
Recent Updates on ALMT Transporters’ Physiology, Regulation, and Molecular Evolution in Plants

1
Department of Biotechnology Engineering, Braude Academic College of Engineering, Snunit 51, Karmiel 2161002, Israel
2
Institute of Agricultural and Nutritional Sciences, Martin Luther University Halle-Wittenberg, Betty-Heimann-Strasse 3, 06120 Halle, Germany
3
Department of Plant Food Products and Biofortification, Institute of Food Biotechnology and Genomics, The National Academy of Sciences of Ukraine, Osipovskogo Str. 2a, 04123 Kyiv, Ukraine
*
Author to whom correspondence should be addressed.
Plants 2023, 12(17), 3167; https://doi.org/10.3390/plants12173167
Submission received: 5 June 2023
/
Revised: 18 August 2023
/
Accepted: 29 August 2023
/
Published: 4 September 2023
(This article belongs to the Section Plant Response to Abiotic Stress and Climate Change)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Aluminium toxicity and phosphorus deficiency in soils are the main interconnected problems of modern agriculture. The aluminium-activated malate transporters (ALMTs) comprise a membrane protein family that demonstrates various physiological functions in plants, such as tolerance to environmental Al3+ and the regulation of stomatal movement. Over the past few decades, the regulation of ALMT family proteins has been intensively studied. In this review, we summarise the current knowledge about this transporter family and assess their involvement in diverse physiological processes and comprehensive regulatory mechanisms. Furthermore, we have conducted a thorough bioinformatic analysis to decipher the functional importance of conserved residues, structural components, and domains. Our phylogenetic analysis has also provided new insights into the molecular evolution of ALMT family proteins, expanding their scope beyond the plant kingdom. Lastly, we have formulated several outstanding questions and research directions to further enhance our understanding of the fundamental role of ALMT proteins and to assess their physiological functions.
1. Introduction
Aluminium-activated malate transporter (ALMT) proteins play vital roles in plant adaptation to aluminium toxicity and acid soils. Soil acidification is characterised by the increased mobility of Al3+ ions, which tend to bind phosphorus into stable complexes, thus combining Al3+ toxicity and decreased phosphate bioavailability. Therefore, the release of malate allows for neutralisation of Al3+ and the disengagement of phosphates from the complex. Early experiments indicated that AtALMT1 is regulated by low pH, H2O2, auxin, and abscisic acid (ABA) [1]; Ca2+ [2]; and γ-Aminobutyric acid (GABA) [3]. Further experiments with soybean ALMT1 demonstrated that the presence of adequate phosphate levels is crucial for soybean adaptation to acid soils [4]. On the contrary, ethylene was shown to act as a negative regulator of the Al3+-induced malate efflux [5].
Recently, the expression of the Arabidopsis thaliana ((L.) Heynh). ALMT gene family under phosphorus-deficient conditions was characterised. In contrast to early experiments, only AtALMT3 was significantly up-regulated in the root by low-phosphate conditions. Specifically, the AtALMT3 gene was expressed in the root hair cells and protein was localised in the plasma membrane and small vesicles. AtALMT3 knock-down and overexpressing lines demonstrated reduced malate exudation and increased malate exudation, respectively, under phosphate-deficient conditions. Therefore, these results indicate the crucial role of plasma membrane-localised AtALMT3 in low-phosphate-inducible malate exudation in Arabidopsis root hair [6]. Interestingly, the ALMT1 homologue from Lupinus albus L., which was expressed in the root apices under phosphorus deficiency, was moderately repressed by Al, involved in Al-independent malate release from root tips, and participated in cation/metal root-to-shoot translocation [7].
The regulatory role of abscisic acid under Al3+ stress was recently updated. Experiments on rice bean (Vigna umbellata (Thunb.) Ohwi and H. Ohashi) and Arabidopsis abi5 (ABA-Insensitive 5) mutants demonstrated that ABA-mediated Al stress response is not associated with known Al tolerance mechanisms (AtALMT1 and MULTI-DRUG AND TOXIC COMPOUND EXTRUSION (AtMATE)). Interestingly, nearly 1/3 of genes activated by Al stress and ABA treatment are the same. Furthermore, ABI5 was identified as a transcription factor that mediates Al stress tolerance via regulation of these genes, most importantly, osmoregulation and cell wall modification-related genes (such as Peroxidase 72, Stress-associated protein 12, XYLOGLUCAN ENDOTRANSGLUCOSYLASE/HYDROLASE 23, and GLUTATHIONE S-TRANSFERASE TAU 8), suggesting the presence of another ABA-dependent mechanism of Al tolerance in plants [8].
Additionally, experimental results obtained from A. thaliana and Amaranthus mangostanus (L.) suggested that the expression of ALMT genes is up-regulated by salt stress, thus increasing the release of malate to the rhizosphere [9,10]. Similarly, Guzmania monostachia (L.) Rusby ex Mez) plants supplied with ammonium and exposed to drought stress showed higher expression of ALMT1, increased accumulation of soluble sugars involved in osmotic adjustment, and higher antioxidant enzyme activity (SOD, CAT, APX, and GR) [11]. Also, the comparison of Al3+-tolerant and Al3+-sensitive wheat (Triticum aestivum L.) lines demonstrated that roots of tolerant lines exuded more malate and GABA under high pH and Al treatment, thus acidifying the rhizosphere faster. While the expression of TaALMT1 did not change under high pH, the expression of GABA synthesis genes (TaGAD and TaGAD1) was increased and the expression of the GABA catabolism gene (GABA-T) was decreased in the tolerant line [12].
Thus, the physiological roles of ALMT transporters are substantially wider than was thought before. This type of anion transporter might play a crucial role in physiological adjustments during adaptation to a wide range of stresses. However, many sides of their functionality, physiology, regulation, and interaction remain largely unknown. In this review, we summarise the recent knowledge on the ALMT family and discuss molecular mechanisms regulating their functioning and involvement in various physiological processes. Finally, we conducted a rigorous bioinformatic and molecular evolution analysis of the ALMT protein family using currently available sequencing data, solved crystal structures, and bioinformatic tools.
2. Bioinformatic Analysis of ALMT Family Proteins
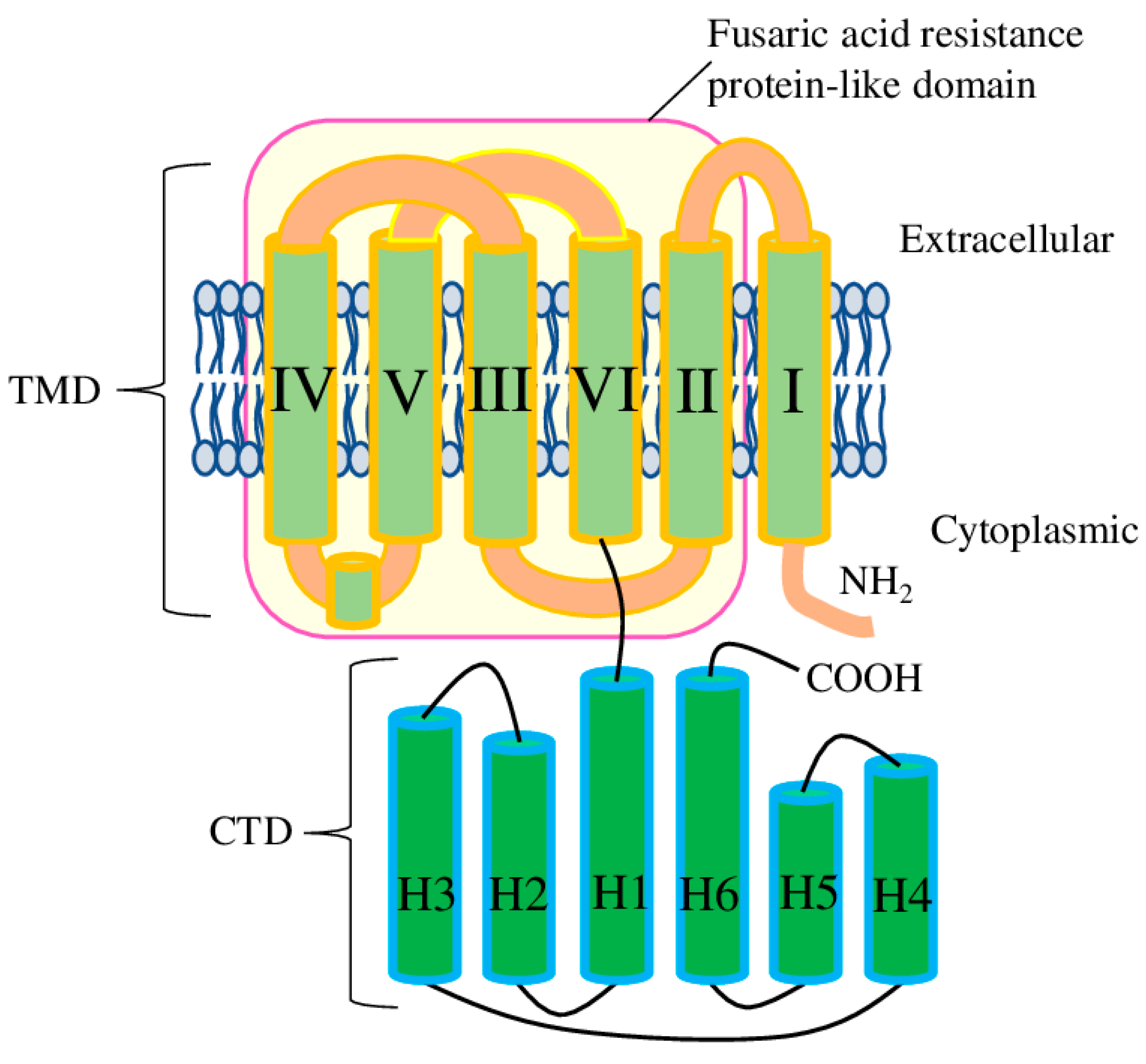
The common plant model organism A. thaliana has 14 ALMT genes which encode 16 proteins (ALMT14 has three isoforms). Examination of ALMTs at the amino acid level suggests the presence of several domains. The aluminium-activated malate transporter (pfam11744) domain (app 400aa) itself is the major component of the ALMT proteins. Further, the ALMT domain could be divided into the N-terminal pore-forming part (transmembrane domain (TMD)), which contains six transmembrane helices, and the C-terminal cytosolic domain (CTD) (Figure 1). ALMT11 has only a partial ALMT domain, and, despite it having well-conserved TM1 and TM2, its ability to transport malate or other substrates is still neither confirmed nor disproved. Therefore, ALMT11 is usually considered as a truncated gene which produces non-functional protein. Also, several ALMT proteins have a deletion in the CTD (in particular, ALMT3-6, 9, 12, and 13 and all isoforms of ALMT14). To the best of our knowledge, the functional significance of this deletion is still unknown (Supplementary Figure S1).
