
Functional Divergence Analysis of AGL6 Genes in Prunus mume

Shandong Provincial Research Center of Demonstration Engineering Technology for Urban and Rural Landscape, College of Forestry, Shandong Agricultural University, Taian 271018, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Plants 2023, 12(1), 158; https://doi.org/10.3390/plants12010158
Submission received: 23 September 2022
/
Revised: 30 November 2022
/
Accepted: 6 December 2022
/
Published: 29 December 2022
(This article belongs to the Special Issue Advancement of Ornamental Flowering Plants)
Abstract
:The AGAMOUS-LIKE6 (AGL6) lineage is an important clade of MADS-box transcription factors that play essential roles in floral organ development. The genome of Prunus mume contains two homoeologous AGL6 genes that are replicated as gene fragments. In this study, two AGL6 homologs, PmAGL6-1 and PmAGL6-2, were cloned from P. mume and then functionally characterized. Sequence alignment and phylogenetic analyses grouped both genes into the AGL6 lineage. The expression patterns and protein–protein interaction patterns showed significant differences between the two genes. However, the ectopic expression of the two genes in Arabidopsis thaliana resulted in similar phenotypes, including the promotion of flowering, alteration of floral organ structure, participation in the formation of the floral meristem and promotion of pod bending. Therefore, gene duplication has led to some functional divergence of PmAGL6-1 and PmAGL6-2 but their functions are similar. We thus speculated that AGL6 genes play a crucial role in flower development in P. mume.
1. Introduction
Prunus mume, the flowers of which occupy the top position among the ten most famous flowers in China, has a planting history of more than 3000 years. The types of its flowers have changed considerably due to its long cultivation. On the basis of the study of flower development in model plants, a series of molecular regulation models for floral organ development have previously been proposed. However, the molecular mechanism regulating flower organ development in P. mume is largely unknown.
According to the ABCDE model, the genes that are responsible for the identity of the five floral organs are divided into five classes: A, B, C, D and E (A for the sepals; A, B and E for the petals; B, C and E for the stamens; C and E for the carpels; and C, D and E for the ovules) [1]. In Arabidopsis, the A-class genes are APETALA1 (AP1) and APETALA2 (AP2), the B-class genes are APETALA3 (AP3) and PISTILLATA (PI), the C-class gene is AGAMOUS (AG), the D-class genes are SHATTERPROOF 1 and 2 (SHP1,2) and SEEDSTICK (STK) and the E-class genes are SEP1, SEP2, SEP3 and SEP4. According to the floral quartet model, the ABCDE class genes regulate the development of different floral organs by forming floral organ-specific tetrameric complexes [2]. In addition to the A, B, C, D and E genes, other MADS-box genes, such as AGL6, play important regulatory roles in plant flower development.
In contrast to the A, B, C, D and E genes, which have been widely studied, there are still few comprehensive studies on the function of the AGL6 genes. AGL6 genes have different expression patterns in different types of plants. In gymnosperms, AGL6 genes are expressed in both vegetative and reproductive organs and are highly expressed in male and female structures. In Picea abies, DAL1 expression is readily detected in male and female cones and in vegetative shoots [3]. In Cryptomeria japonica, CjMADS14 is expressed mainly in the strobili [4]. In dicotyledonous plants, the AGL6 genes are mainly expressed in floral organs, and some are also expressed in vegetative organs. There are two AGL6 genes, AGL6 and AGL13, in the classical model plant Arabidopsis. AGL6 is expressed in ovules, while AGL13 is specifically expressed in pollen and ovules [5,6]. MdMADS11 mRNA expression is high in sepals, petals and ovaries with receptacles [7]. In Actinidia chinensis, two AGL6 homologues have obtained different expression patterns: AcAGL6a is expressed in flower parts and all vegetative tissue samples but mainly in vegetative branches, while AcAGL6b is expressed in vegetative and floral tissues (sepals and petals) but mainly in flower tissues [8]. In monocots, the AGL6 gene is mainly expressed in floral organs. PaAGL6-1 and PaAGL6-2 are expressed throughout all floral organs in Phalaenopsis aphrodite, and PaAGL6-1 is highly expressed specifically in the lip [9]. In Alpinia hainanensis, AhAGL6-like is expressed in all six mature floral organs [10]. Although the expression patterns of AGL6 genes in angiosperms and gymnosperms are slightly different, they are both expressed in floral organs, indicating that AGL6 genes are involved in the development of floral organs. Although AGL6 genes are expressed in the flowers of different plants, there are obvious differences in their expression patterns. PhAGL6 from P. hybrida is present at high levels in developing petals and ovaries, CpAGL6 from Chimonanthus praecox is highly expressed in middle-tepal tissue and HoAGL6 from Hyacinthus orientalis is highly expressed in ovules [11,12,13]. This indicates that, while AGL6 genes may be involved in the regulation of floral organ development in different plants, their functions have undergone species-specific differentiation.
