
Epigenetic Modifications Related to Potato Skin Russeting




1
Institute of Plant Sciences, Agricultural Research Organization, Volcani Institute, 68 HaMacabim Road, P.O. Box 15159, Rishon LeZion 7505101, Israel
2
Department of Horticulture, University of Wisconsin-Madison, Madison, WI 53706, USA
*
Author to whom correspondence should be addressed.
Plants 2023, 12(10), 2057; https://doi.org/10.3390/plants12102057
Submission received: 14 March 2023
/
Revised: 17 May 2023
/
Accepted: 18 May 2023
/
Published: 22 May 2023
(This article belongs to the Special Issue Beyond Mendel's Laws)
Abstract
:Potato tuber skin is a protective corky tissue consisting of suberized phellem cells. Smooth-skinned varieties are characterized by a clean, shiny appearance compared to the darker hue of russeted potatoes. The rough skin of russeted cultivars is a desired, genetically inherited characteristic; however, unwanted russeting of smooth-skinned cultivars often occurs under suboptimal growth conditions. The involvement of epigenetic modifiers in regulating the smooth skin russeting disorder was tested. We used smooth-skin commercial cultivars with and without the russeting disorder and three lines from a breeding population segregating for russeting. Anatomically, the russet skin showed similar characteristics, whether the cause was environmentally triggered or genetically determined. The old outer layers of the corky phellem remain attached to the newly formed phellem layers instead of being sloughed off. Global DNA methylation analysis indicated a significant reduction in the percentage of 5-methylcytosine in mature vs. immature skin and russet vs. smooth skin. This was true for both the smooth-skin commercial cultivars and the russeted lines. The expression level of selected DNA methyltransferases was reduced in accordance. DNA demethylase expression did not change between the skin types and age. Hence, the reduced DNA methylation in mature and russet skin is more likely to be achieved through passive DNA demethylation and loss of methyltransferase activity.
Keywords:
cork; DNA methylation; epigenetic regulation; periderm; phellem; phellogen; russeting; Solanum tuberosum; tuber skin1. Introduction
Potato tubers are covered with a protective corky tissue called the periderm. The periderm comprises three cell types: phellem, phellogen, and phelloderm [1,2]. The phellem forms a series of layers at the outermost level of the periderm and is derived from the meristematic phellogen layer (or cork cambium) below it. As phellem cells develop, they become suberized and die, forming a protective corky layer termed ‘skin’. Suberin biosynthesis in the phellem has been studied extensively [3,4].
During tuber expansion, phellogen cell division continuously adds new skin layers, whereas superficial cork cells are sloughed off, rendering the skin smooth and shiny. Some physiological disorders of the potato tuber are related to abnormal development of the skin, including russeting of smooth-skinned potatoes. The russeted skin appears rough to the touch, with a darker tint compared to the characteristic shiny look of the smooth skin, and is often rejected by consumers. The rough skin of russeted varieties (e.g., the well-known US variety ‘Russet Burbank’) is a desired, genetically determined trait [5].
Impaired skin development leading to russeting is often associated with suboptimal growth conditions (i.e., environmentally triggered russeting) [6]. The phenomenon is not caused by pathogens [7], and the direct causes in the field are not clear. Nevertheless, calcium fertilization mitigates russeting occurrence and severity [7]. In contrast, increased potassium concentration in the soil solution competes with the binding of magnesium and calcium to the tuber skin and is associated with low skin quality [8]. Moreover, the disorder is enhanced in sandy soils (R. Erel, Gilat Center, Israel, unpublished data).
Tubers with russet skin have a thicker layer of phellem than smooth-skinned potatoes [6,7,9,10]. It can result from increased activity of phellem formation—for example, due to high soil temperature [6,11]. Alternatively, thicker phellem can result from strong adherence of “old” phellem to the new layers that form below, so that they are not sloughed off during tuber development; this may be due to increased suberization [10] or increased levels of pectin and hemicellulose [9] in phellem cell walls. Nevertheless, new phellem layers are formed below the old layers, and the latter are cracked, resulting in netted or russeted skin. Whether environmentally triggered and genetically determined russeting share the same genes and biosynthesis pathways is unclear. The roughness of ‘Russet Burbank’ skin depends on growth temperatures [10] and the quantity and source of potassium fertilizer [12], suggesting that common genes might be involved.
Epigenetic factors such as DNA methylation and histone modifications were shown to play a role in phellem differentiation of cork oak, and differed between high and low phellem/cork quality [13,14,15]. Increased expression of DNA methyltransferases (DNMTs) was positively correlated with low cork quality, suggesting that gene silencing leads to a high potential for defects [15]. The russet skin disorder that develops on smooth-skin potato cultivars may be viewed as a cork of low quality [7].
Potato exhibits strong interaction of genotype and environment. Epigenetic regulation governs plant response to extreme growth conditions and allows phenotypic plasticity by regulating gene expression levels [16,17]. The potato periderm is a protective tissue enriched with stress-related factors [18] and responds to stress conditions. For example, temperature stress enhanced the formation and accumulation of skin layers to create a thick protective cover [6]—this demonstrates the strong impact of environmental cues on cork development and the potential for epigenetic modulation of its quality. Agronomical practices may also induce chromatin modifications that affect skin quality [19,20].
Epigenetic mechanisms change the phenotypic traits in the plant without changing the genetic sequences. Examples are DNA methylation, histone modifications, and histone variants. DNA methylation is based on a dynamic addition or removal of a methyl group. In plants, it occurs at the fifth position of the cytosine ring in three contexts: CG, CHG, and CHH (H = C, T, or A), and is mediated by the RNA-directed DNA methylation (RdDM) pathway, involving small interfering RNAs (siRNAs) and scaffold RNAs in addition to protein complexes [21]. Four DNMT families differentially regulate cytosine methylation. METHYLTRANSFERASE1 (MET1) recognizes hemimethylated CG following DNA replication and methylates the naked cytosine in the daughter strand [21,22]. Chromomethylases (CMTs) bind histone H3 lysine 9 (H3K9me2) heterochromatin to methylate non-CG contexts [23]. In flowering plants, CMT3 methylates mostly CHG sites, whereas CMT2 methylates mostly CHH sites [24,25]. The CHH methylation state by CMT2 is maintained by DOMAIN REARRANGED METHYLTRANSFERASE 2 (DRM2) through the RdDM pathway [25,26].