Additionally, another domain has been identified in AtALMTs with a high level of confidence (p-value 0.005 and higher) by NCBI conserved domain database search tool [13]. All ALMT proteins (except ALMT14 isoforms 2 and 3) have a fusaric acid resistance protein-like (pfam13515) domain, which ranges from TM2 to TM6, thus basically corresponding to the pore-forming part (Figure 1). Interestingly, the ALMT14-2 isoform almost completely lacks TM3, while ALMT14-3 lacks TM3 and partially lacks TM4. To the best of our knowledge, the functional properties of these isoforms have never been tested experimentally. Considering that TM3 and TM4 contain the important conservative residues necessary for malate recognition and transport, the functional role of ALMT14 isoforms 2 and 3 would be very interesting to investigate.
2.1. Malate and Al Recognition and Transport
Recently, the structures of Arabidopsis ALMT1 and Glycine max ((L.) Merr.) ALMT12 have been characterised. Also, residues participating in the recognition and transport of malate and Al were identified [14,15]. Besides residues involved in Al binding and coordination (Supplementary Figure S1) located on the TMD, several negatively charged Al-responsive residues were earlier identified on the CTD (Supplementary Figure S1). Interestingly, glutamate 276 (E276 for AtALMT12) was found to be crucial for channel activity, while in TaALMT1 and AtALMT1 (E284 and E256, respectively) this residue was spotted only as required for Al-induced malate transport [16,17].
Based on the conducted analysis (Supplementary Figure S1), residues involved in malate recognition are strongly conserved, while for Al-binding and Al-coordinating residues there were some discrepancies. Thus, conserved D59 (for AtALMT1) could be found only in ALMT1, 2, 7, and10; E156 was only found in ALMT1 and 7; D160 was only found in ALMT1 and 2; and Al coordination M60 was only found in ALMT1, 2, 7, and 10–14 (all isoforms of ALMT14). Additionally, several conservative sites have been identified in GmALMT12, which formed a positively charged ring at the pore entrance (R187 and R198) and a second positively charged ring within the membrane (K109, R113, and R158), mediated an interhelical network within the pore (E100, D168, and Y169), and stabilised the bottom of the T-shaped pore (K164 and K165). Also, the conserved W90 was found to act as a switcher, regulating channel gating (Supplementary Figure S1) [15].
2.2. ALMT/GABA Interplay
The role of GABA as an ALMT regulator and the role of ALMTs in GABA binding and transport has been studied in detail in recent years. The identified conserved GABA-binding motif [3] was shown to exhibit minor variation, which probably greatly affects the interaction of individual ALMTs with GABA. Thus, mutation of the first and second aromatic residues (F213 and F215 for TaALMT1) reduced interaction with and transport of GABA, while preserving positive activation by Al3+ and by external anions [3,18]. Interestingly, in Arabidopsis, only ALMT5 and 6 have two aromatic amino acids in these positions (both F) in addition to ALMT9 (F and Y). However, so far, no studies have confirmed the functional interchangeability of F and Y amino acids. Also, recent research suggests that AtALMT9 does not interact with GABA directly [19]. Furthermore, the significance of these and other sites in the GABA recognition motif involving ALMTs should be specified (for example, FFC in ALMT10 or LFF in ALMT12).
2.3. CaM Binding by ALMTs
Recent experiments on ALMT12 from Brachypodium distachyon, the closest transporter homologous to Arabidopsis thaliana ALMT12, have identified specific residues in the CTD which are responsible for binding with calmodulin (CaM). Thus, substitutions of R335A, R338A, and K342A greatly decreased the ability of malate to activate the channel and binding to CaM. Comparison of different substitution variants (triple and double) helped to emphasise the dominant role of the R335 site [20]. Interestingly, none of the Arabidopsis ALMTs has all three (R335, R338, and K342) conservative sites, while residues corresponding to R335 and K342 are rather well matched (Supplementary Figure S1).
2.4. ALMT Phosphorylation
Earlier research tested several potential protein kinase C (PKC)-mediated phosphorylation sites on TaALMT1 (S56, S183, Y184, T323, S324, S337, S351-352, and S384). However, only the S384A mutation resulted in a noticeable change in transport properties [21]. Recently, the role of ALMTs in abiotic stress response was demonstrated in experiments on plants subjected to drought stress and ABA treatments. Interestingly, AtALMT4 mediated malate efflux during the drought and ABA-induced stomatal closure depended on the phosphorylation of S382 [22]. This site corresponds to S384 in TaALMT1 and is rather conserved in Arabidopsis (except for ALMT9, ALMT6, and ALMT3) (Supplementary Figure S1). However, the significance of phosphorylation at this particular position for the regulation of other Arabidopsis ALMTs was not shown and remains a question for future experiments.
3. Molecular Evolution
3.1. Sequence Identification
To track the evolutionary origin of the ALMT, we have searched for homologous sequences in the Uniprot [23] (http://www.uniprot.org/ accessed on 25 April 2023), Pfam/InterPro [24,25] (http://www.ebi.ac.uk/interpro accessed on 25 April 2023) and NCBI databases [26] (http://www.ncbi.nlm.nih.gov/ accessed on 25 April 2023) using both the amino acid-based BLAST and the domain-based search methods [27]. During the sequence search, the truncated, partial, and identical sequences were removed. In total, 4650 sequences were identified (Viridiplantae: 4646; Amoebozoa: 3; Opisthokonta: 1). Many bacterial and archaea proteins have been annotated as containing an ALMT domain; however, closer examination disproved the presence of a full-length ALMT domain. Also, while AtALMT11 has no full-length ALMT domain, it was not used in the tree reconstruction.
3.2. Phylogenetic Analysis
To understand the evolutionary history of the ALMT proteins, we reconstruct the phylogenetic trees with the maximum likelihood method. Opisthokonta and Amoebozoa ALMT proteins were used as an out-group, and 240 ALMT proteins from different taxa were selected from the Viridiplantae group (Supplementary Figure S2). Because we used only sequences from full-length ALMT proteins, AtALMT11 was not used for tree reconstruction. In general, we can define eight separated clusters and the number of sub-clusters of closely related proteins (Supplementary Figure S2). Therefore, ALMTs from amoeba and Sphaeroforma arctica were located on a separate branch and counted as clade I. Clade II was comprised algae, mosses, and ferns (Chara braunii, Klebsormidium nitens, Physcomitrium patens, Ceratodon purpureus, and Marchantia polymorpha), while sub-clade IIa included only the fern Ceratopteris richardii. Clade III was well separated and formed by ALMT9-like proteins from plants of different taxa (including also both Eudicotyledons and Monocotyledons, such as Oryza species, Phaseolus vulgaris, and Cucurbita maxima) (Supplementary Figure S2).
Clade IV had three well-separated sub-clades, with IV formed by ALMT proteins from Eudicotyledon species (including A. thaliana ALMTs 12, 13, and 14), sub-clade IVa comprising only proteins from Monocotyledon species (such as Oryza sp., Triticum aestivum, Hordeum vulgare and others), and sub-clade IVb including ALMTs from several Eudicotyledon species and species from other taxa (Nicotiana tabacum, Solanum lycopersicum, Nelumbo nucifera, Cinnamomum micranthum, and others). Similarly, clade V was formed mostly by ALMTs from Eudicotyledon species (including A. thaliana ALMT10) and proteins from other taxa (Aquilegia coerulea, Thalictrum thalictroides, and Nelumbo nucifera). Clade Va contained ALMTs from Monocotyledon species, with ALMTs from Cinnamomum micranthum out-grouping the sub-clade consisting of Amborella trichopoda species, i.e., the entire Vth clade (Supplementary Figure S2).
Clades VI and VIa were composed of proteins from Eudicotyledon species, including AtALMTs (ALMT1, 2, and 7 for clade VI and ALMT8 for VIa). Sub-clade VIb was formed mostly by proteins from the Monocotyledon species, and ALMTs from Kalanchoe fedtschenkoi and Cinnamomum micranthum acted as out-groups for this sub-clade. The small sub-clade Vic contained ALMT proteins from different species (Thalictrum thalictroides, Nelumbo nucifera, and Tetracentron sinense). Clade VII had no sub-clade and contained ALMT proteins from Eudicotyledon species and species of other taxa (Prunus avium, Glycine max, Nelumbo nucifera, Cinnamomum micranthum, and others). The VIIIth clade had several well-separated sub-clades, with sub-clades VIII, VIIIa, and VIIIb formed only by ALMTs from Eudicotyledon species (including AtALMT4-6 belonging to VIII), while VIIIc contained only Monocotyledon species. Furthermore, VIIId included AtALMT3 and 9 and various ALMT proteins from different taxa (Eudicotyledons, Monocotyledons, and others). Finally, sub-clade VIIIe was formed by ALMT proteins from Aquilegia coerulea, Thalictrum thalictroides, Wollemia nobilis, and Araucaria cunninghamii. Also, some ALMT proteins (such as Amborella trichopoda, Selaginella moellendorffii, Marchantia polymorpha, and Ceratopteris richardii) were not assigned to any clade/sub-clade and acted as an out-group for corresponding clades and sub-clades (Supplementary Figure S2).
In total, our results confirmed and further expanded the phylogenetic tree published earlier [28,29], where only three “higher plant” species were used for tree reconstruction and Arabidopsis ALMTs were clustered into four groups: (1) ALMT8, 1, 2, and 7; (2) ALMT10; (3) ALMT12, 13, and 14; and (4) ALMT 9 and 3–6 [28]. Thus, our results further expanded these data, separating ALMT8 from ALMT1, 2, and 7 and separating ALMT3 and 9 from ALMT4-6 into different sub-clades (Supplementary Figure S2).
Here, we have to disprove some earlier reports [28,30] that propose that ALMTs are a plant-specific family. We found a full-length ALMT domain in one protein from Sphaeroforma arctica (Opisthokonta) and in three proteins from Amoeba Dictyostelium discoideum (Amoebozoa). Despite obvious differences because of the distant evolutional relation, most conservative sites are presented (Supplementary Figure S1). However, the role of these proteins in malate/GABA/Al3+ transport should be confirmed experimentally. Also, we disprove earlier reports linking ALMTs to the Aromatic Acid Exporter (ArAE) family because we found the ArAE domain only in AtALMT9 and 12 and a limited number of other species (such as A0A2R6WEU1 from Marchantia polymorpha and A0A1Y1HXE2 from Klebsormidium nitens). On the contrary, we can define ALMTs as part of a large group related to the fusaric acid resistance proteins (pfam13515) (Supplementary Figure S1). This group is widely presented in bacteria and fungi, and, considering the presence in other proteins with a fusaric acid resistance protein-like domain and a fusaric acid resistance protein family (PF04632) domain, such as in Arabidopsis and other species (importantly, without an ALMT domain), it is more plausible that ALMTs are part of or evolutionary originated from this group. Also, clear functional separation of the ALMT domain on the N-terminal pore-forming part (thus, acquired from fusaric acid resistance proteins) and the C-terminal CTD (which was, most probably, acquired later from another source and which performs a more pronounced regulatory role) further corroborates this hypothesis. However, this question requires further detailed investigation, which is far beyond the topic of this review.
