
Identification of the Plant Family Caryophyllaceae in Korea Using DNA Barcoding
by
 , ,
, ,
Dong-Pil Jin
1 ,
,
Sunhee Sim
2,
Jong-Won Park
3,
Ji-Eun Choi
2,
Jiwon Yoon
2,
Chae Eun Lim
2 and
Min-Ha Kim
2,* 1
Urban Biodiversity Research Division, Sejong National Arboretum, Korean Arboretum and Gardens Institute, Sejong 30106, Republic of Korea
2
Plant Resources Division, National Institute of Biological Resources, Incheon 22689, Republic of Korea
3
Department of Biology Education, Chonnam National University, Gwangju 61186, Republic of Korea
*
Author to whom correspondence should be addressed.
Plants 2023, 12(10), 2060; https://doi.org/10.3390/plants12102060
Submission received: 6 March 2023
/
Revised: 13 May 2023
/
Accepted: 16 May 2023
/
Published: 22 May 2023
(This article belongs to the Special Issue Diversity and Phylogenetic Relationships of Vascular Plants)
Abstract
:Caryophyllaceae is a large angiosperm family, with many species being utilized as ornamental or medicinal plants in Korea, in addition to several endangered species that are managed by the government. In this study, we used DNA barcoding for the accurate identification of Korean Caryophyllaceae. A total of 78 taxa (n = 215) were sequenced based on three chloroplast regions (rbcL, matK, and psbA–trnH) and nuclear ribosomal internal transcribed spacers (ITS). In the neighbor-joining tree, a higher accuracy of identification was generally observed when using ITS (>73%) rather than chloroplast regions (<62%). The highest resolution was found for rbcL + ITS (77.6%), although resolution varied according to the genus. Among the genera that included two and more species, five genera (Eremogone, Minuartia, Pseudostellaria, Sagina, and Stellaria) were successfully identified. However, the species of five other genera (Cerastium, Gypsophila, Dianthus, Silene, and Spergularia) showed relatively low resolutions (0–61.1%). In the cases of Cerastium, Dianthus, and Silene, ambiguous taxonomic relationships among unidentified species may have been a factor contributing to such low resolutions. However, in contrast to these results, Gypsophila and Spergularia have been identified well in previous studies. Our findings indicate the need of taxonomic reconsideration in Korea.
1. Introduction
Caryophyllaceae is a large family of angiosperms that includes approximately 2200 species across 86 genera, with the majority of plants in the family being found across the Northern Hemisphere [1]. Many members of Caryophyllaceae are commonly planted in gardens (e.g., Dianthus L. (carnation) and Gypsophila L. (baby’s-breath)) or are used as medicinal resources (e.g., Cerastium L. (mouse-ear chickweed), Silene L. (catchfly), and Stellaria L. (chickweed), summarized in [2]). In Korea, 66–91 taxa of 17 genera are recognized, including alien plants [3,4], and these taxa have various uses, similarly to other Caryophyllaceae species. For example, compounds of Gypsophila oldhamiana Miq. (having cytotoxicity against tumor cell), Silene firma Siebold & Zucc. (used for anuria, breast cancer, gonorrhea, and diseases of lactation), Stellaria aquatica (L.) Scope. (applied to pneumonia and high blood pressure), and Pseudostellaria heterophylla (Miq.) Pax (applied to tuberculosis, physical fatigue) have been investigated for their use as medicinal resources [5,6,7,8]. Additionally, to improve the diversity of ornamental crops and farm income, native Korean plants, such as Minuartia laricina (L.) Mattf. and Arenaria juncea M. Bieb., were tested and distributed under the national government’s plan [9]. Furthermore, Lychnis kiusiana Makino, Lychnis wilfordii (Regel) Maxim., and Silene capitata Kom. have been designated as endangered species II by the government [10]. Moreover, several other species are regarded as weeds of agricultural land [11,12], and alien plants of the family have been previously reported [13,14,15,16,17]. Such weeds and alien plants could lead to a decrease in crop yields and the disturbance of native species [18,19]. As a result, endangered, weed, and alien plants must all be continuously monitored; therefore, the precise identification of Caryophyllaceae species is critical for the use of resources and the management of weeds and alien species. However, factors such as morphological variations and polyploidy [20,21], and their commercial products being often sold as plant parts, tea bags, or powders, hinder accurate species identification and may even result in the misuse of resource species or a decrease in product quality.
For accurate and rapid identification of species, numerous previous studies have utilized molecular tools. In particular, DNA barcoding has been the primary method used for the last two decades. DNA barcoding identifies species using short nucleotide sequences or “barcodes” that can be applied across a variety of species [22,23]. In animal models, the cytochrome c oxidase I (CO1) gene is a commonly used DNA barcode [24,25]. However, its use as a DNA barcode in plants is difficult due to its slow and uneven mutation rate. Instead, two genes (rbcL and matK) of chloroplasts were suggested as core barcodes for plant identification. Generally, rbcL shows a high success rate of polymerase chain reaction (PCR) with relatively low resolution, whereas matK exhibits a better resolution despite the relatively low success rate of PCR [26]. However, these genes often show weak discriminatory power for lower taxa [27]; therefore, internal transcribed spacers (ITS) of nuclear ribosomal DNA and non-coding regions (e.g., psbA–trnH, atpF–atpH, and psbK–psbI) of chloroplasts that have more variable sites are suggested as additional regions [28,29]. In practice, DNA barcoding has been applied to the management of plant product quality [30,31], species conservation [32], the control of alien plants [33,34], and exploration of regional flora [35].
With regards to taxonomy, three subfamilies have been recognized in Caryophyllaceae: Alsinoideae, Caryophylloideae, and Paronychioideae [1,36,37]. However, morphological homoplasy within Caryophyllaceae make circumscribing the subfamilies difficult [1,38,39]. Molecular phylogenetic studies of the family also disregarded monophyly of traditional subfamilies; alternatively, the tribal classification was applied [38,39,40]. Similarly, the polyphyly of many genera was detected based on molecular phylogeny, such as Arenaria L., Gypsophila, Minuartia L., and Silene [40]. Therefore, taxonomic revisions on the Caryophyllaceae taxa have been conducted in accordance with molecular results, e.g., [41,42,43,44].
In this study, we aimed to identify Caryophyllaceae species in Korea using DNA barcoding. Considering previous DNA barcode studies, four regions were investigated: two crucial DNA barcodes (rbcL and matK) and two non-coding regions (psbA–trnH and ITS). In addition, information on nucleotide variation, PCR success rate, and discriminatory power was provided using a combination of barcodes. Those results would be utilized as reference of the molecular identification of Korean Caryophyllaceae. In addition, it has been concluded that sequences of various species can provide more detailed taxonomic relationships among taxa. Therefore, we believe that our data will contribute to the taxonomy and phylogeny of Caryophyllaceae.
2. Results
2.1. Characteristics of DNA Barcode Regions
The nucleotide sequences (rbcL, matK, psbA–trnH, and ITS) of 78 taxa including two forma (215 individuals) in Caryophyllaceae from Korea were obtained, and the information for each region is presented in Table S1. In terms of PCR success rate, all analyzed individuals were well amplified (rbcL = 100%, matK = 100%, psbA–trnH = 100%, and ITS = 100%). The sequencing success rate for all regions (rbcL, matK, psbA–trnH, and ITS) reached 100%; however, direct sequencing of the psbA–trnH region was not always successful. These results were clear after reading the region, poly A/T, or short inverted repeat (Figure 1). In the case of ITS, multiple peaks were observed across genera, and direct sequencing of Dianthus, Eremogone, and Stellaria was occasionally limited. When both forward and reverse sequencing results showed multiple peaks at a certain point, we treated that position as a mixed base. Nucleotide sequences were aligned without indels in rbcL (689 bp), whilst length variations were observed in the other regions: matK = 746–782 bp (aligned length = 784 bp), psbA–trnH = 160–404 bp (aligned length = 600 bp), and ITS = 606–637 bp (aligned length = 672 bp) (Table 1). The percentage of variable sites was highest in psbA–trnH (66.3%), followed by ITS (58.3%), matK (55.0%), and rbcL (18.0%) (Table 1). This order was the same for the percentage of Parsimony informative sites: psbA–trnH (61.8%), ITS (56.5%), matK (52.4%), and rbcL (16.7%) (Table 1).
2.2. Evaluation of DNA Barcodes
The mean interspecific pairwise distance was highest in psbA–trnH (0.3245), followed by ITS (0.2259), matK (0.1140), and rbcL (0.0291) (Table 2). Furthermore, the mean intraspecific genetic distance was also the highest in psbA–trnH (0.0078), followed by ITS (0.0037), matK (0.0019), and rbcL (0.0004) (Table 2). These values were 42–73 times larger in interspecific pairwise distances than in intraspecific pairwise distances. When combining the regions, the mean of interspecific pairwise distance is highest in psbA–trnH + ITS (0.2502), and the mean of intraspecific genetic distance is also highest (0.0048) (Table 2). However, an overlap was noted in genetic distance between intraspecific and interspecific pairwise distances for all regions (Figure 2); as a result, there was no distinct barcode gap.
