
The Small Auxin-Up RNA SAUR10 Is Involved in the Promotion of Seedling Growth in Rice
1
School of Life Sciences, Guizhou Normal University, Guiyang 550001, China
2
Key Laboratory of Plant Physiology and Development Regulation, Guizhou Normal University, Guiyang 550001, China
3
Laboratory of State Forestry Administration on Biodiversity Conservation in Mountainous Karst Area of Southwestern China, Guizhou Normal University, Guiyang 550001, China
4
Rice Research Institute, Guangdong Academy of Agricultural Sciences, Guangzhou 510640, China
*
Author to whom correspondence should be addressed.
Plants 2023, 12(22), 3880; https://doi.org/10.3390/plants12223880
Submission received: 23 September 2023
/
Revised: 31 October 2023
/
Accepted: 14 November 2023
/
Published: 17 November 2023
(This article belongs to the Special Issue Crop Breeding: Molecular Genetics and Genomics)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Small auxin-up-regulated RNAs (SAURs) are genes rapidly activated in response to auxin hormones, significantly affecting plant growth and development. However, there is limited information available about the specific functions of SAURs in rice due to the presence of extensive redundant genes. In this study, we found that OsSAUR10 contains a conserved downstream element in its 3′ untranslated region that causes its transcripts to be unstable, ultimately leading to the immediate degradation of the mRNA in rice. In our investigation, we discovered that OsSAUR10 is located in the plasma membrane, and its expression is regulated in a tissue-specific, developmental, and hormone-dependent manner. Additionally, we created ossaur10 mutants using the CRISPR/Cas9 method, which resulted in various developmental defects such as dwarfism, narrow internodes, reduced tillers, and lower yield. Moreover, histological observation comparing wild-type and two ossaur10 mutants revealed that OsSAUR10 was responsible for cell elongation. However, overexpression of OsSAUR10 resulted in similar phenotypes to the wild-type. Our research also indicated that OsSAUR10 plays a role in regulating the expression of two groups of genes involved in auxin biosynthesis (OsYUCCAs) and auxin polar transport (OsPINs) in rice. Thus, our findings suggest that OsSAUR10 acts as a positive plant growth regulator by contributing to auxin biosynthesis and polar transport.
1. Introduction
Small auxin-up RNA (SAUR) is specific to plants and responsive to auxin application. Several other phytohormones and environmental signals also influence SAUR expression [1]. SAUR plays a crucial role in the dynamic and adaptive growth of plants. It was first discovered in the hypocotyl of soybean, which was found to be auxin-inducible for cell elongation. Since then, SAUR has been extensively studied in Arabidopsis thaliana and other plant species [2,3].
The SAURs are a sizeable multigene family and are particularly abundant due to remarkable tandem and segmental duplications [4]. Plants contain various SAUR members that are arranged in clusters in their genomes. The number of members ranges from 3 (Anthoceros angustus) to 308 (Triticum aestivum) among the identified species to date [3,5]. For example, Arabidopsis and rice genomes contain 79 and 58 members [6,7]. They have small open reading frames varying from 180 to 540 bp, including a lack of introns, and encode proteins of 9~20 kDa. Each member has the SAUR-specific domain (SSD), which consists of 60 amino acids and confers a high degree of homology, resulting in functional redundancy [8]. However, the lower conservation and variable ranges of N- and C-terminal sequences may be required for the functional divergence among the SAUR family. It is noteworthy that the SAUR transcripts and their encoded proteins are unstable and degrade quickly after auxin induction, based on the presence of conserved downstream (DST) destabilizing elements (ATAGAT, GTA, or GAT(N)xGTA) in their 3′ untranslated region (UTR), conferring the instability of the SAUR transcripts [9]. Subcellular localization prediction revealed that the SAUR family members could be localized to the plasma membrane, cytoplasm, chloroplast, nucleus, and chloroplast [7]. For example, both OsSAUR39 and OsSAUR45 are localized to the cytoplasm, whereas AtSAUR36, AtSAUR62, and ZmSAUR2 are in the nucleus [10,11,12,13,14]. AtSAUR32 and AtSAUR53 are localized to both the plasma membrane and nucleus [15,16].
SAURs participate in a plethora of developmental processes that have been studied and validated using genetic and molecular tools such as gene editing, RNA interference, or overexpression plants. The first and most prominent function is involved in auxin-mediated cell expansion to regulate plant development by influencing auxin biosynthesis and transport differently. For example, overexpression of AtSAUR41 and AtSAUR63 resulted in pleiotropic and favorable auxin-related phenotypes, consisting of longer hypocotyls, enhanced vegetative biomass, promoted lateral root development, expanded petals, and twisted inflorescence stems [17]. Likewise, overexpression of AtSAUR19 revealed activities in inducing the growth of leaves, stems, and stamen filaments through cell wall expansion and cell elongation, achieved by interacting with the D-clade type 2c phosphatases (PP2C-Ds) protein to stimulate plasma membrane H+-ATPases’ activities, thus resulting in increasing osmotic water flow [18,19]. Similarly, AtSAUR63 was found to have an exclusive expression in root meristems and enhanced expansion by increasing auxin transport [20]. TaSAUR66-5B also promoted root growth and increased biomass and grain yields in wheat by enhancing auxin biosynthesis [5]. Defects in the genes mentioned above lead to auxin response and polar transport disruption, thus promoting cell expansion by regulating auxin transport. However, some SAURs play antagonistic roles in the process of cell expansion. For example, overexpression of AtSAUR32 inhibited hypocotyl growth of Arabidopsis [21]. OsSAUR39 negatively regulated auxin synthesis and transport, since plants with overexpression of OsSAUR39 exhibited reduced lateral root development and shoot length. Similarly, overexpression of OsSAUR45 negatively regulated growth and repressed transcripts of the auxin biosynthesis gene flavin-binding monooxygenase (OsYUCCAs) and auxin transport gene PIN-FORMED family proteins (OsPINs) [10,11,22].
The expression profiles of SAURs respond to internal or environmental cues to regulate plant development and growth. First, SAURs have distinctive expression patterns in various tissues to fine-tune plant development [23]. For instance, AtSAUR51 is expressed explicitly in the meristem, expanding leaves, root tips, and lateral root primordia; the maximum expression of AtSAUR62 is in stamen filaments, petals, sepals, stigmas, styles, pollen tubes, and pollen grains [14]; AtSAUR63 is significantly expressed in hypocotyls, petioles, cotyledons, and flowers. Moreover, AtSAUR41 is exclusively expressed in the microenvironment of the root tip as well as the hypocotyl endodermis, whereas AtSAUR71 and AtSAUR72 located in the same subbranch of the phylogenetic tree are highly expressed in the hypocotyl and the central column of young roots [17]. In addition, AtSAUR71 is also expressed in stomata, and its transcript level is altered with the stage of stomatal development [24]. Secondly, most promoters of SAURs contain multiple cis-acting regulatory elements associated with phytohormones and other signaling pathways, such as ABRE elements, MBS, W-box, and G-box regulated by abscisic acid (ABA), MYB, W-box, bZIP, bHLH, and NAC, respectively [23]. Furthermore, the expressions of AtSAUR20 and AtSAUR63 are responsive to auxin signaling in the ARF-BZR-PIF complex. AtSAUR16, AtSAUR50, and AtSAUR51 are enhanced in response to zeatin [23]. Finally, the transcript levels of many SAURs are circadian, as the DST element is related to circadian control and contributes to the rhythmic expression of SAURs. For example, AtSAUR63 is highly expressed in the morning but decreases in the afternoon through the upstream control of clock genes [25].
Despite plant SAURs being functionally characterized, most SAURs in rice remain poorly understood. Since SAURs are expressed differentially in varied tissues in rice (Oryza sativa), it is necessary to explore the functions of SAURs. In this study, we conducted knockout of OsSAUR10 using the clustered regulatory interspaced short palindromic repeats/CRISPR-associated protein 9 (CRISPR/Cas9) editing technique and overexpression of OsSAUR10. Our study suggested that OsSAUR10 might act as a positive regulator of rice growth.
2. Results
2.1. Structural and Phylogenetic Characterization of OsSAUR10
OsSAUR10 is 1451 bp in length on the second chromosome, consisting of 501 bp cDNA and 3′ UTR with a conserved DST element. OsSAUR10 contains an auxin-inducible structural domain between 26 amino acids (aa) and 112 aa. The conserved motif of the protein sequence confers its ability to be induced by auxin. The motif has also been found in its orthologs (Figure S1). We constructed a phylogenetic tree using the protein sequences from triticale, maize, rice, celery, and Arabidopsis to investigate the phylogenetic relationship of SAUR10. Based on the analysis, the phylogenetic tree was divided into four clusters: group I, II, III, and IV. Among the four clades, group IV contains multiple SAUR members (Figure S2). OsSAUR10 falls into group II, and a more detailed phylogenetic analysis with other SAUR members demonstrated that OsSAUR10 shared a close homolog with ZmSAUR54 (Figure 1A). Furthermore, the most homologous protein in Arabidopsis to OsSAUR10 is AtSAUR53, which has been reported to positively regulate apical hook development and tissue elongation in Arabidopsis [16].