In total, the results of our phylogenetic analysis confirmed the general tree topology reported earlier. At the same time, the position of individual ALMTs has been assigned to separate sub-clades. Because we have identified several non-plant ALMT proteins and the presence of the fusaric acid resistance domain in all ALMT proteins, we have disproved that ALMTs are a plant-specific protein family related to ArAE that possesses an ArAE domain. Therefore, the evolutionary origin of the ALMT family proteins requires further detailed investigation.
4. ALMTs in the Regulation of Stomata/Guard Cells
Anion channels localised on the plasma membrane of guard cells are the key contributors to stomata movement through the release of anions and subsequent membrane depolarisation [31]. Early research has demonstrated that two members of the ALMT family are localised to the tonoplast (AtALMT6 and AtALMT9, but not exclusively) and plasma membrane (AtALMT12) of the guard cells, thus suggesting the role of these channels in vacuolar malate transport or across the plasma membrane, respectively (Figure S1) [32,33,34]. Accordingly, AtALMT6 was characterised as a Ca2+-activated transporter, aiming a malate flux from the cytosol to the vacuole. However, Atalmt6 plants had no distinctive phenotype, possibly because of the functional redundancy of other ALMT-family transporters in guard cells [33]. AtALMT9 was defined as a Cl− channel, which is activated by cytosolic malate and involved in regulating stomata opening, albeit not closing. Interestingly, Atalmt9 plants showed a drought-resistant phenotype, which is congruent with impaired stomatal opening due to decreased uptake of Cl− in guard cells [35]. AtALMT12 was characterised as a quick anion channel, which possesses a high capacity for the transport of Cl−, NO3−, and malate from the guard cells and exhibits negligible reaction on extracellular Al3+. Furthermore, the Atalmt12 mutant inertly closes stomata in response to CO2, ABA, darkness, dehydration, or Ca2+. Thus, these data suggest the involvement of AtALMT12 in regulating stomata closure rather than in Al3+ resistance [32,36].
Recently, the Atalmt6 mutant phenotype was described with Cl−-dependent defective stomatal opening induced by blue light and fusicoccin, a fungal diterpenoid glycoside with a strong ability to activate plasma membrane H+-ATPase [37]. Furthermore, nucleotides were identified as effective modulators of AtALMT9 transport activity. Interestingly, ATP was the most effective cellular blocker and ATP hydrolysis was not required, suggesting that anion transport across the vacuolar membrane is regulated by cytosolic nucleotides and the energetic status of the cell [38]. Furthermore, under NaCl stress, Atalmt9 mutants showed reduced shoot accumulation of both Cl− and Na+, while plants complemented with the E196A channel variant (point mutation that exhibits increased Cl− current activity [39]) showed enhanced channel activity and higher Cl− and Na+ accumulation. Considering that NaCl up-regulated ALMT9 expression in the vasculature of shoots and roots, this suggests it has a crucial role in regulating the entire vacuolar Cl−-loading of the plant immediately after salt-stress is encountered [40]. The Atalmt12 plants responded with slower stomatal closure with a high CO2 concentration and in darkness but also accumulated higher levels of malate and fumarate and had improved mesophyll conductance. Interestingly, these effects were associated with increased photosynthesis and respiration rates, which resulted in improved growth [41]. Also, recent experiments on ALMT12 from the grass Brachypodium distachyon ((L.) P.Beauv.) (BdALMT12) demonstrated that both Ca2+/calmodulin and malate are co-regulators and required for channel activation (Figure 2) [20].
Plant stress hormone ABA is known to regulate plant drought stress adaptation through various pathways. In guard cells, ABA regulates AtALMT12 via kinase OPEN STOMATA 1 (OST1) and protein phosphatase ABSCISIC ACID-INSENSITIVE-1 (ABI1) [42]. Later experiments have defined that xylem-delivered sulphate could trigger drought-mediated stomata closure in several pathways (Figure 2). First, sulphate can reach guard cells and activate AtALMT12 directly. Second, sulphate in guard cells can induce the expression of 9-cis-epoxycarotenoid dioxygenase (NCED3), the key step in chloroplastic ABA biosynthesis. Third, sulphate can stimulate ABA biosynthesis through cysteine synthesis, thus promoting the activity of abscisic aldehyde oxidase 3 (AAO3)—the last step in ABA biosynthesis [43]. Furthermore, ABA regulates AtALMT4, another member of the ALMT family mediating Mal2- release from the vacuole in an ABA-dependent way. Atalmt4 plants demonstrated impaired stomatal closure in response to ABA and increased wilting in response to drought stress and ABA. Interestingly, the activity of AtALMT4 depended on phosphorylation on the C-terminal S382, which can be phosphorylated by mitogen-activated protein kinases in vitro. Dephosphomimetic mutants of AtALMT4 S382 showed increased channel activity and Mal2- efflux, while phosphomimetic mutants were electrically inactive and phenocopied the almt4 mutants [22].
Because ALMT proteins have a GABA-binding motif [3], it was suggested that GABA can directly regulate ALMT activity. Experiments on plant protoplasts and monkey kidney cells confirmed that GABA treatment resulted in AtALMT12 current reduction [44]. Interestingly, experiments on Triticum aestivum (L.) root tips and TaALMTs-expressing Xenopus laevis oocytes demonstrated that ALMTs can transport both GABA and anions in planta, as well as that an intact GABA-binding motif is crucial for its normal functioning [18]. Similarly, GABA was shown to reduce the anion channel opening frequency, thus regulating anions passing through the pore under stress [45]. Further research suggested that GABA can affect stomata aperture (thus regulating water use efficiency and drought resilience) by modulating the activity of ALMT family proteins [46]. However, it is most likely that GABA acted on ALMTs not directly, but through other unknown regulatory factors or vacuolar ion channels/transporters (Figure 2) [19]. This point of view is supported by the substantial R-type anion currents in the double (Atalmt12/13 and almt12/14) and triple (Atalmt12/13/14) mutants [47]. Indeed, our data analysis of co-localised, co-expressed, and domain-sharing proteins/genes of ALMTs suggested 27 various transporters which could potentially be involved in regulating stomata aperture. In this list, we can find some well-known transporters (like members of the ABC family, SLAH, SKOR, SWEET), as well as several unknown and uncharacterised transporters (Supplementary Table S1). Thus, further study of the interaction of ALMTs with other membrane transport proteins will perhaps shed light on the coordinated mechanism of transmembrane ion movement during stomata movement.
Therefore, the current model suggests the key role of vacuolar ALMTs during stomatal opening. When K+ enters the cells, starch is degraded to malate for the further compensation of charges [28]. As a result of starch degradation, the elevated cytosolic concentration of malate leads to the activation of ALMT9 combined with subsequent intensification of malate transport into the vacuole. In addition, ALMT9 is also activated to mediate the transport of Cl− into the vacuole and cause an accumulation of solutes in the vacuole of guard cells, with the final result being stomatal opening [28,35]. Furthermore, existing experimental data support the point of view that, as concerns the vacuolar chloride channel CLCa, the activity of AtALMT9 is crucial in stomata opening through the transport of Cl− into the vacuole [35,40,48,49].
5. Transcriptional Regulation
Regulation of the transcription activity of ALMT-encoded genes is the most crucial process in the physiological and biochemical adjustment of plants during growth and development and stress adaptation. One of the central roles in the transcriptional regulation of ALMT genes has been appointed to transcription factors (TFs) such as STOP1 (SENSITIVE TO PROTON RHIZOTOXICITY) (Figure 3). STOP1 belongs to C2H2-type TFs, and it was identified as a crucial regulator of Al-induced expression of the ALMT1 gene in Arabidopsis and other plant species [50,51]. Accordingly, the WRKY46 transcription factor was identified as a repressor of AtALMT1. Wrky46 plants demonstrated higher ALMT1 expression and increased malate exudation, with a subsequently higher tolerance to Al [52]. In the following years, the mechanism regulating transcriptional activation/repression of ALMT was further elucidated.
5.1. STOP1/CAMTA2/WRKY Regulatory Node
In planta complementation assays of AtALMT1 demonstrated that thepromoter region between −540 and the ATG codon contains eight cis elements essential for Al induction and STOP1 regulation. In particular, the region around the −297 site (cis-D) is recognised by four zinc finger domains of STOP1; thus, it is necessary for STOP1 binding. Some other cis elements have also been characterised, with cis-B shown to interact with an unknown repressor and cis-C with the activator CALMODULIN-BINDING TRANSCRIPTION ACTIVATOR 2 (CAMTA2). Accordingly, cis-H is crucial for the early phase, while cis-A and cis-C are important for the late phase of Al3+-stress response [53]. Also, calcineurin B-like protein (CBL) was shown to up-regulate ALMT1 expression and regulate root malate efflux [54]. CALMODULIN-LIKE24 (CML24) is another protein involved in Al-induced cytosolic Ca2+ signal transmission to regulate ALMT1 expression in a STOP1-independent way. Upon activation, CML24 interacted with CAMTA2 to promote ALMT1-mediated malate exudation from roots (Figure 3). Also, CML24 suppressed WRKY46, thus preventing its repression of ALMT1. Therefore, these results provide a novel player in Ca2+-mediated signalling in ALMT1-dependent Al stress resistance [55].
Interestingly, the application of phosphatidylinositol 4-kinase (PI4K) and phospholipase C (PLC) inhibitors suppressed AtALMT1 transcription and reduced Al-activated malate transport. Furthermore, AZD7762, a human protein kinase inhibitor, suppressed late-phase Al-induced AtALMT1 expression, presumably by inhibiting homologous calcineurin B-like protein (CBL)-interacting protein kinase (CIPK) and/or Ca-dependent protein kinase (CDPK). Also, AZD7762 induced the expression of WRKY46 (ALMT1 repressor) and suppressed CAMTA2 (an ALMT1 activator) (Figure 3). These data suggest that phosphatidylinositol metabolism is involved in regulating malate secretion in plants under Al stress [56].
Recent research has demonstrated that transgenic plants expressing the WRKY21 TF from soybean (Glycine max (L.) Merr.), when treated with AlCl3, showed higher induction of Al stress-associated genes (STOP1, ALMT1, MATE, and ALS3), along with up-regulated expression of other stress-related genes (from the COR, DREB, LEA, and RD families). Also, GmWRKY21-expressing transgenic Arabidopsis plants had increased root growth, lower MDA, and higher proline accumulation under AlCl3 stress. These results suggest that other TFs may promote tolerance to Al stress through the STOP1–ALMT1 node as a part of the general abiotic stress response [57].