Passive DNA demethylation results from an absence of DNMTs, a reduction in their activity, or a shortage of methyl donors following DNA replication [27]. In plants, active DNA demethylation is mediated by 5-methylcytosine DNA glycosylases through a DNA base-excision repair pathway [28]. There are four 5-methylcytosine DNA glycosylases in Arabidopsis: REPRESSOR OF SILENCING 1 (ROS1), DEMETER (DME), DEMETER-LIKE 2 (DML2), and DEMETER-LIKE 3 (DML3) [28].
To test whether epigenetic modification is involved in potato skin maturation and russeting, we used smooth-skin potato commercial cultivars with and without the russeting disorder (environmentally triggered russeting), in addition to three lines from a breeding population segregating for russeting (i.e., genetically determined russeting)—one smooth and two russeted to different levels. An anatomical study was used to define the different russeting phenotypes, global DNA methylation (GDM) was determined for smooth and russet skin types, and the expression level of selected DNA modifier genes was monitored.
2. Results
2.1. Description of Russet Skin
The smooth-skinned cultivars used in the study are characterized by a clean, shiny appearance (Figure 1a–c); Dèsirèe and Rosanna also exhibit a red/purplish tint due to the accumulation of anthocyanins in their periderm. Skin with environmentally triggered russeting that develops on the surface of the cultivars Jazzy and Georgina appears with a darker hue without the shiny look, as well as fine cracking (Figure 1b,c). The russet on the surface of the 43_wR and 25_hR lines is coarser, rough to the touch, and with netting-like morphology (Figure 1d); the latter exhibits a darker tint than the former.
2.2. Anatomical Study of Russeted Skin
An anatomical study was conducted to distinguish between the morphologies of the two skin types, smooth vs. russet. Sections through the surface of Rosanna tuber demonstrate typical morphology of the skin made of phellem cells. The phellem are rectangular flattened cells arranged in columns of ca. ten layers, and their suberized cell walls are auto-fluorescing under UV light (Figure 2a).
Smooth skin of cv. Jazzy shows similar morphology of skin organization, albeit fewer cell layers (ca. five layers; Figure 2b). The russet skin of Jazzy seems more condensed than the smooth skin. New phellem cell layers are formed underneath the russet layers, although the skin was already cured and adhering firmly to the tuber flesh (Figure 2c). The old phellem cells remain attached to the newly formed skin layers. During normal skin development, these cells are sloughed off, rendering the skin smooth and shiny.
The skin of the smooth line, 16_S, exhibits the characteristic phellem cell columns that are nicely arranged and with numerous cell layers (Figure 3a). The russet skin of lines 43_wR and 25_hR has patches of dead phellem that adhered to the newer phellem layers underneath them—in 43_wR with weak russeting, this phenomenon is finer compared to 25_hR with heavy russeting (Figure 3b,c, respectively). Note the autofluorescence signal of the russet skin fragments under UV light is weaker than that of the native phellem.
2.3. Global DNA Methylation of Russeted Skin
To examine whether russeting is associated with epigenetic regulation of gene expression, GDM was determined in DNA extracted from russet and smooth skin. First, a comparison was made between the immature and mature skin of smooth cultivars Dèsirèe and Rosanna to monitor the status of the GDM at two developmental stages of the skin. This was done for two years for each cultivar. Results showed a significant reduction in GDM status in mature skin compared to immature skin (Figure 4a).
Comparison of the smooth and russet skin of Jazzy and Georgina showed a significant reduction in GDM levels in the russet skin (Figure 4b). Reduced GDM levels were also monitored using the breeding lines 16_S, 43_wR, and 25_hR—each line showed significantly lower levels in the mature compared to immature developmental stage of the skin, irrespective of smooth or russet skin type (Figure 4c). Moreover, in the stage of skin immaturity, the russet skin type of 43_wR, and 25_hR had reduced GDM levels compared to the smooth skin type of 16_S (Figure 4c). Thus, the results obtained for the tested breeding lines supported the previous data obtained for the smooth skin cultivars with and without the environmentally triggered russeting—reduced GDM levels in maturing and in russeted skin.
2.4. Expression of Epigenetic Modifiers in Russeted Skin
A literature survey was conducted to identify genes coding for DNMT and DNA demethylases that may be responsible for the DNA epigenetic modification associated with russeting—both the environmentally triggered and genetically determined traits. Similar genes were also identified in previous work on potato phellogen transcriptome (Table S1; [11]). Gene expression of selected epigenetic modifiers was tested in the same skin samples as above. This included the DNA demethylases DML1, DML3, and DME, and the DNMTs MET1/2, CMT3, and DRM2. The expression of two markers for suberization, CER6, and CYP86A33, was used as a reference to phellem cell activity. A comparison of Dèsirèe’s immature and mature skin over two years indicated similar activity of the suberization markers in both developmental stages (Figure 5a,b). Met1/2 was significantly downregulated in the mature skin in both years; DRM2 and DME were strongly downregulated in 2017 and CMT3 in 2018. When gene expression was monitored in the smooth vs. russet skin of Jazzy and Georgina, results indicated reduced expression of suberization genes in the russet skin, but the differences were not always statistically significant (Figure 5c,d). In both cultivars, DRM2 was significantly downregulated in the russet skin.
Comparisons of immature vs. mature skin for the smooth line 16_S and for the russeted line 25_hR pointed again at the association of reduced expression of MET1/2 and DRM2 in mature skin, albeit not always significantly (Figure 6a,b). DML3 was also downregulated in the mature skin of 16_S (Figure 6a). When all three breeding lines were compared at their immature stage (when skin formation is most active), DML3 was significantly reduced in the russeted genotypes. CMT3 and DRM2 were significantly reduced in 43_wR and MET1/2 in 25_hR when compared to the smooth genotype 16_S (Figure 6c). Overall, it can be concluded that skin aging and russeting are similarly associated with reduced activity of DNMTs, resulting in reduced GDM levels in the respective skin types.
3. Discussion
Environmentally triggered russeting of smooth skin cultivars and genetically determined russeting appear as brownish regions on the surface of the tubers. Tissue color or roughness, and the degree of surface coverage, may differ between genotypes, but russet tissue morphology has a common characteristic: the old outer layers of the corky phellem remain attached to the newly formed phellem layers instead of being sloughed off. These superficial layers have a compressed structure that is less organized than the phellem layers of the smooth skin type, and they crack as the tuber expands [6,7]. The outcome is russeted skin that looks like flakes of dry skin adhered to the surface of the tuber or as netted rough skin.
During recent years, we applied diverse experimental approaches to cope with potato skin disorders, including various protocols for haulm desiccation [29] and mineral fertilization [7,8,30]. These were complemented with proteomic and transcriptomic studies of skin development [6,18,31] and the identification of skin/suberin-related genes [32]. Although the treatments we have deployed slightly improved skin appearance, none “cured” the costly problem of the environmentally triggered russeting in smooth skin cultivars. This motivated us to look for an epigenetic mechanism that may indicate tuned regulated expression of known genes during russet development and of unknown regulating sequences that could not be detected by the omics approaches we deployed previously.