To investigate the detailed subcellular location of the OsSAUR10, a GFP vector fused with OsSAUR10 was expressed transiently in rice leaf protoplasts. The results showed that the protoplasts transformed with 35S::GFP, as a positive control, displayed green fluorescent signals in the entire cell, including the membrane, cytoplasm, and nucleus under an exciting 488 nm light (Figure 1B). By contrast, when transformed with 35S::OsSAUR10:GFP, the GFP signals were only distributed in the cell membrane, precisely at the stained position using chloromethyl-benzamidodialkyl carbocyanine (CM-Dil), which was exclusively stained in the plasma membrane. Hence, we inferred that OsSAUR10 was localized in the plasma membrane.
2.2. OsSAUR10 Had a Distinctive Expression Pattern and Was Expressed in a Circadian Manner
To explore the expression pattern of OsSAUR10, various organs of Oryza sativa subsp. Japonica cv. Zhonghua11 (ZH11) at different periods were collected for qRT-PCR analysis, including seeds, seedlings 7 days after germination (DAGs), roots, leaves, sheaths, panicles at stage 3 and stage 8, and florets (Figure 2A). OsSAUR10 was higher in young seedlings, especially roots, in which levels were 14.24 times and 15.86 times higher than those expressed in the matured seeds, individually. Moreover, OsSAUR10 was highly expressed in the early developing stage of the panicle, and its transcript degraded immediately after further development, based on the fact that the transcript in Panicle 3 was 11.44 times than that in Panicle 8, suggesting that OsSAUR10 might play a crucial role in seedling development in the early stages.
Meanwhile, the promoter activity of OsSAUR10 was analyzed using the β-Glucuronidase (GUS) reporter. A total of 2000 bp promoter sequences of OsSAUR10 were amplified using primers listed in Table S1 for the GUS assay. The qRT-PCR results illustrated that strong staining signals of 7 DAGs seedlings were observed in the whole root, suggesting the accumulation of OsSAUR10 (Figure 2B), especially in root hair and elongation regions. We further magnified the root and observed staining signals in the apical meristem compared with the root cap, suggesting its predominant expression in elongating tissues of roots (Figure 2C,D). The histological observation further suggested that OsSAUR10 was strongly expressed in meristematic tissues, including parenchymal cells, primordia, and endosperms with the well-differentiated zones, indicating that OsSAUR10 might promote cell elongation (Figure 2E,F). However, some weak signals were noticed in leaves (Figure 2G) and root hairs (Figure 2H).
The expression profiles of OsSAUR10 were further investigated during the early germinal periods. Young seedlings were collected at intervals of 12 h from 0 to 120 h after seed germination (HAGs) and used to test the transcripts of OsSAUR10 (Figure 2I). OsSAUR10 transcript was significantly decreased from 0 to 24 HAGs according to the qRT-PCR analysis. It rapidly elevated and reached its peak at 60 HAGs. Interestingly, the expression was immediately reduced at 96 HAGs and subsequently enhanced again until 120 HAGs, suggesting that OsSAUR10 was expressed in a circadian manner.
2.3. OsSAUR10 Positively Responded to Varied Phytohormone Signals
The promoter of OsSAUR10 was found to reveal the potential regulated factors. Apart from the conserved sequences, the cis-acting elements of OsSAUR10 have been identified to be associated with transcriptional factors (bZIP, MYB, and AP2) (Table S2). The results also revealed that OsSAUR10 could be responsive to auxin, ABA, GA, and cytokinin (CK) signals.
We further investigated the expression of OsSAUR10 in 7 DAGs seedlings under various treatments, including auxin, polar auxin inhibitors (PAT), ABA, SA, and 6-BA. The results demonstrated that OsSAUR10 was remarkably enhanced by auxin, its analogs, or the PAT inhibitors. Among them, indole-3-acetic acid (IAA) was the conspicuous factor in influencing OsSAUR10, followed by naphthaleneacetic acid (NAA) and 2,4-dichlorophenoxyacetic acid (2,4-D). Besides auxin influx inhibitors naphthoxyacetic acid (NOA) and 3-chloro-4-hydroxyphenylacetic acid (CHPAA), the auxin efflux inhibitor 2,3,5-triiodobenzoic acid (TIBA) enhanced the transcripts of OsSAUR10 despite no significant change with auxin efflux inhibitor naphthylthalamic acid (NPA). In addition, OsSAUR10 was also significantly increased after ABA and gibberellin acid (GA3) treatments and did not differ with salicylic acid (SA) and 6-benzyladenine (6-BA) (Figure 3A). We further used a histochemical staining assay, and GUS staining could be found in the cell elongation zone of the root. After the application of IAA and ABA for five hours, more obvious staining signals were found in the same part, and no significant change was observed when the root was treated with 6-BA, indicating that IAA and ABA substantially enhanced OsSAUR10, according to the qRT-PCR results (Figure 3B).
2.4. The OsSAUR10 Mutants Impeded Seedling Growth
To evaluate the role of OsSAUR10 in rice development, a CRISPR-Cas9 construct expressing gRNA targeted to the exon of OsSAUR10 was generated and transformed into rice. Two knockout lines were obtained and proved to belong to homozygous mutant plants based on the sequencing analysis of DNA fragments using OsSAUR10 genomic-specific primers. The ossaur10#6 was produced by a guanine deletion at 192 within the exon, and the other ossaur10#15 was generated by deleting five bases from 192 to 196 (Figure 4A).
The seedlings of ossaur10 mutants displayed obvious morphological defects, such as reduced whole-plant height, decreased shoot length, narrower leaves, and smaller panicles. The whole seedlings of the two knockout lines were significantly dwarfed (Figure 4B). A comparison of four internode lengths of the stems (first, second, third, and fourth) of ossaur10 was conducted. They exhibited prominently shorter and slower growth than ZH11, especially the first internode (Figure 4C). We conducted histological observation using periodic acid–Schiff (PAS) reagent staining and found that the primordia of leaf sheaths (triangle) and shoot apical stems (asterisk) were substantially larger in wild-type compared to ossaur10 mutants, suggesting that OsSAUR10 might promote cell size and improve the growth and development of rice (Figure 4D). We gathered the 7 DAGs seedlings and analyzed their agronomic characteristics. Our results revealed that two ossaur10 mutants had a lower seedling height, 12.89% and 15.74% less than the wild-type (Figure 4E). Additionally, the root lengths of two ossaur10 mutants decreased by 17.26% and 8.59%, respectively (Figure 4F). The overall plant height and the number of tillers were also reduced by 3.59% and 26.67%, respectively (Figure 4G,J). Despite no significant change in matured seeds and seed setting rate between wild-type and two ossaur10 mutants (Figure S3 and Figure 4H), the findings revealed that the average seed germination rates of two ossaur10 mutants were only 83% and 70%, respectively, which were both lower compared to the 85% germination rate of the wild-type (Figure 4I). Therefore, it can be deduced that OsSAUR10 plays a critical positive role in seedling growth and is responsible for cell elongation.
2.5. OsSAUR10-Overexpression Displayed No Apparent Effect on Seedling Growth
We further transformed 35S::OsSAUR10 into ZH11 and obtained OsSAUR10-overexpressing plants. Two overexpression lines, OsSAUR10-ox#3 and OsSAUR10-ox#12, were compared with the wild-type and showed no significant phenotype divergences (Figure 5A). The qRT-PCR results confirmed that the OsSAUR10 transcripts in OsSAUR10-ox#3 and OsSAUR10-ox#12 were 12.84 times and 7.54 times increased in the wild-type, respectively (Figure 5B). However, we found no substantial changes between the overexpression and wild-type lines when measuring the seedling height and root length of 7 DAGs seedlings using at least fifteen plants (Figure 5C,D).
2.6. OsSAUR10 Influenced Auxin Synthesis and Transport
Since SAURs act as early response genes of auxin, they can potentially affect auxin biosynthesis and polar transport. Herein, we explored the effects of OsSAUR10 on the transcripts of the auxin synthesis OsYUCCA gene and auxin efflux carrier OsPIN gene family. The qRT-PCR showed that only expression of OsYUCCA1 among the seven family members was significantly promoted in the OsSAUR10-ox lines. However, the expressions of the other four members, OsYUCCA3, OsYUCCA4, OsYUCCA6, and OsYUCCA7, were remarkedly promoted in ossaur10 lines, despite no noticeable change in the remaining two OsYUCCAs (Figure 6A). Regarding auxin transport, the expressions of 12 OsPIN members were determined. We obtained that the transcripts of OsPIN1a, OsPIN1b, OsPIN5a, OsPIN5b, OsPIN5c, and OsPIN10b were significantly enhanced in the knockout line, whereas only OsPIN5b and OsPIN8 were increased in OsSAUR10-ox lines (Figure 6B). The results demonstrated that the up-regulation of most OsPIN members could influence the knockout of ossaur10 lines.