5.2. STOP1/STOP2 Pathway
Recently, analysis of GFP-tagged STOP1 proteins helped to identify other genes rapidly induced upon Al treatment and containing STOP1-binding sites, such as STOP2 and GLUTAMATE-DEHYDROGENASE1 and 2 (GDH1 and 2). Furthermore, the GDH1 and GDH2 genes were suppressed in stop1 mutant lines treated with AlCl3, and the gdh1/2 double mutant showed increased sensitivity to Al stress, suggesting the involvement of both genes in Al tolerance in a STOP1-dependent way (Figure 3) [58]. Indeed, the role of STOP2, a STOP1 homologue, in Al stress tolerance and low pH was demonstrated in Arabidopsis. Expression of STOP2 in stop1 plants partially rescued growth and root tip viability by activating the expression of some genes regulated by STOP1. Thus, polygalacturonase-inhibiting protein 1 and 2 (PGIP1 and PGIP2) genes, which are known to stabilise cell walls at low pH, were activated, while the expression of AtALMT1 was not recovered [59].
Recently, an MADS-box TF from soybean (GsMAS1) was shown to participate in Al stress tolerance. Accordingly, the expression of GsMAS1 in Arabidopsis enhanced tolerance to Al stress, improved root length, and increased proline accumulation under AlCl3 treatment. In particular, STOP1, ALMT1, STOP2, AtMATE, and PGIP2 were the key genes activated by GsMAS1, suggesting that it may increase resistance to Al stress through several pathways in Arabidopsis [60].
5.3. STOP1/Phosphate Interplay
The low level of external phosphate is another factor regulating the transcription of STOP1 and its direct target ALMT1. Phenotypically, limited phosphate (lPi) supply is manifested by inhibiting primary root growth in many plant species. Mechanically, lPi enhances expression ALMT1, which subsequently promotes quick primary root growth arrest together with LOW PHOSPHATE ROOT (LPR) ferroxidases and peroxidase-dependent cell wall stiffening (Figure 4). Furthermore, the accumulation of Fe and callose in the stem cell niche inhibited cell proliferation while accelerating cell differentiation, which resulted in the meristem’s reduction. Importantly, malate exudation is crucial for Fe accumulation in the apoplast of meristematic cells and the subsequent peroxidase-dependent activation differentiation of meristematic cells in response to lPi supply [61,62]. Later research defined that the tonoplast-localised ALUMINIUM-SENSITIVE3/SENSITIVE TO ALUMINIUM RHIZOTOXICITY1 transporter complex (ALS3/STAR1) repressed nuclear STOP1 accumulation, thus inhibiting the STOP1–ALMT1 pathway [63]. Furthermore, Al3+ can act similarly to Fe and inhibit proteasomal degradation of STOP1, thus facilitating its stability, accumulation in the nucleus, and ALMT1 expression [64].
The MEDIATOR (MED) complex is a multi-protein assembly that controls gene expression through interaction with basal RNA polymerase II transcription machinery, cis regulatory elements, and enhancers. The MED16 subunit was shown to regulate plant development and response to multiple biotic and abiotic stresses, including Fe uptake (Figure 4) [65]. As was recently shown, MED16 is also involved in lPi response through direct interaction with STOP1 and co-regulation of STOP1-regulated genes. Accordingly, the expression of ALMT1 was greatly reduced in med16 plants in both conditions (lPi and normal Pi levels). These data add another transcriptional activator to the STOP1–ALMT1 node, further linking Fe uptake, lPi, and Al stress response to malate efflux [66].
Recently, the AtWRKY33 TF was identified as a negative regulator of lPi-induced root architecture remodelling (Figure 4). Atwrky33 plants showed increased sensitivity to lPi via inhibition of primary root growth and the promotion of root hair formation. In particular, AtWRKY33 negatively regulated ALMT1 transcription (but not STOP1) under lPi conditions, thus mediating Fe3+ accumulation in root tips to inhibit root growth [67]. Also, lPi down-regulated BRASSINAZOLE-RESISTANT 1 (BZR1) expression in A. thaliana, a crucial regulator of the brassinosteroid signalling pathway [68]. As it was shown on Nicotiana benthamiana and Oryza sativa, nuclear-localised BZR1 competed with STOP1, thus suppressing ALMT1 activation and malate secretion [69].
Furthermore, lPi inhibited primary root growth via post-transcriptional regulation. Particularly, under lPi conditions, histone H3 acetylation was increased, and HDC1 protein abundance was reduced because of 26S proteasome-mediated degradation. At the same time, the histone deacetylase complex 1 (hdc1) plants were hypersensitive to Pi deficiency, with inhibited primary root growth and an increased number of root hairs (Figure 4). Also, under lPi conditions, hdc1 plants accumulated a higher amount of Fe3+ in the root tips and showed increased expression of ALMT1, STOP1, PHOSPHATE TRANSPORTER1, and LPR1 and 2. Additionally, lPi conditions enriched the histone H3 acetylation of ALMT1 and LPR1. These results suggest a new chromatin-level control mechanism via repression of the STOP1–ALMT1 node and associated genes in response to lPi conditions through HDC1-mediated histone H3 deacetylation [70].
5.4. TFs Regulating ALMTs in Berries/Fruits
Despite the role in Al stress and lPi response, malate greatly affects the palatability of the edible tissues of many agricultural species, such as apples, tomatoes, pears, kiwis, and others [71]. Several recent papers have defined different TFs regulating malate accumulation in the fruits of several species. Thus, in apple (Malus domestica Borkh.), MdMYB73 was found to activate MdALMT9, vacuolar ATPase subunit A (MdVHA-A), and vacuolar pyrophosphatase 1 (MdVHP1). Interestingly, cold-induced bHLH1 (MdCIbHLH1) was also shown to interact with MdMYB73 and enhanced its activity, thus modulating vacuolar malate accumulation and pH in apple [72].
Furthermore, the member of the plant-specific BTB-TAZ DOMAIN PROTEIN 2 (BT2), which contains an N-terminal bric-a-brac/tramtrack/broad complex (BTB), a transcriptional adapter zinc finger (TAZ) domain, and a C-terminal calmodulin-binding (CaMB) domain, was shown to regulate malate accumulation and vacuolar pH in response to nitrate. Thus, in response to nitrate treatment, MdBT2 directly interacted with and ubiquitinated MdCIbHLH1 and MdMYB73 through the ubiquitin/26S proteasome pathway in both in vitro and in vivo conditions. Subsequently, degradation of the MdCIbHLH1 and MdMYB73 proteins reduced the levels of downstream genes, such as MdVHA-A, MdVHP1, and MdALMT9. These results provided the direct link between high nitrate supply and malate accumulation and vacuolar acidification in apples [73,74].
5.5. Regulation of STOP1 Proteasomal Degradation
STOP1 Ubiquitination. Interestingly, STOP1 regulated its degradation by up-regulating the RAE1 gene (regulation of Atalmt1 expression 1), which ubiquitinated and degraded STOP1, thus forming a negative feedback loop between STOP1 and RAE1 (Figure 4) [75]. Later, similar biological functions were assigned to RAE1 homologue 1 (RAH1), which can also directly interact with STOP1 and promote its ubiquitination and degradation. As expected, double mutant rah1rae1 shows reduced plant growth, while introduction of the stop1 mutation (i.e., rah1rae1stop1) rescued plant growth [76].
STOP1-THO/TREX complex. Recent research has established the involvement of the THO/TREX complex in the regulation of nucleocytoplasmic STOP1 mRNA export, thus suggesting its regulation at a post-transcriptional level [77]. THO/TREX complex is a conserved eukaryotic multi-protein complex regulating transcription, mRNA processing, and export [78]. Thus, a mutant of hyperrecombination protein 1 (hpr1), a subunit of the THO/TREX complex, demonstrated reduced expression of STOP1-regulated genes and the associated Al resistance. STOP1 mRNA was retained in the nucleus, which further resulted in decreased STOP1 protein abundance. Interestingly, the introduction of the rae1 mutation to the hpr1 background (i.e., hpr1rae1 double mutant) partially rescued the reduced Al resistance and limited phosphate (lPi) response in single hpr1 mutants [77]. Similar biological functions have also been described for TEX1—another subunit of the THO/TREX complex. Mutant tex1 plants exhibited reduced expression of STOP1-regulated genes, including AtALMT1. Unlike hpr1, tex1 plants had no STOP1 mRNA accumulation in the nucleus; however, these plants had reduced levels of STOP1 protein. Similarly, double mutants (tex1rae1) showed partially rescued Al resistance and lPi response phenotypes in comparison to single tex1 plants [79]. In total, these results demonstrated that HPR1 and TEX1 have overlapping but distinct roles in the modulation of Al resistance and lPi response through regulation of the expression of STOP1 downstream genes.
STOP1 SUMOylation. SUMOylation (SMALL UBIQUITIN-LIKE MODIFIER) is the process of reversible conjunction of SUMO to target proteins to modulate their function, stability, localisation, and/or activity [80]. Specifically, ELS-type SUMO protease 4 (AtESD4) was demonstrated to interact and deSUMOylate STOP1. esd4 plants had increased level of STOP1 SUMOylation, which caused its increased association with the AtALMT1 promoter and, subsequently, enhanced AtALMT1 expression. Three sites for SUMOylation were identified (K40, K212, and K395), and inhibition of STOP1 SUMOylation reduced STOP1 stability and the expression of STOP1 downstream genes [81]. Similarly, E3 SUMO ubiquitin ligase (AtSIZ1) is one of the proteins responsible for STOP1 SUMOylation. siz1 plants showed reduced levels of both STOP1 SUMOylation and STOP1 protein, while the expression of STOP1-regulated genes (including ALMT1) was increased. Interestingly, siz1 plants demonstrated increased Al stress resistance and lPi response, which suggests STOP1-independent regulation of AtALMT1 expression [82].
5.6. The Role of Jasmonic Acid in ALMT Regulation
Jasmonic acid (JA) is another player involved in the regulation of Al stress-induced root growth inhibition. The expression of the major JA receptor CORONATINE-INSENSITIVE1 (COI1) and the signalling regulator bHLH-III-type transcription factor (MYC2) was up-regulated in response to Al stress in the root tips. Accordingly, coi1 and myc2 plants showed reduced root growth inhibition in response to Al stress, while exogenous JA (in the form of methyl jasmonate) enhanced the Al-induced inhibition of root growth in WT plants. Interestingly, WT and coi1 plants reacted to the treatment with other toxins (La3+, Cd2+, Cu2+, and Na+) in the same way. Furthermore, the expression level of ALMT1 was two times higher in the roots of coi1 plants, while the expression of other related genes (STOP1, MATE, and ALS1 and 3) was not changed. Subsequently, malate exudation was higher in coi1 plants after Al treatment. However, ALMT1 expression and the level of malate exudation after Al stress were not affected in the myc2 plants. These data provide an additional JA-mediated signalling pathway regulating root growth inhibition in response to Al stress [83].
