
Pollen Morphology in Sorbus L. (Rosaceae) and Its Taxonomic Implications
by
 , and
, and
Meng Li
1,†,
Chang-Fen Tian
1,†,
Muhammad Idrees
2,
Mitra Pathak
3,
Xian-Hua Xiong
4,
Xin-Fen Gao
5 and
Xian-Rong Wang
1,* 1
Co-Innovation Center for Sustainable Forestry in Southern China, College of Biology and the Environment, Nanjing Forestry University, Nanjing 210037, China
2
College of Life Science, Neijiang Normal University, Neijiang 641000, China
3
Plant Research Center, Salyan, Department of Plant Resources, Ministry of Forests and Environment, Kathmandu 44710, Nepal
4
College of Life Science and Biotechnology, Mianyang Teachers’ College, Mianyang 621000, China
5
Chengdu Institute of Biology, Chinese Academy of Sciences, Chengdu 610041, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Plants 2023, 12(18), 3318; https://doi.org/10.3390/plants12183318
Submission received: 31 August 2023
/
Revised: 8 September 2023
/
Accepted: 14 September 2023
/
Published: 20 September 2023
(This article belongs to the Special Issue Origin and Evolution of the East Asian Flora (EAF))
Abstract
:The genus Sorbus L. in the Rosaceae family is taxonomically challenging due to its morphological variation, polyploidy, and interspecific hybridization. In this study, we used scanning electron microscopy (SEM) to observe the pollen morphology of eighty species, representing six subgenera, in order to assess the differences within the genus Sorbus and its pollen characteristics. We conducted a cluster analysis on three qualitative and four quantitative characteristics. The results demonstrated that the pollen grains of the studied Sorbus species are isopolar and tricolporate. We identified five types of pollen shapes: suboblate, spheroidal, subprolate, prolate, and perprolate. The pollen ornamentation of the investigated species could be classified into five types: striate-perforate, striate, cerebroid-perforate, cerebroid, and foveolate. Interestingly, within the same subgenera, different species exhibited multiple types of characters. The cluster analysis indicated that all 80 species could be divided into six groups, with group B consisting exclusively of species from the subgenus Sorbus. Although pollen micro-morphologies alone do not provide sufficient evidence to establish the taxonomic relationships of the subgenera within Sorbus, they do offer valuable information for species-level taxonomic treatment.
1. Introduction
The genus Sorbus L., which belongs to the family Rosaceae, is of significant economic and ornamental importance. It comprises 258 species and is distributed across Europe, Africa, Asia, and America [1,2]. The East Asia region is considered the center of distribution for Sorbus. Due to factors such as intraspecific variation, polyploidy, and interspecific hybridization, species circumscription within this genus is highly challenging [1,3,4]. Initially, Linnaeus only included two compound-leaved species when he first published about this genus [5]. However, with the advancement in systematic botany, further studies have identified additional species. Some taxa of Sorbus have been merged into other genera of Maleae, such as Pyrus L. and Crataegus L., and considered as subgenera or sections [2,6]. There has been a debate among taxonomists regarding whether Sorbus solely comprises compound-leaved species [7,8,9,10,11] or includes both single-leaved and compound-leaved taxa [3,12,13,14,15,16].
The genus Sorbus has been classified into six subgenera (Sorbus, Cormus Spach, Aria (Pers.) Host, Micromeles Decne., Torminaria M. Roem., and Chamaemespilus Medik.) based on morphological characteristics [1]. However, other studies suggest that Sorbus species can be organized into three sections [16], eleven sections [17], or five groups [6,18]. Recent studies utilizing DNA sequence data in phylogenetic analyses have shown that Sorbus sensu lato is polyphyletic and Sorbus sensu stricto is monophyletic [19,20,21,22,23]. Nevertheless, the infraspecific relationship among the other five subgenera remains unclear. For instance, subgenus Micromeles has been considered both a subgenus within Sorbus [1,16] and a distinct genus [6,24]. Recent research has indicated that hybridization may have occurred between the ancestral lineages of Sorbus and Aria prior to the early Miocene, suggesting that they were the most likely parents of Micromeles [20]. As a result, five hybridogenous genera of Sorbus s.l. have been established based on phylogenetic studies, and the name Sorbomeles has been proposed for hybrids of Sorbus and Micromeles [2]. Rushforth has accepted the six genera published by previous taxonomists, as well as the additional five genera described by Sennikov and Kurtto [2] and has proposed an additional five new genera of Asiatic whitebeams [25]. Consequently, further study is needed to provide reliable taxonomic evidence for the classification of Sorbus.
Pollen morphology is a conserved feature that remains unaffected by changes in environmental conditions, making it a valuable tool for taxonomic classification and identification of plants. Recent advancements in microscopic technology have enabled the observation of pollen surface sculptures, facilitating the resolution of taxonomic problems. Numerous studies in palynology have demonstrated that pollen observation provides effective evidence for the classification and identification of plants at both the generic and specific levels [26,27,28,29,30,31,32,33,34].
Although some research papers have examined and published the pollen morphology of certain Sorbus species [35,36,37,38,39,40], the overall body of work in this area remains limited, often focusing on minority species or specific taxa. Currently, there is a lack of comprehensive documentation regarding the pollen morphology of the Sorbus genus in both China and the rest of the world. Therefore, our study aimed to address these gaps by sampling eighty species across all six subgenera of Sorbus. The specific objectives were to: (1) observe and describe the pollen morphology using scanning electron microscopy (SEM), (2) analyze all collected data and discuss the taxonomic significance of the pollen morphology within Sorbus subgenera and among different species, and (3) explore the correlations between the observed pollen morphology and previous taxonomic research conducted on Sorbus.
2. Results
2.1. Pollen Size and Shape
The pollen morphology of all studied Sorbus species was thoroughly described and illustrated with SEM photographs (Figure 1, Figure 2, Figure 3, Figure 4, Figure 5 and Figure 6), revealing predominantly small- to medium-sized pollen grains. Among the six subgenera examined, medium-sized pollen grains were observed in all, while small-sized pollen grains were specifically found in subgenera Sorbus, Micromeles, and Aria.