3. Discussion
3.1. OsSAUR10 Positively Regulates Seedling Growth in Rice
OsSAUR10 shares a common conserved core region responsive to auxin, thus positively regulating seedling growth [3]. Similarly, AtSAUR41 and AtSAUR71, in the same clade as OsSAUR10, promote cell growth and organ elongation in Arabidopsis [24]. Despite overexpression of OsSAUR39 and OsSAUR45 being reported to inhibit plant growth, here, we revealed that knockout of other SAURs resulted in the same phenotype by affecting auxin synthesis and carriers through the gene expression of OsYUCCAs and OsPINs [10,11]. Both knockout lines exhibited defective phenotypes, such as dwarf seedlings, decreased tiller number, lower germination rate, and lower yield compared with wild-type, indicating that OsSAUR10, different from the two previously reported SAUR members, was a positive regulator of rice growth and development.
The phylogenetic relationships of SAUR proteins from five species showed that four groups were clustered. OsSAUR10, situated in cluster II, displayed distant evolution from OsSAUR45 and OsSAUR39, categorized into group III. Likewise, an early study showed that OsSAUR10 was distantly separated from OsSAUR45 and OsSAUR39 among 58 members of rice [6]. Additionally, from a perspective of genomic distribution, OsSAUR10 localized on the second chromosome, whereas OsSAUR45 and OsSAUR39 were located on the ninth chromosome. Moreover, subcellular localization results showed that OsSAUR10 localized in the plasma membrane, whereas OsSAUR39 and OsSAUR45 were both in the cytoplasm, indicating that they may take part in different biological processes to be responsive to auxin signals.
3.2. Tissue-Specific and Circadian-Regulated OsSAUR10 Is Inducible by Varied Phytohormones
OsSAUR10 exhibited diverse expression profiles throughout plant development in varied tissues. An early study showed it was highly expressed in 3 DAGs seedlings [3]. OsSAUR10 was also significantly expressed in the 7 DAGs seedlings in our research, especially in the region of root elongation and panicles in the early development stage. Furthermore, the histological observation displayed strong GUS staining signals in parenchymal cells, the endodermis, and the epidermis (Figure 2), thus suggesting that tissue-specific expression of OsSAUR10 enhanced cell expansion and division to promote rice growth [23].
The 3′UTR of OsSAUR10 contains a highly conserved DST element that determines the stability of its transcripts. The putative DST element regulated the OsSAUR10 transcript circadian in the early germination stage. Moreover, two OsSAUR10-ox lines exhibited similar agronomic traits to the wild-type, possibly due to the DST element, which confer the unstable OsSAUR10 and degradation within minutes after transcription. Previous research has indicated that the overexpression of AtSAURs containing the DST element leads to phenotypes that are less severe when compared to those where the DST element is absent [13,23].
OsSAUR10 is involved in different hormone response pathways. It has been found that gibberellin treatment degrades DELLA, leading to the release of BRASSINAZOLE-RESISTANT1 (BZR1) and ARF6, which facilitates the triggering of downstream SAURs [26,27]. Our results also showed that OsSAUR10 was remarkably increased by auxin and GA3 application, and the promotion might be through the ARF-BZR-PIF signaling module [28]. In addition, its transcript was also substantially enhanced by ABA, suggesting crosstalk between auxin and ABA. Similarly, AtSAUR41 and AtSAUR32 were also ABA-inducible, strengthening the plant’s resilience to harsh environmental conditions by harnessing the potential of cell expansion modulation, including drought and salt tolerance [15,29]. The decreased seed germination and dwarf seedlings in the ossaur10 mutants may result from the altered hormones, which impede seedling development. Several reports in the past revealed the interplay between auxin and the other phytohormones, and their pathways impinged on each other. In rice, the network of inter-regulation between different hormones is very complex. Therefore, the comprehensive interactions result in defective phenotypes in the ossaur10 mutants.
3.3. OsSAUR10 Regulates Seedlings’ Growth by Impacting Auxin Production and Its Polar Transport
Auxin accumulation and distribution are closely linked to the expression of auxin synthesis genes and their transporter genes [30]. The YUCCA gene family, which catalyzes the N-oxidation of tryptamine to form N-hydroxyl tryptamine in vitro, involves embryogenesis and seedling development and is responsible for auxin biosynthesis in shoots [31]. Overexpression of AtYUCCA2 and AtYUCCA3 resulted in a similar phenotype with functional redundancy between the different YUCCA genes. The functional redundancy was further confirmed through simultaneous analysis of the AtYUCCA2, AtYUCCA4, and AtYUCCA6 genes, which were partially overlapping in their expression in the stem tip and flower of Arabidopsis, and the mutation of a single gene did not have a significant phenotype. Among seven OsYUCCAs in rice [32], OsYUCCA1 might be the dominant one that promotes auxin biosynthesis, according to the fact that overexpression of OsYUCCA1 resulted in a high-growth-hormone phenotype in rice. In contrast, the antisense gene expression increased rice growth hormone insensitivity [33]. Our study found that OsYUCCA1 was significantly promoted in the overexpression seedlings despite the enhancement of OsYUCCA3, OsYUCCA4, OsYUCCA6, and OsYUCCA7 in the ossaur10 lines. This result revealed that OsSAUR10 promoted auxin biosynthesis mainly through the increased expression of OsYUCCA1.
PINs play roles in the polarity of auxin transporter, impacting auxin homeostasis and metabolism, and also belong to a multigene family [34]. Eight members of the PIN family were identified in Arabidopsis. At the same time, different members might play different roles [35]. For example, PIN5 and PIN8 have antagonistic/compensatory activity [36]. There are 12 members of the PIN family identified in rice [37]. OsPIN1a is highly accumulated in the root base of the stem base and mid-column sheath cells, and suppression of OsPIN1a expression confers an increase in the number of tillers [38]. OsPIN2 takes part in basipetal polar auxin transport, promoting the tillers’ spreading growth [39]. The expression of OsPIN1a, OsPIN1b, OsPIN5a, OsPIN5b, OsPIN5c, and OsPIN10b was significantly up-regulated in the CRISPR lines of OsSAUR10, implying the repressive regulation effect of OsSAUR10 on some OsPINs. The increased OsPINs might contribute to the decreased number of tillers in ossaur10 mutants. In summary, we learned that the divergence of seed germination and seedling development is probably altered due to the up-regulation of OsYUCCA and varied OsPIN expression.
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
The ossaur10 seedlings produced through genome editing using the CRISRP-Cas9 and OsSAUR10-ox lines were grown in the same place as the wild-type in the following conditions: 60~70% humidity, a photoperiod of 12 h light (28 °C) /12 h dark (24 °C), and 450 μmol/m2/s light intensity. For phytohormone treatment, seeds were put on sterile gauze saturated with Yoshida’s nutrient solution and grown in darkness at 37 °C until germination. We collected 7 DAG seedlings and incubated them in a nutrition solution containing 1 μM 2,4-D, 1 μM NAA, 10 μM IAA, 1 μM 6-BA, 10 μM ABA, 1 mM SA, 10 μM GA3, and growth hormone inhibitors, including 1 μM Naphthylthalamic acid NPA, 1 μM TIBA, 1 μM NOA, and 1 mM CHPAA. After being treated for five hours, the samples were collected, frozen in liquid nitrogen immediately, and stored at −80 °C.
4.2. Phylogenetic and Promoter Analysis
We used the SAUR protein sequences from Arabidopsis (79), Oryza sativa (58), Amborella trichopoda (26), Zea mays (79), and Cercis canadensis (48) listed in Table S3 to construct a phylogenetic tree. Multiple sequence alignment was performed using ClustalW2 (https://www.ebi.ac.uk/Tools/msa/clustalw2/, accessed on 16 November 2023). The phylogenic tree was displayed using the maximum likelihood method in MEGA 11 (https://www.megasoftware.net, accessed on 16 November 2023) and bootstraps with 1000 replicates to evaluate the reliability of nodes. Then, we used the iTOL (https://itol.embl.de/, accessed on 16 November 2023) online tool to annotate and visualize the resulting tree.
The 2 kb genomic DNA sequences upstream of the transcriptional initiation site of OsSAUR10 were retrieved from the Rice Genome Annotation Project database (http://rice.uga.edu/, accessed on 16 November 2023). Afterward, the promoter sequences were submitted into the New PLACE Web database (https://www.dna.affrc.go.jp/PLACE/?action=newplace, accessed on 16 November 2023) to obtain cis-acting regulatory elements.
4.3. Subcellular Localization Assay
To analyze subcellular localization, a GFP reporter tag was constructed. The open reading frame of OsSAUR10 was amplified and fused into the N-terminal start of GFP under the control of the CaMV 35S promoter (35S::OsSAUR10:GFP) of pM999 [40]. The reconstruction vector was then introduced via polyethylene glycol (PEG)-mediated transient transformation into rice protoplasts, and fusion protein localization was monitored in the following 24 h [41]. Meanwhile, the plasma membrane was labeled with CM-Dil at a concentration of 10 μmol/L for 15~20 min. Fluorescence images were captured using the LSM710 laser scanning microscope (Carl Zeiss, Jena, Germany) with a 20 × objective, 500 gain, 488 nm excitation wavelength, and 507 nm emission wavelength.
