9, e114196. [Google Scholar] [CrossRef] [PubMed]
- Nosrati, R.; Owlia, P.; Saderi, H.; Rasooli, I.; Malboobi, M.A. Phosphate solubilization characteristics of efficient nitrogen fixing soil Azotobacter strains. Iran. J. Microbiol. 2014, 6, 285–295. [Google Scholar]
- Niu, Y.F.; Chai, R.S.; Jin, G.L.; Wang, H.; Tang, C.X.; Zhang, Y.S. Responses of root architecture development to low phosphorus availability: A review. Ann. Bot. 2013, 112, 391. [Google Scholar] [CrossRef]
- Kim, H.J.; Li, X. Effects of phosphorus on shoot and root growth, partitioning, and phosphorus utilization efficiency in Lantana. HortScience 2016, 51, 1001–1009. [Google Scholar] [CrossRef]
- Gaur, R.; Shani, N.; Johri, B.N.; Rossi, P.; Aragno, M. Diacetylphloroglucinol-producing pseudomonads do not influence AM fungi in wheat rhizosphere. Curr. Sci. 2004, 86, 453–457. [Google Scholar]
- Board, J.E.; Tan, Q. Assimilatory capacity effects on soybean yield components and pod number. Crop. Sci. 1995, 35, 846–851. [Google Scholar] [CrossRef]
- Vogel, J.T.; Liu, W.; Olhoft, P.; Crafts-Brandner, S.J.; Pennycooke, J.C.; Christiansen, N. Soybean yield formation physiology—A foundation for precision breeding based improvement. Front. Plant Sci. 2021, 12, 2483. [Google Scholar] [CrossRef]
- Aziz, A.L.A.; Ahiabor, B.D.K.; Opoku, A.; Abaidoo, R.C.; Bhat, R. Contributions of Rhizobium inoculants and phosphorus fertilizer to biological nitrogen fixation, growth and grain yield of three soybean varieties on a fluvic luvisol. J. Exp. Agric. Int. 2016, 10, 1–11. [Google Scholar] [CrossRef]
- Si, Z.; Yang, Q.; Liang, R.; Chen, L.; Chen, D.; Li, Y. Digalactosyldiacylglycerol synthase gene MTDGD1 plays an essential role in nodule development and nitrogen fixation. MPMI 2019, 32, 1196–1209. [Google Scholar] [CrossRef] [PubMed]
- Grennan, A.K. Phosphate accumulation in plants: Signaling. Plant Physiol. 2008, 148, 3. [Google Scholar] [CrossRef] [PubMed]
- Tagore, G.S.; Namdeo, S.L.; Sharma, S.K.; Kumar, N. Effect of Rhizobium and phosphate solubilizing bacterial inoculants on symbiotic traits, nodule leghemoglobin, and yield of chickpea genotypes. Int. J. Agron. 2013, 2013, 1–8. [Google Scholar] [CrossRef]
- Sindhu, S.S.; Sharma, R.; Sindhu, S.; Sehrawat, A. Soil fertility improvement by symbiotic rhizobia for sustainable agriculture. In Soil Fertility Management for Sustainable Development; Springer: Singapore, 2019; pp. 101–155. [Google Scholar]
- Basu, A.; Prasad, P.; Das, S.N.; Kalam, S.; Sayyed, R.Z.; Reddy, M.S.; El Enshasy, H. Plant growth promoting rhizobacteria (pgpr) as green bioinoculants: Recent developments, constraints, and prospects. Sustainability 2021, 13, 1140. [Google Scholar] [CrossRef]
- Waghmare, Y.M.; Gokhale, D.N.; Pawar, H.D. Effect of integrated nutrient management on yield, yield attributes and quality of soybean (Glycine max (L.) Merill.). J. Agric. Res. 2012, 37, 370–372. [Google Scholar]
- Jaga, P.K.; Sharma, S. Effect of bio-fertilizer and fertilizers on productivity of soybean. APSR 2015, 17, 171–174. [Google Scholar]