On average, the length of the polar axis (P) ranged from 16.64 μm to 50.70 μm, with the entire range extending from 15.59 μm to 54.08 μm. Notably, the longest polar axis length was observed in S. devoniensis of subgenus Aria, while the shortest was observed in S. sargentiana of subgenus Sorbus. Regarding the equatorial axis (E), the mean value varied between 11.58 μm and 32.38 μm, with the entire range spanning from 10.53 μm to 35.06 μm. The species with the longest equatorial axis length was S. tauricola within subgenus Aria, whereas the shortest was found in S. albopilosa of subgenus Sorbus.
The length of colpi ranged from 12.92 μm (S. graeca) to 43.60 μm (S. devoniensis). The C/P ratio demonstrated relative stability, with the majority of species (82.5%) falling within the range of 0.71 to 0.90 μm.
All pollen grains were isopolar and the of P/E ratio varied from 0.83 to 2.09. According to the classification rules, more than half of the pollen grains showed prolate shapes (51.25%). The others appeared in four shapes: suboblate (8.75%), spheroidal (16.25%), subprolate (13.75%), and perprolate (10.00%). Except for subgenera Cormus, Torminaria, and Chamaemespilus, the other subgenera (Micromeles, Sorbus, and Aria) revealed three or five different pollen types.
2.2. Pollen Ornamentation
The variations of all characters selected in this study across subgenera and detailed pollen morphological feature data of the investigated taxa are shown (Table 1 and Supplementary Table S1). According to the diverse morphological characteristics of the examined specimens, the pollen ornamentation was classified into five types (Figure 7): I, striate-perforate, II, striate, III, cerebroid-perforate, IV, cerebroid, and V, foveolate.
Type I: Striate-perforate pollen grains were characterized by irregularly distributed parallel stripes and holes of different sizes on the surface. This type was observed in 29 species belonging to subgenera Aria, Sorbus, and Micromeles, with a hole density of less than 2.54 per μm.
Type II: Striate pollen grains were similar to type I in terms of parallel stripes on the surface, but they lacked perforations. The stripes and grooves ran parallel to the polar axis and often formed fingerprint-like twists. Twenty-five taxa from three subgenera were assigned to type II, including species like S. persica from subgenus Aria, S. foliolosa, and S. multijuga from subgenus Sorbus, and S. thomsonii from subgenus Micromeles.
Type III: Pollen grains of this type exhibited an exine ornamentation that bent into a brain shape with small holes of less than 1 μm in diameter uniformly distributed between lines and grooves on the pollen surface. Twelve species from four subgenera were found to have this type of ornamentation.
Type IV: This type was characterized by regular brain-like stripes on the pollen surface without any holes. Seven species from subgenus Sorbus, as well as S. japonica from subgenus Micromeles, were assigned to this type.
Type V: Foveolate pollen grains were distinct, with evenly distributed holes across the entire surface and few prominent lines. Seven species, including S. torminalis from subgenus Torminaria, S. domestica from subgenus Cormus, and five species from subgenus Sorbus such as S. sitchensis, exhibited this type of ornamentation.
2.3. Cluster Analysis of Sorbus Based on Measured Data
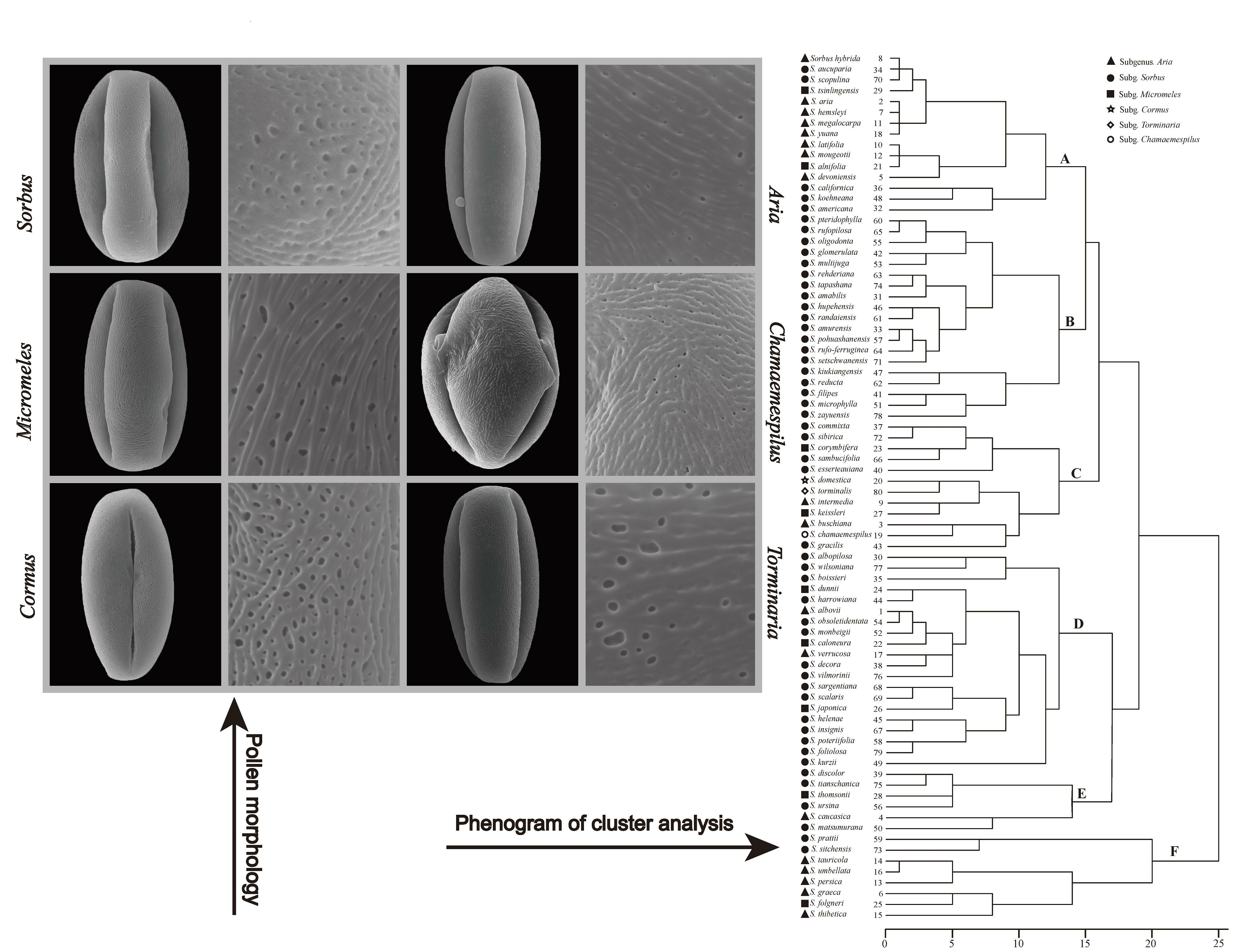
Cluster analysis was conducted using three qualitative and four quantitative characters (Figure 8), resulting in the classification of the examined Sorbus species into six distinct groups (A, B, C, D, E, F). In the cluster tree, all tested species showed intersections with one another, and taxa from the same subgenus did not form separate branches. With the exception of Group B, the other groups consisted of taxa from multiple subgenera.