The ability to identify species using the DNA barcode regions was also evaluated using the phylogenetic analysis—specifically, the Neighbor-Joining (NJ) method (Figure 3, Figure 4 and Figure 5). Generally, a higher degree of species resolution was observed in cases using ITS (>73%) compared with those using only chloroplast regions (<62%) (Table 3); therefore, the best resolution was shown in ITS + rbcL (77.63%) (Figure 5). The best close match also showed similar results with phylogenetic analysis (Table 3). Overall, a higher success rate was observed in combined DNA barcode regions than a single region. Especially, cases using ITS (>69%) showed a higher degree of species identification (“correct” in Table 3) than those using only chloroplast regions (<68%). Differing with the phylogenetic analysis, using the combined four regions and ITS + matK + psbA–trnH showed the best resolution (76.74%), while lower resolution was observed in ITS + rbcL (70.69%). As with the NJ and best close match analyses, better species partitions were observed in cases using combined regions compared to a single region. Under the best asap-score, the highest species partition was observed when using all regions (number of subsets, 40; asap-score, 8.00). In addition, the ASAP exhibited better resolution in cases using ITS compared to those using only chloroplast regions (Figures S1–S14).
Based on the best resolution tree (ITS + rbcL) (Figure 5), most genera were identified on all NJ trees, except for Minuartia and Silene. The ability for species identification varied according to the genus. Among the genera that included two or more taxa, all five genera (Eremogone Fenzl, Minuartia, Pseudostellaria Pax, Sagina L., and Stellaria) formed a clade, indicating successful identification (Figure 5). Each individual for Eremogone capillaris (Poir.) Fenzl and Minuartia arctica (Steven ex Ser.) Graebn. was investigated on the NJ tree; these were thought to have been distinguishable due to sufficient branch divergence from related species. In contrast, species of five genera (Cerastium, Gypsophila, Dianthus, Silene, and Spergularia (Pers.) J. Presl & C. Presl) were moderately or hardly distinguished at all (0–61.11%). Two species of Gypsophila (Gypsophila oldhamiana and Gypsophila pacifica Kom.) showed no separation from each other (0%), whilst Cerastium and Spergularia also showed low resolution (33.33%). Cerastium glomeratum Thuill. was shown to be separated from the two subspecies of Cerastium fontanum Baumg. (subsp. hallaisanense (Nakai) J.S. Kim and subsp. vulgare (Hartm.) Greuter & Burdet); however, these subspecies were not distinguishable from each other. Similarly, in Spergularia, only Spergularia rubra (L.) J. Presl & C. Presl could be identified from Spergularia bocconei (Scheele) Asch. & Graebn. and Spergularia marina (L.) Griseb. The DNA barcodes showed better discriminatory power for Dianthus (60.00%) and Silene (61.11%). In Dianthus, Dianthus armeria L., Dianthus barbatus L. var. asiaticus Nakai, and Dianthus japonicus Thunb. were all clustered in each clade. However, Dianthus chinensis var. serpens Y. N. Lee, Dianthus chinensis var. morii (Nakai) Y. C. Chu, Dianthus superbus L. var. superbus, and Dianthus superbus var. speciosus Rchb. diverged into a branch separate from other taxa and were sequenced based on only one individual. In addition, the majority of Dianthus species showed multiple types on the ITS; therefore, we concluded that these four taxa may not be distinguishable. In Silene, Silene capitata Kom., Silene firma, Silene seoulensis Nakai, Silene baccifera (L.) Roth, Silene gallica L., Silene repens Patrin ex Pers., Silene antirrhina L., Silene koreana Kom., and Silene takesimensis Uyeki & Sakata were all clustered into each clade according to morphological identification. Only one individual was sequenced in the case of Silene conoidea L., although this was clearly separated from the other. Despite Silene aprica Turcz. Ex Fisch. & C. A. Mey. being distinguishable at the species level, it was more difficult to distinguish at the intraspecies level.
3. Discussion
In this study, we examined four DNA barcoding regions in Korean Caryophyllaceae species. Ideal DNA barcodes allow for easy amplification and have sufficient variable sites to identify species [22]. Across taxa, all regions were well amplified with universal primers and sequences were obtained successfully. These results corresponded with the criteria for easy amplification. However, when considering their discriminatory ability, the combination of ITS was deemed as the optimal DNA barcode, with 71.43 to 75.32% in the phylogenetic analyses (Table 3). The best close match supported such a result of phylogenetic analysis, although there are some differences between the two analyses (Table 3). This identification resolution was comparable to that of other angiosperm taxa [45,46,47]. ITS is well known to have many informative sites; therefore, this region has previously been suggested as a DNA barcode in other plants because of its high resolution [47,48,49,50,51]. These results may have been influenced by the mutation rate of ITS being higher than that of chloroplast genes (rbcL and matK) [52,53]. Similar to the results of previous studies, our results showed a higher discriminatory power when using ITS (>72%) than when using only chloroplast DNA regions (<61%) (Table 3). A low amplification rate of ITS has occasionally been reported according to the taxon [54], although such a problem was not observed in this study.
Although the addition of the psbA–trnH region (74.03%) did not improve the ability of species identification when compared with using only ITS (74.03%), this region did enhance discriminatory power at the genus level (Table 3). In the ITS and ITS + rbcL trees, Silene oliganthella Nakai ex Kitag. was not well distinguished from the congeneric species Silene fasciculata Nakai and Silene jenisseensis Willd. because of its short branch (Figure 4 and Figure 5). Meanwhile, Silene oliganthella diverged deeply from the latter with a long branch on the ITS + psbA–trnH tree (Figure S15). However, such expectations require caution, considering that only one sample of Silene oliganthella was included in the present study. In addition, with regard to taxonomy, the species was regarded as a variety or synonym of Silene jenisseensis [55]. Despite considerable sequence variation in psbA–trnH attributed to divergence among Silene species, simultaneous high variation within species disturbed precise identification in the case of Pseudostellaria. Sequence variations within species were detected, although some were shared with individuals of other species. Such a low resolution of psbA–trnH has also been observed in other lineages [56,57]. In addition, bi-directional sequencing of psbA–trnH often failed, which was thought to be due to long mononucleotide repeats [58,59] or a potential loop structure [60]. In fact, slippage after long mononucleotide repeats was observed in Dianthus and Saponaria, whilst a pair of inverted repeats that could form a cruciform shape was found in Cerastium, Sagina, and Spergularia (Figure 1). Therefore, in this study, we suggest ITS as a DNA barcode for Korean Caryophyllaceae species when considering both cost efficiency and discriminatory power, such as previous DNA barcoding studies [47,48,49,50,51]. Furthermore, using rbcL is recommended for better resolution (Table 3). In addition, psbA–trnH could be used as a supplementary region for the identification of a genus such as Silene. It is probable when considering the high degree of species identification in the best close match (Table 3). When considering cases where two DNA barcode regions were combined, it was supported by the ASAP result that the highest degree of species partitioning (37) was observed in ITS + psbA–trnH (under the best ASAP score).
Most of the genera in Korea were accurately identified in our phylogenetic tree, with the exception of Minuartia and Silene, which were identified as polyphyletic groups (Figure 5). However, this was not due to a lack of discriminatory power in the DNA regions used in this study. Molecular phylogenies have previously suggested that these two genera are polyphyletic [44,61]. In the case of Minuartia, the three species (Minuartia arctica, Minuartia laricina, and Minuartia verna (L.) Hiern var. leptophylla (Rchb.) Nakai) analyzed in this study each belong to different sections within the genus (summarized in [61]). The molecular phylogeny also showed that each of these species are included in different clades [61]. In fact, Minuartia laricina and Minuartia verna var. leptophylla were not analyzed in the analysis but their position in the phylogenetic tree could be inferred based on closely related taxa. As a result, they have been treated as Cherleria arctica (Steven ex Ser.) A. J. Moore & Dillenb., and Pseudocherleria laricina (L.) Dillenb. & Kadereit [61,62]. Minuartia verna var. leptophylla was not mentioned in the previous study [61]; it may be treated as a taxon of Sabulina Rchb. Silene has also been suggested as a polyphyletic group. In that study, Silene armeria L. (=Atocion armeria (L.) Raf.) were placed within the sister group of Silene. Lychnis species were posited within Silene; so, they were treated as a section of Silene. In light of the polyphyly of Minuartia and Silene, further investigation may be necessary to evaluate the validity of their current taxonomic classification in Korea.
Although most species of Caryophyllaceae were identified well using DNA barcodes, five genera (Cerastium, Gypsophila, Dianthus, Silene, and Spergularia) showed a low resolution (0–61.11%). Such a low resolution may have been caused by wide morphological variations within species and hybridization among species. In the case of Cerastium, subspecies of Cerastium fontanum (subsp. hallaisanense and subsp. vulgare) were not distinguished and were instead considered taxa [63]. Similar to Cerastium, unidentified Silene species in the NJ tree were thought to be undistinctive taxa (Silene takesimensis, Silene fasciculata, Silene oliganthella) [64]. For Dianthus, many species could have intercrossed both naturally and through cultivation (summarized in [65]). In reality, multiple peaks in the ITS region are frequently detected in the genus, which can make identification difficult. Therefore, the low rate of identification of these genera is not surprising considering the taxonomic similarities between them. In contrast, Spergularia bocconei could be distinguished from Spergularia marina by its dense glandular hair on stems and leaves [16], in addition to having smaller capsules/seeds than the latter [66]. Gypsophila pacifica and Gypsophila oldhamiana could also be distinguished by their leaf (ovate vs. oblong), inflorescence (diffuse vs. dense), and flower morphologies (pedicels, 2–5 mm vs. 5–10 mm; apex of petals, rounded vs. truncate or retuse; stamens and styles, shorter than petals vs. longer than petals) [3]. The Spergularia and Gypsophila species have been well identified using ITS2 and trnL–F [67], and ISSR and RAPD [68]. However, previous studies are in contrast to our results. There might be various reasons for such results, such as genetic variation across different regions or misconceptions about the taxonomic classification of these two genera. However, it is difficult to determine the exact cause based on the current study alone. Therefore, further investigation of morphological and genetic variations is necessary to better understand the relationships between these two genera in Korea.