GDM analysis of potato skin indicated a significant reduction in the percentage of 5-methylcytosine in maturing skin compared to immature skin, and in russet compared to smooth skin. This was true for smooth skin commercial cultivars Dèsirèe, Rosanna, Jazzy, and Georgina and the breeding lines segregating for russeting. In accordance, the gene expression level of selected DNMTs was reduced in mature vs. immature skin and in russet vs. smooth skin in all the potatoes tested. In most cases, DRM2 was dramatically downregulated, albeit not always significantly. MET1/2 was mainly downregulated in mature vs. immature smooth skin and in genetically determined heavy russeting but not in environmentally triggered russeting. The CMT3 expression profile was not conclusive among the tested samples. Of the DNA demethylases, DML3 was downregulated in maturing smooth skin of 16_S and the russeted lines compared to the smooth line.
DNA demethylation at a subset of regions relies on active DNA demethylation initiated by DNA glycosylases ROS1, DML2, and DML3 but can also occur passively [33]. In potato skin, reduced DNA methylation level in aging or russeted skin is more likely to be achieved through passive DNA demethylation and loss of DNMT activity.
Overall it is concluded that skin aging and russeting involve reduced DNA methylation associated with passive demethylation and reduced activity of DNMTs.
Tuber skin is a model system for plant corky tissues and may be compared to the cork of trees. Epigenetic factors such as DNA methylation and histone modification were shown to play a role in cork differentiation of cork oak (Quercus suber) and with altered patterns between cork of high and low quality [13,14,15]. Orthologs of genes involved in epigenetic modifications of oak phellogen and cork quality were found in the transcriptome of potato skin phellogen [11]. In general, the russet skin of smooth potato cultivars can also be viewed as a cork of low quality [7].
In potato skin, DNA methylation level and DRM2 expression were positively associated—high DRM2 gene expression and high DNA methylation level—while the contribution of other DNMT activities seems less consistent between samples. These high activities were associated with smooth skin (corky skin of high quality). In accordance, in the cork of Q. suber, DRM2 was the most active methyltransferase compared to MET1, MET2, and CMT3 [15]. However, contrary to what was shown for the potato skin, Q. suber cork of high quality was associated with lower levels of DNMTs gene expression. Yet, the trend of GDM was similar in potato and Q. suber—high in cork of high quality. Thus, in Q. suber there was a negative correlation between the relative levels of expression of DNMTs and GDM [15]. Nevertheless, it was suggested that gene silencing is higher in cork of low quality, leading to a higher potential of defects [15].
Conversely, in potato, the level of GDM and related gene expression were lower in maturing and russeted skin (corky skin of low quality). Therefore, it can be speculated that skin maturation requires fewer cellular specifications, hence less gene silencing by methylation, compared to the early stages of skin formation. Moreover, reduced gene regulation by methylation leads to “abnormal” development, hence to russeting.
DNA methylation provides stringent regulation or fine-tuning of gene expression in controlling phenotypic variation in response to environmental factors or at critical developmental stages [17,27]. In potato, increased DNA methylation was reported for photoperiod-sensitive tuberization [34] and in response to heat [35] and salt stress [36]. Accordingly, we hypothesize that reduced DNA methylation in aging or russeted skin reflects weak control of skin developmental processes and, thus, the formation of russeted skin instead of smooth. This is further demonstrated when we examine skin/periderm developmental stages.
The transcriptome of potato phellogen included DNA markers for epigenetic regulation [11]. The highest expression (given by FPKM) was related to genes involved in chromatin assembly/remodeling, as expected for cambial tissue that is extensively dividing. Components of RdDM pathway were also expressed, however, to a lesser extent. This included DRM2, AGO4, DDM1, DRD1, and RDR2 [11,25,27]. The expression of DML1/ROS1 with demethylation activity was very low, as well as histone demethylation activity. Transcripts for MET1, MET2, and CMT3 that maintain the methylation sites were not detected at that early stage of development. Overall, it is suggested that in the phellogen, initiation of methylation activity allows the specific programming of developmental pathways towards the formation of suberized phellem (skin; external layers of the periderm) and the parenchyma-like phelloderm (internal periderm layers). Thus, in smooth skin cultivars, high DNA methylation directs the formation of the skin of high quality. Reduction in DNA methylation suggests a change of program or looseness of controlled processes that leads to russeting.
Data also imply that skin russeting may share a similar DNA methylation state as the skin maturation process. Whether the same set of structural and regulatory genes are epigenetically regulated requires further clarification.
4. Materials and Methods
4.1. Plant Material
The study included four commercial cultivars of potato (Solanum tuberosum L.)—Dèsirèe, Rosanna, Jazzy, and Georgina—and three UW-Madison breeding lines from North American russet germplasm, which were selected from two populations segregating for russeting: W17071 = AW07791-2rus x AW07791-2rus; and W17069 = AW06108-rus x AW07791-2rus. The three breeding lines were as follows: W17069–16 (abbreviated 16_S) had smooth skin; W17071-43 (abbreviated 43_wR) had weak russeting; and W17071-25 (abbreviated 25_hR) had heavy russeting (Figure 1). Dèsirèe and Rosanna are red-skin cultivars and were used in this work interchangeably.
Tubers of Jazzy and Georgina were sampled from commercial fields in the northern part of the Negev region of Israel at harvest time in 2019–2020. These were sorted into two groups—tubers with the cultivar characteristic smooth skin and tubers with an environmentally triggered russeting. Plants of Dèsirèe, Rosanna, and the 16_S, 43_wR, and 25_hR genotypes were grown in 20-L containers in a greenhouse in Volcani Institute, in 2016-2018 and 2022, respectively, under natural winter conditions (November–January, average temperature range of 10–18 °C). Tubers were collected at two developmental stages—at 7–8 weeks post sprout emergence when the skin is immature and can be easily separated by the hand from the tuber flesh, and at 10–12 weeks post sprout emergence during skin maturation and skin-set. For each cultivar and the breeding lines, three individual plants were harvested at each time point. Tubers of the same plant were pooled for sampling.
At sampling, tissue blocks were taken from the tuber surface for anatomical study of the skin, and skin tissue was collected for the molecular analyses described below. The latter was frozen in liquid nitrogen and stored at −80 °C until use.