Group A consisted of fifteen species from three subgenera: Sorbus, Micromeles, and Aria, and significant differences were observed among them. The majority (86.67%) of species in Group A exhibited striate-perforate ornamentation and had prolate or perprolate pollen shapes.
Group B was the only group that exclusively consisted of 19 species from subgenus Sorbus. It was characterized by striate or striate-perforate ornamentation with prolate-shaped pollen grains. The quantitative traits of pollen from these 19 taxa were greater than those of other subgenus Sorbus taxa (P: 23.54–35.38 μm, E: 13.77–27.59 μm, C: 21.27–30.93 μm, C/P: 0.81–0.98 μm).
Group C comprised twelve species from all six subgenera of Sorbus, and was characterized by cerebroid-perforate or foveolate ornamentation. The pollen grains in this group were prolate with longer polar axis lengths (27.07–40.86 μm) and high hole density.
Group D consisted of fifteen taxa from subgenus Sorbus and five taxa from subgenera Aria and Micromeles, and was characterized by imperforate pollen grains.
Group E included species from subgenera Sorbus, Micromeles, and Aria. In comparison to other branches, this group exhibited shorter polar axis lengths.
Group F comprised five taxa from subgenera Aria, Micromeles, and Sorbus, with pollen grains exhibiting suboblate or spheroidal shapes.
3. Discussion
In this study, our focus was on the pollen micro-morphology of the genus Sorbus. The observed variations in pollen ornamentation have provided valuable information. Overall, there is a high degree of similarity in pollen ornamentation at the genus level. However, certain differences have been identified at the subgenus or species level when compared to previous studies.
3.1. Pollen Size and Shape
Regarding pollen size and shape, all the investigated species had small- or medium-sized pollen grains with three apertures, consistent with earlier research. The shape of the pollen grains displayed significant variation, with the majority being spheroidal, subprolate, or prolate, while a few species exhibited suboblate or perprolate shapes.
The mean value of the polar axis length ranged from 16.64 μm to 50.70 μm, and the equatorial axis length ranged from 11.58 μm to 32.38 μm. Previous studies on the pollen morphology of Sorbus have reported mean polar axis lengths ranging from 16.47 μm to 45.0 μm, and equatorial axis lengths varying from 9.89 μm to 34.2 μm [35,36,37,39,40].
Bednorz et al. [36] reported that the polar axis length of S. aria, S. torminalis, S. intermedia, and S. chamaemespilus was typically over 28 μm with some slight overlap. However, in our study, the polar axis length of these species was consistently over 32 μm with relatively stable ranges of variability. In comparison, Yang’s observations of pollen grains showed larger polar and equatorial axis lengths, with D values of up to 12 μm [35]. It is evident that there are some discrepancies between our results and previous studies, particularly in terms of the significant variation in polar and equatorial axis lengths within the same Sorbus species. Previous articles have primarily focused on a limited number of taxa within Sorbus, which may explain the major differences found between our findings and theirs. These findings suggest that pollen size is not a stable characteristic, and that there exists individual variation in pollen grains among Sorbus species. The availability of materials and the selection of species to study are likely critical factors in palynological research on Sorbus.
3.2. Pollen Ornamentation
Ornamentation is a significant distinguishing characteristic in rosaceous pollen, and researchers have regarded pollen ornamentation as the foundation for Sorbus classification [30,35,39,40,41]. In this study, striate sculpture was observed in 54 species, with the highest occurrence in subgenera Sorbus and Aria. Cerebroid sculpture was found in nineteen species belonging to four subgenera (Sorbus, Micromeles, Aria, and Chamaemespilus), and faveolated pollen was found in seven species belonging to subgenera Sorbus, Torminaria, and Cormus. Previous studies by other scholars have likewise identified sculpture, cerebroid, and occasional faveolated ornamentation in the pollen of Sorbus species [30,35,39,40]. Overall, our findings are consistent with previous palynology studies on Sorbus, but there are some variations at the subgenus and species levels.
Five Sorbus species from Poland belonging to subgenera Sorbus, Aria, Chamaemespilus, and Torminaria were reported to have pollen morphology that aligns with their systematic classification, with the exception of subgenus Aria, which exhibited different pollen morphology. The other four subgenera could be easily differentiated based on pollen morphology [36]. However, our study did not confirm this conclusion. There was a high degree of similarity in pollen morphology among the six subgenera. Species within subgenus Sorbus displayed five different ornamentations and shapes, while subgenera Torminaria and Chamaemespilus exhibited ornamentations and shapes that were distinct from subgenus Sorbus. Subgenus Aria showed no distinctive pollen characters, and it encompassed five different pollen shapes and three types of ornamentation.
3.3. The Comparison of Pollen Morphology, Molecular Systematics, and Morphological Classification
Molecular phylogeny and morphology have provided evidence to support the notion that Sorbus s.l. is a polyphyletic genus [20,21,22,23]. Previous studies have also identified five groups within Sorbus s.l. [2,18,42], and morphological data have been categorized into three or six subgenera [1,16]. However, when conducting cluster analysis, the taxa from different subgenera did not form the expected clades. Group B consisted of nineteen species belonging to subgenus Sorbus, while the remaining five groups had individuals from at least three subgenera. Therefore, the pollen morphology does not align with the taxonomic relationships inferred from molecular phylogenies and morphology analysis.
It has been previously demonstrated that Sorbus s.s. (subgenus Sorbus) is a monophyletic group, forming a distinct phylogenetic lineage with species from the temperate zone of the northern hemisphere [19,20,43,44]. Furthermore, subgenus Sorbus is believed to have originated from the primitive Aria [42,45]. There were no significant differences in pollen morphology between subgenera Sorbus and Aria. Although nineteen species from subgenus Sorbus formed a single branch on the cluster tree, the other sixty-one species from six subgenera were intertwined in different branches. Therefore, the claim of subgenus Sorbus being monophyletic cannot be supported by pollen morphology alone.