- Filipini, L.D.; Pilatti, F.K.; Meyer, E.; Ventura, B.S.; Lourenzi, C.R.; Lovato, P.E. Application of Azospirillum on seeds and leaves, associated with Rhizobium inoculation, increases growth and yield of common bean. Arch. Microbiol. 2021, 203, 1033–1038. [Google Scholar] [CrossRef]
- Estrada-Bonilla, G.A.; Durrer, A.; Cardoso, E.J.B.N. Use of compost and phosphate-solubilizing bacteria affect sugarcane mineral nutrition, phosphorus availability, and the soil bacterial community. Appl. Soil Ecol. 2021, 157, 103760. [Google Scholar] [CrossRef]
- Liang, J.; Fang, H.; Hao, G.; Liang, J.; Fang, H.; Hao, G. Effect of plant roots on soil nutrient distributions in shanghai urban landscapes. Am. J. Plant Sci. 2016, 7, 296–305. [Google Scholar] [CrossRef]
- Chen, B.J.W.; During, H.J.; Vermeulen, P.J.; de Kroon, H.; Poorter, H.; Anten, N.P.R. The analysis of plant root responses to nutrient concentration, soil volume and neighbour presence: Different statistical approaches reflect different underlying basic questions. Funct. Ecol. 2020, 34, 2210–2217. [Google Scholar] [CrossRef]
- Alori, E.T.; Glick, B.R.; Babalola, O.O. Microbial phosphorus solubilization and its potential for use in sustainable agriculture. Front. Microbiol. 2017, 2, 971. [Google Scholar] [CrossRef] [PubMed]
- Timofeeva, A.; Galyamova, M.; Sedykh, S. Prospects for using phosphate-solubilizing microorganisms as natural fertilizers in agriculture. Plants 2022, 11, 2119. [Google Scholar] [CrossRef] [PubMed]
- Bhodiwal, S.; Barupal, T. Phosphate Solubilizing Microbes: An Incredible Role for Plant Supplements; Springer: Berlin, Germany, 2013; Volume 2, pp. 1–14. [Google Scholar]
- Ahmad, E.; Zaidi, A.; Khan, M.S. Response of PSM inoculation to certain legumes and cereal crops. In Phosphate Solubilizing Microorganisms: Principles and Application of Microphos Technology; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Wickerham, L.J. Taxonomy of Yeasts. Technical Bulletin of the U.S. Department of Agriculture. 1951, Volume 1029, pp. 1–55. Available online: http://www.mycobank.org/BioloMICS.aspx?TableKey=14682616000000061&Rec=19613&Fields=All (accessed on 13 October 2019).
- Galloway, D.L.; Burgess, R. Applied Mycology and Bacteriology, 3rd ed.; Leonard Hill, Ltd.: London, UK, 1940; Volume 58, p. 1939. [Google Scholar]
- Somasegaran, P.; Hoben, H.J. Handbook for Rhizobia: Methods and Legumes-Rhizobium Technology; Springer: Berlin, Germany, 1994; p. 450. [Google Scholar]
- Rao, W.; Sinha, M. Phosphate dissolving microorganisms in the soil and rhizosphere. Indian J. Agric. Sci. 1963. [Google Scholar]
- Watanabe, F.S.; Olsen, S.R. Test of an ascorbic acid method for determining phosphorus in water and NaHCO3 extracts from soil. SSSAJ 1965, 29, 677–678. [Google Scholar] [CrossRef]
- Mehlich, A.; Mehlich, A.A.-M. Mehlich 3 soil test extractant: A modification of Mehlich 2 extractant. Commun. Soil Sci. Plant Anal. 1984, 15, 1409–1416. [Google Scholar] [CrossRef]
- Premono, M.E.; Moawad, A.M. Effect of Phosphate-Solubilizing Pseudomonas Putida on the Growth of Maize and Its Survival in the Rhizosphere. 1996. Available online: https://knowledgecenter.cimmyt.org/cgi-bin/koha/opac-detail.pl?biblonumber=25521&shelfbrowse_itemnumber=19186 (accessed on 16 July 2019).