4.2. Anatomical Studies
Tissue blocks from the surface of the tubers were fixed in FAA (50% ethanol, 5% acetic acid, and 3.7% formaldehyde, v/v, in water), dehydrated in an ethanol/xylene series, and embedded in paraplast (Surgipath Paraplast Plus, Leica Biosystems Richmond Inc., Richmond, IL, USA), according to standard methods [37]. Tissue sections (16 µm) were stained with Safranin-O/Fast green (Sigma-Aldrich, Jerusalem, Israel) [38] and were observed under the microscope (Leica DMLB, Germany) through a CCD camera (Leica DC2000) by using the Leica IM1000 program. Visible light was used to examine skin tissue morphology, and UV light to view the autofluorescence of the phellem-suberized cell walls.
4.3. Total DNA Extraction and Global DNA Methylation (GDM) Assay
Total DNA was extracted from skin tissue using the DNeasy® Plant Pro Kit (QIAGEN GmbH, Germany). Purified DNA was quantified in the NanoDropTM 2000/2000c (Thermo Scientific, Waltham, MA, USA). Determination of GDM was performed using the MethylFlash™ Global DNA Methylation (5-mC) ELISA kit (Colorimetric) (Epigentek, USA catalog no. # P-1030). Tested genomic DNA was used at 100 ng per reaction according to the kit protocol. A standard curve was prepared with a solution of methylated DNA containing 5% of 5-methylcytosine at 5 ng/μL and 50 μg/mL. The negative control was a solution of unmethylated DNA. Both solutions were provided in the kit. According to the assay protocol, a standard curve was prepared at 0, 0.2%, 1%, and 5% of 5-methylcytosine using the standard solutions. Absorbance at 450 nm was recorded using the Synergy H1 Hybrid Reader Photometer (BioTek, Agilent Technologies Inc., Santa Clara, CA, USA). Analyses were performed with three biological replicates, each with two technical replicates. Data were analyzed for significance by Tukey’s HSD test (p < 0.05) using the JMP16 software (http://www.jmp.com)
4.4. Total RNA Extraction, cDNA Synthesis, and Quantitative Reverse Transcriptase PCR (qRT-PCR)
Total RNA was extracted from skin samples according to Ginzberg et al. [6]. cDNA synthesis was done using QuantiTect Reverse Transcription Kit (QIAGEN GmbH, Germany). LightCycler® 480 SYBR Green I Master (Roche Diagnostics GmbH, Mannheim, Germany) was used for qRT-PCR according to the manufacturer’s protocol with specific primers (Table 1). Gene expression data was quantified using a standard curve—a mixture of the tested cDNA samples at equal amounts was diluted 1:30, 1:240, and 1:1920 and used in each qRT-PCR with the specific primers. Reactions with the individual tested cDNAs and the mixed samples for the standard curve were run in a Rotor-Gene 6000 Real-Time PCR machine (Corbett Research Ltd., Sydney, NSW, Australia), and the build-in software calculated relative gene expression values. Each qRT-PCR was performed with three biological replicates, each with three technical replicates. Values in each sample were normalized to the expression levels of the reference gene, α-CHAIN OF THE NASCENT POLYPEPTIDE-ASSOCIATED COMPLEX (α-NAC) [6]. Data were analyzed for significance by Student’s t-test (p < 0.05) using the JMP16 software (http://www.jmp.com).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Primers used in the study.
| Gene Code | Gene | Potato ID | Forward Primers | Reverse Primers |
|---|---|---|---|---|
| DML1 | DEMETER-LIKE DNA GLYCOSYLASE 1 | Soltu.DM.09G004240 | CCACCAAATGTTTCTCGACC | TGTTGTGGCTTCAATACCCT |
| DML3 | DEMETER-LIKE DNA GLYCOSYLASE 3 | Soltu.DM.04G017430 | CTGAGCACACAGTTTTGAGG | ATTGCTCTCAGCCTGTAACC |
| DME | DNA GLYCOSYLASE DEMETER | Soltu.DM.11G005260 | ATTCAAAGGCCTAACCACAG | CTGGCTTTCCTTTTGTCCTA |
| MET1 | DNA (CYTOSINE-5)-METHYLTRANSFERASE 1 | Soltu.DM.11G013230 | TAAGGTGGGGATGTGCTTTC | GTGCATATGCCAAAGGAGGT |
| CMT3 | CHROMOMETHYLASE 3 | Soltu.DM.12G000130 | ACTTGCCTGGAGTTCGTGTT | GTAGTTCCTCAAAACCGTTT |
| DRM2 | DOMAINS REARRANGED | Soltu.DM.02G006560 | GGATCAGGAAAGCTGTGGAG | GGTTCTTTGGAAACCCCAAT |
| NAC | NASCENT POLYPEPTIDE-ASSOCIATED COMPLEX | Soltu.DM.09G002650 | ATATAGAGCTGGTGATGACT | TCCATGATAGCAGAGACTA |
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants12102057/s1. Table S1. Epigenetic markers in the transcriptome of potato phellogen.
Author Contributions
Conceptualization, I.G.; formal analysis, P.K. and Y.K.; resources, I.G. and J.B.E.; writing—original draft preparation, I.G. and P.K.; writing—review and editing, I.G. and J.B.E. All authors have read and agreed to the published version of the manuscript.
Funding
Development of the UW-Madison potato breeding lines was supported by the USDA National Institute of Food and Agriculture Award 2019-34141-30284.
Data Availability Statement
The data presented in this study is contained within the article and the Supplementary Material Table S1.