4.4. GUS Staining
The 2 kb promoter sequence of OsSAUR10 was amplified from the rice genomic DNA using primers (Table S1), digested using BamH I and Pst I (TaKaRa, Kusatsu, Japan), and inserted into the pDX2181 binary vector containing the GUS reporter gene [42]. This vector was transformed into ZH11 plants via an Agrobacterium-mediated transformation procedure [43]. Different tissues of seedlings at 7 DAGs were collected and put into a 2 mL tube, and reaction solution containing 20 mL 100 mM sodium phosphate, pH 7.0, 0.5 mM potassium ferrocyanide, 1 mL 0.5 M EDTA, 0.5 mM potassium ferricyanide, 5 mg chloramphenicol, 50 mg X-Gluc, 0.1% Triton X-100, 10 mL methanol, and ddH2O up to 50 mL was added. After being treated for 24 h at 37 °C, X-Gluc stained tissues were decolorized in solution (glacial acetic acid: ethanol: H2O = 1:3:6) for 2 h, washed 2~3 times, and examined using a stereo microscope (SZX16, Olympus, Hamburg, Japan).
4.5. RNA Isolation and qRT-PCR Analysis
Total RNA extracted from various tissues and seedlings of wild-type and OsSAUR10-ox was reverse-transcribed into cDNA using an RT-PCR Kit® with an oligo dT-adaptor primer (TaKaRa, Kusatsu, Japan). The qRT-PCR was performed in a LightCycler480 instrument (Roche, Rotkreuz, Switzerland) with the FastStart DNA Master SYBR Green I kit. The procedure was performed at 95 °C for 5 min, 40 cycles of 95 °C for 25 s, annealing at 57 °C for 15 s, 72 °C for 25 s, and 4 °C for 10 min. To estimate the expression levels relative to the control, we calculated ΔΔCt and then analyzed the data using the 2−ΔΔCt method. We used a ubiquitin gene (LOC_Os03g13170) as an internal standard gene to normalize cDNA starting amounts. All samples were subjected to three biological repeats.
4.6. Vector Construction and Transgenic Rice Plants
A nucleotide sequence with specific gRNA was designed from the exon sequence of OsSAUR10 (LOC_Os02g30810) and transformed into a pCXUN-CAS9 backbone vector [44]. A 501 bp cDNA fragment of OsSAUR10 was amplified and inserted into the pU2301-Flag using gateway (Thermo Fisher, Waltham, MA, USA) [45]. These constructed vectors were transformed into Agrobacterium tumefaciens EHA105 and subsequently transferred into ZH11 calli as previously described [46]. Different transgenic lines were obtained, and their agronomic traits were investigated.
4.7. Histological Observation
The roots were cut into small pieces, fixed in FAA (10% formalin (v/v), 5% acetic acid, 50% ethanol) for vacuum pumping, and stained in hematoxylin for at least 48 h before use. After rinsing the samples with distilled water twice, they were dehydrated using a series of ethanol solutions (30%, 50%, 70%, 85%, and 95% for 1 h each, before being placed in 100% ethanol for 2 h). Afterward, the samples were made transparent using a series of chloroform solutions (1/5, 2/5, 3/5, 4/5, and 5/5 for 1 h each) and then impregnated with broken paraffin wax for 3 days. Finally, the samples were embedded in melted paraffin wax, and sections of samples (8 mm) were made using an RM2265 microtome (Leica, Wetzlar, Germany). In addition, shoot apex samples of wild-type and ossaur10 mutants were made for histological observation and stained with PAS reagent. After washing with distilled water three times, all slides were examined using a BX53 microscope (Olympus, Hamburg, Germany).
4.8. Statistical Analysis
The data were represented as mean ± standard deviation and compared using analysis of variance (ANOVA) followed by a comparison with Tukey’s test using the Statistical Program for Social Sciences (SPSS) program version 20.0 (SPSS Inc., Chicago, IL, USA). * denotes significant difference at p < 0.05 and (**) represents significant difference at p < 0.01.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants12223880/s1, Figure S1: Characterization and gene structure of OsSAUR10. (A) The gene and protein structures of OsSAUR10. Only one exon encodes the auxin-inducible domain (PF02519). The presence of the DST element was shown in its 3′ UTR region. (B) OsSAUR10 homologous alignment in Oryza brachyantha, Sorghum bicolor, Zea mays, Panicum miliaceum, Setaria italica, and Brachypodium distachyon. The conserved domains are shown above the same amino acids displayed in the red background; Figure S2: A phylogenetic tree of SAUR proteins from Arabidopsis, rice, triticale, maize, and celery was constructed using MEGA11 software through the maximum-likelihood method with 1000 bootstrap replicates. The proteins of the SAUR family belonging to varied groups are highlighted in different groups. Species abbreviations are as follows: At: A. thaliana; Os: O. sativa; AmTr: Amborella trichopoda; Zm: Zea mays; Cerca: Cercis canadensis; Figure S3: Phenotypes of seedlings (A–C) and matured seeds (D–I) of wild-type and two ossaur10 mutants. Bar = 2 mm; Table S1: The list of primers used in this study; Table S2: The list of conserved cis-acting elements of OsSAUR10; Table S3: The identified SAUR proteins from Arabidopsis thaliana (79), Oryza sativa (58), Amborella trichopoda (26), Zea mays (79), and Cercis canadensis (48).
Author Contributions
Methodology, X.H.; writing—original draft preparation, H.Y. and N.L.; project administration, Z.L.; funding acquisition, X.H. and L.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the National Natural Science Foundation of China (Grant No. 32160699 and 32060587), the Joint Fund of the National Natural Science Foundation of China and the Karst Science Re-search Center of Guizhou province (Grant No. U1812401), Guizhou Normal University Qian Shi Xin Miao ([2022]18), and the Province Natural Science Foundation (QKHJC-ZK [2023]ZD026).
Data Availability Statement
Data are contained within the article and Supplementary Materials.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Stortenbeker, N.; Bemer, M. The SAUR gene family: The plant’s toolbox for adaptation of growth and development. J. Exp. Bot. 2019, 70, 17–27. [Google Scholar] [CrossRef] [PubMed]
- McClure, B.A.; Guilfoyle, T. Characterization of a class of small auxin-inducible soybean polyadenylated RNAs. Plant Mol. Biol. 1987, 9, 611–623. [Google Scholar] [CrossRef]
- Zhang, H.; Yu, Z.; Yao, X.; Chen, J.; Chen, X.; Zhou, H.; Lou, Y.; Ming, F.; Jin, Y. Genome-wide identification and characterization of small auxin-up RNA (SAUR) gene family in plants: Evolution and expression profiles during normal growth and stress response. BMC Plant Biol. 2021, 21, 4. [Google Scholar] [CrossRef]
- Chen, Y.; Hao, X.; Cao, J. Small auxin upregulated RNA (SAUR) gene family in maize: Identification, evolution, and its phylogenetic comparison with Arabidopsis, rice, and sorghum. J. Integr. Plant Biol. 2014, 56, 133–150. [Google Scholar] [CrossRef]
- Lv, W.; He, X.; Guo, H.; Lan, H.; Jiao, Y.; Li, L.; Lian, Y.; Wang, Z.; Xin, Z.; Ren, Y.; et al. Genome-wide identification of tasaur gene family members in hexaploid wheat and functional characterization of TaSAUR66-5B in improving nitrogen use efficiency. Int. J. Mol. Sci. 2022, 23, 7574. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.; Tyagi, A.K.; Khurana, J.P. Genome-wide analysis, evolutionary expansion, and expression of early auxin-responsive SAUR gene family in rice (Oryza sativa). Genomics 2006, 88, 360–371. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Gray, W.M. SAUR proteins as effectors of hormonal and environmental signals in plant growth. Mol. Plant 2015, 8, 1153–1164. [Google Scholar] [CrossRef] [PubMed]
- Park, J.E.; Park, J.Y.; Kim, Y.S.; Staswick, P.E.; Jeon, J.; Yun, J.; Kim, S.Y.; Kim, J.; Lee, Y.H.; Park, C.M. GH3-mediated auxin homeostasis links growth regulation with stress adaptation response in Arabidopsis. J. Biol. Chem. 2007, 282, 10036–10046. [Google Scholar] [CrossRef]
- Sullivan, M.L.; Green, P.J. Mutational analysis of the DST element in tobacco cells and transgenic plants: Identification of residues critical for mRNA instability. RNA 1996, 2, 308–315. [Google Scholar]
- Kant, S.; Bi, Y.M.; Zhu, T.; Rothstein, S.J. SAUR39, a small auxin-up RNA gene, acts as a negative regulator of auxin synthesis and transport in rice. Plant Physiol. 2009, 151, 691–701. [Google Scholar] [CrossRef]
- Xu, Y.X.; Xiao, M.Z.; Liu, Y.; Fu, J.L.; He, Y.; Jiang, D.A. The small auxin-up RNA OsSAUR45 affects auxin synthesis and transport in rice. Plant Mol. Biol. 2017, 94, 97–107. [Google Scholar] [CrossRef]