3.4. Interspecific Clustering of Different Subgenera Based on Pollen Morphology
Sorbus s.l. includes both simple-leaved and compound-leaved taxa. Some taxonomists have divided the simple-leaved taxa into two genera (Aria and Micromeles) or merged them into one genus (Aria). Subgenus Aria mainly occurs in Europe and Asia. Initially, Persoon considered Aria to be a subgenus of Sorbus [1,46]. However, later taxonomic studies placed it in Pyrus or Sorbus as a section [9]. Host treated it as a separate genus [47], and this treatment was followed by Sennikov and Kurtto, who proposed five hybridogenous genera with simple leaves [2]. The instability of the carpel number and free top of the ovary were considered unique characteristics of the Aria and Aucuparia groups [42]. Rushforth described five new genera of simple leaves based on references in the literature [25]. However, the species within the simple-leaved taxa exhibited multiple types in pollen morphology. The majority of the group (70.73%) exhibited striate sculptures, with over half of the species (51.85%) being medium-sized. Different subgenera or genera described by taxonomists do not exhibit distinct characteristics in terms of pollen.
Subgenus Aria consists of some simple-leaved taxa of Sorbus s.l. and mainly occurs in Europe. Scholars have classified the species of subgenus Aria into five (Aria, Griffitharia Rushforth, Wilsonaria Rushforth, Micromeles Decaisne, and Alniaria Rushforth) or four (Aria Host, Karpatiosorbus Sennikov & Kurtto, Hedlundia Sennikov & Kurtto, and Borkhausenia Sennikov & Kurtto) different genera [2,25]. The pollen characteristics of subgenus Aria observed in this study show no obvious regularity. In the cluster analysis results, Aria and other subgenera are often clustered together, but some species of Aria can form a small single branch. The results of biological research indicate that S. torminalis and S. aucuparia are likely involved in the speciation of S. tauricola. However, the pollen morphology of these species shows distinct differences, enabling their differentiation, and the intergroup distance between them is considerable.
Subgenus Micromeles is endemic to Asia and differs from Sorbus in terms of style and carpel structures [6,20,45]. Previously, Micromeles was classified under Aria due to its similarity in fruit structure [48]. Rushforth divided the species of subgenus Micromeles into four genera: Thomsonaria Rushforth, Alniaria Rushforth, Micromeles Decaisne, and Dunniaria Rushforth [25]. In this study, the examination of pollen morphology revealed a variety of shapes and ornamentations. Species of subgenus Micromeles formed no more than two species per branch, and species within subgenus Micromeles were scattered throughout the phenogram. The pollen morphology of Sorbus was insufficient in elucidating the taxonomic relationships among subgenus Micromeles species.
Subgenus Sorbus, also known as Sorbus s.s., consists of species with compound-leaves. Both molecular phylogeny and morphology studies have confirmed its monophyly [15,19,23,43]. No distinct patterns were observed, and the majority of species demonstrated considerable variations in pollen morphology within this subgenus. Except for 19 species that formed a well-defined clade, other taxa within the subgenus were scattered in the cluster analysis tree. The significant differences in pollen characteristics between species also provide useful information for their identification. For instance, S. koehneana was previously considered a synonym or variant of S. multijuga [12]; however, they exhibit different pollen shapes as observed in this study.
Various studies have been reported on the subgenera Torminaria, Chamaemespilus, and Cormus. Clustering analysis has indicated that subgenus Torminaria has an isolated position within Sorbus [45]. However, our data reveal that it cannot be separated from other subgenera. In terms of morphological characters such as flowers and petals, subgenus Chamaemespilus differs from all other subgenera of Sorbus [42]. Species within subgenera Torminaria and Chamaemespilus were mistakenly placed in Crataegus due to the special character differences of Sorbus. While the vegetative parts of subgenus Cormus closely resemble subgenus Sorbus, they can be distinguished by flower and fruit characters. Some studies have reported that subgenus Cormus is distantly related to Sorbus [2,45]. Additionally, the hole density of subgenus Cormus species is slightly larger than that of species from subgenus Torminaria, making it easier to distinguish between the two subgenera. The pollen shape and ornamentation of subgenus Chamaemespilus show a different type compared to subgenera Torminaria and Cormus. In our cluster analysis, these three subgenera were grouped together (Group C). Previous studies have shown that S. intermedia contains apigenin O-glucuronide, which reflects its close affinity to S. torminalis [4,45]. The cluster phenogram also indicates that S. intermedia and S. torminalis share the same branch, indicating a closely related pollen morphology between the two species.
The intersubgeneric and interspecific classification of Sorbus, as determined by morphology and molecular phylogeny, did not align with the findings of pollen morphology. Subgenera Torminaria, Chamaemespilus, and Sorbus have the capacity to hybridize with Aria, indicating that certain Sorbus species derive from intersubgeneric hybridization. The phenogram analysis based on pollen statistics can serve as supporting evidence for interspecific relationships between some hybrid species and their parent species. Notably, certain sibling species that were difficult to differentiate using traditional taxonomic classification exhibited distinct pollen characteristics, such as S. tapashana and S. tianschanica, S. folgneri, and S. hemsleyi. Additionally, numerous species assigned to different subgenera or with distant genetic relationships clustered together in the same branch of the phenogram, including S. domestica, S. keissleri, S. buschiana, and S. gracilis. These results suggest that pollen morphology in Sorbus may evolve in diverse patterns.
4. Materials and Methods
4.1. Sample Collection
A total of eighty species from six subgenera, following the classification of Phipps et al. [1], were collected. Sixty pollen samples were obtained from fresh collections or collected from various herbaria mentioned in Supplementary Table S2. The remaining 20 species’ pollen data were sourced from Jing and Yang [35,39].
4.2. Pollen Morphological Characteristics
Dried pollen grains were mounted on stubs and coated with gold at 10 mA for 1 min using an ion-sputtering device. The morphological features of the pollen grains were observed using an environmental scanning electron microscope (Quanta 200, FEI company, Shanghai, China) at a 10 kV accelerating voltage at Nanjing Forestry University. For species with wide distribution, at least two samples from different areas were scanned. Twenty pollen grains were randomly selected, and their equatorial axis length (E), polar axis length (P), colpi length (C), and hole density were measured using digital SEM images processed with Image J 1.53 [49]. The ratios of colpi length to polar axis length (C/P), polar axis length to equatorial axis length (P/E), as well as the mean values of P and E, were calculated using Excel.