4. Materials and Methods
4.1. Taxon Sampling
A total of 215 individuals representing 78 taxa across 17 genera were collected through field surveys, and dried specimens were collected from the herbarium of the National Institute of Biological Resources (NIBR). Leaves sampled from the field were immediately dried using silica gel. We attempted to cover the majority of species in Caryophyllaceae by citing the National List of Species of Korea [4]. For plants that grow in North Korea, we utilized samples collected from nearby regions, such as China and Russia. To represent genetic variation within species, we collected three or more samples per taxon from different populations, excluding plants that grow in North Korea. The information of samples is shown in Table S1.
4.2. DNA Extraction, Polymerase Chain Reaction (PCR), and Sequencing
Total genomic DNA of the abovementioned plants was extracted from silica-dried leaves using NucleoSpin® Plant II (Macherey-Nagel, Düren, Germany), according to the manufacturer’s protocol, although the incubation times during cell lysis and elution of DNA were manually modified. The concentration of extracted DNA was subsequently measured using a Synergy LX microplate reader (BioTek Instruments, Winooski, VT, USA).
PCR was carried out to amplify DNA barcode regions, and each reaction mixture contained approximately 10 ng of DNA, 10 μL of AccuPower® Taq PCR PreMix (Bioneer, Daejeon, Republic of Korea), distilled water, and appropriate volume of primers (usually 0.3 μM) in a total volume of 20 μL. The primers were selected with reference to previous studies so that they could be applied to the taxa of Caryophyllaceae (Table 4). The reaction was conducted after initial denaturation at 95 °C for 3 min: denaturation step at 95 °C for 30 s, annealing step at 52 °C (for rbcL, matK, psbA–trnH) to 55 °C (for ITS) for 30–45 s, and extension step at 72 °C for 45–80 s according to product size in 35 cycles. The final extension was performed at 72 °C for 7 min. PCR results were then confirmed via high-resolution capillary electrophoresis on a QIAxcel Advanced Instrument (Qiagen, Hilden, Germany). Successfully amplified products were sequenced using an ABI 3730XL sequencer (Applied Biosystems, Foster City, CA, USA), and sequencing was performed by Macrogen (Seoul, Republic of Korea).
4.3. Data Analysis
Chromatogram files of sequencing were imported into Geneious R 10.2.6 (Biomatters, Auckland, New Zealand), and the nucleotide sequences were curated, analyzed, and aligned using MAFFT [74], with some parts being manually edited. All nucleotide sequences produced in this study were deposited in GenBank (Table S1, ITS: OQ150537–OQ150751, rbcL: OQ172533–OQ172747, matK: OQ172318–OQ172532, psbA–trnH: OQ172748–OQ172962O).
The characteristics such as length, number of variable sites, and G/C content of nucleotide sequences were measured using Geneious R 10.2.6 (Biomatters), whilst the number of parsimony-informative sites was measured using PAUP 4.0b10 [75]. To evaluate DNA barcoding regions, the barcode gap was checked, which indicated whether or not interspecific and intraspecific distances overlapped. In this process, according to various combinations of the four regions, the pairwise genetic distance matrix for each individual was calculated based on the Kimura 2-parameter (K2P) method [76], which was conducted using MEGA 11 [77]. To identify species, the NJ tree was also constructed, using the K2P method throughout MEGA 11, with the bootstrap value of each node being calculated with 2000 replications. Amaranthus spinosus L. was used as an outgroup in this NJ analysis. (ITS: KY968964, matK: MF159529, rbcL: MF135474, psbA–trnH: MF143791). The NJ analysis uses a distance matrix based on nucleotide differences between pairs of taxa [78], making it well-suited to DNA barcoding. Compared with other methods of phylogenetic tree construction, the NJ analysis is relatively insensitive to issues such as multiple substitutions and missing data, and it is also faster [78,79]. The success of species identification was determined by whether individuals within the species were clustered into a clade. In this process, the forma was not counted as an individual taxon. When only one individual within a species was analyzed, we decided manually by considering branch length.
Along with the NJ analysis, we used the “best close match” function in TAXONDNA to assess the success of species identification [80]. This analysis assigned a query to the species name of its best-matching barcode, regardless of the degree of similarity between the query and barcode sequences, within a distance threshold of 95% [80]. Additionally, we utilized Assemble Species by Automatic Partitioning (ASAP), an unsupervised Operational Taxonomic Unit (OTU) picking method based on pairwise sequence distance [81]. The ASAP suggests optimal species partitions with an asap-score that reflects the confidence level of the clustering. Generally, a lower score implies a better partition. This program is available online (https://bioinfo.mnhn.fr/abi/public/asap/asapweb.html (accessed on 5 May 2023)). In this process, we used default parameters, except for the substitution model (K2P method).
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants12102060/s1, Figure S1: Species partitions through ASAP algorithm based on rbcL; Figure S2: Species partitions through ASAP algorithm based on matK; Figure S3: Species partitions through ASAP algorithm based on psbA–trnH; Figure S4: Species partitions through ASAP algorithm based on internal transcribed spacer (ITS); Figure S5: Species partitions through ASAP algorithm based on ITS + matK; Figure S6: Species partitions through ASAP algorithm based on ITS + psbA–trnH; Figure S7: Species partitions through ASAP algorithm based on ITS + rbcL; Figure S8: Species partitions through ASAP algorithm based on matK + psbA–trnH; Figure S9: Species partitions through ASAP algorithm based on matK + rbcL; Figure S10: Species partitions through ASAP algorithm based on psbA–trnH + rbcL; Figure S11: Species partitions through ASAP algorithm based on ITS + matK + rbcL; Figure S12: Species partitions through ASAP algorithm based on ITS + psbA–trnH + rbcL; Figure S13: Species partitions through ASAP algorithm based on rbcL + psbA–trnH + rbcL; Figure S14: Species partitions through ASAP algorithm based on ITS + matK + psbA–trnH + rbcL; Figure S15: The neighbor-joining phylogenetic tree of the Korean Caryophyllaceae based on combined sequence from ITS + psbA–trnH; Table S1: List of species sequenced in this study and GenBank accession number.
Author Contributions
C.E.L. and M.-H.K. designed this study and acquired funding; D.-P.J., S.S., J.-E.C. and J.Y. performed DNA experiments; D.-P.J., S.S., J.-W.P. and J.-E.C. analyzed the data and visualized the results; D.-P.J. and J.-W.P. validated the results; D.-P.J. wrote and edited manuscript; J.-W.P., C.E.L. and M.-H.K. reviewed the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a grant from the National Institute of Biological Resources funded by the Ministry of Environment of the Republic of Korea (no: NIBR202212101, NIBR202313101).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The four chloroplast genomes, newly sequenced in this study, were archived in NCBI with accession numbers OQ150537–OQ150751 and OQ172318–OQ172962.
Acknowledgments
The authors are grateful to the concerned staff at the Korea National Arboretum, National Institute of Ecology (Korea), and Warm Temperate and Subtropical Forest Research Center (Korea) for permitting the examination and collection of specimens. The authors also would like to thank the members of the society for Korean Peninsula plants (han-ban-do-sig-mul-yeon-gu-hoe) for their assistance in collecting samples.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bittrich, V. Caryophyllaceae. In The Families and Genera of Vascular Plants; Kubitzki, K., Bittrich, V., Rohwer, J., Eds.; Springer: Berlin, Germany, 1993; Volume 2, pp. 206–230. [Google Scholar]
- Chandra, S.; Rawat, D.S. Medicinal plants of the family Caryophyllaceae: A review of ethno-medicinal uses and pharmacological properties. Integr. Med. Res. 2015, 4, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.P.; Choi, K. Caryophyllaceae. In Flora of Korea; Flora of Korea Editorial Committee, Ed.; National Institute of Biological Resources: Incheon, Republic of Korea, 2018; Volume 3, pp. 30–61. [Google Scholar]
- National Institute of Biological Resources. National List of Species of Korea. 2021. Available online: https://species.nibr.go.kr/index.do (accessed on 7 February 2023).
- Han, Y.S.; Yim, D.S.; Lee, S.Y. Studies on biological activities of the roots of Pseudostellaria heterophylla. Saengyak Hakhoe Chi 2000, 31, 45–50, (In Korean with English Abstract). [Google Scholar]
- Xie, L.X.; Zhang, H.C.; Wang, H.Y.; Wang, Y.; Wang, F.L.; Sun, J.Y. Two new triterpenoids from Gypsophila oldhamiana. Nat. Prod. Res. 2016, 30, 1068–1074. [Google Scholar] [CrossRef]
- Cho, H.D.; Kang, W.S.; Kim, D.H.; Ku, J.J.; Seo, K.I. Comparison of biological activity between Stellaria aquatica seed extracts. Korean J. Food Preserv. 2019, 26, 228–237. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Widyowati, R.; Sulistyowaty, M.I.; Sugimoto, S.; Yamano, Y.; Kawakami, S.; Otsuka, H.; Matsunami, K. Firmosides A and B: Two new sucrose ferulates from the aerial parts of Silene firma and evaluation of radical scavenging activities. J. Nat. Med. 2020, 74, 796–803. [Google Scholar]
- Gil, M.; Kwon, H.H.; Kwon, Y.H.; Jung, M.J.; Kim, S.Y.; Rhie, Y.H. Growth of Minuartia laricina, Arenaria juncea, and Corydalis speciose in field with various soil water contents. J. Bio-Envrion. Control 2020, 29, 344–353, (In Korean with English Abstract). [Google Scholar] [CrossRef]
- National Institute of Ecology. Endangered Wildlife. Available online: https://www.nie.re.kr/nieEng/main/contents.do?menuNo=400052 (accessed on 7 February 2023).