- Knauss, S.; Rohrmeier, T.; Lehle, L. The auxin-induced maize gene ZmSAUR2 encodes a short-lived nuclear protein expressed in elongating tissues. J. Biol. Chem. 2003, 278, 23936–23943. [Google Scholar] [CrossRef] [PubMed]
- Hou, K.; Wu, W.; Gan, S.S. SAUR36, a small auxin up RNA gene, is involved in the promotion of leaf senescence in Arabidopsis. Plant Physiol. 2013, 161, 1002–1009. [Google Scholar] [CrossRef] [PubMed]
- He, S.L.; Hsieh, H.L.; Jauh, G.Y. SMALL AUXIN UP RNA62/75 are required for the translation of transcripts essential for pollen tube growth. Plant Physiol. 2018, 178, 626–640. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Liu, Y.; Li, M.; Lamin-Samu, A.T.; Yang, D.; Yu, X.; Izhar, M.; Jan, I.; Ali, M.; Lu, G. The Arabidopsis SMALL AUXIN UP RNA32 protein regulates ABA-mediated responses to drought stress. Front. Plant Sci. 2021, 12, 625493. [Google Scholar] [CrossRef]
- Kathare, P.K.; Dharmasiri, S.; Dharmasiri, N. SAUR53 regulates organ elongation and apical hook development in Arabidopsis. Plant Signal. Behav. 2018, 13, e1514896. [Google Scholar] [CrossRef]
- Kong, Y.; Zhu, Y.; Gao, C.; She, W.; Lin, W.; Chen, Y.; Han, N.; Bian, H.; Zhu, M.; Wang, J. Tissue-specific expression of SMALL AUXIN UP RNA41 differentially regulates cell expansion and root meristem patterning in Arabidopsis. Plant Cell Physiol. 2013, 54, 609–621. [Google Scholar] [CrossRef]
- Spartz, A.K.; Lee, S.H.; Wenger, J.P.; Gonzalez, N.; Itoh, H.; Inze, D.; Peer, W.A.; Murphy, A.S.; Overvoorde, P.J.; Gray, W.M. The SAUR19 subfamily of SMALL AUXIN UP RNA genes promote cell expansion. Plant J. 2012, 70, 978–990. [Google Scholar] [CrossRef]
- Spartz, A.K.; Ren, H.; Park, M.Y.; Grandt, K.N.; Lee, S.H.; Murphy, A.S.; Sussman, M.R.; Overvoorde, P.J.; Gray, W.M. SAUR Inhibition of PP2C-D phosphatases activates plasma membrane H+-ATPases to promote cell expansion in Arabidopsis. Plant Cell 2014, 26, 2129–2142. [Google Scholar] [CrossRef]
- Gastaldi, V.; Lucero, L.E.; Ferrero, L.V.; Ariel, F.D.; Gonzalez, D.H. Class-I TCP Transcription factors activate the SAUR63 Gene subfamily in gibberellin-dependent stamen filament elongation. Plant Physiol. 2020, 182, 2096–2110. [Google Scholar] [CrossRef]
- Park, J.E.; Kim, Y.S.; Yoon, H.K.; Park, C.M. Functional characterization of a small auxin-up RNA gene in apical hook development in Arabidopsis. Plant Sci. 2007, 172, 150–157. [Google Scholar] [CrossRef]
- Chae, K.; Isaacs, C.G.; Reeves, P.H.; Maloney, G.S.; Muday, G.K.; Nagpal, P.; Reed, J.W. Arabidopsis SMALL AUXIN UP RNA63 promotes hypocotyl and stamen filament elongation. Plant J. Cell Mol. Biol. 2012, 71, 684–697. [Google Scholar] [CrossRef] [PubMed]
- van Mourik, H.; van Dijk, A.D.J.; Stortenbeker, N.; Angenent, G.C.; Bemer, M. Divergent regulation of Arabidopsis SAUR genes: A focus on the SAUR10-clade. BMC Plant Biol. 2017, 17, 245. [Google Scholar] [CrossRef] [PubMed]
- Qiu, T.; Chen, Y.; Li, M.; Kong, Y.; Zhu, Y.; Han, N.; Bian, H.; Zhu, M.; Wang, J. The tissue-specific and developmentally regulated expression patterns of the SAUR41 subfamily of small auxin up RNA genes: Potential implications. Plant Signal. Behav. 2013, 8, e25283. [Google Scholar] [CrossRef]
- Nakamichi, N.; Kiba, T.; Henriques, R.; Mizuno, T.; Chua, N.H.; Sakakibara, H. PSEUDO-RESPONSE REGULATORS 9, 7, and 5 are transcriptional repressors in the Arabidopsis circadian clock. Plant Cell 2010, 22, 594–605. [Google Scholar] [CrossRef]
- Bai, M.Y.; Shang, J.X.; Oh, E.; Fan, M.; Bai, Y.; Zentella, R.; Sun, T.P.; Wang, Z.Y. Brassinosteroid, gibberellin and phytochrome impinge on a common transcription module in Arabidopsis. Nat. Cell Biol. 2012, 14, 810–817. [Google Scholar] [CrossRef]
- Favero, D.S.; Le, K.N.; Neff, M.M. Brassinosteroid signaling converges with SUPPRESSOR OF PHYTOCHROME B4-#3 to influence the expression of SMALL AUXIN UP RNA genes and hypocotyl growth. Plant J. 2017, 89, 1133–1145. [Google Scholar] [CrossRef]
- Franklin, K.A.; Lee, S.H.; Patel, D.; Kumar, S.V.; Spartz, A.K.; Gu, C.; Ye, S.; Yu, P.; Breen, G.; Cohen, J.D.; et al. Phytochrome-interacting factor 4 (PIF4) regulates auxin biosynthesis at high temperature. Proc. Natl. Acad. Sci. USA 2011, 108, 20231–20235. [Google Scholar] [CrossRef]
- Qiu, T.; Qi, M.; Ding, X.; Zheng, Y.; Zhou, T.; Chen, Y.; Han, N.; Zhu, M.; Bian, H.; Wang, J. The SAUR41 subfamily of SMALL AUXIN UP RNA genes is abscisic acid inducible to modulate cell expansion and salt tolerance in Arabidopsis thaliana seedlings. Ann. Bot. 2020, 125, 805–819. [Google Scholar] [CrossRef]
- Strader, L.C.; Zhao, Y. Auxin perception and downstream events. Curr. Opin. Plant Biol. 2016, 33, 8–14. [Google Scholar] [CrossRef]
- Ishida, J.K.; Wakatake, T.; Yoshida, S.; Takebayashi, Y.; Kasahara, H.; Wafula, E.; de Pamphilis, C.W.; Namba, S.; Shirasu, K. Local auxin biosynthesis mediated by a YUCCA flavin monooxygenase regulates haustorium development in the parasitic plant Phtheirospermum japonicum. Plant Cell 2016, 28, 1795–1814. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Kamiya, N.; Morinaka, Y.; Matsuoka, M.; Sazuka, T. Auxin biosynthesis by the YUCCA genes in rice. Plant Physiol. 2007, 143, 1362–1371. [Google Scholar] [CrossRef]
- Zhang, T.; Li, R.; Xing, J.; Yan, L.; Wang, R.; Zhao, Y. The YUCCA-Auxin-WOX11 module controls crown root development in Rice. Front. Plant Sci. 2018, 9, 523. [Google Scholar] [CrossRef] [PubMed]
- Simon, S.; Skupa, P.; Viaene, T.; Zwiewka, M.; Tejos, R.; Klima, P.; Carna, M.; Rolcik, J.; De Rycke, R.; Moreno, I.; et al. PIN6 auxin transporter at endoplasmic reticulum and plasma membrane mediates auxin homeostasis and organogenesis in Arabidopsis. New Phytol. 2016, 211, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Wisniewska, J.; Xu, J.; Seifertova, D.; Brewer, P.B.; Ruzicka, K.; Blilou, I.; Rouquie, D.; Benkova, E.; Scheres, B.; Friml, J. Polar PIN localization directs auxin flow in plants. Science 2006, 312, 883. [Google Scholar] [CrossRef]
- Dal Bosco, C.; Dovzhenko, A.; Palme, K. Intracellular auxin transport in pollen: PIN8, PIN5 and PILS5. Plant Signal. Behav. 2012, 7, 1504–1505. [Google Scholar] [CrossRef]
- Wang, J.R.; Hu, H.; Wang, G.H.; Li, J.; Chen, J.Y.; Wu, P. Expression of PIN genes in rice (Oryza sativa L.): Tissue specificity and regulation by hormones. Mol. Plant 2009, 2, 823–831. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Zhu, L.; Shou, H.; Wu, P. A PIN1 family gene, OsPIN1, involved in auxin-dependent adventitious root emergence and tillering in rice. Plant Cell Physiol. 2005, 46, 1674–1681. [Google Scholar] [CrossRef]
- Chen, Y.; Fan, X.; Song, W.; Zhang, Y.; Xu, G. Over-expression of OsPIN2 leads to increased tiller numbers, angle and shorter plant height through suppression of OsLAZY1. Plant Biotechnol. J. 2012, 10, 139–149. [Google Scholar] [CrossRef]
- Wu, L.; Luo, Z.; Shi, Y.; Jiang, Y.; Li, R.; Miao, X.; Yang, F.; Li, Q.; Zhao, H.; Xue, J.; et al. A cost-effective tsCUT&Tag method for profiling transcription factor binding landscape. J. Integr. Plant Biol. 2022, 64, 2033–2038. [Google Scholar] [CrossRef]
- Huang, X.L.; Lu, Z.H.; Wang, X.; Ouyang, Y.D.; Chen, W.; Xie, K.L.; Wang, D.Y.; Luo, M.; Luo, J.; Yao, J.L. Imprinted gene OsFIE1 modulates rice seed development by influencing nutrient metabolism and modifying genome H3K27me3. Plant J. 2016, 87, 305–317. [Google Scholar] [CrossRef]
- He, Y.; Zhang, T.; Sun, H.; Zhan, H.; Zhao, Y. A reporter for noninvasively monitoring gene expression and plant transformation. Hortic. Res. 2020, 7, 152. [Google Scholar] [CrossRef]
- Xu, Y.; Zong, W.; Hou, X.; Yao, J.; Liu, H.; Li, X.; Zhao, Y.; Xiong, L. OsARID3, an AT-rich interaction domain-containing protein, is required for shoot meristem development in rice. Plant J. 2015, 83, 806–817. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Liu, S.S.; Liu, Y.Q.; Ling, S.; Chen, C.S.; Yao, J.L. HOTHEAD-Like HTH1 is involved in anther cutin biosynthesis and is required for pollen fertility in rice. Plant Cell Physiol. 2017, 58, 1238–1248. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Zhou, T.; Hu, G.; Wu, H.; Han, Z.; Xiao, J.; Li, X.; Xing, Y. An ethyl methanesulfonate-induced neutral mutant-bridging method efficiently identifies spontaneously mutated genes in rice. Plant J. Cell Mol. Biol. 2020, 104, 1129–1141. [Google Scholar] [CrossRef]
- Wu, C.; Li, X.; Yuan, W.; Chen, G.; Kilian, A.; Li, J.; Xu, C.; Li, X.; Zhou, D.X.; Wang, S.; et al. Development of enhancer trap lines for functional analysis of the rice genome. Plant J. 2003, 35, 418–427. [Google Scholar] [CrossRef]
Figure 1.