Acknowledgments
The authors thank Mike Thornton from the University of Idaho for reviewing the manuscript. The research is a contribution of the Agricultural Research Organization, Volcani Institute.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Reeve, R.M.; Hautala, E.; Weaver, M.L. Anatomy and compositional variation within potatoes 1. Developmental histology of the tuber. Am. Potato J. 1969, 46, 361–373. [Google Scholar] [CrossRef]
- Kumar, P.; Ginzberg, I. Potato periderm development and tuber skin quality. Plants 2022, 11, 2099. [Google Scholar] [CrossRef] [PubMed]
- Woolfson, K.N.; Esfandiari, M.; Bernards, M.A. Suberin biosynthesis, assembly, and regulation. Plants 2022, 11, 555. [Google Scholar] [CrossRef]
- Nomberg, G.; Marinov, O.; Arya, G.C.; Manasherova, E.; Cohen, H. The key enzymes in the suberin biosynthetic pathway in plants: An update. Plants 2022, 11, 392. [Google Scholar] [CrossRef] [PubMed]
- De Jong, H. Inheritance of russeting in cultivated diploid potatoes. Potato Res. 1981, 24, 309–313. [Google Scholar] [CrossRef]
- Ginzberg, I.; Barel, G.; Ophir, R.; Tzin, E.; Tanami, Z.; Muddarangappa, T.; De Jong, W.; Fogelman, E. Transcriptomic profiling of heat-stress response in potato periderm. J. Exp. Bot. 2009, 60, 4411–4421. [Google Scholar] [CrossRef]
- Ginzberg, I.; Minz, D.; Faingold, I.; Soriano, S.; Mints, M.; Fogelman, E.; Warshavsky, S.; Zig, U.; Yermiyahu, U. Calcium mitigated potato skin physiological disorder. Am. J. Potato Res. 2012, 89, 351–362. [Google Scholar] [CrossRef]
- Keren-Keiserman, A.; Baghel, R.S.; Fogelman, E.; Faingold, I.; Zig, U.; Yermiyahu, U.; Ginzberg, I. Effects of polyhalite fertilization on skin quality of potato tuber. Front. Plant Sci. 2019, 10, 1379. [Google Scholar] [CrossRef]
- Okazawa, Y.; Iriuda, N. On occurrence of defected potato tubers with rough russeted skin. Jpn. J. Crop Sci. 1980, 49, 58–65. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Timm, H.; Spurr, A.R. Effects of soil temperature on growth and nutrition of potato plants and tuberization, composition, and periderm structure of tubers. Proc. Am. Soc. Hortic. Sci. 1964, 84, 412–423. [Google Scholar]
- Vulavala, V.K.R.; Fogelman, E.; Faigenboim, A.; Shoseyov, O.; Ginzberg, I. The transcriptome of potato tuber phellogen reveals cellular functions of cork cambium and genes involved in periderm formation and maturation. Sci. Rep. 2019, 9, 10216. [Google Scholar] [CrossRef] [PubMed]
- Murphy, H.; Goven, M. Influence of source of potash on yield, specific gravity, and surface russeting of the Russet Burbank variety in Maine. Am. Potato J. 1965, 42, 192–194. [Google Scholar] [CrossRef]
- Inácio, V.; Barros, P.M.; Costa, A.; Roussado, C.; Gonçalves, E.; Costa, R.; Graça, J.; Oliveira, M.M.; Morais-Cecílio, L. Differential DNA methylation patterns are related to phellogen origin and quality of Quercus suber cork. PLoS ONE 2017, 12, e0169018. [Google Scholar] [CrossRef] [PubMed]
- Inácio, V.; Martins, M.T.; Graça, J.; Morais-Cecílio, L. Cork oak young and traumatic periderms show PCD typical chromatin patterns but different chromatin-modifying genes expression. Front. Plant Sci. 2018, 9, 1194. [Google Scholar] [CrossRef] [PubMed]
- Ramos, M.; Rocheta, M.; Carvalho, L.; Inácio, V.; Graça, J.; Morais-Cecilio, L. Expression of DNA methyltransferases is involved in Quercus suber cork quality. Tree Genet. Genomes 2013, 9, 1481–1492. [Google Scholar] [CrossRef]
- Chinnusamy, V.; Zhu, J.-K. Epigenetic regulation of stress responses in plants. Curr. Opin. Plant Biol. 2009, 12, 133–139. [Google Scholar] [CrossRef]
- Lieberman-Lazarovich, M.; Kaiserli, E.; Bucher, E.; Mladenov, V. Natural and induced epigenetic variation for crop improvement. Curr. Opin. Plant Biol. 2022, 70, 102297. [Google Scholar] [CrossRef]
- Barel, G.; Ginzberg, I. Potato skin proteome is enriched with plant defense components. J. Exp. Bot. 2008, 59, 3347–3357. [Google Scholar] [CrossRef]
- Kooke, R.; Johannes, F.; Wardenaar, R.; Becker, F.; Etcheverry, M.; Colot, V.; Vreugdenhil, D.; Keurentjes, J.J.B. Epigenetic basis of morphological variation and phenotypic plasticity in Arabidopsis thaliana. Plant Cell 2015, 27, 337–348. [Google Scholar] [CrossRef]
- Secco, D.; Whelan, J.; Rouached, H.; Lister, R. Nutrient stress-induced chromatin changes in plants. Curr. Opin. Plant Biol. 2017, 39, 1–7. [Google Scholar] [CrossRef]
- Law, J.A.; Jacobsen, S.E. Establishing, maintaining and modifying DNA methylation patterns in plants and animals. Nat. Rev. Genet. 2010, 11, 204–220. [Google Scholar] [CrossRef] [PubMed]