4.3. Cluster Analysis
Three qualitative and four quantitative variables were chosen for cluster analysis, which was performed using IBM SPSS Statistics 25 software. The terminology for pollen shape and ornamentation adhered to Erdtman [50], Wang et al. [51], and Halbritter et al. [52]. The hole density was coded based on the average value of actual measurements for each pollen grain. The pollen shape (P/E) was categorized into five types: suboblate, spheroidal, subprolate, prolate, and perprolate. The five types of pollen grain ornamentation included: striate-perforate, striate, cerebroid-perforate, cerebroid, and foveolate (Figure 7 and Table 2).
5. Conclusions
The importance of SEM studies for accurate and efficient identification of Sorbus s.l. using various palyno-morphological characters has been demonstrated in this study. It has been concluded that there is a high diversity pattern in the pollen of Sorbus. With the exception of subgenera. Cormus, Torminaria, and Chamaemespilus, there are no unique pollen morphologies for any of the subgenera or species, due to the presence of overlapping characters among these subgenera and taxa.
While pollen morphology alone is insufficient to fully elucidate or reconstruct the taxonomic relationships within Sorbus at the sub-generic or sectional level, it can provide valuable information for further taxonomic treatment at the specific level.
This study presents the first comprehensive analysis of Sorbus in terms of pollen morphology. However, the results do not support the monophyly of the six subgenera. For future investigations, it is recommended to increase the sample size and conduct more extensive research.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants12183318/s1, Table S1: Summary of pollen morphological data for investigated Sorbus taxa title; Table S2: Voucher information of genus Sorbus species examined in this study.
Author Contributions
Methodology, M.L.; software, C.-F.T., X.-H.X. and M.I.; writing—original draft preparation, M.L. and C.-F.T.; writing—review and editing, M.L., M.I. and M.P.; visualization, C.-F.T.; supervision, X.-F.G.; project administration, X.-R.W.; funding acquisition, M.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the National Natural Science Foundation of China (Grant No. 32000165) and by the 2016 Shiu-Ying Hu Student/Post-Doctoral Exchange Award from Arnold Arboretum of Harvard University, awarded to Meng Li.
Data Availability Statement
All data and materials used in this study are included in this paper.
Acknowledgments
We thank the curators and staff at A, CDBI, E, GH, MO, KATH, KUN, NF, and PE for allowing us to examine specimens. We are grateful to Faye Rosin (Arnold Arboretum of Harvard University), Yuan Zhao, and Wen-Qi Jiang (Nanjing Forestry University) for their help with experiments and in taking pollen photos.
Conflicts of Interest
The author declares no conflict of interest.
References
- Phipps, J.B.; Robertson, K.R.; Smith, P.G.; Rohrer, J.R. A Checklist of the Subfamily Maloideae (Rosaceae). Can. J. Bot. 1990, 68, 2209–2269. [Google Scholar] [CrossRef]
- Sennikov, A.N.; Kurtto, A. A Phylogenetic Checklist of Sorbus s.l. (Rosaceae) in Europe. Memo. Soc. Fauna Flora Fenn. 2017, 93, 1–78. Available online: https://journal.fi/msff/article/view/64741 (accessed on 15 March 2022).
- Lu, L.D.; Spongberg, S.A. Sorbus. In Flora of China; Wu, Z.Y., Peter, H.R., Hong, D., Eds.; Science Press & Missouri Botanical Garden Press: Beijing, China; Saint Louis, MI, USA, 2003; Volume 9, pp. 144–170. [Google Scholar]
- Németh, C.; Papp, N.; Nosková, J.; Höhn, M. Speciation by Triparental Hybridization in Genus Sorbus (Rosaceae). Biol. Futura 2020, 71, 209–222. [Google Scholar] [CrossRef] [PubMed]
- Linnaeus, C. Species Plantarum; Impensis Laurentii Salvii: Holmiae, Sweden, 1753; Volume 2, p. 1200. [Google Scholar]
- Kovanda, M.; Challice, J. The Genus Micromeles Revisited. Folia Geobot. 1981, 16, 181–193. [Google Scholar] [CrossRef]
- Hooker, J.D. Rosaceae. In The Flora of British India; L. Reeve: London, UK, 1879; Volume 2, pp. 307–388. [Google Scholar]
- Lindley, J.X. Observations on the Natural Group of Plants Called Pomaceae. Trans. Linn. Soc. Lond. 1821, 13, 88–106. [Google Scholar] [CrossRef]
- de Candolle, A.; de Candolle, A.P. Rosaceae. In Prodromus Systematis Naturalis Regni Vegetabilis, Sive, Enumeratio Contracta Ordinum Generum Specierumque Plantarum Huc Usque Cognitarium, Juxta Methodi Naturalis, Normas Digesta; Sumptibus Sociorum Treuttel & Würtz: Paris, France, 1825; Volume 2, pp. 525–639. [Google Scholar]
- Focke, W.O. Rosaceae. In Die Natürlichen Pflanzenfamilien Nebst Ihren Gattungen Und Wichtigeren Arten, Insbesondere Den Nutzpflanzen, Unter Mitwirkung Zahlreicher Hervorragender Fachgelehrten Begründet; Engler, A., Krause, K., Pilger, R., Prantl, K., et al., Eds.; Verlag von Wilhelm Engelman: Leipzig, Germany, 1894; Volume 3, pp. 1–61. [Google Scholar]
- Roemer, M.J. Familiarum Naturalium Regni Vegetabilis Synopses Monographicae; Landes-Industrie-Comptoir: Weimar, Germany, 1847; p. 314. [Google Scholar]
- Rehder, A.; Koehne, B.A.E. Rosaceae. In Plantae Wilsonianae: An Enumeration of the Woody Plants Collected in Western China for the Arnold Arboretum of Harvard University During the Years 1907, 1908, and 1910 by E. H. Wilson; Sargent, C.S., Wilson, E.H., Eds.; The University Press: Cambridge, UK, 1913; Volume 2, pp. 434–483. [Google Scholar]
- Wenzig, T. Die Pomaceen. Charaktere Der Gattungen Und Arten. Jahrb.; Notizblatt des Königl. Bot. Gart. Berlin 1883, 2, 287–307. [Google Scholar]
- Hedlund, T. Monographie Der Gattung Sorbus; Kessinger Publishing: Whitefish, MT, USA, 1901; p. 163. [Google Scholar]
- Wang, G.X.; Zhang, M.L. A Molecular Phylogeny of Sorbus (Rosaceae) Based on ITS Sequence. Yuan Yi Xue Bao 2011, 38, 2387–2394. [Google Scholar] [CrossRef]
- Yu, D.J.; Lu, L.D.; Gu, C.Z.; Guan, K.J.; Jiang, W.F. Rosaceae. In Flora of China; Editorial Committee of Flora of China; Chinese Academy of Sciences, Ed.; Science Press: Beijing, China, 1974; Volume 36, pp. 1–404. [Google Scholar]
- McAllister, H.; Mathew, B. The Genus Sorbus: Mountain Ash and Other Rowans; Royal Botanic Gardens: London, UK, 2005; p. 256. [Google Scholar]
- Li, Q.Y.; Guo, W.; Liao, W.B.; Macklin, J.A.; Li, J.H. Generic Limits of Pyrinae: Insights from Nuclear Ribosomal DNA Sequences. Bot. Stud. 2012, 53, 151–164. Available online: https://www.researchgate.net/publication/254607803 (accessed on 20 February 2022).