(A) Phylogenetic tree of OsSAUR10. Numbers next to each node represent confidence percentages. (B) Subcellular location of OsSAUR10. 35S::OsSAUR10:GFP and the control 35S::GFP were transiently expressed in rice protoplasts individually. Bar = 10 μm.
Figure 1.
(A) Phylogenetic tree of OsSAUR10. Numbers next to each node represent confidence percentages. (B) Subcellular location of OsSAUR10. 35S::OsSAUR10:GFP and the control 35S::GFP were transiently expressed in rice protoplasts individually. Bar = 10 μm.
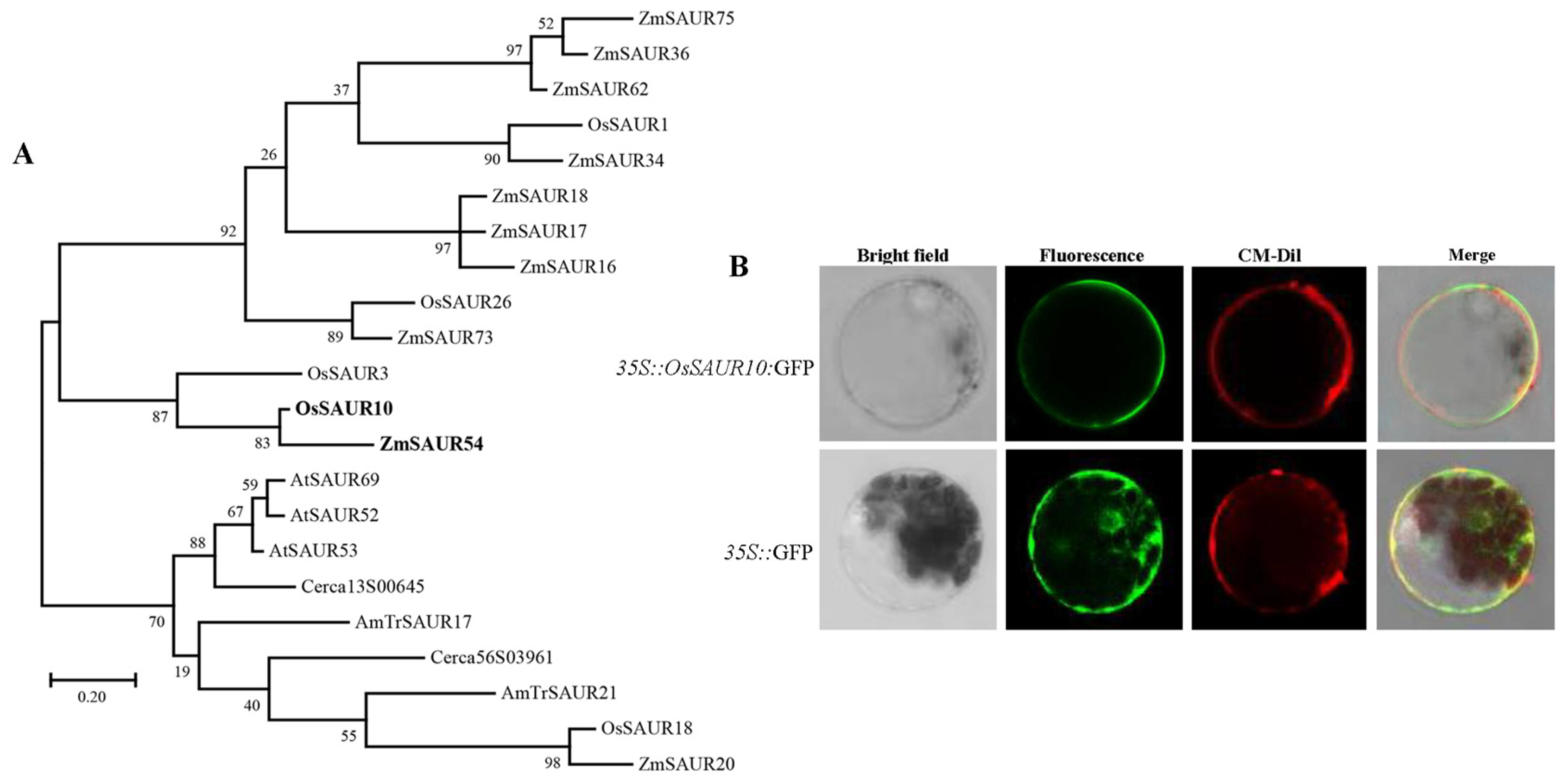
Figure 2.
The expression profiles of OsSAUR10. (A) qRT-PCR analysis of OsSAUR10 in different tissues of rice. (B) GUS staining of seedling 7 days after germination, and the squares individually indicate the elongation regions and the root cap. (C) The enlargement of elongation regions and (D) the root cap of the root. (E) The cross-section of histological observation, (F) the longitudinal section of histological observation, (G) leaf, and (H) root hair. (I) OsSAUR10 transcripts at intervals of 12 h from 0 to 120 HAGs (hours after germination). (B–D,G,H) bar = 0.1 cm; (E,F) bar = 100 μm.
Figure 2.
The expression profiles of OsSAUR10. (A) qRT-PCR analysis of OsSAUR10 in different tissues of rice. (B) GUS staining of seedling 7 days after germination, and the squares individually indicate the elongation regions and the root cap. (C) The enlargement of elongation regions and (D) the root cap of the root. (E) The cross-section of histological observation, (F) the longitudinal section of histological observation, (G) leaf, and (H) root hair. (I) OsSAUR10 transcripts at intervals of 12 h from 0 to 120 HAGs (hours after germination). (B–D,G,H) bar = 0.1 cm; (E,F) bar = 100 μm.