Recently, these results have been further elaborated in experiments on tomato (Solanum lycopersicum L.) where the crosstalk between JA and Al stress was analysed. Al stress up-regulated genes related to JA biosynthesis and signalling (such as OXOPHYTODIENOIC ACID REDUCTASE 3, ALLENE OXIDE CYCLASE, COI1, and MYC2), which further enhanced Al-induced root growth inhibition. Interestingly, both Al stress and JA treatment up-regulated several WRKY TFs (3, 6, 16, 37, 39, and 71) and ALMTs (3, 6, and 7), which were further analysed in MYC2-silenced and JASMONIC ACID-INSENSITIVE 1 (jai1) plants. The obtained results suggest that Al stress and JA cross-talk in root growth inhibition is mediated via SlALMT3 and six SlWRKY TFs as its upstream regulators [84].
In total, these data suggest a very complex and sophisticated network regulating STOP1 and ALMTs expression and, subsequently, Al stress resistance and lPi response. This network includes various TFs, hormones (such as GABA, JA, and ABA), and post-translational modifications (phosphorylation, SUMOylation, and ubiquitination) of the regulatory proteins. Despite the recent progress in our understanding of STOP1/ALMT regulation, further research in this direction may help to produce plants that are more tolerant to drought stress, acid, and Al-contaminated and low-phosphorus soils and more productive under adverse environmental conditions.
6. Conclusions and Future Prospects
Recent studies confirm the complex regulation of the ALMT family of proteins. In addition to the STOP1 TF, the expression and functioning of ALMTs are also regulated by plant hormones (primarily by GABA, but also JA and possibly others), ROS, metals (such as Al, Fe, and probably others), phosphorus availability in soil, and various post-translational modifications. Furthermore, the expression of ALMT genes affects plant immunity, proton tolerance, root development, and tolerance to abiotic stress factors. Further studies should characterise the molecular mechanisms underlying the crosstalk between these ALMT-regulating components.
Based on the discussed literature and the bioinformatic and evolutionary analysis conducted, we can formulate several directions for future investigation:
- The functional role of ALMT11 is still unknown. While it misses most conserved residues required for Al/malate/GABA recognition and transport, the presented sequence similarity (especially in the TM1 region) may be sufficient to form heterodimers with other ALMTs and, thus, deactivate them to act as a negative regulator.
- Currently, the role of ALMT14 isoforms 2 and 3 is unknown. Therefore, any research investigating the functions of these proteins would be beneficial, particularly their ability to recognise/transport Al3+ ions and/or malate and the ability to form homo/hetero dimers with other ALMTs. Recent results on full-length and truncated versions of rice (Oryza sativa L.) and wheat ALMTs suggest that they function as multimeric proteins, where combinations of ALMT subunits can affect channel function [85]. Similar experiments on ALMT11 and ALMT14 would greatly advance our understanding of ALMT functionality.
- The deletion in the CTD has been identified in several ALMTs. It would be interesting to identify how this deletion affects Al3+/malate recognition/transport function, or how it correlates with protein localisation and stability.
- The presence of the fusaric acid resistance protein-like (pfam13515) domain as the core of the TMD suggests that it may be the original form of the ALMT protein (before it acquired the CTD). So far, the ability of other Arabidopsis proteins possessing the fusaric acid resistance protein-like (pfam13515) domain to recognise/transport Al3+ or malate has not been studied.
- We have discussed several plant hormones interacting and regulating the STOP1/ALMT pathway of Al stress and lPi response (GABA, JA, and ABA). However, analysis of the ALMT interactome suggests that other hormones may be involved. For example, ALMT1 was shown to interact with Arabidopsis histidine kinase 4, a cytokinin receptor [86], thus suggesting possible regulation by the cytokinin.
- Also, ALMT1 interacts with a cell wall-associated receptor-like protein kinase (WAK1) [87], which is known as a receptor of oligogalacturonides, is involved in wounding response, and is a regulator of cell wall synthesis.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants12173167/s1, Supplementary Figure S1: The alignment of selected ALMT protein sequences. Brown and blue bars represent transmembrane helices (1 to 6) and fusaric acid resistance protein-like (pfam13515) domains, respectively. Green bars represent ALMT proteins where the ArAE_1 domain signature was identified. Magenta, green, and blue squares depict the gap in the ALMT sequences, GABA-binding motif, and crucial Glutamate residues, respectively. Dots of different colours represent various residues, important for the channel’s activity, regulation, and substrate selectivity. Multiple sequence alignments of ALMT domain sequences were performed using MUSCLE [88] with default settings in UGENE [89] software; Supplementary Figure S2: Phylogeny estimation of the ALMT proteins. Full-length proteins from selected taxa were used for phylogeny reconstruction with the maximum likelihood method [90] and JTT [91] substitution model in the MEGAX [92] program. For the maximum likelihood tree, the JTT substitution model was selected, assuming an estimated proportion of invariant sites and four gamma-distributed rate categories to account for rate heterogeneity across sites. The gamma shape parameter was estimated directly from the data. Reliability for the internal branch was assessed using the bootstrapping method (1000 bootstrap replicates); Supplementary Table S1: List of transporters that are co-localised, co-expressed, and/or share domains with ALMTs.
Author Contributions
Conceptualization, methodology, formal analysis: S.A.D. and S.V.I.; writing—original draft preparation: S.A.D.; supervision: S.V.I.; writing—review and editing: S.A.D. and S.V.I. All authors have read and agreed to the published version of the manuscript.
Funding
This research was partially supported by a Philipp Schwartz Fellowship from the Alexander von Humboldt Foundation.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Used sequences may be provided upon request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kobayashi, Y.; Kobayashi, Y.; Sugimoto, M.; Lakshmanan, V.; Iuchi, S.; Kobayashi, M.; Bais, H.P.; Koyama, H. Characterization of the Complex Regulation of AtALMT1 Expression in Response to Phytohormones and Other Inducers. Plant Physiol. 2013, 162, 732–740. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, Y.; Kobayashi, Y.; Watanabe, T.; Shaff, J.E.; Ohta, H.; Kochian, L.V.; Wagatsuma, T.; Kinraide, T.B.; Koyama, H. Molecular and Physiological Analysis of Al3+ and H+ Rhizotoxicities at Moderately Acidic Conditions. Plant Physiol. 2013, 163, 180–192. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, S.A.; Tyerman, S.D.; Xu, B.; Bose, J.; Kaur, S.; Conn, V.; Domingos, P.; Ullah, S.; Wege, S.; Shabala, S.; et al. GABA Signalling Modulates Plant Growth by Directly Regulating the Activity of Plant-Specific Anion Transporters. Nat. Commun. 2015, 6, 7879. [Google Scholar] [CrossRef]
- Liang, C.; Piñeros, M.A.; Tian, J.; Yao, Z.; Sun, L.; Liu, J.; Shaff, J.; Coluccio, A.; Kochian, L.V.; Liao, H. Low pH, Aluminum, and Phosphorus Coordinately Regulate Malate Exudation through GmALMT1 to Improve Soybean Adaptation to Acid Soils. Plant Physiol. 2013, 161, 1347–1361. [Google Scholar] [CrossRef] [PubMed]
- Tian, Q.; Zhang, X.; Ramesh, S.; Gilliham, M.; Tyerman, S.D.; Zhang, W.-H. Ethylene Negatively Regulates Aluminium-Induced Malate Efflux from Wheat Roots and Tobacco Cells Transformed with TaALMT1. J. Exp. Bot. 2014, 65, 2415–2426. [Google Scholar] [CrossRef]
- Maruyama, H.; Sasaki, T.; Yamamoto, Y.; Wasaki, J. AtALMT3 Is Involved in Malate Efflux Induced by Phosphorus Deficiency in Arabidopsis thaliana Root Hairs. Plant Cell Physiol. 2019, 60, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Neuhäuser, B.; Neumann, G.; Ludewig, U. LaALMT1 Mediates Malate Release from Phosphorus-Deficient White Lupin Root Tips and Metal Root to Shoot Translocation. Plant Cell Environ. 2020, 43, 1691–1706. [Google Scholar] [CrossRef]
- Fan, W.; Xu, J.M.; Wu, P.; Yang, Z.X.; Lou, H.Q.; Chen, W.W.; Jin, J.F.; Zheng, S.J.; Yang, J.L. Alleviation by Abscisic Acid of Al Toxicity in Rice Bean Is Not Associated with Citrate Efflux but Depends on ABI5-Mediated Signal Transduction Pathways: Cross-Talk between Al and ABA Signals. J. Integr. Plant Biol. 2019, 61, 140–154. [Google Scholar] [CrossRef]
- Guo, S.-H.; Jiang, L.-Y.; Xu, Z.-M.; Li, Q.-S.; Wang, J.-F.; Ye, H.-J.; Wang, L.-L.; He, B.-Y.; Zhou, C.; Zeng, E.Y. Biological Mechanisms of Cadmium Accumulation in Edible Amaranth (Amaranthus mangostanus L.) Cultivars Promoted by Salinity: A Transcriptome Analysis. Environ. Pollut. 2020, 262, 114304. [Google Scholar] [CrossRef]