- Kim, S.C.; Oh, Y.J.; Kwon, Y.W. Weed flora of agricultural area in Korea. KJWS 1992, 12, 317–334. [Google Scholar]
- Lee, I.Y.; Oh, Y.J.; Hong, S.H.; Heo, S.J.; Lee, C.Y.; Park, K.W.; Cho, S.H.; Kwon, O.D.; Im, I.B.; Kim, S.K. Occurrence of weed flora and changes in weed vegetation in orchard fields of Korea. Weed Turf. Sci. 2017, 6, 21–27, (In Korean with English Abstract). [Google Scholar] [CrossRef]
- Lee, Y.M.; Park, S.H.; Choi, H.S.; Yang, J.C.; Nam, G.H.; Chung, G.Y.; Choi, H.J. Floristic study of Dalmasan and its adjacent regions. Korean J. Environ. Ecol. 2009, 23, 1–21, (In Korean with English Abstract). [Google Scholar]
- Bak, G.J.; Koh, J.K.; Pak, J.H. Newly recorded naturalized species in Korea, Silene Antirrhina L. (Caryophyllaceae). Korean J. Plant Taxon. 2011, 41, 171–174, (In Korean with English Abstract). [Google Scholar] [CrossRef]
- Jung, S.Y.; Hong, J.K.; Park, S.H.; Yang, J.C.; Yun, S.M.; Kang, Y.S. Two unrecorded alien plants of South Korea: Geranium Dissectum L. (Geraniaceae) and Dianthus Armeria L. Korean J. Plant Taxon. 2015, 45, 272–277, (In Korean with English Abstract). [Google Scholar] [CrossRef]
- Choi, J.E.; Kim, J.H.; Hong, J.K.; Kim, J.S. Two new naturalized species from South Korea, Polygonum ramosissimum Michx. (Polygonaceae) and Spergularia Bocconi (Scheele) Graebn. (Caryophyllaceae). Korean J. Plant Taxon. 2016, 46, 326–330, (In Korean with English Abstract). [Google Scholar] [CrossRef]
- Hong, J.K.; Shim, S.D.; Kim, H.S.; Sim, S.; Hyun, C.W.; Kim, J.H. New record of an alien plant, Petrorhagia nanteuilii (Caryophyllaceae) in Korea. Korean J. Plant Taxon. 2021, 51, 386–390. [Google Scholar] [CrossRef]
- Rembold, K.; Mangopo, H.; Tjitrosoedirdjo, S.S.; Kreft, H. Plant diversity, forest dependency, and alien plant invasions in tropical agricultural landscapes. Biol. Conserv. 2017, 213, 234–242. [Google Scholar] [CrossRef]
- Son, S.H.; Jo, A.R.; Kim, D.E. Current status of alert alien species management for the establishment of proactive management systems in Korea. J. Ecol. Environ. 2021, 45, 26. [Google Scholar] [CrossRef]
- Brysting, A.K.; Elven, R. The Cerastium alpinum-C. arcticum complex (Caryophyllaceae): Numerical analyses of morphological variation and a taxonomic revision of C. arcticum Lange sl. Taxon 2000, 49, 189–216. [Google Scholar] [CrossRef]
- Farsi, M.; Behroozian, M.; Vaezi, J.; Joharchi, M.R.; Memariani, F. The evolution of Dianthus polylepis complex (Caryophyllaceae) inferred from morphological and nuclear DNA sequence data: One or two species? Plant Syst. Evol. 2013, 299, 1419–1431. [Google Scholar] [CrossRef]
- Kress, W.J.; Wurdack, K.J.; Zimmer, E.A.; Weigt, L.A.; Janzen, D.H. Use of DNA barcodes to identify flowering plants. Proc. Natl. Acad. Sci. USA 2005, 102, 8369–8374. [Google Scholar] [CrossRef]
- CBOL Plant Working Group. A DNA barcode for land plants. Proc. Natl. Acad. Sci. USA 2009, 106, 12794–12797. [Google Scholar] [CrossRef]
- Kress, W.J.; Erickson, D.L. DNA barcodes: Genes, genomics, and bioinformatics. Proc. Natl. Acad. Sci. USA 2008, 105, 2761–2762. [Google Scholar] [CrossRef]
- Meusnier, I.; Singer, G.A.C.; Landry, J.F.; Hickey, D.A.; Hebert, P.D.N.; Hajibabaei, M. A universal DNA mini-barcode for biodiversity analysis. BMC Genom. 2008, 9, 214. [Google Scholar] [CrossRef]
- Naeem, A.; Khan, A.A.; Cheema, H.M.N.; Khan, I.A.; Buerkert, A. DNA barcoding for species identification in the Palmae family. Genet. Mol. Res. 2014, 13, 10341–10348. [Google Scholar] [CrossRef] [PubMed]
- Gernandt, D.S.; López, G.G.; García, S.O.; Liston, A. Phylogeny and classification of Pinus. Taxon 2005, 54, 29–42. [Google Scholar] [CrossRef]
- Hollingsworth, P.M.; Graham, S.W.; Little, D.P. Choosing and using a plant DNA barcode. PLoS ONE 2011, 6, e19254. [Google Scholar] [CrossRef]
- Dong, W.; Xu, C.; Li, C.; Sun, J.; Zuo, Y.; Shi, S.; Cheng, T.; Guo, J.; Zhou, S. ycf1, the most promising plastid DNA barcode of land plants. Sci. Rep. 2015, 5, 8348. [Google Scholar] [CrossRef] [PubMed]
- Mishra, P.; Kumar, A.; Nagireddy, A.; Mani, D.N.; Shukla, A.K.; Tiwari, R.; Sundaresan, V. DNA barcoding: An efficient tool to overcome authentication challenges in the herbal market. Plant Biotechnol. J. 2016, 14, 8–21. [Google Scholar] [CrossRef]
- Gesto-Borroto, R.; Medina-Jiménez, K.; Lorence, A.; Villarreal, M.L. Application of DNA barcoding for quality control of herbal drugs and their phytopharmaceuticals. Rev. Bras. Farmacogn. 2021, 31, 127–141. [Google Scholar] [CrossRef]
- Hartvig, I.; Czako, M.; Kjær, E.D.; Nielsen, L.R.; Theilade, I. The use of DNA barcoding in identification and conservation of rosewood (Dalbergia spp.). PLoS ONE 2015, 10, e0138231. [Google Scholar] [CrossRef]
- Van De Wiel, C.C.M.; Van Der Schoot, J.; Van Valkenburg, J.; Duistermaat, H.; Smulders, M.J.M. DNA barcoding discriminates the noxious invasive plant species, floating pennywort (Hydrocotyle ranunculoides L.f.), from non-invasive relatives. Mole. Ecol. Resour. 2009, 9, 1086–1091. [Google Scholar] [CrossRef]
- Jin, D.P.; Kim, J.H.; Sim, S.; Suh, H.J.; Kim, J.S. New record of an alien plant, Desmodium paniculatum (Fabaceae), in Korea based on a morphological examination and DNA barcoding. Korean J. Plant Taxon. 2021, 51, 133–140. [Google Scholar] [CrossRef]
- Mosa, K.A.; Gairola, S.; Jamdade, R.; El-Keblawy, A.; Al Shaer, K.I.; Al Harthi, E.K.; Shabana, H.A.; Mahmoud, T. The promise of molecular and genomic techniques for biodiversity research and DNA barcoding of the Arabian Peninsula flora. Front. Plant Sci. 2019, 9, 1929. [Google Scholar] [CrossRef]
- Pax, F.; Hoffmann, K. Caryophyllaceae. In Die Natürlichen Pflanzenfamilien, 2nd ed.; Engler, A., Prantl, K., Eds.; Engelmann: Leipzig, Germany, 1934; pp. 275–364. [Google Scholar]
- Rabeler, R.K.; Bittrich, V. Suprageneric nomenclature in the Caryophyllaceae. Taxon 1993, 42, 857–863. [Google Scholar] [CrossRef]
- Fior, S.; Karis, P.O.; Casazza, G.; Minuto, L.; Sala, F. Molecular phylogeny of the Caryophyllaceae (Caryophyllales) inferred from chloroplast matK and nuclear rDNA ITS sequences. Am. J. Bot. 2006, 93, 399–411. [Google Scholar] [CrossRef] [PubMed]
- Harbaugh, D.T.; Nepokroeff, M.; Rabeler, R.K.; McNeill, J.; Zimmer, E.A.; Wagner, W.L. A new lineage-based tribal classification of the family Caryophyllaceae. Int. J. Plant Sci. 2010, 171, 185–198. [Google Scholar] [CrossRef]
- Greenberg, A.K.; Donoghue, M.J. Molecular systematics and character evolution in Caryophyllaceae. Taxon 2011, 60, 1637–1652. [Google Scholar] [CrossRef]
- Frajman, B.; Thollesson, M.; Oxelman, B. Taxonomic revision of Atocion and Viscaria (Sileneae, Caryophyllaceae). Bot. J. Linn. Soc. 2013, 173, 194–210. [Google Scholar] [CrossRef]
- Madhani, H.; Rabeler, R.; Pirani, A.; Oxelman, B.; Heubl, G.; Zarre, S. Untangling phylogenetic patterns and taxonomic confusion in tribe Caryophylleae (Caryophyllaceae) with special focus on generic boundaries. Taxon 2018, 67, 83–112. [Google Scholar] [CrossRef]
- Sharples, M.T.; Tripp, E.A. Phylogenetic relationships within and delimitation of the cosmopolitan flowering plant genus Stellaria L. (Caryophyllaceae): Core stars and fallen stars. Syst. Bot. 2019, 44, 857–876. [Google Scholar] [CrossRef]
- Jafari, F.; Zarre, S.; Gholipour, A.; Eggens, F.; Rabeler, R.K.; Oxelman, B. A new taxonomic backbone for the infrageneric classification of the species-rich genus Silene (Caryophyllaceae). Taxon 2020, 69, 337–368. [Google Scholar] [CrossRef]
- Luo, K.; Chen, S.; Chen, K.; Song, J.; Yao, H.; Ma, X.; Zhu, Y.; Pang, X.; Yu, H.; Li, X.; et al. Assessment of candidate plant DNA barcodes using the Rutaceae family. Sci. China Life Sci. 2010, 53, 701–708. [Google Scholar] [CrossRef]