- Goll, M.G.; Bestor, T.H. Eukaryotic cytosine methyltransferases. Annu. Rev. Biochem. 2005, 74, 481–514. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Zhong, X.; Bernatavichute, Y.V.; Stroud, H.; Feng, S.; Caro, E.; Vashisht, A.A.; Terragni, J.; Chin, H.G.; Tu, A. Dual binding of chromomethylase domains to H3K9me2-containing nucleosomes directs DNA methylation in plants. Cell 2012, 151, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Stroud, H.; Do, T.; Du, J.; Zhong, X.; Feng, S.; Johnson, L.; Patel, D.J.; Jacobsen, S.E. Non-CG methylation patterns shape the epigenetic landscape in Arabidopsis. Nat. Struct. Mol. Biol. 2014, 21, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Zemach, A.; Kim, M.Y.; Hsieh, P.-H.; Coleman-Derr, D.; Eshed-Williams, L.; Thao, K.; Harmer, S.L.; Zilberman, D. The Arabidopsis nucleosome remodeler DDM1 allows DNA methyltransferases to access H1-containing heterochromatin. Cell 2013, 153, 193–205. [Google Scholar] [CrossRef]
- Yaari, R.; Katz, A.; Domb, K.; Harris, K.D.; Zemach, A.; Ohad, N. RdDM-independent de novo and heterochromatin DNA methylation by plant CMT and DNMT3 orthologs. Nat. Commun. 2019, 10, 1613. [Google Scholar] [CrossRef]
- Zhang, H.; Lang, Z.; Zhu, J.-K. Dynamics and function of DNA methylation in plants. Nat. Rev. Mol. Cell Biol. 2018, 19, 489–506. [Google Scholar] [CrossRef]
- Zhu, J.-K. Active DNA Demethylation Mediated by DNA Glycosylases. Annu. Rev. Genet. 2009, 43, 143–166. [Google Scholar] [CrossRef]
- Ginzberg, I.; Gerchikov, N.; Ziv, E.; Fogelman, E.; Tanami, Z.; Warschavsky, S. Potato tuber skin development: The effect of hot climate and plant desiccation. Acta Hortic. 2005, 684, 93–98. [Google Scholar] [CrossRef]
- Vulavala, V.K.; Elbaum, R.; Yermiyahu, U.; Fogelman, E.; Kumar, A.; Ginzberg, I. Silicon fertilization of potato: Expression of putative transporters and tuber skin quality. Planta 2016, 243, 217–229. [Google Scholar] [CrossRef]
- Fogelman, E.; Tanami, S.; Ginzberg, I. Anthocyanin synthesis in native and wound periderms of potato. Physiol. Plant. 2015, 153, 616–626. [Google Scholar] [CrossRef] [PubMed]
- Vulavala, V.K.R.; Fogelman, E.; Rozental, L.; Faigenboim, A.; Tanami, Z.; Shoseyov, O.; Ginzberg, I. Identification of genes related to skin development in potato. Plant Mol. Biol. 2017, 94, 481–494. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Sun, L.; Huang, R.-Z.; Liang, W.; Liu, X.; He, H.; Fukuda, H.; He, X.-Q.; Qian, W. Active DNA demethylation regulates tracheary element differentiation in Arabidopsis. Sci. Adv. 2020, 6, eaaz2963. [Google Scholar] [CrossRef] [PubMed]
- Ai, Y.; Jing, S.; Cheng, Z.; Song, B.; Xie, C.; Liu, J.; Zhou, J. DNA methylation affects photoperiodic tuberization in potato (Solanum tuberosum L.) by mediating the expression of genes related to the photoperiod and GA pathways. Hortic. Res. 2021, 8, 181. [Google Scholar] [CrossRef]
- Dutta, M.; Raturi, V.; Gahlaut, V.; Kumar, A.; Sharma, P.; Verma, V.; Gupta, V.K.; Sood, S.; Zinta, G. The interplay of DNA methyltransferases and demethylases with tuberization genes in potato (Solanum tuberosum L.) genotypes under high temperature. Front. Plant Sci. 2022, 13, 933740. [Google Scholar] [CrossRef]
- Sabbah, S.; Raise, M.; Tal, M. Methylation of DNA in NaCl-adapted cells of potato. Plant Cell Rep. 1995, 14, 467–470. [Google Scholar] [CrossRef]
- Ruzin, S.E. Plant Microtechnique and Microscopy; Oxford University Press: New York, NY, USA, 1999. [Google Scholar]
- Johansen, D.A. Plant Microtechniques; McGraw-Hill Book Company, Inc.: New York, NY, USA, 1940. [Google Scholar]
Figure 1.
Images of potatoes used in the study. (a) The red cultivar Dèsirèe at immature stage. Rosanna tubers are not shown here but look similar. (b,c) Tubers of the white cultivars Jazzy and Georgina, respectively. On the left of each frame are tubers with the characteristic shiny and smooth skin; on the right are tubers with environmentally triggered russeting. (d) Three lines from a breeding population segregating for russeting; left to right (in pairs) are tubers with smooth-skin, weak russeting, and heavy russeting. Enlarged skin images are given below the respective tubers.
Figure 1.
Images of potatoes used in the study. (a) The red cultivar Dèsirèe at immature stage. Rosanna tubers are not shown here but look similar. (b,c) Tubers of the white cultivars Jazzy and Georgina, respectively. On the left of each frame are tubers with the characteristic shiny and smooth skin; on the right are tubers with environmentally triggered russeting. (d) Three lines from a breeding population segregating for russeting; left to right (in pairs) are tubers with smooth-skin, weak russeting, and heavy russeting. Enlarged skin images are given below the respective tubers.
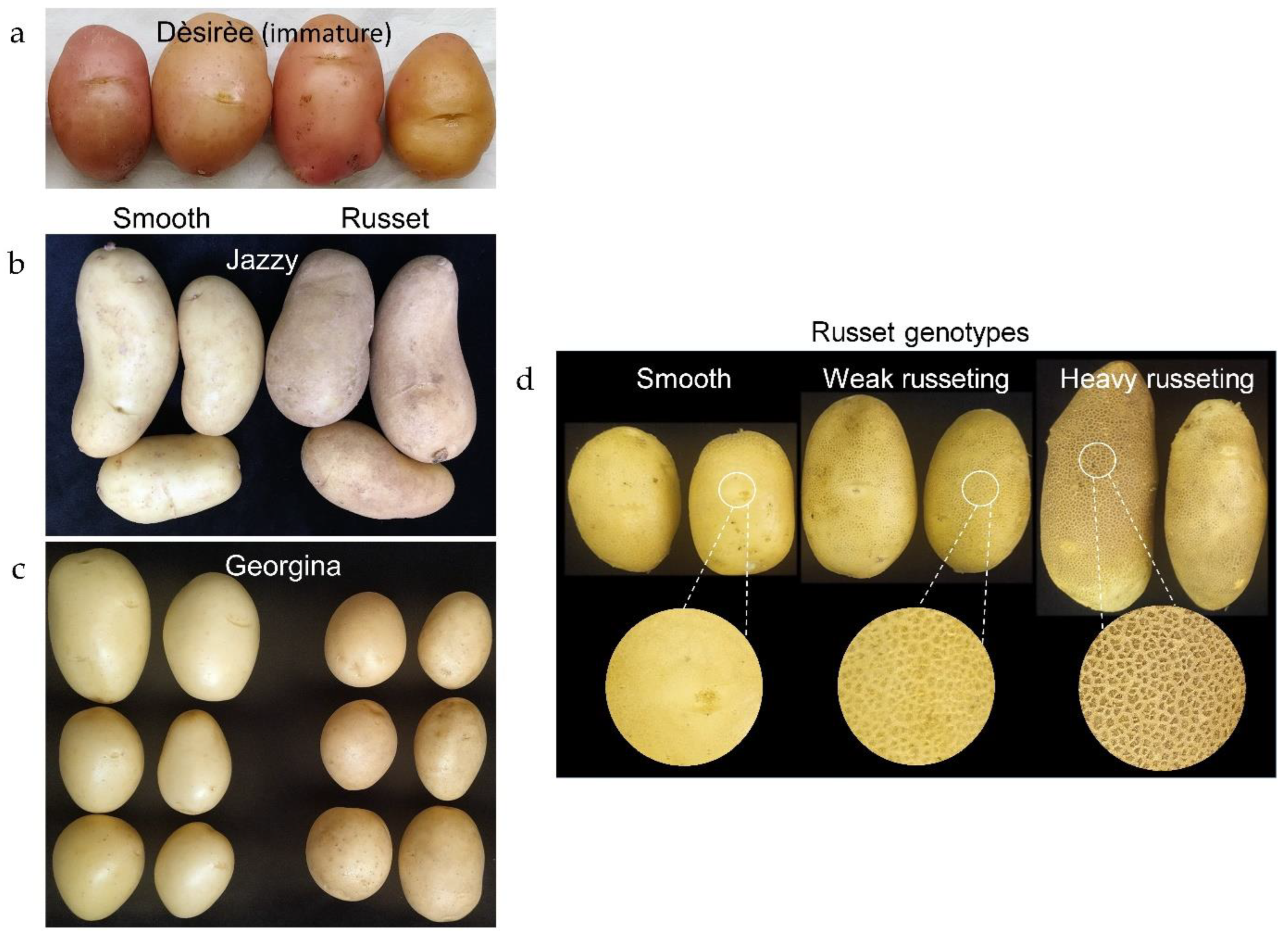
Figure 2.