- Zarei, A.; Chiu, G.Z.; Yu, G.; Trobacher, C.P.; Shelp, B.J. Salinity-Regulated Expression of Genes Involved in GABA Metabolism and Signaling. Botany 2017, 95, 621–627. [Google Scholar] [CrossRef]
- Pereira, P.N.; Gaspar, M.; Smith, J.A.C.; Mercier, H. Ammonium Intensifies CAM Photosynthesis and Counteracts Drought Effects by Increasing Malate Transport and Antioxidant Capacity in Guzmania Monostachia. J. Exp. Bot. 2018, 69, 1993–2003. [Google Scholar] [CrossRef] [PubMed]
- Kamran, M.; Ramesh, S.A.; Gilliham, M.; Tyerman, S.D.; Bose, J. Role of TaALMT1 malate-GABA Transporter in Alkaline pH Tolerance of Wheat. Plant Cell Environ. 2020, 43, 2443–2459. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Derbyshire, M.K.; Gonzales, N.R.; Lu, S.; Chitsaz, F.; Geer, L.Y.; Geer, R.C.; He, J.; Gwadz, M.; Hurwitz, D.I.; et al. CDD: NCBI’s Conserved Domain Database. Nucleic Acids Res. 2015, 43, D222–D226. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yu, X.; Ding, Z.J.; Zhang, X.; Luo, Y.; Xu, X.; Xie, Y.; Li, X.; Yuan, T.; Zheng, S.J.; et al. Structural Basis of ALMT1-Mediated Aluminum Resistance in Arabidopsis. Cell Res. 2022, 32, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Tang, L.; Xu, J.; Zhang, X.; Zhu, Y.; Zhang, C.; Wang, M.; Liu, X.; Li, F.; Sun, F.; et al. Cryo-EM Structure and Electrophysiological Characterization of ALMT from Glycine max Reveal a Previously Uncharacterized Class of Anion Channels. Sci. Adv. 2022, 8, eabm3238. [Google Scholar] [CrossRef]
- Mumm, P.; Imes, D.; Martinoia, E.; Al-Rasheid, K.A.S.; Geiger, D.; Marten, I.; Hedrich, R. C-Terminus-Mediated Voltage Gating of Arabidopsis Guard Cell Anion Channel QUAC1. Mol. Plant 2013, 6, 1550–1563. [Google Scholar] [CrossRef] [PubMed]
- Ligaba, A.; Dreyer, I.; Margaryan, A.; Schneider, D.J.; Kochian, L.; Piñeros, M. Functional, Structural and Phylogenetic Analysis of Domains Underlying the Al Sensitivity of the Aluminum-Activated Malate/Anion Transporter, TaALMT1. Plant J. 2013, 76, 766–780. [Google Scholar] [CrossRef]
- Ramesh, S.A.; Kamran, M.; Sullivan, W.; Chirkova, L.; Okamoto, M.; Degryse, F.; McLaughlin, M.; Gilliham, M.; Tyerman, S.D. Aluminum-Activated Malate Transporters Can Facilitate GABA Transport. Plant Cell 2018, 30, 1147–1164. [Google Scholar] [CrossRef]
- Jaślan, J.; De Angeli, A. Heterologous Expression Reveals That GABA Does Not Directly Inhibit the Vacuolar Anion Channel At ALMT9. Plant Physiol. 2022, 189, 469–472. [Google Scholar] [CrossRef]
- Luu, K.; Rajagopalan, N.; Ching, J.C.H.; Loewen, M.C.; Loewen, M.E. The Malate-Activated ALMT12 Anion Channel in the Grass Brachypodium distachyon Is Co-Activated by Ca2+/Calmodulin. J. Biol. Chem. 2019, 294, 6142–6156. [Google Scholar] [CrossRef]
- Ligaba, A.; Kochian, L.; Piñeros, M. Phosphorylation at S384 Regulates the Activity of the TaALMT1 Malate Transporter That Underlies Aluminum Resistance in Wheat. Plant J. 2009, 60, 411–423. [Google Scholar] [CrossRef]
- Eisenach, C.; Baetz, U.; Huck, N.V.; Zhang, J.; De Angeli, A.; Beckers, G.J.M.; Martinoia, E. ABA-Induced Stomatal Closure Involves ALMT4, a Phosphorylation-Dependent Vacuolar Anion Channel of Arabidopsis. Plant Cell 2017, 29, 2552–2569. [Google Scholar] [CrossRef]
- The UniProt Consortium; Bateman, A.; Martin, M.-J.; Orchard, S.; Magrane, M.; Agivetova, R.; Ahmad, S.; Alpi, E.; Bowler-Barnett, E.H.; Britto, R.; et al. UniProt: The Universal Protein Knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam Protein Families Database in 2019. Nucleic Acids Res. 2019, 47, D427–D432. [Google Scholar] [CrossRef]
- Mitchell, A.L.; Attwood, T.K.; Babbitt, P.C.; Blum, M.; Bork, P.; Bridge, A.; Brown, S.D.; Chang, H.-Y.; El-Gebali, S.; Fraser, M.I.; et al. InterPro in 2019: Improving Coverage, Classification and Access to Protein Sequence Annotations. Nucleic Acids Res. 2019, 47, D351–D360. [Google Scholar] [CrossRef]
- Sayers, E.W.; Bolton, E.E.; Brister, J.R.; Canese, K.; Chan, J.; Comeau, D.C.; Connor, R.; Funk, K.; Kelly, C.; Kim, S.; et al. Database Resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2022, 50, D20–D26. [Google Scholar] [CrossRef]
- Boratyn, G.M.; Schäffer, A.A.; Agarwala, R.; Altschul, S.F.; Lipman, D.J.; Madden, T.L. Domain Enhanced Lookup Time Accelerated BLAST. Biol. Direct 2012, 7, 12. [Google Scholar] [CrossRef]
- Sharma, T.; Dreyer, I.; Kochian, L.; Piñeros, M.A. The ALMT Family of Organic Acid Transporters in Plants and Their Involvement in Detoxification and Nutrient Security. Front. Plant Sci. 2016, 7, 1488. [Google Scholar] [CrossRef]
- Dreyer, I.; Gomez-Porras, J.L.; Riaño-Pachón, D.M.; Hedrich, R.; Geiger, D. Molecular Evolution of Slow and Quick Anion Channels (SLACs and QUACs/ALMTs). Front. Plant Sci. 2012, 3, 263. [Google Scholar] [CrossRef] [PubMed]
- Daspute, A.A.; Sadhukhan, A.; Tokizawa, M.; Kobayashi, Y.; Panda, S.K.; Koyama, H. Transcriptional Regulation of Aluminum-Tolerance Genes in Higher Plants: Clarifying the Underlying Molecular Mechanisms. Front. Plant Sci. 2017, 8, 1358. [Google Scholar] [CrossRef]
- Sussmilch, F.C.; Roelfsema, M.R.G.; Hedrich, R. On the Origins of Osmotically Driven Stomatal Movements. New Phytol. 2019, 222, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Meyer, S.; Mumm, P.; Imes, D.; Endler, A.; Weder, B.; Al-Rasheid, K.A.S.; Geiger, D.; Marten, I.; Martinoia, E.; Hedrich, R. AtALMT12 Represents an R-Type Anion Channel Required for Stomatal Movement in Arabidopsis Guard Cells: AtALMT12-Mediated Release of Anions in Guard Cells. Plant J. 2010, 63, 1054–1062. [Google Scholar] [CrossRef] [PubMed]
- Meyer, S.; Scholz-Starke, J.; De Angeli, A.; Kovermann, P.; Burla, B.; Gambale, F.; Martinoia, E. Malate Transport by the Vacuolar AtALMT6 Channel in Guard Cells Is Subject to Multiple Regulation: AtALMT6 Mediates Malate Transport in Guard Cells. Plant J. 2011, 67, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Kovermann, P.; Meyer, S.; Hörtensteiner, S.; Picco, C.; Scholz-Starke, J.; Ravera, S.; Lee, Y.; Martinoia, E. The Arabidopsis Vacuolar Malate Channel Is a Member of the ALMT Family. Plant J. 2007, 52, 1169–1180. [Google Scholar] [CrossRef]
- De Angeli, A.; Zhang, J.; Meyer, S.; Martinoia, E. AtALMT9 Is a Malate-Activated Vacuolar Chloride Channel Required for Stomatal Opening in Arabidopsis. Nat. Commun. 2013, 4, 1804. [Google Scholar] [CrossRef]
- Sasaki, T.; Mori, I.C.; Furuichi, T.; Munemasa, S.; Toyooka, K.; Matsuoka, K.; Murata, Y.; Yamamoto, Y. Closing Plant Stomata Requires a Homolog of an Aluminum-Activated Malate Transporter. Plant Cell Physiol. 2010, 51, 354–365. [Google Scholar] [CrossRef]
- Ye, W.; Koya, S.; Hayashi, Y.; Jiang, H.; Oishi, T.; Kato, K.; Fukatsu, K.; Kinoshita, T. Identification of Genes Preferentially Expressed in Stomatal Guard Cells of Arabidopsis Thaliana and Involvement of the Aluminum-Activated Malate Transporter 6 Vacuolar Malate Channel in Stomatal Opening. Front. Plant Sci. 2021, 12, 744991. [Google Scholar] [CrossRef]
- Zhang, J.; Martinoia, E.; De Angeli, A. Cytosolic Nucleotides Block and Regulate the Arabidopsis Vacuolar Anion Channel AtALMT9. J. Biol. Chem. 2014, 289, 25581–25589. [Google Scholar] [CrossRef]
- Zhang, J.; Baetz, U.; Krügel, U.; Martinoia, E.; De Angeli, A. Identification of a Probable Pore-Forming Domain in the Multimeric Vacuolar Anion Channel AtALMT9. Plant Physiol. 2013, 163, 830–843. [Google Scholar] [CrossRef]
- Baetz, U.; Eisenach, C.; Tohge, T.; Martinoia, E.; De Angeli, A. Vacuolar Chloride Fluxes Impact Ion Content and Distribution during Early Salinity Stress. Plant Physiol. 2016, 172, 1167–1181. [Google Scholar] [CrossRef]
- Medeiros, D.B.; Martins, S.C.V.; Cavalcanti, J.H.F.; Daloso, D.M.; Martinoia, E.; Nunes-Nesi, A.; DaMatta, F.M.; Fernie, A.R.; Araújo, W.L. Enhanced Photosynthesis and Growth in Atquac1 Knockout Mutants Are Due to Altered Organic Acid Accumulation and an Increase in Both Stomatal and Mesophyll Conductance. Plant Physiol. 2016, 170, 86–101. [Google Scholar] [CrossRef] [PubMed]
- Imes, D.; Mumm, P.; Böhm, J.; Al-Rasheid, K.A.S.; Marten, I.; Geiger, D.; Hedrich, R. Open Stomata 1 (OST1) Kinase Controls R-Type Anion Channel QUAC1 in Arabidopsis Guard Cells. Plant J. 2013, 74, 372–382. [Google Scholar] [CrossRef]
- Malcheska, F.; Ahmad, A.; Batool, S.; Müller, H.M.; Ludwig-Müller, J.; Kreuzwieser, J.; Randewig, D.; Hänsch, R.; Mendel, R.R.; Hell, R.; et al. Drought-Enhanced Xylem Sap Sulfate Closes Stomata by Affecting ALMT12 and Guard Cell ABA Synthesis. Plant Physiol. 2017, 174, 798–814. [Google Scholar] [CrossRef] [PubMed]
- Domingos, P.; Dias, P.N.; Tavares, B.; Portes, M.T.; Wudick, M.M.; Konrad, K.R.; Gilliham, M.; Bicho, A.; Feijó, J.A. Molecular and Electrophysiological Characterization of Anion Transport in Arabidopsis Thaliana Pollen Reveals Regulatory Roles for pH, Ca 2+ and GABA. New Phytol. 2019, 223, 1353–1371. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.; Tyerman, S.D.; Gilliham, M. Cytosolic GABA Inhibits Anion Transport by Wheat ALMT1. New Phytol. 2020, 225, 671–678. [Google Scholar] [CrossRef]