- Jin, D.P.; Park, J.W.; Park, J.S.; Choi, B.H. DNA barcode and phylogenetic study of the tribe Desmodieae (Fabaceae) in Korea. Korean J. Plant Taxon. 2019, 49, 224–239. [Google Scholar] [CrossRef]
- Zhang, D.; Jiang, B. Species identification in complex groups of medicinal plants based on DNA barcoding: A case study on Astragalus Spp. (Fabaceae) from Southwest China. Conserv. Genet. Resour. 2020, 12, 469–478. [Google Scholar] [CrossRef]
- Feng, S.; Jiang, Y.; Wang, S.; Jiang, M.; Chen, Z.; Ying, Q.; Wang, H. Molecular identification of Dendrobium species (Orchidaceae) based on the DNA barcode ITS2 region and its application for phylogenetic study. Int. J. Mol. Sci. 2015, 16, 21975–21988. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.J.; Ji, Y.; Choi, G.; Kang, Y.M.; Yang, S.; Moon, B.C. Molecular identification and phylogenetic analysis of important medicinal plant species in genus Paeonia based on rDNA-ITS, matK, and rbcL DNA barcode sequences. Genet. Mol. Res. 2016, 15, 1–12. [Google Scholar] [CrossRef]
- Yao, P.C.; Gao, H.Y.; Wei, Y.N.; Zhang, J.H.; Chen, X.Y.; Li, H.Q. Evaluating sampling strategy for DNA barcoding study of coastal and inland halo-tolerant Poaceae and Chenopodiaceae: A case study for increased sample size. PLoS ONE 2017, 12, e0185311. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Xie, W.; Li, X.; Shi, S.; Guo, Z. Establishing community-wide DNA barcode references for conserving mangrove forests in China. BMC Plant Biol. 2021, 21, 571. [Google Scholar] [CrossRef] [PubMed]
- Aubriot, X.; Lowry, P.P., II; Cruaud, C.; Couloux, A.; Haevermans, T. DNA barcoding in a biodiversity hot spot: Potential value for the identification of Malagasy Euphorbia L. listed in CITES Appendices I and II. Mole. Ecol. Resour. 2013, 13, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Ng, W.L.; Mahat, M.N.; Nazre, M.; Mohamed, R. DNA barcoding of the endangered Aquilaria (Thymelaeaceae) and its application in species authentication of agarwood products traded in the market. PLoS ONE 2016, 11, e0154631. [Google Scholar] [CrossRef]
- Chen, S.; Yao, H.; Han, J.; Liu, C.; Song, J.; Shi, L.; Zhu, Y.; Ma, X.; Gao, T.; Pang, X.; et al. Validation of the ITS2 region as a novel DNA barcode for identifying medicinal plant species. PLoS ONE 2010, 5, e8613. [Google Scholar] [CrossRef]
- Zhou, L.; Wu, Z.; Magnus, L.; Bengt, O. Silene Linnaeus. In Flora of China; Wu, C.Y., Raven, P.H., Hong, D.Y., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 2001; Volume 6, pp. 66–100. [Google Scholar]
- Edwards, D.; Horn, A.; Taylor, D.; Savolainen, V.; Hawkins, J.A. DNA barcoding of a large genus, Aspalathus L. (Fabaceae). Taxon 2008, 57, 1317–1327. [Google Scholar] [CrossRef]
- Castro, C.; Hernandez, A.; Alvarado, L.; Flores, D. DNA barcodes in fig cultivars (Ficus carica L.) using ITS regions of ribosomal DNA, the psbA–trnH spacer and the matK coding sequence. Am. J. Plant Sci. 2015, 6, 95–102. [Google Scholar] [CrossRef]
- Fazekas, A.J.; Kuzmina, M.L.; Newmaster, S.G.; Hollingsworth, P.M. DNA barcoding methods for land plants. In DNA Barcodes: Methods and Protocols; Kress, W.J., Erickson, D.L., Eds.; Humana Press: Totowa, NJ, USA, 2012; pp. 223–252. [Google Scholar]
- Ma, X.Y.; Xie, C.X.; Liu, C.; Song, J.Y.; Yao, H.; Luo, K.; Zhu, Y.J.; Gao, T.; Pang, X.H.; Qian, J.; et al. Species identification of medicinal Pteridophytes by a DNA barcode marker, the chloroplast psbA–trnH intergenic region. Biol. Phar. Bull. 2010, 33, 1919–1924. [Google Scholar] [CrossRef] [PubMed]
- Štorchová, H.; Olson, M.S. The architecture of the chloroplast psbA–trnH non-coding region in angiosperms. Plant Syst. Evol. 2007, 268, 235–256. [Google Scholar] [CrossRef]
- Dillenberger, M.S.; Kadereit, J.W. Maximum polyphyly: Multiple origins and delimitation with plesiomorphic characters require a new circumscription of Minuartia (Caryophyllaceae). Taxon 2014, 63, 64–88. [Google Scholar] [CrossRef]
- Moore, A.J.; Dillenberger, M.S. A conspectus of the genus Cherleria (Minuartia s.l., Caryophyllaceae). Willdenowia 2017, 47, 5–14. [Google Scholar] [CrossRef]
- Lu, D.; Morton, J.K. Cerastium Linnaeus. In Flora of China; Wu, C.Y., Raven, P.H., Hong, D.Y., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 2001; Volume 6, pp. 31–37. [Google Scholar]
- Hong, S.P.; Han, M.J.; Kim, K.J. Systematic significance of seed coat morphology in Silene L. s. str. (Sileneae-Caryophyllaceae) from Korea. J. Plant Biol. 1999, 42, 146–150. [Google Scholar] [CrossRef]
- Onozaki, T. Dianthus. In Ornamental Crops; Van Huylenbroeck, J., Ed.; Springer International Publishing: Cham, Switzerland, 2018; pp. 349–381. [Google Scholar]
- Adams, L.G.; West, J.G.; Cowley, K.J. Revision of Spergularia (Caryophyllaceae) in Australia. Aust. Syst. Bot. 2008, 21, 251–270. [Google Scholar] [CrossRef]
- Alonso, M.Á.; Crespo, M.B.; Martínez-Azorín, M.; Mucina, L. Taxonomic identity and evolutionary relationships of South African taxa related to the Spergularia media group (Caryophyllaceae). Plant Syst. Evol. 2021, 307, 24. [Google Scholar] [CrossRef]
- Kolodziej, B.; Okon, S.; Nucia, A.; Ociepa, T.; Luchowska, K.; Sugier, D.; Gevrenova, R.; Henry, M. Morphological, chemical, and genetic diversity of Gypsophila L. (Caryophyllaceae) species and their potential use in the pharmaceutical industry. Turk. J. Botany 2018, 42, 257–270. [Google Scholar]
- Muir, G.; Schlötterer, C. Limitations to the Phylogenetic Use of ITS Sequences in Closely Related Species and Populations—A Case Study in Quercus Petraea (Matt.) Liebl. Available online: http://webdoc.sub.gwdg.de/ebook/y/1999/whichmarker/m11/Chap11.htm (accessed on 7 February 2023).
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Fay, M.F.; Swensen, S.M.; Chase, M.W. Taxonomic affinities of Medusagyne oppositifolia (Medusagynaceae). Kew Bull. 1997, 52, 111–120. [Google Scholar] [CrossRef]
- Sang, T.; Crawford, D.J.; Stuessy, T.F. Chloroplast DNA phylogeny, reticulate evolution, and biogeography of Paeonia (Paeoniaceae). Am. J. Bot. 1997, 84, 1120–1136. [Google Scholar] [CrossRef]
- Tate, J.A. Systematics and Evolution of Tarasa (Malvaceae): An Enigmatic Andean Polyploid Genus. Ph.D. Dissertation, The University of Texas at Austin, Austin, TX, USA, 2002. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Swofford, D.L. PAUP: Phylogenetic Analysis Using Parsimony, Version 4.0b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Desper, R.; Gascuel, O. Fast and accurate phylogeny reconstruction algorithms based on the minimum-evolution principle. J. Comput. Biol. 2002, 9, 687–705. [Google Scholar] [CrossRef]
- Meier, R.; Kwong, S.; Vaidya, G.; Ng, P.K.L. DNA barcoding and taxonomy in Diptera: A tale of high intraspecific variability and low identification success. Syst. Biol. 2006, 55, 715–728. [Google Scholar] [CrossRef]
- Puillandre, N.; Brouillet, S.; Achaz, G. ASAP: Assemble species by automatic partitioning. Mol. Ecol. Resour. 2021, 21, 609–620. [Google Scholar] [CrossRef]
Figure 1.
Alignment of psbA–trnH sequences in Korean Caryophyllaceae. Short inverted repeats are indicated with color-shaded rectangles (blue and pink). Each color-shaded rectangle matches with the same color-shaded rectangles of the opposite side.
Figure 1.
Alignment of psbA–trnH sequences in Korean Caryophyllaceae. Short inverted repeats are indicated with color-shaded rectangles (blue and pink). Each color-shaded rectangle matches with the same color-shaded rectangles of the opposite side.