- Strzelczyk, E.; Pokojska, B.A. Production of auxins and gibberellin-like substances by mycorrhizal fungi, bacteria and actinomycetes isolated from soil and the mycorrhizosphere of pine (Pinus silvestris L.). Plant Soil 1984, 81, 185–194. [Google Scholar] [CrossRef]
- Tang, Y.W.; Bonner, J. The enzymatic inactivation of indoleacetic acid; some characteristics of the enzyme contained in pea seedlings. Arch. Biochem. 2019, 13, 11–25. [Google Scholar]
- Anderson, J.M. Tropical Soil Biology and Fertility, 2nd ed.; CAB International: Wallingfold, UK, 1989; Volume 2. [Google Scholar]
- Van Reeuwijk, L. Procedures for Soil Analysis; International Soil Reference and Information Center: Wageningen, The Netherland, 2002. [Google Scholar]
- Bray, R.H.; Kurtz, L.T. Determination of total organic and available forms of phosphorus in soils. Soil Sci. 1945, 59, 39–45. [Google Scholar] [CrossRef]
- Kuo, S. Phosphorus. In Methods of Soil Analysis, Part 3: Chemical Methods; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Soltanpour, P.N., Tabatabai, M.A., Johnston, C.T., Sumner, M.E., Eds.; American Society of Agronomy: Madison, MI, USA, 1996; pp. 36–37. [Google Scholar]
- Bremner, J.M.; Mulvaney, C.S. Total nitrogen. In Methods of Soil Analysis. Part 2. Chemical and Microbiological Properties; Page, A.L., Miller, R.H., Keeny, D.R., Eds.; American Society of Agronomy; Soil Science Society of America: Madison, MI, USA, 1982; pp. 595–624. [Google Scholar]
- Okalebo, J.R.; Gathua, K.W.; Woomer, P.L. Laboratory methods of soil and plant analysis: A working manual. In Tropical Soil Biology and Fertility, 2nd ed.; SACRED Africa: Nairobi, Kenya, 2002; Volume 14. [Google Scholar]
- Jegathambigai, V.; Wijeratnam, R.S.W. Trichoderma as a seed treatment to control Helminthosporium leaf spot disease of Chrysalidocarpus lutescens. World J. Agric. Sci. 2009, 5, 720–728. [Google Scholar]
- Cloete, K.J.; Valentine, A.J.; Stander, M.A.; Blomerus, L.M.; Botha, A. Evidence of symbiosis between the soil yeast Cryptococcus laurentii and a Sclerophyllous medicinal shrub, Agathosma betulina (berg.) pillans. Microb. Ecol. 2009, 57, 624–632. [Google Scholar] [CrossRef]
- Peoples, M.B.; Faizah, A.W.; Rerkasem, B.; Herridge, D.F. Methods for evaluating nitrogen fixation by nodulated legumes in the field. ACIAR 1989, 11, 76. [Google Scholar]
- Young, G.E.; Conway, C.F. Estimation of allantoin. J. Biol. Chem. 1942, 142, 839–843. [Google Scholar] [CrossRef]
- Kamphake, L.J.; Hannah, S.A.; Cohen, J.M. Automated analysis for nitrate by hydrazine reduction. Water Res. 1967, 1, 205–216. [Google Scholar] [CrossRef]
- Herridge, D.F. Effects of nitrate and plant development on the abundance of nitrogenous solutes in root-bleeding and vacuum-extracted exudates of soybean. Crop. Sci. 1984, 24, 173. [Google Scholar] [CrossRef]
- Yemm, E.W.; Cocking, E.C.; Ricketts, R.E. The determination of amino-acids with ninhydrin. Analyst 1995, 80, 209. [Google Scholar] [CrossRef]