- Xu, B.; Long, Y.; Feng, X.; Zhu, X.; Sai, N.; Chirkova, L.; Betts, A.; Herrmann, J.; Edwards, E.J.; Okamoto, M.; et al. GABA Signalling Modulates Stomatal Opening to Enhance Plant Water Use Efficiency and Drought Resilience. Nat. Commun. 2021, 12, 1952. [Google Scholar] [CrossRef]
- Jaślan, J.; Marten, I.; Jakobson, L.; Arjus, T.; Deeken, R.; Sarmiento, C.; De Angeli, A.; Brosché, M.; Kollist, H.; Hedrich, R. ALMT-independent Guard Cell R-type Anion Currents. New Phytol. 2023, 239, 2225–2234. [Google Scholar] [CrossRef]
- Wege, S.; De Angeli, A.; Droillard, M.-J.; Kroniewicz, L.; Merlot, S.; Cornu, D.; Gambale, F.; Martinoia, E.; Barbier-Brygoo, H.; Thomine, S.; et al. Phosphorylation of the Vacuolar Anion Exchanger AtCLCa Is Required for the Stomatal Response to Abscisic Acid. Sci. Signal. 2014, 7, ra65. [Google Scholar] [CrossRef]
- Eisenach, C.; De Angeli, A. Ion Transport at the Vacuole during Stomatal Movements. Plant Physiol. 2017, 174, 520–530. [Google Scholar] [CrossRef]
- Iuchi, S.; Koyama, H.; Iuchi, A.; Kobayashi, Y.; Kitabayashi, S.; Kobayashi, Y.; Ikka, T.; Hirayama, T.; Shinozaki, K.; Kobayashi, M. Zinc Finger Protein STOP1 Is Critical for Proton Tolerance in Arabidopsis and Coregulates a Key Gene in Aluminum Tolerance. Proc. Natl. Acad. Sci. USA 2007, 104, 9900–9905. [Google Scholar] [CrossRef]
- Iuchi, S.; Kobayashi, Y.; Koyama, H.; Kobayashi, M. STOP1, a Cys2/His2 Type Zinc-Finger Protein, Plays Critical Role in Acid Soil Tolerance in Arabidopsis. Plant Signal. Behav. 2008, 3, 128–130. [Google Scholar] [CrossRef]
- Ding, Z.J.; Yan, J.Y.; Xu, X.Y.; Li, G.X.; Zheng, S.J. WRKY46 Functions as a Transcriptional Repressor of ALMT1, Regulating Aluminum-Induced Malate Secretion in Arabidopsis. Plant J. 2013, 76, 825–835. [Google Scholar] [CrossRef]
- Tokizawa, M.; Kobayashi, Y.; Saito, T.; Kobayashi, M.; Iuchi, S.; Nomoto, M.; Tada, Y.; Yamamoto, Y.Y.; Koyama, H. SENSITIVE TO PROTON RHIZOTOXICITY1, CALMODULIN BINDING TRANSCRIPTION ACTIVATOR2, and Other Transcription Factors Are Involved in ALUMINUM-ACTIVATED MALATE TRANSPORTER1 Expression. Plant Physiol. 2015, 167, 991–1003. [Google Scholar] [CrossRef] [PubMed]
- Ligaba-Osena, A.; Fei, Z.; Liu, J.; Xu, Y.; Shaff, J.; Lee, S.; Luan, S.; Kudla, J.; Kochian, L.; Piñeros, M. Loss-of-function Mutation of the Calcium Sensor CBL 1 Increases Aluminum Sensitivity in Arabidopsis. New Phytol. 2017, 214, 830–841. [Google Scholar] [CrossRef]
- Zhu, X.; Wang, P.; Bai, Z.; Herde, M.; Ma, Y.; Li, N.; Liu, S.; Huang, C.; Cui, R.; Ma, H.; et al. Calmodulin-like Protein CML24 Interacts with CAMTA2 and WRKY46 to Regulate ALMT1 -dependent Al Resistance in Arabidopsis thaliana. New Phytol. 2022, 233, 2471–2487. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Sadhukhan, A.; Kobayashi, Y.; Ogo, N.; Tokizawa, M.; Agrahari, R.K.; Ito, H.; Iuchi, S.; Kobayashi, M.; Asai, A.; et al. Involvement of Phosphatidylinositol Metabolism in Aluminum-Induced Malate Secretion in Arabidopsis. J. Exp. Bot. 2019, 70, 3329–3342. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Wang, J.; Wang, X.; Zhang, X.; Cheng, Y.; Cai, Z.; Nian, H.; Ma, Q. GmWRKY21, a Soybean WRKY Transcription Factor Gene, Enhances the Tolerance to Aluminum Stress in Arabidopsis thaliana. Front. Plant Sci. 2022, 13, 833326. [Google Scholar] [CrossRef]
- Tokizawa, M.; Enomoto, T.; Ito, H.; Wu, L.; Kobayashi, Y.; Mora-Macías, J.; Armenta-Medina, D.; Iuchi, S.; Kobayashi, M.; Nomoto, M.; et al. High Affinity Promoter Binding of STOP1 Is Essential for Early Expression of Novel Aluminum-Induced Resistance Genes GDH1 and GDH2 in Arabidopsis. J. Exp. Bot. 2021, 72, 2769–2789. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Ohyama, Y.; Kobayashi, Y.; Ito, H.; Iuchi, S.; Fujita, M.; Zhao, C.-R.; Tanveer, T.; Ganesan, M.; Kobayashi, M.; et al. STOP2 Activates Transcription of Several Genes for Al- and Low PH-Tolerance That Are Regulated by STOP1 in Arabidopsis. Mol. Plant 2014, 7, 311–322. [Google Scholar] [CrossRef]
- Zhang, X.; Li, L.; Yang, C.; Cheng, Y.; Han, Z.; Cai, Z.; Nian, H.; Ma, Q. GsMAS1 Encoding a MADS-Box Transcription Factor Enhances the Tolerance to Aluminum Stress in Arabidopsis thaliana. IJMS 2020, 21, 2004. [Google Scholar] [CrossRef]
- Balzergue, C.; Dartevelle, T.; Godon, C.; Laugier, E.; Meisrimler, C.; Teulon, J.-M.; Creff, A.; Bissler, M.; Brouchoud, C.; Hagège, A.; et al. Low Phosphate Activates STOP1-ALMT1 to Rapidly Inhibit Root Cell Elongation. Nat. Commun. 2017, 8, 15300. [Google Scholar] [CrossRef] [PubMed]
- Mora-Macías, J.; Ojeda-Rivera, J.O.; Gutiérrez-Alanís, D.; Yong-Villalobos, L.; Oropeza-Aburto, A.; Raya-González, J.; Jiménez-Domínguez, G.; Chávez-Calvillo, G.; Rellán-Álvarez, R.; Herrera-Estrella, L. Malate-Dependent Fe Accumulation Is a Critical Checkpoint in the Root Developmental Response to Low Phosphate. Proc. Natl. Acad. Sci. USA 2017, 114, E3563–E3572. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, Z.; Zheng, Z.; Dong, J.; Song, L.; Sui, L.; Nussaume, L.; Desnos, T.; Liu, D. Genetic Dissection of Fe-Dependent Signaling in Root Developmental Responses to Phosphate Deficiency. Plant Physiol. 2019, 179, 300–316. [Google Scholar] [CrossRef] [PubMed]
- Godon, C.; Mercier, C.; Wang, X.; David, P.; Richaud, P.; Nussaume, L.; Liu, D.; Desnos, T. Under Phosphate Starvation Conditions, Fe and Al Trigger Accumulation of the Transcription Factor STOP1 in the Nucleus of Arabidopsis Root Cells. Plant J. 2019, 99, 937–949. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wu, H.; Wang, N.; Fan, H.; Chen, C.; Cui, Y.; Liu, H.; Ling, H. Mediator Subunit 16 Functions in the Regulation of Iron Uptake Gene Expression in Arabidopsis. New Phytol. 2014, 203, 770–783. [Google Scholar] [CrossRef]
- Raya-González, J.; Ojeda-Rivera, J.O.; Mora-Macias, J.; Oropeza-Aburto, A.; Ruiz-Herrera, L.F.; López-Bucio, J.; Herrera-Estrella, L. MEDIATOR16 Orchestrates Local and Systemic Responses to Phosphate Scarcity in Arabidopsis Roots. New Phytol. 2021, 229, 1278–1288. [Google Scholar] [CrossRef] [PubMed]
- Shen, N.; Hou, S.; Tu, G.; Lan, W.; Jing, Y. Transcription Factor WRKY33 Mediates the Phosphate Deficiency-Induced Remodeling of Root Architecture by Modulating Iron Homeostasis in Arabidopsis Roots. IJMS 2021, 22, 9275. [Google Scholar] [CrossRef]
- Lee, H.-S.; Kim, Y.; Pham, G.; Kim, J.W.; Song, J.-H.; Lee, Y.; Hwang, Y.-S.; Roux, S.J.; Kim, S.-H. Brassinazole Resistant 1 (BZR1)-Dependent Brassinosteroid Signalling Pathway Leads to Ectopic Activation of Quiescent Cell Division and Suppresses Columella Stem Cell Differentiation. EXBOTJ 2015, 66, 4835–4849. [Google Scholar] [CrossRef]
- Liu, T.; Deng, S.; Zhang, C.; Yang, X.; Shi, L.; Xu, F.; Wang, S.; Wang, C. Brassinosteroid Signaling Regulates Phosphate Starvation-induced Malate Secretion in Plants. JIPB 2023, 65, 1099–1112. [Google Scholar] [CrossRef]
- Xu, J.M.; Wang, Z.Q.; Wang, J.Y.; Li, P.F.; Jin, J.F.; Chen, W.W.; Fan, W.; Kochian, L.V.; Zheng, S.J.; Yang, J.L. Low Phosphate Represses Histone Deacetylase Complex1 to Regulate Root System Architecture Remodeling in Arabidopsis. New Phytol. 2020, 225, 1732–1745. [Google Scholar] [CrossRef]
- Huang, X.-Y.; Wang, C.-K.; Zhao, Y.-W.; Sun, C.-H.; Hu, D.-G. Mechanisms and Regulation of Organic Acid Accumulation in Plant Vacuoles. Hortic. Res. 2021, 8, 227. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.-G.; Li, Y.-Y.; Zhang, Q.-Y.; Li, M.; Sun, C.-H.; Yu, J.-Q.; Hao, Y.-J. The R2R3-MYB Transcription Factor MdMYB73 Is Involved in Malate Accumulation and Vacuolar Acidification in Apple. Plant J. 2017, 91, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.-Y.; Gu, K.-D.; Cheng, L.; Wang, J.-H.; Yu, J.-Q.; Wang, X.-F.; You, C.-X.; Hu, D.-G.; Hao, Y.-J. BTB-TAZ Domain Protein MdBT2 Modulates Malate Accumulation and Vacuolar Acidification in Response to Nitrate. Plant Physiol. 2020, 183, 750–764. [Google Scholar] [CrossRef]
- Zhang, Q.-Y.; Gu, K.-D.; Wang, J.-H.; Yu, J.-Q.; Wang, X.-F.; Zhang, S.; You, C.-X.; Hu, D.-G.; Hao, Y.-J. BTB-BACK-TAZ Domain Protein MdBT2-Mediated MdMYB73 Ubiquitination Negatively Regulates Malate Accumulation and Vacuolar Acidification in Apple. Hortic. Res. 2020, 7, 151. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, J.; Guo, J.; Zhou, F.; Singh, S.; Xu, X.; Xie, Q.; Yang, Z.; Huang, C.-F. F-Box Protein RAE1 Regulates the Stability of the Aluminum-Resistance Transcription Factor STOP1 in Arabidopsis. Proc. Natl. Acad. Sci. USA 2019, 116, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Fang, Q.; Zhou, F.; Zhang, Y.; Singh, S.; Huang, C. Degradation of STOP1 Mediated by the F-box Proteins RAH1 and RAE1 Balances Aluminum Resistance and Plant Growth in Arabidopsis thaliana. Plant J. 2021, 106, 493–506. [Google Scholar] [CrossRef]
- Guo, J.; Zhang, Y.; Gao, H.; Li, S.; Wang, Z.; Huang, C. Mutation of HPR1 Encoding a Component of the THO/TREX Complex Reduces STOP1 Accumulation and Aluminium Resistance in Arabidopsis thaliana. New Phytol. 2020, 228, 179–193. [Google Scholar] [CrossRef]