The analysis of AGL6 gene mutants or plants overexpressing the AGL6 gene also proved that the AGL6 gene is involved in floral organ regulation. The lodicules, stamens and carpels were all found to be absent in an OsMADS6 mutant, and many green and lemma-like organs appeared in their place. However, overexpression of OsMADS6 results in the overproduction of palea, lodicules, stamens and carpels [14]. bde mutants exhibit multiple defects in floral development, but these defects do not appear during earlier stages of inflorescence or vegetative development [15]. In Arabidopsis, the flowering time and fertility of agl6-2-deletion mutant plants are normal, which may be due to the redundant function of another AGL6 gene, AGL13. Arabidopsis plants overexpressing AGL6 produce terminal flowers at the top of the inflorescence, and the number and morphology of the floral organs of the terminal flowers are altered [5], suggesting that AGL6 regulates the differentiation of floral organs in Arabidopsis. The ectopic expression of OMADS1, an orchid (Oncidium Gower Ramsey) AGL6-like gene, results in the production of terminal flowers in transgenic Arabidopsis plants [16].
In addition to regulating the development of floral organs, AGL6 genes also regulate flowering time: AGL6 can shorten the growth period of young plants and promote early flowering. Arabidopsis plants overexpressing the AGL6 gene show an early-flowering phenotype under both long-day and short-day conditions [17]. Arabidopsis plants overexpressing the AGL6 genes of different plant species, such as wintersweet and hyacinth, show shorter vegetative growth times and earlier flowering times [11,13]. The AGL6 gene DAL1 plays an important regulatory role in the juvenile-to-adult phase transition in Norway spruce. Arabidopsis plants transformed with the DAL1 gene show an early-flowering phenotype, and plants with an extremely early-flowering phenotype bud and flower as soon as the cotyledons are unearthed [3]. The above results suggest that promoting the expression of the AGL6 gene is a feasible approach for shortening the juvenile growth period and promoting early flowering of plants, especially woody plants.
Gene duplication plays an important role in the evolution of diversity and novel functions and is especially prevalent in the nuclear genomes of flowering plants [18]. Duplicated genes (paralogs) may experience the following fates: (1) nonfunctionalization through silencing or null mutation, (2) neofunctionalization through the acquisition of a novel function or (3) subfunctionalization through the partitioning of functional modules such that the complementation of the two copies provides the functional capability of the ancestral gene [18]. Extensive duplication events have resulted in the expansion of MADS-box genes, and the study of MADS-box gene evolution provides examples of all three postduplication fates mentioned above [19,20].
According to the existing research, there is more than one AGL6 gene within individual species. There is only one AGL6 gene in Brachypodium distachyon, ginger and petunia [12,21,22,23,24], while two AGL6-type genes are found in Arabidopsis, Magnolia wufengensis and rice [6,17,25,26,27] and three AGL6-class genes are present in wheat and Cymbidium goeringii [28,29]. In Rosa chinensis, the number of AGL6 genes reaches five [30]. The current research on AGL6 genes is not sufficient, and more AGL6 genes remain to be found.
The two AGL6 genes of P. mume are replicated as gene fragments and, therefore, provide a good model for studying the functional divergence and evolution of duplicated genes in plants. AGL6 genes do not belong to any of the ABDCE gene categories, but they play important roles in flower development and floral organ determination in plants. In this study, we cloned two AGL6 homologs, PmAGL6-1 and PmAGL6-2, from P. mume. We studied their phylogeny, expression patterns and protein–protein interaction patterns and analyzed their functions. On this basis, the model of the molecular regulation of P. mume organ development was improved to provide a theoretical basis for using the AGL6 gene to improve the flower types of P. mume.
2. Results
2.1. Cloning and Sequence Analysis of PmAGL6-1 and PmAGL6-2
The full-length CDSs of PmAGL6-1 and PmAGL6-2 were cloned and then confirmed by sequencing (Figure 1). PmAGL6-1 contained a 738 bp open reading frame encoding 245 amino acids, whereas PmAGL6-2 contained a 720 bp open reading frame encoding 239 amino acids. The two genes also included a highly conserved MADS domain and a less conserved K domain (Figure 1). In addition, AGL6 motifs I and II [26] were identified in the C-terminal domains of PmAGL6-1 and PmAGL6-2. The phylogenetic analysis of AGL6 proteins showed that PmAGL6-1 and PmAGL6-2 belonged to two separate subclades (Figure 2).