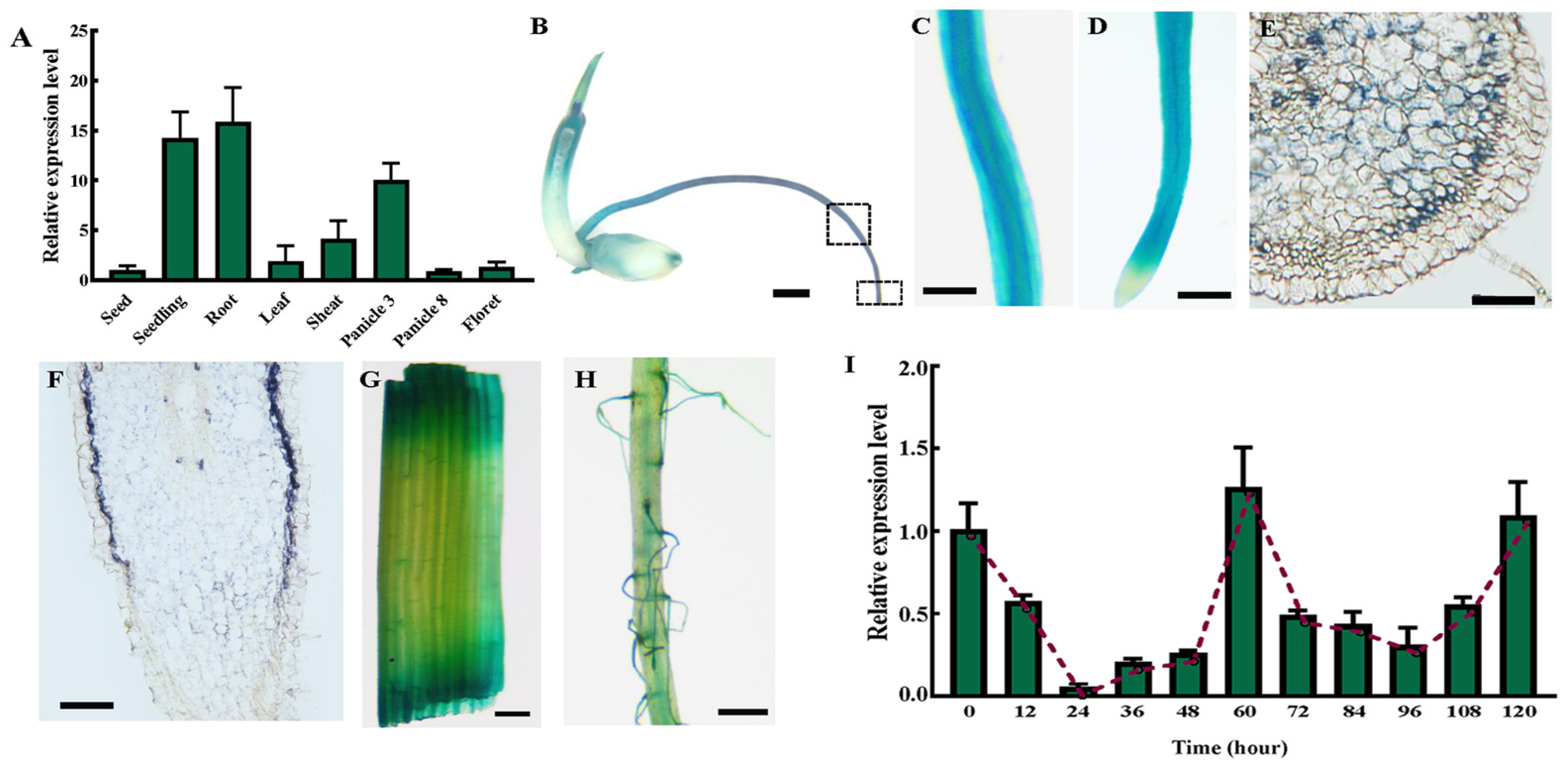
Figure 3.
The inducible responses of OsSAUR10 to various phytohormone treatments. (A) Analysis of OsSAUR10 expressions in 7 DAGs seedlings treated with different phytohormones. * p < 0.05, ** p < 0.01. (B) GUS staining of 7 DAGs seedlings treated with 10 μM IAA, 10 μM ABA, and 1 μM 6-BA for 5 h.
Figure 3.
The inducible responses of OsSAUR10 to various phytohormone treatments. (A) Analysis of OsSAUR10 expressions in 7 DAGs seedlings treated with different phytohormones. * p < 0.05, ** p < 0.01. (B) GUS staining of 7 DAGs seedlings treated with 10 μM IAA, 10 μM ABA, and 1 μM 6-BA for 5 h.
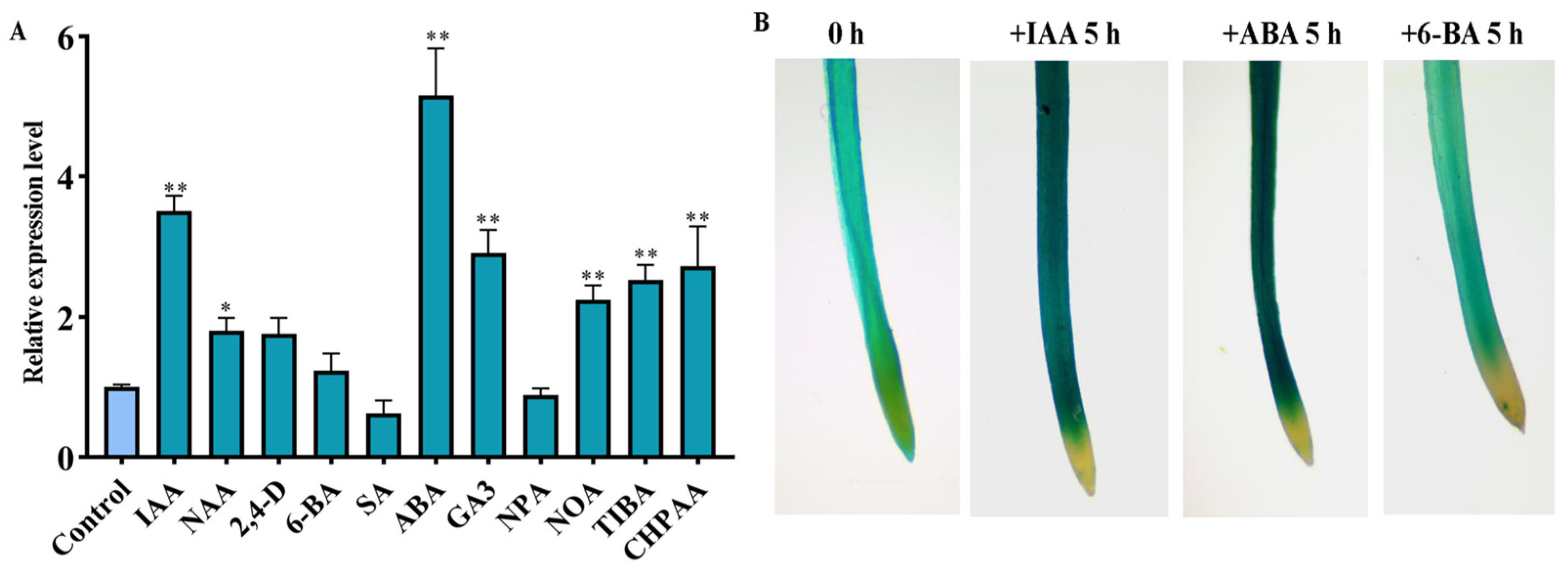
Figure 4.
Phenotypic analysis of ossaur10 mutants. (A) The ossaur10 mutants were generated using CRISPR-Cas9. The upper panel shows the gene model of OsSAUR10, indicating the positions of single nucleotide deletions in the ossaur10 mutant lines. The lower panel displays the mutation sites of ossaur10 compared to the wild-type (WT) sequences. Four color indicate four nucleotides (B) Seedlings of 60 DAGs wild-type and ossaur10 plants. (C) Panicles and different internode lengths of wild-type and ossaur10 plants. (D) Longitudinal sections of histological observation through shoot apex from wild-type and ossaur10 mutants using periodic acid–Schiff staining (PAS) reagent and enlargement of the blue box. The asterisk denotes the shoot apical meristem, and the triangle represents leaf sheath cells, bar = 100 μm. Wild-type and ossaur10 plants after heading. (E) Seedling height of wild-type and ossaur10 plants, as well as (F) root length, (G) plant height, (H) seed setting rate, (I) germination rate, and (J) the number of tillers. (*) denotes significant difference at p < 0.05, (**) represents significant difference at p < 0.01 and the circle dot indicates the specific agronomic value of each seedling.
Figure 4.
Phenotypic analysis of ossaur10 mutants. (A) The ossaur10 mutants were generated using CRISPR-Cas9. The upper panel shows the gene model of OsSAUR10, indicating the positions of single nucleotide deletions in the ossaur10 mutant lines. The lower panel displays the mutation sites of ossaur10 compared to the wild-type (WT) sequences. Four color indicate four nucleotides (B) Seedlings of 60 DAGs wild-type and ossaur10 plants. (C) Panicles and different internode lengths of wild-type and ossaur10 plants. (D) Longitudinal sections of histological observation through shoot apex from wild-type and ossaur10 mutants using periodic acid–Schiff staining (PAS) reagent and enlargement of the blue box. The asterisk denotes the shoot apical meristem, and the triangle represents leaf sheath cells, bar = 100 μm. Wild-type and ossaur10 plants after heading. (E) Seedling height of wild-type and ossaur10 plants, as well as (F) root length, (G) plant height, (H) seed setting rate, (I) germination rate, and (J) the number of tillers. (*) denotes significant difference at p < 0.05, (**) represents significant difference at p < 0.01 and the circle dot indicates the specific agronomic value of each seedling.