Figure 2.
A test of barcode gap in Korean Caryophyllaceae based on variable barcode combinations. X-axes indicate Kimura 2-parameter (K2P) distances and y-axes represent the occurrences.
Figure 2.
A test of barcode gap in Korean Caryophyllaceae based on variable barcode combinations. X-axes indicate Kimura 2-parameter (K2P) distances and y-axes represent the occurrences.
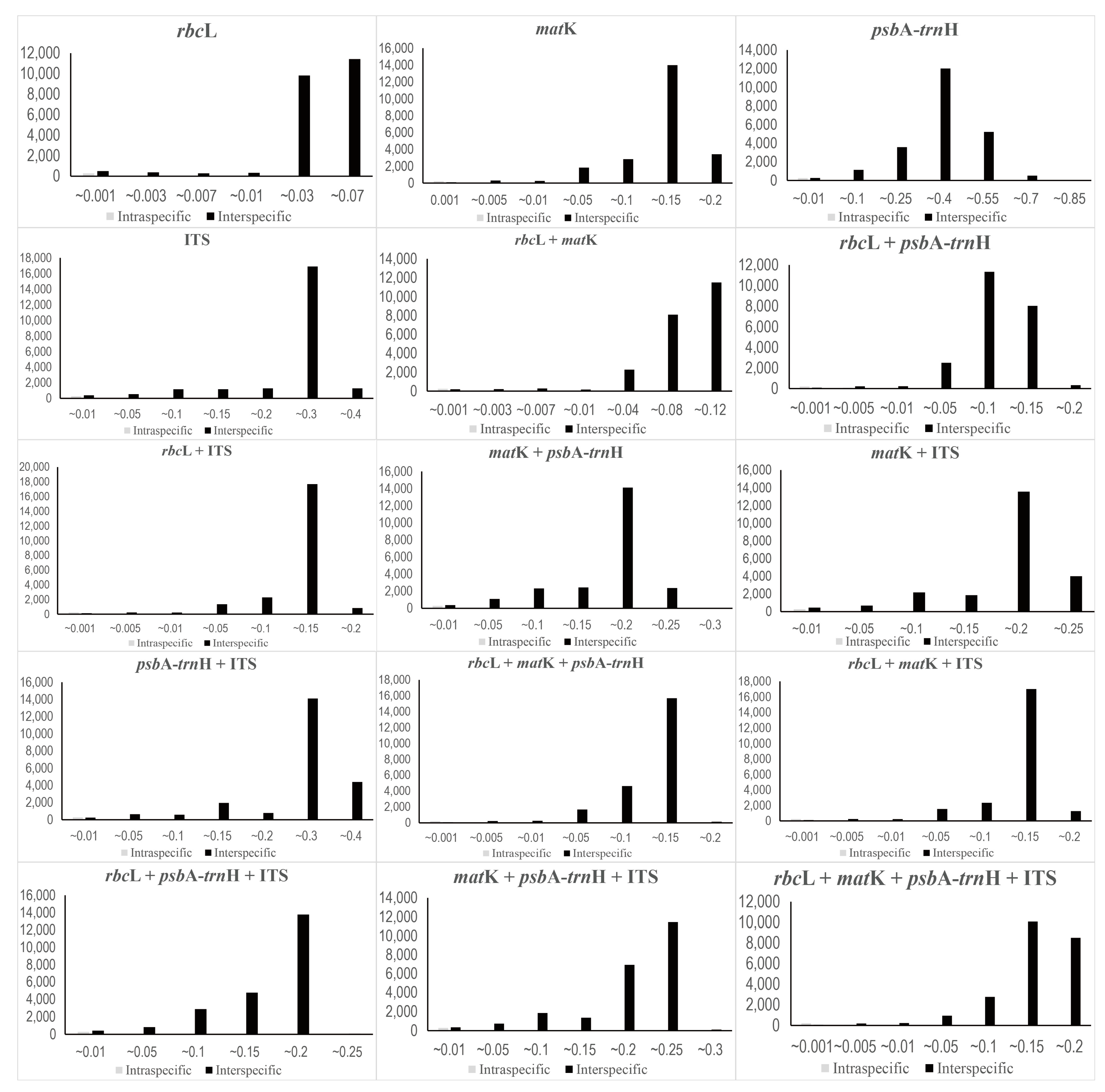
Figure 3.
The neighbor-joining phylogenetic tree of the Korean Caryophyllaceae based on combined sequence from three chloroplast DNA regions (rbcL + matK + psbA–trnH). This tree is split into (a,b). The schematic phylogenetic tree in the top left represents the combined topology (a,b). The circles on the nodes represent bootstrap values (black dot = >80%, gray dot = >50%).
Figure 3.
The neighbor-joining phylogenetic tree of the Korean Caryophyllaceae based on combined sequence from three chloroplast DNA regions (rbcL + matK + psbA–trnH). This tree is split into (a,b). The