2.2. Expression Pattern Analysis of PmAGL6-1 and PmAGL6-2
The expression patterns of PmAGL6-1 and PmAGL6-2 were analyzed by qRT-PCR (Table 1). The transcripts of PmAGL6-1 and PmAGL6-2 were not detected in the vegetative organs (roots, stems and leaves), and the expression domains of the two genes were limited to reproductive organs. The tissue-specific expression of these two genes in floral tissues has diverged. The PmAGL6-1 gene is expressed in sepals, petals, stamens and pistils, but its expression levels are very different. The expression levels are the highest in sepals and petal and low in pistils and stamens. The PmAGL6-2 gene is expressed at the highest level in sepals and at a low level in pistils. PmAGL6-1 and PmAGL6-2 were all highly expressed in the three fruit stages and the expression level decreased gradually with the development of fruit (Figure 3a).
During the differentiation of flower buds, the expression levels of PmAGL6-1 and PmAGL6-2 first increased until they reached a peak and then decreased. PmAGL6-1 reached its highest expression level in S6, while PmAGL6-2 was expressed at the highest level in S5 (Figure 3b).
2.3. Ectopic Expression of PmAGL6-1 and PmAGL6-2 in Arabidopsis
The seeds of the T2-generation transgenic plants of different lines were screened and sown with wild-type Arabidopsis at the same time. The phenotypes of T2-generation transgenic plants obtained under the same environmental conditions were observed. The ectopic expression of PmAGL6-1 and PmAGL6-2 caused early flowering (Figure 4IA,IIA and Table 2). Extremely early flowering was observed in PmAGL6-1 transgenic plants, which showed curling leaves and shorter plant heights (Figure 4IB) than wild-type Arabidopsis plants. In addition to the changes in flowering time, petal numbers and flower architecture were also changed in the transgenic plants. The number of petals in the transgenic plants varied from three to five (Figure 4ID,IE,IIB,IIC). The overexpression of PmAGL6-1 and PmAGL6-2 resulted in bilaterally symmetrical petals in transgenic Arabidopsis plants (Figure 4IF,IID) and changed the arrangement of the petals, resulting in the partial overlap of petals (Figure 4IJ,IIH). In addition, in PmAGL6-2 transgenic plants, the petal tip became pointed (Figure 4IIJ). Terminal flowers were observed in PmAGL6-1 and PmAGL6-2 transgenic plants in which two to four flowers were combined (Figure 4IG–II,IIE–IIG). Both types of transgenic plants exhibited curved fruit pods (Figure 4IK,III).
We analyzed the expression profiles of PmAGL6-1 in four 35S::PmAGL6-1 transgenic Arabidopsis lines and PmAGL6-2 in five 35S::PmAGL6-2 transgenic Arabidopsis lines by qRT-PCR. The results showed that PmAGL6-1 and PmAGL6-2 were expressed in the transgenic plants, but their expression levels were different in different lines (Figure 5a,b).
CYC genes play an important role in flower symmetry [31,32]. To further understand the reasons why the flowers became bilaterally symmetrical in the PmAGL6-1 and PmAGL6-2 transgenic Arabidopsis plants, we collected bilaterally symmetrical flowers from 35S::PmAGL6-1 and 35S::PmAGL6-1 transgenic Arabidopsis plants. Then, the expression of the CYC gene was analyzed. In bilaterally symmetrical flowers from the 35S::PmAGL6-1 and 35S::PmAGL6-2 transgenic Arabidopsis plants, CYC was upregulated (Figure 5c).
To further understand the mechanisms responsible for the early flowering of the PmAGL6-1 and PmAGL6-2 transgenic Arabidopsis lines, the expressions of some of the endogenous flowering-related genes were analyzed in the 35S::PmAGL6-1 transformant A1 line and the 35S::PmAGL6-2 transformant B4 line, which exhibited an early-flowering phenotype. Relative to the wild-type, AP1, CO, FLC, FT, SOC and LFY expression was upregulated in the two early-flowering lines, whereas the expression of FLC was downregulated (Figure 5d).