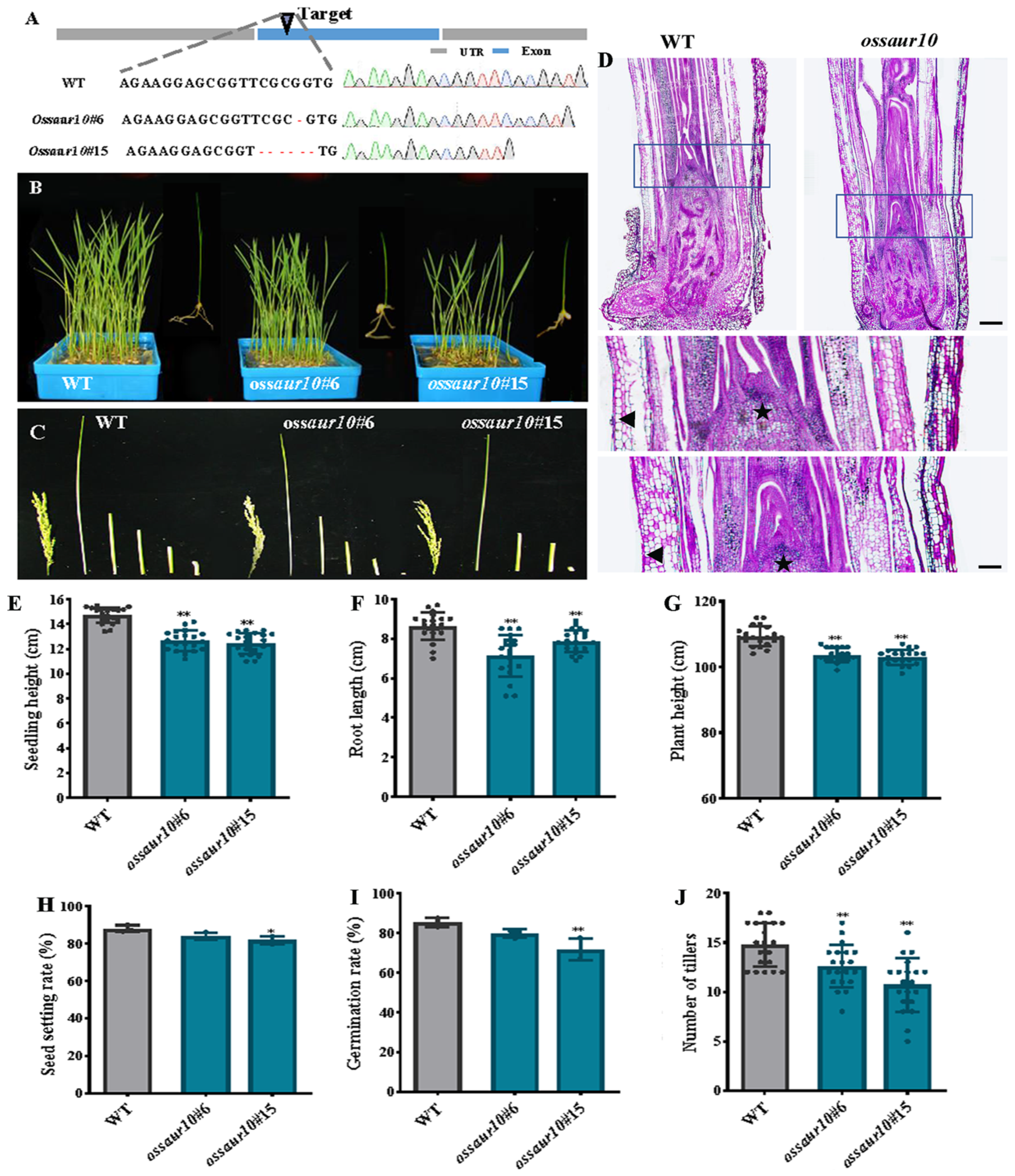
Figure 5.
OsSAUR10-ox seedlings shared the same growth as wild-type (WT). (A) The phenotype of 60 DAGs seedlings. (B) The relative expression level, (C) seedling height, and (D) root length of wild-type and OsSAUR10-ox seedlings. (**) represents significant difference at p < 0.01 and the circle dot indicates the specific agronomic value of each seedling.
Figure 5.
OsSAUR10-ox seedlings shared the same growth as wild-type (WT). (A) The phenotype of 60 DAGs seedlings. (B) The relative expression level, (C) seedling height, and (D) root length of wild-type and OsSAUR10-ox seedlings. (**) represents significant difference at p < 0.01 and the circle dot indicates the specific agronomic value of each seedling.
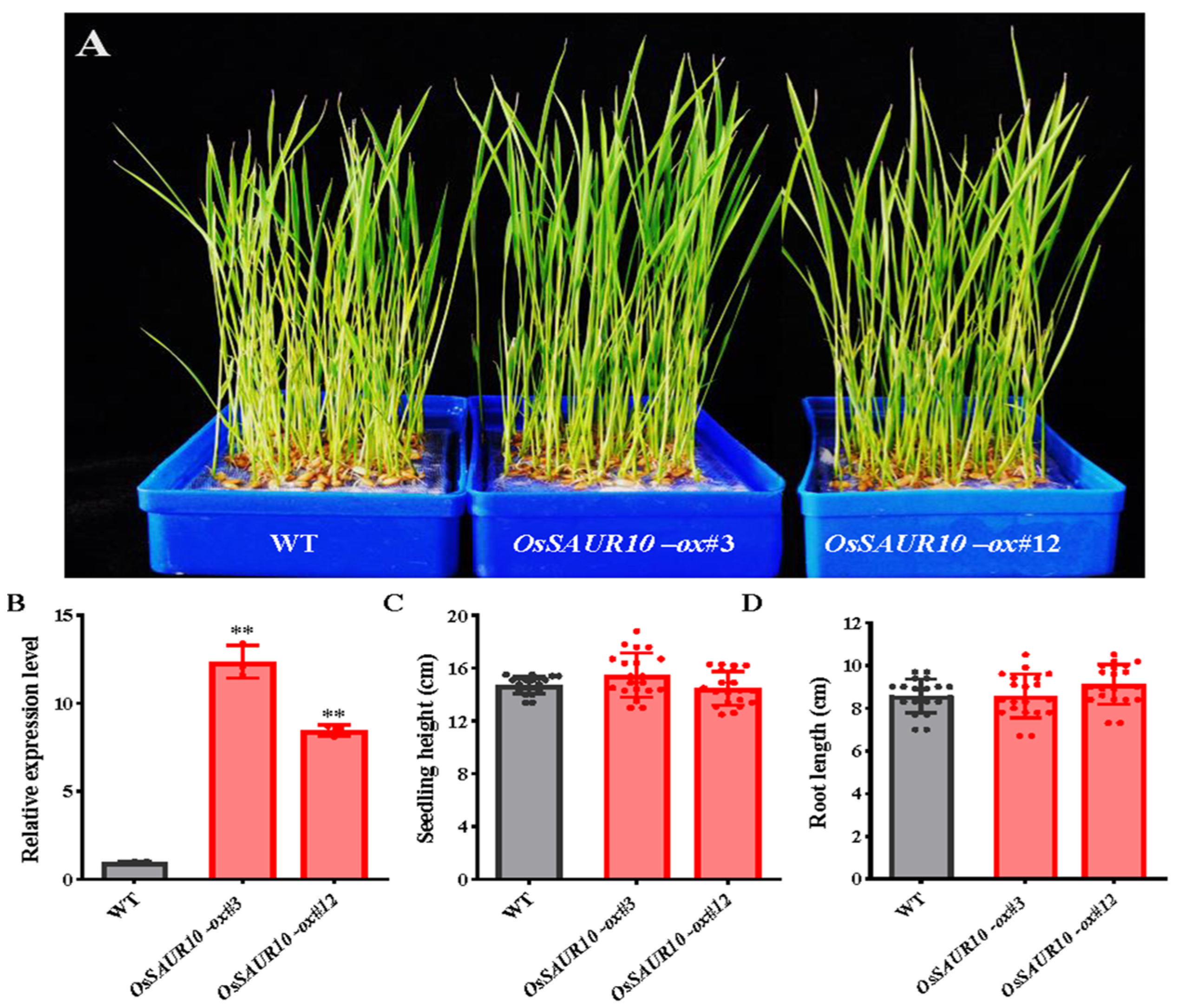
Figure 6.
qRT-PCR analysis of auxin-related genes in three genotypes. (A) Transcripts of auxin growth hormone synthesis OsYUCCA gene. (B) Auxin efflux carrier OsPINs gene family expression in WT, ossaur10, and OsSAUR10-ox seedlings. (**) represents significant difference at p < 0.01.
Figure 6.
qRT-PCR analysis of auxin-related genes in three genotypes. (A) Transcripts of auxin growth hormone synthesis OsYUCCA gene. (B) Auxin efflux carrier OsPINs gene family expression in WT, ossaur10, and OsSAUR10-ox seedlings. (**) represents significant difference at p < 0.01.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Huang, X.; Lu, Z.; Zhai, L.; Li, N.; Yan, H. The Small Auxin-Up RNA SAUR10 Is Involved in the Promotion of Seedling Growth in Rice. Plants 2023, 12, 3880. https://doi.org/10.3390/plants12223880
AMA Style
Huang X, Lu Z, Zhai L, Li N, Yan H. The Small Auxin-Up RNA SAUR10 Is Involved in the Promotion of Seedling Growth in Rice. Plants. 2023; 12(22):3880. https://doi.org/10.3390/plants12223880
Chicago/Turabian StyleHuang, Xiaolong, Zhanhua Lu, Lisheng Zhai, Na Li, and Huiqing Yan. 2023. "The Small Auxin-Up RNA SAUR10 Is Involved in the Promotion of Seedling Growth in Rice" Plants 12, no. 22: 3880. https://doi.org/10.3390/plants12223880
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.