- Herridge, D.F.; Peoples, M.B. Ureide assay for measuring nitrogen fixation by nodulated soybean calibrated by 15N methods. Plant Physiol. 1990, 93, 495–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunnett, C.W. Pairwise multiple comparisons in the homogeneous variance, unequal sample size case. J. Am. Stat. Assoc. 1980, 75, 789–795. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Seedling Vigor Index | Specific Root Length (cm mg−1) | Germination Percentage (%) |
|---|---|---|---|
| T1 | 17.9 d ± 0.20 | 0.03 a ± 0 | 97.5 a ± 0.3 |
| T2 | 22.9 c ± 0.18 | 0.03 a ± 0 | 97.7 a ± 0.2 |
| T3 | 24.6 b ± 0.26 | 0.03 a ± 0 | 97.7 a ± 0.2 |
| T4 | 24.5 b ± 0.20 | 0.03 a ± 0 | 98 a ± 0.1 |
| T5 | 32.2 a ± 0.35 | 0.03 a ± 0 | 98 a ± 0.1 |
| T6 | 31.5 a ± 0.71 | 0.03 a ± 0 | 97.7 a ± 0.2 |
| Bacteria | ** | ns | ns |
| Yeast | *** | ns | ns |
| Bacteria × yeast | **** | ns | ns |
| Treatment | RUN% | RUN% | %Ndfa | %Ndfa |
|---|---|---|---|---|
| V5 | R3.5 | V5 | R3.5 | |
| T1 | 5.4 c ± 0.91 | 9.4 c ± 0.33 | −3.6 c ± 1.42 | −10.2 c ± 1.52 |
| T2 | 50.1 b ± 2.50 | 62.3 b ± 0.86 | 66.2 b ± 3.90 | 72.4 b ± 1.34 |
| T3 | 6.8 c ± 0.40 | 11.3 c ± 1.19 | 0 c ± 0.63 | −7.3 c ± 1.85 |
| T4 | 6.5 c ± 0.90 | 10.3 c ± 1.49 | −2 c ± 1.40 | −8.8 c ± 2.33 |
| T5 | 61.3 a ± 2.21 | 77.8 a ± 1.91 | 83.7 a ± 3.44 | 90.7 a ± 1.29 |
| T6 | 58.2 a ± 1.85 | 75 a ± 0.98 | 78.8 a ± 2.88 | 88.3 a ± 2.13 |
| Bacteria | **** | **** | **** | **** |
| Yeast | ns | ns | ns | ns |
| Bacteria × yeast | **** | **** | **** | **** |
| Treatment | Straw N Concentration (mg g−1) | Straw P Concentration (mg g−1) | Seed Nitrogen Concentration (mg g−1) | Seed P Concentration (mg g−1) |
|---|---|---|---|---|
| T1 | 28.51 d ± 3.41 | 72.91 d ± 3.21 | 50.57 d ± 3.43 | 102.91 d ± 3.21 |
| T2 | 58.73 b ± 2.16 | 80.78 c ± 2.16 | 75.83 b ± 6.15 | 114.78 c ± 2.16 |
| T3 | 38.06 c ± 2.03 | 97.23 b ± 4.38 | 64.77 c ± 3.55 | 123.23 b ± 4.38 |
| T4 | 37.26 c ± 2.63 | 95.08 b ± 3.21 | 63.42 c ± 3.12 | 121.08 b ± 3.21 |
| T5 | 69.18 a ± 2.48 | 121.66 a ± 5.59 | 85.32 a ± 1.84 | 151.66 a ± 5.59 |
| T6 | 67.24 a ± 1.97 | 128.36 a ± 4.81 | 84.98 a ± 1.17 | 148.36 a ± 4.81 |
| Bacteria | *** | * | **** | * |
| Yeast | * | ** | *** | *** |
| Bacteria × yeast | **** | **** | **** | **** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zveushe, O.K.; de Dios, V.R.; Zhang, H.; Zeng, F.; Liu, S.; Shen, S.; Kang, Q.; Zhang, Y.; Huang, M.; Sarfaraz, A.; et al. Effects of Co-Inoculating Saccharomyces spp. with Bradyrhizobium japonicum on Atmospheric Nitrogen Fixation in Soybeans (Glycine max (L.)). Plants 2023, 12, 681. https://doi.org/10.3390/plants12030681
Zveushe OK, de Dios VR, Zhang H, Zeng F, Liu S, Shen S, Kang Q, Zhang Y, Huang M, Sarfaraz A, et al. Effects of Co-Inoculating Saccharomyces spp. with Bradyrhizobium japonicum on Atmospheric Nitrogen Fixation in Soybeans (Glycine max (L.)). Plants. 2023; 12(3):681. https://doi.org/10.3390/plants12030681
Chicago/Turabian StyleZveushe, Obey Kudakwashe, Victor Resco de Dios, Hengxing Zhang, Fang Zeng, Siqin Liu, Songrong Shen, Qianlin Kang, Yazhen Zhang, Miao Huang, Ahmed Sarfaraz, and et al. 2023. "Effects of Co-Inoculating Saccharomyces spp. with Bradyrhizobium japonicum on Atmospheric Nitrogen Fixation in Soybeans (Glycine max (L.))" Plants 12, no. 3: 681. https://doi.org/10.3390/plants12030681