2.4. Protein–Protein Interactions among the Products of Two AGL6 Genes and Other Floral Organ Identity-Determining Genes in P. mume
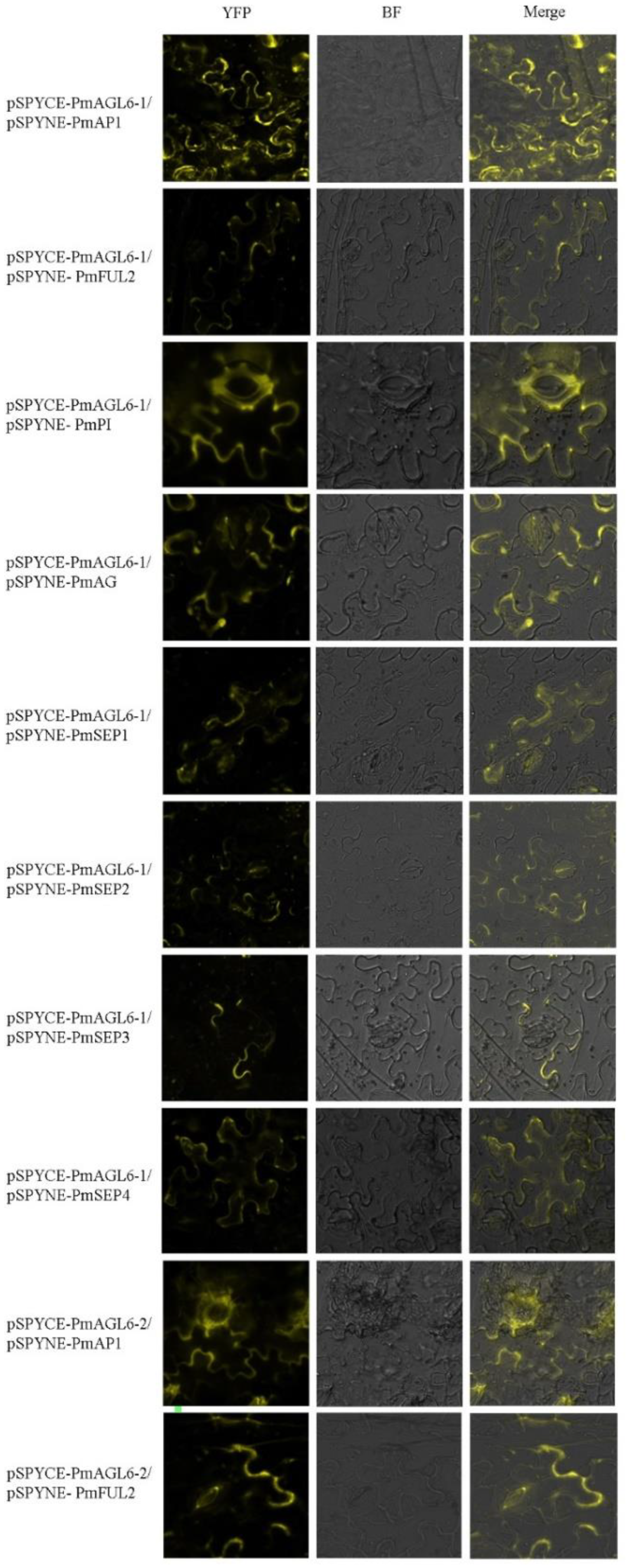
We used yeast two-hybrid assays and bimolecular fluorescence complementation experiments to study protein–protein interactions among the products of two AGL6 genes and other floral organ identity-determining genes in P. mume. The Y2H experiment revealed that neither PmAGL6-1 nor PmAGL6-2 could form a homodimer and that they could not interact with each other. There were differences in the types and numbers of interacting proteins between the products of the two AGL6 proteins. Nine proteins interacted with PmAGL6-1 among the A, B, C and E gene products, while only two class A genes showed interactions with PmAGL6-2. PmAGL6-1 could strongly dimerize with PmAP1 and moderately interacted with PmFUL2, PmPI, PmAG, PmSEP1, PmSEP2 and PmSEP4. PmAGL6-1 could only interact weakly with PmFUL1 and PmSEP3. PmAGL6-2 also showed strong interactions with PmAP1 but weaker interactions with PmFUL2 (Figure 6).
To further verify the results of the yeast two-hybrid assays, we carried out fluorescence complementation experiments. The results obtained via the two experimental methods were basically the same. Yellow fluorescent protein (YFP) signals were detected in the tobacco epidermis cell membrane when the tested combination showed interaction in the Y2H analysis, except for YCE-PmAGL6-1/YNE-PmFUL1, possibly because the interaction between PmAGL6-1 and PmFUL1 was too weak or nonexistent (Figure 7).