- Luna, R.; Rondón, A.G.; Aguilera, A. New Clues to Understand the Role of THO and Other Functionally Related Factors in MRNP Biogenesis. Biochim. Et Biophys. Acta (BBA)—Gene Regul. Mech. 2012, 1819, 514–520. [Google Scholar] [CrossRef]
- Zhu, Y.-F.; Guo, J.; Zhang, Y.; Huang, C.-F. The THO/TREX Complex Component RAE2/TEX1 Is Involved in the Regulation of Aluminum Resistance and Low Phosphate Response in Arabidopsis. Front. Plant Sci. 2021, 12, 698443. [Google Scholar] [CrossRef]
- Augustine, R.C.; Vierstra, R.D. SUMOylation: Re-Wiring the Plant Nucleus during Stress and Development. Curr. Opin. Plant Biol. 2018, 45, 143–154. [Google Scholar] [CrossRef]
- Fang, Q.; Zhang, J.; Zhang, Y.; Fan, N.; Van Den Burg, H.A.; Huang, C.-F. Regulation of Aluminum Resistance in Arabidopsis Involves the SUMOylation of the Zinc Finger Transcription Factor STOP1. Plant Cell 2020, 32, 3921–3938. [Google Scholar] [CrossRef] [PubMed]
- Fang, Q.; Zhang, J.; Yang, D.-L.; Huang, C.-F. The SUMO E3 Ligase SIZ1 Partially Regulates STOP1 SUMOylation and Stability in Arabidopsis thaliana. Plant Signal. Behav. 2021, 16, 1899487. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.-B.; He, C.; Ma, Y.; Herde, M.; Ding, Z. Jasmonic Acid Enhances Al-Induced Root Growth Inhibition. Plant Physiol. 2017, 173, 1420–1433. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, L.; Su, H.; Guo, L.; Zhang, J.; Li, Y.; Xu, J.; Zhang, X.; Guo, Y.-D.; Zhang, N. Jasmonate and Aluminum Crosstalk in Tomato: Identification and Expression Analysis of WRKYs and ALMTs during JA/Al-Regulated Root Growth. Plant Physiol. Biochem. 2020, 154, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Hu, Z.; Luo, Y.; Feng, C.; Long, Y. Multiple ALMT Subunits Combine to Form Functional Anion Channels: A Case Study for Rice ALMT7. Front. Plant Sci. 2022, 13, 1012578. [Google Scholar] [CrossRef] [PubMed]
- Dortay, H.; Gruhn, N.; Pfeifer, A.; Schwerdtner, M.; Schmülling, T.; Heyl, A. Toward an Interaction Map of the Two-Component Signaling Pathway of Arabidopsis thaliana. J. Proteome Res. 2008, 7, 3649–3660. [Google Scholar] [CrossRef]
- Jones, A.M.; Xuan, Y.; Xu, M.; Wang, R.-S.; Ho, C.-H.; Lalonde, S.; You, C.H.; Sardi, M.I.; Parsa, S.A.; Smith-Valle, E.; et al. Border Control—A Membrane-Linked Interactome of Arabidopsis. Science 2014, 344, 711–716. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: A Multiple Sequence Alignment Method with Reduced Time and Space Complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef]
- Okonechnikov, K.; Golosova, O.; Fursov, M. Unipro UGENE: A Unified Bioinformatics Toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef]
- Whelan, S.; Goldman, N. A General Empirical Model of Protein Evolution Derived from Multiple Protein Families Using a Maximum-Likelihood Approach. Mol. Biol. Evol. 2001, 18, 691–699. [Google Scholar] [CrossRef]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The Rapid Generation of Mutation Data Matrices from Protein Sequences. Comput. Appl. Biosci. 1992, 8, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic representation of the domain arrangement in one AtALMT1 subunit. The TMD (transmembrane domain) contains six TMs (transmembrane helices I to VI); the CTD (C-terminal cytosolic domain) contains six Hs (α-helix bundles 1 to 6); and the magenta rectangle represents the fusaric acid resistance protein-like (pfam13515) domain.
Figure 1.
Schematic representation of the domain arrangement in one AtALMT1 subunit. The TMD (transmembrane domain) contains six TMs (transmembrane helices I to VI); the CTD (C-terminal cytosolic domain) contains six Hs (α-helix bundles 1 to 6); and the magenta rectangle represents the fusaric acid resistance protein-like (pfam13515) domain.

Figure 2.
The role of ALMTs in stomatal opening and closure. ALMT proteins are responsible for the influx/efflux of compounds (blue arrows); black arrows represent activation and blunt arrows represent inhibition, while the dotted arrow represents indirect regulation. ROS—reactive oxygen species, Mal2-—malate, ABA—abscisic acid, PP2C—protein phosphatases type 2C, MPKs—mitogen-activated protein kinases, GABA—γ-aminobutyric acid, CPKs—calcium-dependent kinases.
Figure 2.
The role of ALMTs in stomatal opening and closure. ALMT proteins are responsible for the influx/efflux of compounds (blue arrows); black arrows represent activation and blunt arrows represent inhibition, while the dotted arrow represents indirect regulation. ROS—reactive oxygen species, Mal2-—malate, ABA—abscisic acid, PP2C—protein phosphatases type 2C, MPKs—mitogen-activated protein kinases, GABA—γ-aminobutyric acid, CPKs—calcium-dependent kinases.

Figure 3.
Schematic representation of STOP-mediated transcriptional regulation of target genes under Al stress and normal conditions. Genes responsible for early-phase Al response are depicted in a pink rectangle, and late-phase Al response genes are shown within a pale green rectangle. Regulators of STOP1 are depicted in a dark green rectangle. Black arrows represent activation, and blunt arrows represent inhibition.
Figure 3.
Schematic representation of STOP-mediated transcriptional regulation of target genes under Al stress and normal conditions. Genes responsible for early-phase Al response are depicted in a pink rectangle, and late-phase Al response genes are shown within a pale green rectangle. Regulators of STOP1 are depicted in a dark green rectangle. Black arrows represent activation, and blunt arrows represent inhibition.

Figure 4.
Schematic model for the role of ALMT1 expression under low-Pi conditions. Under low-Pi and acidic conditions, Fe and Al accumulate in the cytoplasm, where they decrease the proteasomal degradation of STOP1, thereby promoting ALMT1 transcription. The tonoplast-located ALS2–STAR1 proteins transport Fe/Al to the vacuole, thus reducing their concentration in the cytosol and, subsequently, reducing STOP1 accumulation in the nucleus and ALMT1 transcription. ALMT1 exuded malate where it coupled with Fe and LOW PHOSPHATE ROOT 1 (LPR1); they generate ROS which inhibit cell wall expansion. Also, low Pi induced LPR1/2 expression, which converts Fe2+ to Fe3+. MEDIATOR 16 (MED16) interacted with STOP1 and linked with RNA polymerase II (RNA Pol II) to promote the expression of ALMT1 and malate efflux. The F-box protein Regulation of AtALMT1 Expression 1 (RAE1) interacted with STOP1 and promoted STOP1 ubiquitination and its further proteasomal degradation. At the same time, STOP1 promoted RAE1 transcription, thus creating a negative feedback loop between RAE1 and STOP1 (depicted in magenta arrows). Black arrows represent activation, blunt black arrows represent repression, dashed arrows represent transfer between compartments, and blue arrows indicate that the reaction involved other proteins.
Figure 4.
Schematic model for the role of ALMT1 expression under low-Pi conditions. Under low-Pi and acidic conditions, Fe and Al accumulate in the cytoplasm, where they decrease the proteasomal degradation of STOP1, thereby promoting ALMT1 transcription. The tonoplast-located ALS2–STAR1 proteins transport Fe/Al to the vacuole, thus reducing their concentration in the cytosol and, subsequently, reducing STOP1 accumulation in the nucleus and ALMT1 transcription. ALMT1 exuded malate where it coupled with Fe and LOW PHOSPHATE ROOT 1 (LPR1); they generate ROS which inhibit cell wall expansion. Also, low Pi induced LPR1/2 expression, which converts Fe2+ to Fe3+. MEDIATOR 16 (MED16) interacted with STOP1 and linked with RNA polymerase II (RNA Pol II) to promote the expression of ALMT1 and malate efflux. The F-box protein Regulation of AtALMT1 Expression 1 (RAE1) interacted with STOP1 and promoted STOP1 ubiquitination and its further proteasomal degradation. At the same time, STOP1 promoted RAE1 transcription, thus creating a negative feedback loop between RAE1 and STOP1 (depicted in magenta arrows). Black arrows represent activation, blunt black arrows represent repression, dashed arrows represent transfer between compartments, and blue arrows indicate that the reaction involved other proteins.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dabravolski, S.A.; Isayenkov, S.V. Recent Updates on ALMT Transporters’ Physiology, Regulation, and Molecular Evolution in Plants. Plants 2023, 12, 3167. https://doi.org/10.3390/plants12173167
AMA Style
Dabravolski SA, Isayenkov SV. Recent Updates on ALMT Transporters’ Physiology, Regulation, and Molecular Evolution in Plants. Plants. 2023; 12(17):3167. https://doi.org/10.3390/plants12173167
Chicago/Turabian StyleDabravolski, Siarhei A., and Stanislav V. Isayenkov. 2023. "Recent Updates on ALMT Transporters’ Physiology, Regulation, and Molecular Evolution in Plants" Plants 12, no. 17: 3167. https://doi.org/10.3390/plants12173167
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.