- Zheng, D.M.; Zhang, M.L. A Cladistic and Phenetic Analysis of the Infrageneric Relationships of Sorbus s. l. (Maloideae, Rosaceae) Based on the Morphological Characters. Yuan Yi Xue Bao 2007, 34, 723–728. [Google Scholar] [CrossRef]
- Lo, E.Y.Y.; Donoghue, M.J. Expanded Phylogenetic and Dating Analyses of the Apples and Their Relatives (Pyreae, Rosaceae). Mol. Phylogenet Evol. 2012, 63, 230–243. [Google Scholar] [CrossRef]
- Zhang, S.D.; Jin, J.J.; Chen, S.Y.; Chase, M.W.; Soltis, D.E.; Li, H.T.; Yang, J.B.; Li, D.Z.; Yi, T.S. Diversification of Rosaceae since the Late Cretaceous Based on Plastid Phylogenomics. New Phytol. 2017, 214, 1355–1367. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Huang, C.H.; Hu, Y.; Wen, J.; Li, S.; Yi, T.; Chen, H.; Xiang, J.; Ma, H. Evolution of Rosaceae Fruit Types Based on Nuclear Phylogeny in the Context of Geological Times and Genome Duplication. Mol. Biol. Evol. 2017, 34, 262–281. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.H.; Shi, S.; Li, J.L.; Yu, J.; Wang, L.; Yang, X.Y.; Guo, L.; Zhou, S.L. Phylogeny of Maleae (Rosaceae) Based on Multiple Chloroplast Regions: Implications to Genera Circumscription. BioMed. Res. Int. 2018, 2018, 7627191. [Google Scholar] [CrossRef] [PubMed]
- Gabrielyan, E.T. Sorbus, L. In Western Asia and the Himalayas; Izdatel’stvo AN ArmSSR: Erevan, Armenia, 1978; p. 329. [Google Scholar]
- Rushforth, K. The Whitebeam Problem, and a Solution. Phytologia 2018, 100, 222–247. Available online: https://www.phytologia.org/uploads/2/3/4/2/23422706/100_4_222-247rushforthmalinae__rosaceae_12-5-18.pdf (accessed on 19 May 2022).
- Lechowicz, K.; Wrońska-Pilarek, D.; Bocianowski, J.; Maliński, T. Pollen Morphology of Polish Species from the Genus Rubus L. (Rosaceae) and Its Systematic Importance. PLoS ONE 2020, 15, e0237625. [Google Scholar] [CrossRef]
- Lu, L.; Fritsch, P.W.; Wang, H.; Li, H.T.; Li, D.Z.; Chen, J.Q. Pollen Morphology of Gaultheria L. And Related Genera of Subfamily Vaccinioideae: Taxonomic and Evolutionary Significance. Rev. Palaeobot. Palyno. 2009, 154, 106–123. [Google Scholar] [CrossRef]
- Wan, C.Y.; Liu, J.X. The Morphology and Systematics of the Pollen of Stellaria. Palynology 2017, 41, 533–546. [Google Scholar] [CrossRef]
- Xiong, X.H.; Zhou, X.M.; Li, M.; Xu, B.; Deng, H.N.; Yu, Q.; Gao, X.F. Pollen Morphology in Rubus (Rosaceae) and Its Taxonomic Implications. Plant Syst. Evol. 2019, 305, 705–716. [Google Scholar] [CrossRef]
- Song, J.H.; Moon, H.K.; Hong, S.P. Pollen Morphology of the Tribe Sorbarieae (Rosaceae). Plant Syst. Evol. 2016, 302, 853–869. [Google Scholar] [CrossRef]
- Sarwar, A.K.M.G.; Hoshino, Y.; Araki, H. Pollen Morphology and Its Taxonomic Significance in the Genus Bomarea Mirb. (Alstroemeriaceae)—I. Subgenera Baccata, Sphaerine, and Wichuraea. Acta Bot. Bras. 2015, 29, 425–432. [Google Scholar] [CrossRef]
- Banks, H.I.; Forest, F.; Lewis, G. Evolution and Diversity of Pollen Morphology in Tribe Cercideae (Leguminosae). Taxon 2014, 63, 299–314. [Google Scholar] [CrossRef]
- Hebda, R.J.; Chinnappa, C.C.; Smith, B.M. Pollen Morphology of the Rosaceae of Western Canada. Grana 1988, 27, 95–113. [Google Scholar] [CrossRef]
- Liao, M.; Ullah, F.; Deng, H.N.; Zhang, J.Y.; Xu, B.; Gao, X.F. Pollen Morphology of the Genus Sophora (Fabaceae) and Its Taxonomic Implications. Microsc. Res. Techniq. 2022, 85, 1723–1741. [Google Scholar] [CrossRef]
- Yang, L.H.; Wu, Y.H.; Pei, X.; Guan, X.L.; Zheng, J. Pollen Morphological Characteristics and Cluster Analysis on Some Species in Sorbus Linn. J. Plant Resour. Eviron. 2019, 28, 84–90. [Google Scholar] [CrossRef]
- Bednorz, L.; Maciejewska, I.; Wronska-Pilarek, D.; Fujiki, T. Pollen Morphology of the Polish Species of the Genus Sorbus L. Acta Soc. Bot. Pol. 2005, 74, 315–322. [Google Scholar] [CrossRef]
- Bednorz, L.; Fujiki, T. Pollen Morphology of Some European Sorbus Species. Rocz. AR. Pozn. 2003, 6, 3–7. Available online: https://www.researchgate.net/publication/238084984 (accessed on 13 September 2023).