Environmentally triggered russeting. Anatomy of smooth and russet skin of potato cultivars. (a) Smooth skin of cv. Rosanna, (b) smooth skin of cv. Jazzy, (c) russet skin of cv. Jazzy with newly formed phellem layers (encircled) below it. The left and right panels are images from light and UV microscopes, respectively. The red bar marks the phellem layers (the skin). Scale bar = 100 µm.
Figure 2.
Environmentally triggered russeting. Anatomy of smooth and russet skin of potato cultivars. (a) Smooth skin of cv. Rosanna, (b) smooth skin of cv. Jazzy, (c) russet skin of cv. Jazzy with newly formed phellem layers (encircled) below it. The left and right panels are images from light and UV microscopes, respectively. The red bar marks the phellem layers (the skin). Scale bar = 100 µm.

Figure 3.
Genetically determined russeting. Anatomy of smooth and russet skin of potato breeding lines. Lines with smooth (a), weak russeted (b), and heavy russeted skin (c). The left and right panels are images from light and UV microscopes, respectively. The red bar marks the phellem layers. The orange bars mark the russet layers. Scale bar = 100 µm.
Figure 3.
Genetically determined russeting. Anatomy of smooth and russet skin of potato breeding lines. Lines with smooth (a), weak russeted (b), and heavy russeted skin (c). The left and right panels are images from light and UV microscopes, respectively. The red bar marks the phellem layers. The orange bars mark the russet layers. Scale bar = 100 µm.
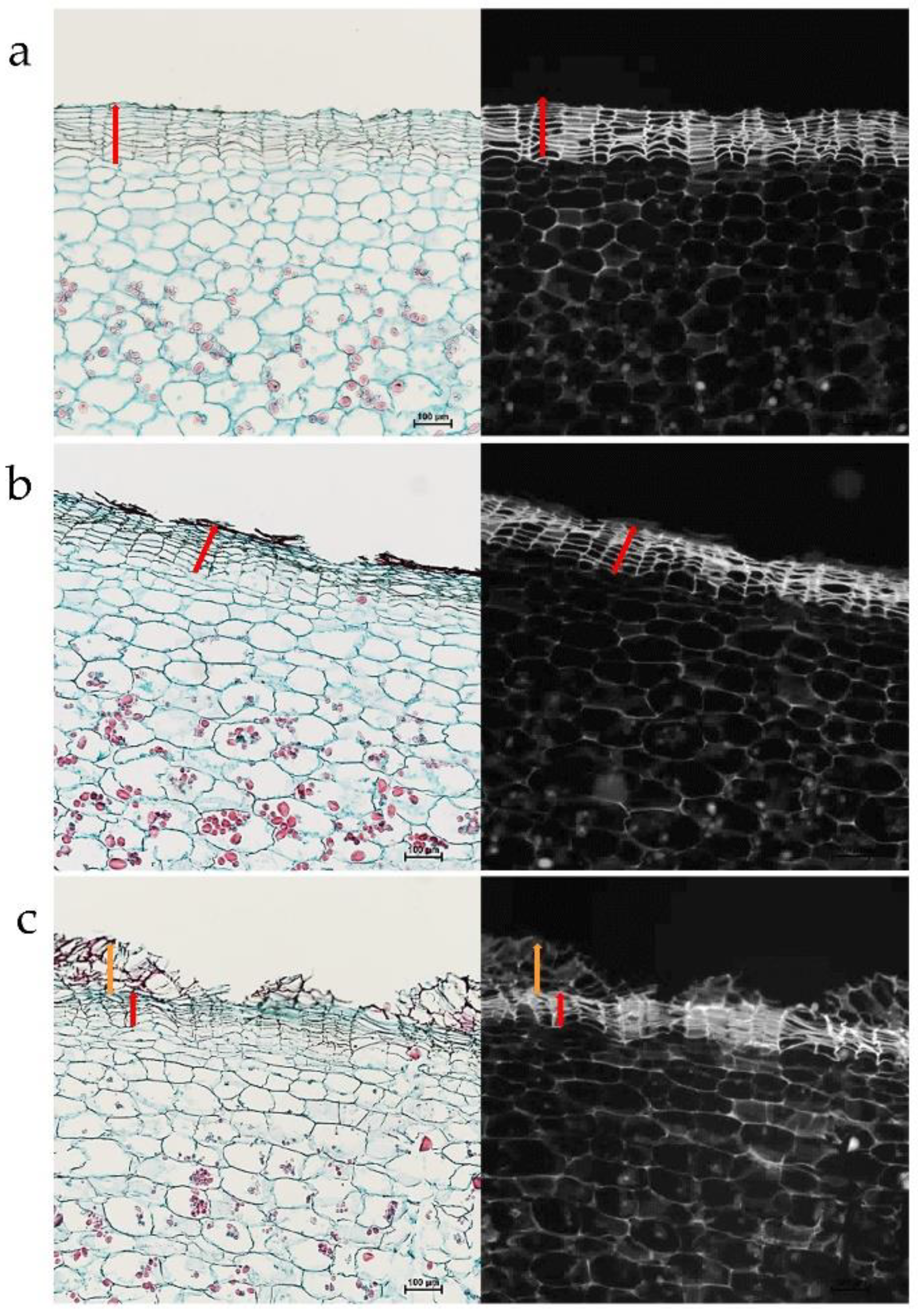
Figure 4.
Global DNA methylation level in tuber skin, (a) at two developmental stages, (b) following environmentally triggered russeting, and (c) in the skin of russet and smooth breeding lines. (a) Samples were collected at immature and mature developmental stages from tubers of Dèsirèe and Rosanna cultivars for two successive years. (b) Smooth and russet regions of the skin were collected from tubers of two potato cultivars, Jazzy and Georgina. (c) The skin was collected from breeding lines with smooth skin (16_S), weak russeting (43_wR), and heavy russeting (25_hR). Each bar represents the averages of three biological replicates with SE. Asterisks indicate statistical significance between two means of the same sample. Capital letters compare the immature skin of the breeding lines, and small letters are for mature skin. Statistical analysis was done by Tukey-Kramer HSD (p < 0.05).