schematic phylogenetic tree in the top left represents the combined topology (a,b). The circles on the nodes represent bootstrap values (black dot = >80%, gray dot = >50%).
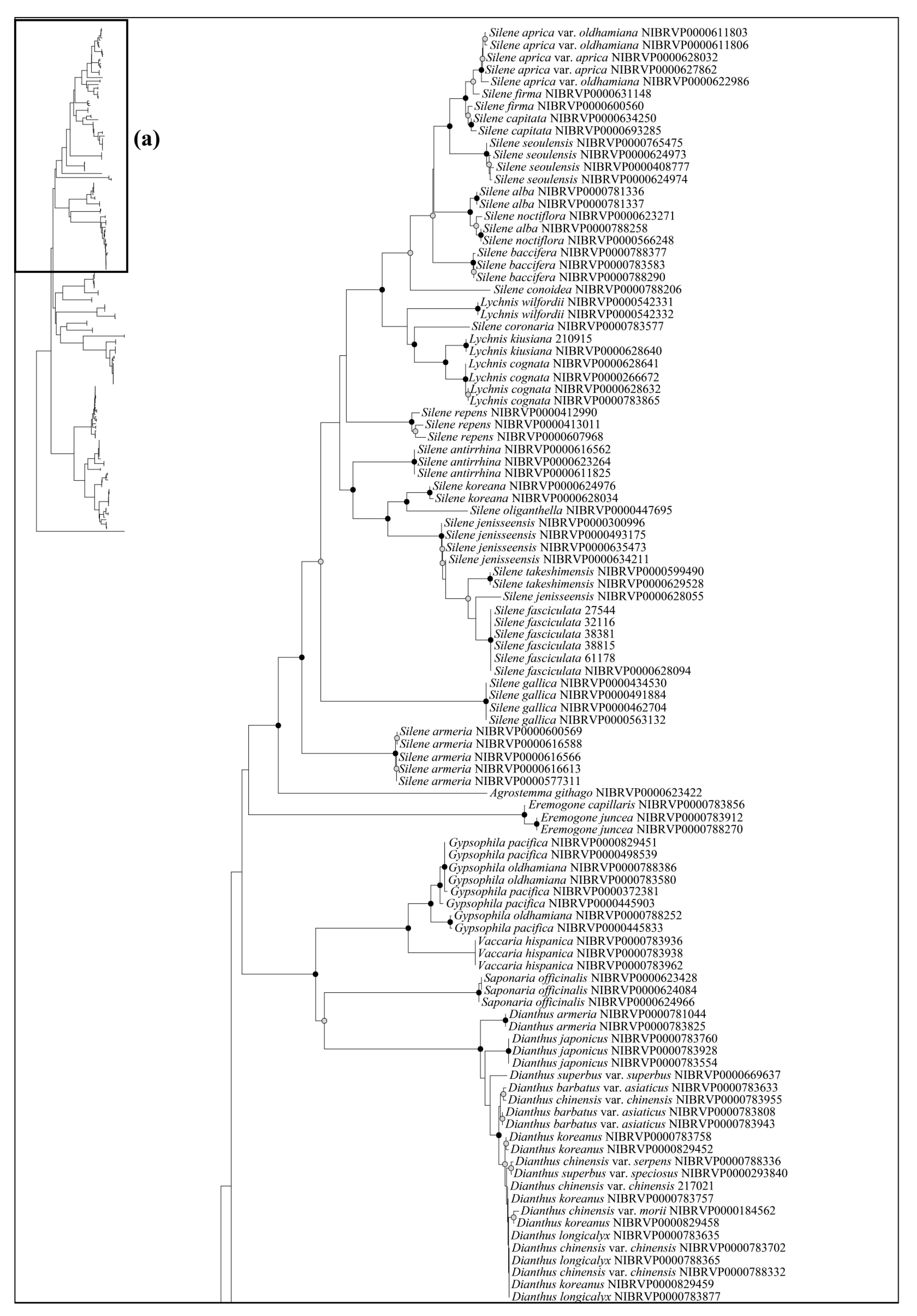
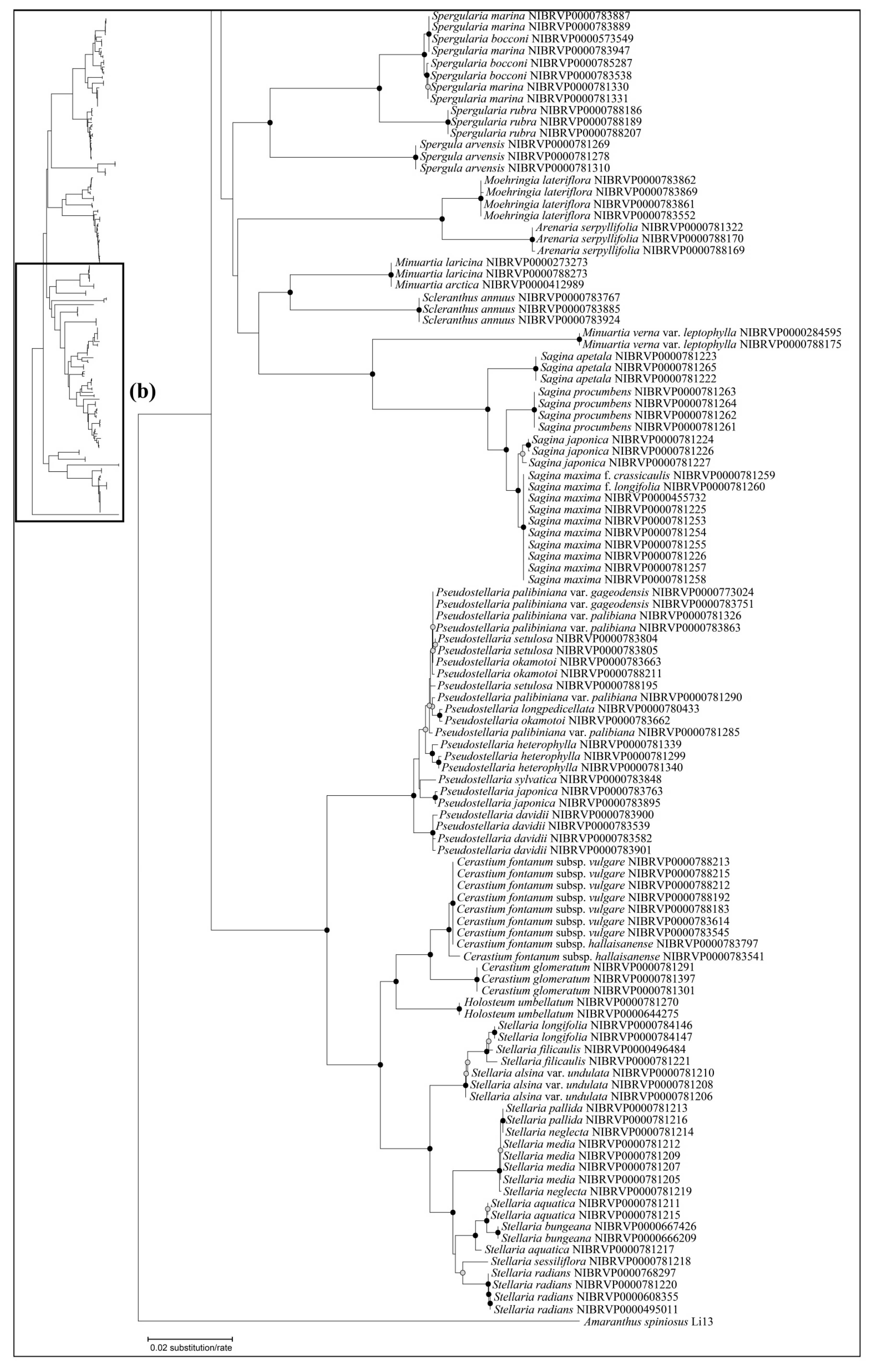
Figure 4.
The neighbor-joining phylogenetic tree of the Korean Caryophyllaceae based on internal transcribed spacer. This tree is split into (a,b). The schematic phylogenetic tree in the top left represents the combined topology (a,b). The circles on the nodes represent bootstrap values (black dot = >80%, gray dot = >50%).
Figure 4.
The neighbor-joining phylogenetic tree of the Korean Caryophyllaceae based on internal transcribed spacer. This tree is split into (a,b). The schematic phylogenetic tree in the top left represents the combined topology (a,b). The circles on the nodes represent bootstrap values (black dot = >80%, gray dot = >50%).
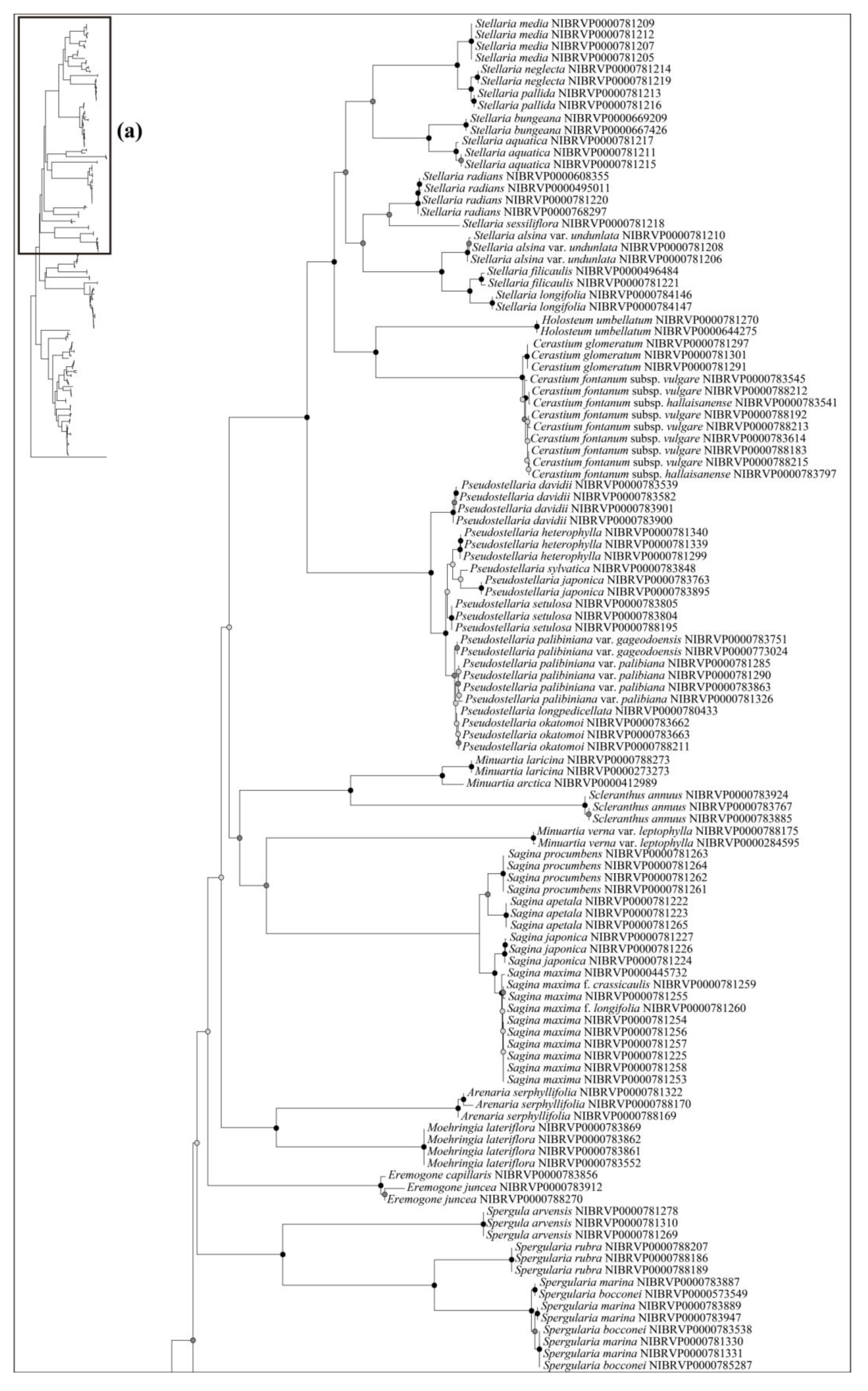
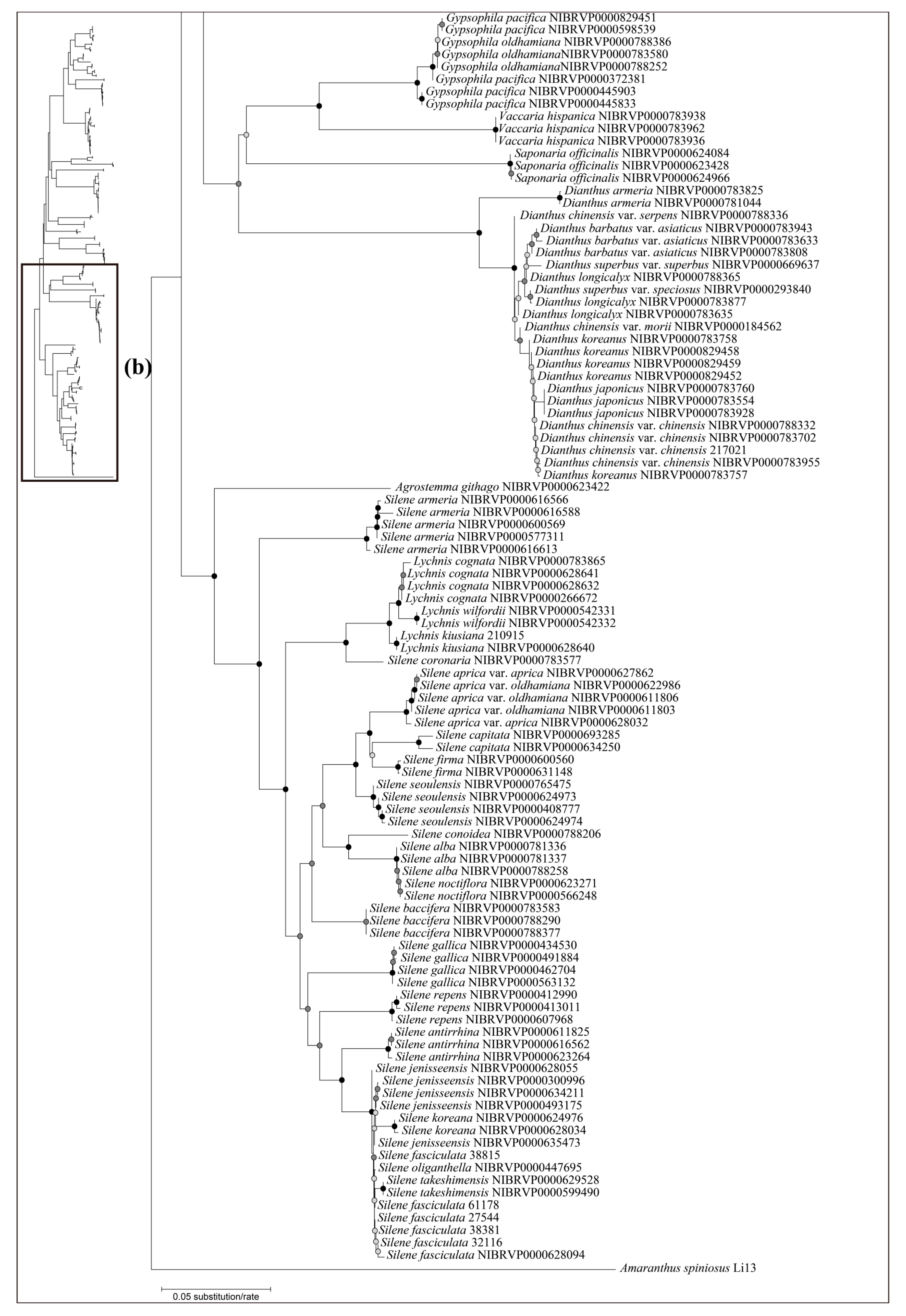
Figure 5.