- Hebda, R.J.; Chinnappa, C.C. Studies on Pollen Morphology of Rosaceae. Acta Bot. Gallica 1994, 141, 183–193. [Google Scholar] [CrossRef]
- Jing, H.J. Taxonomic Revision of Sorbus L. (Rosaceae). Ph.D. Thesis, Sun Yat sen University, Guangzhou, China, 2015. [Google Scholar]
- Eide, F. Key for Northwest European Rosaceae Pollen. Grana 1981, 20, 101–118. [Google Scholar] [CrossRef]
- Shi, S.; Li, J.; Sun, J.; Yu, J.; Zhou, S. Phylogeny and Classification of Prunus Sensu Lato (Rosaceae). J. Integr. Plant Biol. 2013, 55, 1069–1079. [Google Scholar] [CrossRef]
- Kovanda, M. Flower and Fruit Morphology of Sorbus in Correlation to the Taxonomy of the Genus. Preslia 1961, 33, 1–16. [Google Scholar]
- Li, M.; Toma, T.; Gao, Y.D.; Xu, B.; Zhu, Z.M.; Ju, W.B.; Gao, X.F. Molecular Phylogenetics and Historical Biogeography of Sorbus Sensu Stricto (Rosaceae). Mol. Phylogenet. Evol. 2017, 111, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Z.R. Leaf Characters of Sorbus Section Sorbus from China and Their Taxonomic Significance. Master’s Thesis, Nanjing Forestry University, Nanjing, China, 2019. [Google Scholar]
- Challice, J.; Kovanda, M. Flavonoids as Markers of Taxonomic Relationships in the Genus Sorbus in Europe. Preslia 1978, 50, 305–320. Available online: https://www.preslia.cz/archive/Preslia_50_1978_305-320.pdf (accessed on 22 February 2023).
- Persoon, C.H. Synopsis Plantarum, Seu Enchiridium Botanicum, Complectens Enumerationem Systematicam Specierum Hucusque Cognitarum; Parisiis Lutetiorum, C.F. Cramerum: Paris, France, 1807; Volume 2, p. 656. [Google Scholar]
- Host, N.T. Flora Austriaca; Arkose Press: Viennae, Austria, 1831; Volume 2, pp. 7–17. [Google Scholar]
- Robertson, K.R.; Phipps, J.B.; Rohrer, J.R.; Smith, P.G. A Synopsis of Genera in Maloideae (Rosaceae). Syst. Bot. 1991, 16, 376–394. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. Nih Image to Imagej: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Erdtman, G. Handbook of Palynology-An Introduction to the Study of Pollen Grains and Spores; Munksgaard: Copenhagen, Denmark, 1969; p. 496. [Google Scholar]
- Wang, F.S.; Chien, N.S.; Zhang, Y.L.; Yang, H.Q. Pollen Flora of China; Science Press: Beijing, China, 1995; pp. 3–10. [Google Scholar]
- Halbritter, H.; Ulrich, S.; Grímsson, F.; Weber, M.; Zetter, R.; Hesse, M.; Buchner, R.; Svojtka, M.; Frosch-Radivo, A. Illustrated Pollen Terminology, 2nd ed.; Springer: Cham, Switzerland, 2018. [Google Scholar]
Figure 1.
SEM micrographs of pollen grains of studied Sorbus taxa. (1-a–10-a) Pollen grains in polar view. (1-b–10-b) Pollen ornamentation. 1, S. albovii; 2, S. alnifolia; 3, S. americana; 4, S. amurensis; 5, S. aira; 6, S. aucuparia; 7, S. boissieri; 8, S. buschiana; 9, S. californica; 10, S. caloneura.
Figure 1.
SEM micrographs of pollen grains of studied Sorbus taxa. (1-a–10-a) Pollen grains in polar view. (1-b–10-b) Pollen ornamentation. 1, S. albovii; 2, S. alnifolia; 3, S. americana; 4, S. amurensis; 5, S. aira; 6, S. aucuparia; 7, S. boissieri; 8, S. buschiana; 9, S. californica; 10, S. caloneura.

Figure 2.
SEM micrographs of pollen grains of studied Sorbus taxa. (1-a–10-a) Pollen grains in polar view. (1-b–10-b) Pollen ornamentation. 1, S. caucasica; 2, S. chamaemespilus; 3, S. commixta; 4, S. corymbifera; 5, S. decora; 6, S. devoniensis; 7, S. discolor; 8, S. domestica; 9, S. dunnii; 10, S. esserteauiana.
Figure 2.
SEM micrographs of pollen grains of studied Sorbus taxa. (1-a–10-a) Pollen grains in polar view. (1-b–10-b) Pollen ornamentation. 1, S. caucasica; 2, S. chamaemespilus; 3, S. commixta; 4, S. corymbifera; 5, S. decora; 6, S. devoniensis; 7, S. discolor; 8, S. domestica; 9, S. dunnii; 10, S. esserteauiana.

Figure 3.
SEM micrographs of pollen grains of studied Sorbus taxa. (1-a–10-a) Pollen grains in polar view. (1-b–10-b) Pollen ornamentation. 1, S. folgneri; 2, S. foliolosa; 3, S. gracilis; 4, S. graeca; 5, S. harrowiana; 6, S. helenae; 7, S. hemsleyi; 8, S. hupehensis; 9, S. hybrida; 10, S. insignis.
Figure 3.
SEM micrographs of pollen grains of studied Sorbus taxa. (1-a–10-a) Pollen grains in polar view. (1-b–10-b) Pollen ornamentation. 1, S. folgneri; 2, S. foliolosa; 3, S. gracilis; 4, S. graeca; 5, S. harrowiana; 6, S. helenae; 7, S. hemsleyi; 8, S. hupehensis; 9, S. hybrida; 10, S. insignis.

Figure 4.