Figure 4.
Global DNA methylation level in tuber skin, (a) at two developmental stages, (b) following environmentally triggered russeting, and (c) in the skin of russet and smooth breeding lines. (a) Samples were collected at immature and mature developmental stages from tubers of Dèsirèe and Rosanna cultivars for two successive years. (b) Smooth and russet regions of the skin were collected from tubers of two potato cultivars, Jazzy and Georgina. (c) The skin was collected from breeding lines with smooth skin (16_S), weak russeting (43_wR), and heavy russeting (25_hR). Each bar represents the averages of three biological replicates with SE. Asterisks indicate statistical significance between two means of the same sample. Capital letters compare the immature skin of the breeding lines, and small letters are for mature skin. Statistical analysis was done by Tukey-Kramer HSD (p < 0.05).
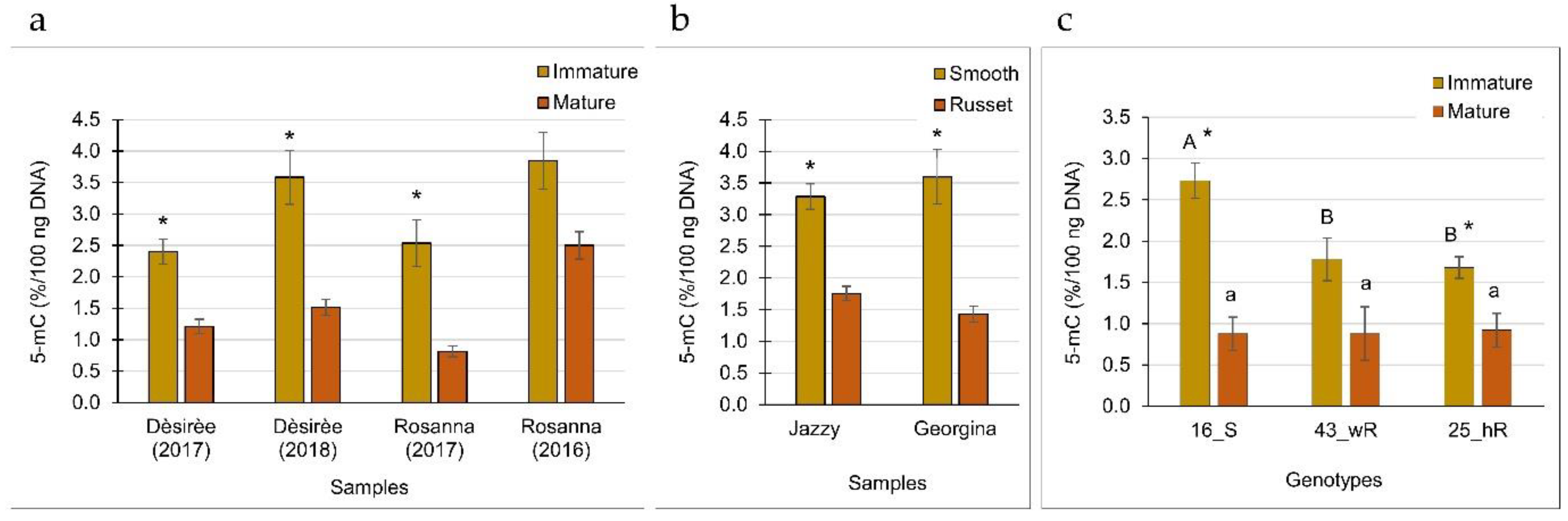
Figure 5.
The expression level of genes coding for epigenetic modifiers that may determine skin maturation and russeting (listed in Table 1). (a,b) Immature and mature skin of the cultivar Dèsirèe collected in 2017 and 2018, respectively. (c,d) Smooth (S) and russet (R) skin of the cultivars Jazzy and Georgina, respectively. Each bar represents the averages of three biological replicates with SE. Asterisks indicate statistical significance between two means of immature vs. mature or smooth vs. russet. Statistical analysis was done by Tukey-Kramer HSD (p < 0.05).
Figure 5.
The expression level of genes coding for epigenetic modifiers that may determine skin maturation and russeting (listed in Table 1). (a,b) Immature and mature skin of the cultivar Dèsirèe collected in 2017 and 2018, respectively. (c,d) Smooth (S) and russet (R) skin of the cultivars Jazzy and Georgina, respectively. Each bar represents the averages of three biological replicates with SE. Asterisks indicate statistical significance between two means of immature vs. mature or smooth vs. russet. Statistical analysis was done by Tukey-Kramer HSD (p < 0.05).

Figure 6.
The expression level of genes coding for epigenetic modifiers that may determine skin maturation and russeting in immature and mature skin of smooth and russeted genotypes. (a) The smooth genotype 16_S; (b) the heavy russeted genotype 25_hR; and (c) a comparison of the immature state of the three genotypes, the smooth 16_S, the weak russeted 43_wR, and the heavy russeted 25_hR. Each bar represents the averages of three biological replicates with SE. Significant difference among means is indicated by an asterisk or different letters. Statistical analysis was done by Tukey-Kramer HSD (p < 0.05).
Figure 6.
The expression level of genes coding for epigenetic modifiers that may determine skin maturation and russeting in immature and mature skin of smooth and russeted genotypes. (a) The smooth genotype 16_S; (b) the heavy russeted genotype 25_hR; and (c) a comparison of the immature state of the three genotypes, the smooth 16_S, the weak russeted 43_wR, and the heavy russeted 25_hR. Each bar represents the averages of three biological replicates with SE. Significant difference among means is indicated by an asterisk or different letters. Statistical analysis was done by Tukey-Kramer HSD (p < 0.05).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kumar, P.; Kaplan, Y.; Endelman, J.B.; Ginzberg, I. Epigenetic Modifications Related to Potato Skin Russeting. Plants 2023, 12, 2057. https://doi.org/10.3390/plants12102057
AMA Style
Kumar P, Kaplan Y, Endelman JB, Ginzberg I. Epigenetic Modifications Related to Potato Skin Russeting. Plants. 2023; 12(10):2057. https://doi.org/10.3390/plants12102057
Chicago/Turabian StyleKumar, Pawan, Yulia Kaplan, Jeffrey B. Endelman, and Idit Ginzberg. 2023. "Epigenetic Modifications Related to Potato Skin Russeting" Plants 12, no. 10: 2057. https://doi.org/10.3390/plants12102057
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.