The neighbor-joining phylogenetic tree of the Korean Caryophyllaceae based on combined sequence (internal transcribed spacer + rbcL). This tree is split into (a,b). The schematic phylogenetic tree in the top left represents combined topology (a,b). The circles on the nodes represent bootstrap values (black dot = >80%, gray dot = >50%).
Figure 5.
The neighbor-joining phylogenetic tree of the Korean Caryophyllaceae based on combined sequence (internal transcribed spacer + rbcL). This tree is split into (a,b). The schematic phylogenetic tree in the top left represents combined topology (a,b). The circles on the nodes represent bootstrap values (black dot = >80%, gray dot = >50%).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Statistics of four DNA barcode loci of Caryophyllaceae in Korea.
| rbcL | matK | psbA–trnH | ITS | Total | |
|---|---|---|---|---|---|
| Sequence length (bp) | 689 | 746–782 | 160–404 | 606–637 | - |
| Aligned length (bp) | 689 | 784 | 600 | 672 | 2747 |
| G + C ratio (%) | 43.0–44.3 | 29.6–33.4 | 21.1–33.3 | 50.7–63.2 | - |
| Number of variable sites (% variable sites) | 124 (18.0) | 431 (55.0) | 398 (66.3) | 393 (58.3) | 1346 (49.0) |
| Number of informative sites (% variable sites) | 115 (16.7) | 411 (52.4) | 371 (61.8) | 381 (56.5) | 1278 (46.5) |
Table 2.
Summary of the pairwise intraspecific and interspecific distances in the DNA barcode loci of Caryophyllaceae species.
Table 2.
Summary of the pairwise intraspecific and interspecific distances in the DNA barcode loci of Caryophyllaceae species.
| Region | Intraspecific K2P Distance | Interspecific K2P Distance | ||||
|---|---|---|---|---|---|---|
| Min. | Mean | Max. | Min. | Mean | Max. | |
| rbcL | 0 | 0.0004 | 0.0313 | 0 | 0.0291 | 0.0576 |
| matK | 0 | 0.0019 | 0.1296 | 0 | 0.1140 | 0.1754 |
| psbA–trnH | 0 | 0.0078 | 0.4690 | 0 | 0.3245 | 0.7045 |
| ITS | 0 | 0.0037 | 0.2478 | 0 | 0.2259 | 0.3531 |
| rbcL + matK | 0 | 0.0012 | 0.0817 | 0 | 0.0726 | 0.1126 |
| rbcL + psbA–trnH | 0 | 0.0022 | 0.1284 | 0 | 0.0888 | 0.1888 |
| rbcL + ITS | 0 | 0.0019 | 0.1246 | 0 | 0.1141 | 0.1657 |
| matK + psbA–trnH | 0 | 0.0033 | 0.2015 | 0 | 0.1548 | 0.2599 |
| matK + ITS | 0 | 0.0008 | 0.0072 | 0 | 0.1609 | 0.2340 |
| psbA–trnH + ITS | 0 | 0.0048 | 0.3077 | 0 | 0.2502 | 0.3754 |
| rbcL + matK + psbA–trnH | 0 | 0.0021 | 0.1289 | 0 | 0.1003 | 0.1658 |
| rbcL + matK + ITS | 0 | 0.0019 | 0.1265 | 0 | 0.1140 | 0.1622 |
| rbcL + psbA–trnH + ITS | 0 | 0.0028 | 0.1724 | 0 | 0.1405 | 0.2113 |
| matK + psbA–trnH + ITS | 0 | 0.0011 | 0.0110 | 0 | 0.1809 | 0.2666 |
| rbcL + matK + psbA–trnH + ITS | 0 | 0.0008 | 0.0077 | 0 | 0.1314 | 0.1936 |
Table 3.
Species identification based on phylogenetic analysis and the best close match.
| Region | Phylogenetic Analysis | Best Close Match | |||
|---|---|---|---|---|---|
| Correct | Ambiguous | Incorrect | No Match | ||
| rbcL | 44.74% | 84 (39.06%) | 121 (56.27%) | 10 (4.65%) | 0 (0.00%) |
| matK | 56.58% | 115 (53.48%) | 74 (34.41%) | 25 (11.62%) | 1 (0.46%) |
| psbA–trnH | 59.21% | 139 (64.65%) | 50 (23.25%) | 23 (10.69%) | 3 (1.39%) |
| ITS | 76.32% | 149 (69.30%) | 48 (22.32%) | 16 (7.44%) | 2 (0.93%) |
| ITS + matK | 73.68% | 160 (74.41%) | 34 (15.81%) | 20 (9.30%) | 1 (0.46%) |
| ITS + psbA–trnH | 76.32% | 164 (76.27%) | 18 (8.37%) | 30 (13.95%) | 3 (1.39%) |
| ITS + rbcL | 77.63% | 152 (70.69%) | 45 (20.93%) | 17 (7.90%) | 1 (0.46%) |
| matK + psbA–trnH | 61.84% | 141 (65.58%) | 43 (20.00%) | 28 (13.02%) | 3 (1.39%) |
| rbcL + matK | 60.53% | 124 (57.67%) | 64 (29.76%) | 27 (12.55%) | 0 (0.00%) |
| rbcL + psbA–trnH | 61.84% | 141 (65.58%) | 44 (20.46%) | 28 (13.02%) | 2 (0.93%) |
| ITS + matK + psbA–trnH | 73.68% | 165 (76.74%) | 20 (9.30%) | 28 (13.02%) | 2 (0.93%) |
| ITS + rbcL + matK | 73.68% | 163 (75.81%) | 28 (13.02%) | 23 (10.69%) | 1 (0.46%) |
| ITS + rbcL + psbA–trnH | 76.32% | 164 (76.27%) | 17 (7.90%) | 32 (14.88%) | 2 (0.93%) |
| rbcL + matK + psbA–trnH | 59.21% | 146 (67.9%) | 32 (14.88%) | 36 (16.74%) | 1 (0.46%) |
| ITS + rbcL + matK + psbA–trnH | 75.00% | 165 (76.74%) | 18 (8.37%) | 31 (14.41%) | 1 (0.46%) |
Table 4.
List of primers for the four regions sequenced in this study.
| Region | Primer Name | Primer Sequence (′5-′3) | Reference |
|---|---|---|---|
| ITS | ITS18S-A | CCTTMTCATYTAGAGGAAGGAG | Muir & Schlötterer [69] |
| ITS4 | TCCTCCGCTTATTGATATGC | White et al. [70] | |
| rbcL | rbcL 1F | ATGTCACCACAAACAGAAAC | Fay et al. [71] |
| rbcL 724R | TCGCATGTACCTGCAGTAGC | ||
| matK | matK_3F | CGTACAGTACTTTTGTGTTTACGAG | Kim K.J. (unpublished work) |
| matK_1R | ACCCAGTCCATCTGGAAATCTTGGTTC | ||
| psbA–trnH | psbA_sang | GTTATGCATGAACGTAATGCTC | Sang et al. [72] |
| trnH_tate | CGCGCATGGTGGATTCACAATCC | Tate [73] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Jin, D.-P.; Sim, S.; Park, J.-W.; Choi, J.-E.; Yoon, J.; Lim, C.E.; Kim, M.-H. Identification of the Plant Family Caryophyllaceae in Korea Using DNA Barcoding. Plants 2023, 12, 2060. https://doi.org/10.3390/plants12102060
AMA Style
Jin D-P, Sim S, Park J-W, Choi J-E, Yoon J, Lim CE, Kim M-H. Identification of the Plant Family Caryophyllaceae in Korea Using DNA Barcoding. Plants. 2023; 12(10):2060. https://doi.org/10.3390/plants12102060
Chicago/Turabian StyleJin, Dong-Pil, Sunhee Sim, Jong-Won Park, Ji-Eun Choi, Jiwon Yoon, Chae Eun Lim, and Min-Ha Kim. 2023. "Identification of the Plant Family Caryophyllaceae in Korea Using DNA Barcoding" Plants 12, no. 10: 2060. https://doi.org/10.3390/plants12102060
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.