SEM micrographs of pollen grains of studied Sorbus taxa. (1-a–10-a) Pollen grains in polar view. (1-b–10-b) Pollen ornamentation. 1, S. intermedia; 2, S. japonica; 3, S. keissleri; 4, S. koehneana; 5, S. kurzii; 6, S. latifolia; 7, S. matsumurana; 8, S. monbeigii; 9, S. mougeotii; 10, S. persica.
Figure 4.
SEM micrographs of pollen grains of studied Sorbus taxa. (1-a–10-a) Pollen grains in polar view. (1-b–10-b) Pollen ornamentation. 1, S. intermedia; 2, S. japonica; 3, S. keissleri; 4, S. koehneana; 5, S. kurzii; 6, S. latifolia; 7, S. matsumurana; 8, S. monbeigii; 9, S. mougeotii; 10, S. persica.

Figure 5.
SEM micrographs of pollen grains of studied Sorbus taxa. (1-a–10-a) Pollen grains in polar view. (1-b–10-b) Pollen ornamentation. 1, S. pohuashanensis; 2, S. prattii; 3, S. rehderiana; 4, S. rufo-ferruginea; 5, S. sambucifolia; 6, S. sargentiana; 7, S. scopulina; 8, S. sibirica; 9, S. sitchensis; 10, S. tauricola.
Figure 5.
SEM micrographs of pollen grains of studied Sorbus taxa. (1-a–10-a) Pollen grains in polar view. (1-b–10-b) Pollen ornamentation. 1, S. pohuashanensis; 2, S. prattii; 3, S. rehderiana; 4, S. rufo-ferruginea; 5, S. sambucifolia; 6, S. sargentiana; 7, S. scopulina; 8, S. sibirica; 9, S. sitchensis; 10, S. tauricola.

Figure 6.
SEM micrographs of pollen grains of studied Sorbus taxa. (1-a–10-a) Pollen grains in polar view. (1-b–10-b) Pollen ornamentation. 1, S. thibetica; 2, S. thomsonii; 3, S. tianschanica; 4, S. torminalis; 5, S. tsinlingensis; 6, S. umbellate; 7, S. ursine; 8, S. verrucosa; 9, S. vilmorinii; 10, S. yuana.
Figure 6.
SEM micrographs of pollen grains of studied Sorbus taxa. (1-a–10-a) Pollen grains in polar view. (1-b–10-b) Pollen ornamentation. 1, S. thibetica; 2, S. thomsonii; 3, S. tianschanica; 4, S. torminalis; 5, S. tsinlingensis; 6, S. umbellate; 7, S. ursine; 8, S. verrucosa; 9, S. vilmorinii; 10, S. yuana.

Figure 7.
Scanning Electron Microphotographs of pollen ornamentation types. (Type I), striate-perforate; (Type II), striate; (Type III), cerebroid-perforate; (Type IV), cerebroid; (Type V), foveolate.
Figure 7.
Scanning Electron Microphotographs of pollen ornamentation types. (Type I), striate-perforate; (Type II), striate; (Type III), cerebroid-perforate; (Type IV), cerebroid; (Type V), foveolate.

Figure 8.
The phenogram of cluster analysis is based on 7 characters.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Distribution of subgenus pollen morphology.
| Characters | Subgenus (Number of Species) | ||||||
|---|---|---|---|---|---|---|---|
| Aira | Chamaemespilus | Cormus | Micromeles | Sorbus | Torminaria | Total | |
| Ornamentation Type I | 10 | 0 | 0 | 2 | 17 | 0 | 29 |
| Ornamentation Type II | 5 | 0 | 0 | 2 | 18 | 0 | 25 |
| Ornamentation Type III | 3 | 1 | 0 | 4 | 4 | 0 | 12 |
| Ornamentation Type IV | 0 | 0 | 0 | 1 | 6 | 0 | 7 |
| Ornamentation Type V | 0 | 0 | 1 | 0 | 5 | 1 | 7 |
| Suboblate | 3 | 0 | 0 | 1 | 3 | 0 | 7 |
| Spheroidal | 3 | 0 | 0 | 0 | 10 | 0 | 13 |
| Subprolate | 1 | 1 | 0 | 2 | 7 | 0 | 11 |
| Prolate | 8 | 0 | 1 | 6 | 25 | 1 | 41 |
| Perprolate | 3 | 0 | 0 | 0 | 5 | 0 | 8 |
| Hole density (0) | 5 | 0 | 0 | 3 | 24 | 0 | 32 |
| Hole density (0.18–2.81) | 12 | 0 | 0 | 5 | 25 | 1 | 43 |
| Hole density (3.74–5.70) | 1 | 1 | 1 | 1 | 1 | 0 | 5 |
Table 2.
Morphological characters used in cluster analysis.
| Characters | Type of Traits | Code |
|---|---|---|
| Length polar axis (P) | Quantitative | μm |
| Length of equatorial axis (E) | Quantitative | μm |
| Length of colpi (C) | Quantitative | μm |
| The ratio of colpus length to Polar axis length (C/P) | Quantitative | ratio |
| Pollen shape (P/E) | Qualitative | suboblate = 1; spheroidal = 2; subprolate = 3; prolate = 4; perprolate = 5 |
| Pollen ornamentation | Qualitative | striate-perforate = 1; striate = 2; cerebroid-perforate = 3; cerebroid = 4; foveolate = 5 |
| Hole density | Qualitative | absent = 0; 0.18–2.81/μm2 = 1; 3.74–5.70/μm2 = 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, M.; Tian, C.-F.; Idrees, M.; Pathak, M.; Xiong, X.-H.; Gao, X.-F.; Wang, X.-R. Pollen Morphology in Sorbus L. (Rosaceae) and Its Taxonomic Implications. Plants 2023, 12, 3318. https://doi.org/10.3390/plants12183318
AMA Style
Li M, Tian C-F, Idrees M, Pathak M, Xiong X-H, Gao X-F, Wang X-R. Pollen Morphology in Sorbus L. (Rosaceae) and Its Taxonomic Implications. Plants. 2023; 12(18):3318. https://doi.org/10.3390/plants12183318
Chicago/Turabian StyleLi, Meng, Chang-Fen Tian, Muhammad Idrees, Mitra Pathak, Xian-Hua Xiong, Xin-Fen Gao, and Xian-Rong Wang. 2023. "Pollen Morphology in Sorbus L. (Rosaceae) and Its Taxonomic Implications" Plants 12, no. 18: 3318. https://doi.org/10.3390/plants12183318
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.