3. Discussion
In this study, two AGL6 genes, PmAGL6-1 and PmAGL6-2, from P. mume were cloned and analyzed. The open reading frame of PmAGL6-2 was 18 bases shorter than that of PmAGL6-1, and the missing bases were located mainly in the C region. PmAGL6-1 shared 51.02% homology with PmAGL6-2. The amino acid sequences of the K region, AGL6-I motif and AGL6-II motif of the two genes were very different. Phylogenetic tree analysis also showed that the two genes were not clustered together. This suggested that the sequences of the two genes diverged after the duplication events.
PmAGL6-1 and PmAGL6-2 were both highly expressed in fruit, indicating that the two AGL6 genes may be involved in the development of fruits in P. mume. Neither PmAGL6-1 nor PmAGL6-2 was expressed in vegetative tissues, and their expressions patterns in floral organs were also different. PmAGL6-1 was expressed in sepals, petals, stamens and pistils, and it was highly expressed in sepals and petals. PmAGL6-2 was only expressed in sepals and carpels and was highly expressed in sepals. This expression pattern in floral organs is similar to PmAP1 in P. mume [33]. The expression patterns of PmAGL6-1 and PmAGL6-2 in different flower bud differentiation stages were the same, first increasing and then decreasing after they reached a maximum. This expression pattern is consistent with those of the AGL6 genes of wheat and wintersweet [13,28] and also similar to the expression pattern of PmAP1 in P. mume [33]. It can be seen that PmAGL6-1 and PmAGL6-2 are involved in the development of floral organs in P. mume, and their expression patterns are similar to the expression pattern of PmAP1.
PmAGL6-1 can interact with the class A, B, C and E genes in P. mume, while PmAGL6-2 only interacts with class A genes. In wheat, three TaAGL6 proteins interact with TaAP3, TaAG and TaMADS13, and redundant functions for these proteins have been suggested [28]. Both MawuAGL6-1 and MawuAGL6-2 show strong interactions with MawuAGL2, MawuAGL9 and MawuAG-1 and can form homodimers and interact with each other; one difference is that MawuAGL6-2 can interact with MawuAP1 [25], and their protein–protein interaction patterns differ slightly. In contrast, the protein–protein interaction patterns of PmAGL6-1 and PmAGL6-2 are very different. Studies have shown that the K region of the MADS-box gene plays a significant role in protein–protein interactions [34,35]. The amino acid sequences of the K regions of the three AGL6 genes in wheat are identical, which may cause them to interact with the same proteins. There are 14 amino acid differences in the K region of the amino acid sequence encoded by MawuAGL6-1 and MawuAGL6-2, and 28 amino acid differences in the K region of the amino acid sequence encoded by PmAGL6-1 and PmAGL6-2; this may be the fundamental reason why the difference in the protein–protein interaction patterns between PmAGL6-1 and PmAGL6-2 was greater than that between MawuAGL6-1 and MawuAGL6-2. The similarity in the protein–protein interaction patterns between PmAGL6-1 and PmAGL6-2 was that they could interact with PmAP1 and PmFUL2. The protein–protein interaction patterns of PmAGL6-1 and PmAP1 were very similar, further implying the similarity of their functions [33].
The overexpression of PmAGL6-1 and PmAGL6-2 resulted in early-flowering phenotypes in Arabidopsis. Moreover, a very early-flowering phenotype was found in PmAGL6-1 transgenic plants, which was very similar to the very early-flowering phenotype caused by the ectopic expression of OMADS7, OMADS1 and DAL in Arabidopsis in terms of plant appearance [3,36]. OMADS1 significantly promotes flowering by positively activating the flowering time genes FT and SOC1 and the floral initiation genes LFY and AP1 in transgenic Arabidopsis plants [16]. Upregulated expression of HoAGL6 causes early flowering by increasing the expression of SOC1 and LFY [11]. The ectopic expression of CpAGL6 in Arabidopsis leads to precocious flowering, which is mainly correlated with the inhibition of the floral repressor FLC and the promotion of the floral promoters AP1 and FT [13]. Endogenous overexpression of AGL6 increases the expression of the known floral regulators FT and AP1 in Arabidopsis [5]. FT, SOC and CO are flowering-time genes that are close to the top of the regulatory hierarchy in flower development and mediate the switch from vegetative to reproductive development by activating meristem identity genes or by repressing genes that maintain the vegetative phase. AP1 and LFY are meristem identity genes that control the transition from vegetative growth to inflorescence and floral meristem development and function as upstream regulators of floral organ identity genes [37,38,39,40]. FLC is an inhibitor of flowering in Arabidopsis [41]. The results of the detection of flowering gene expression in transgenic plants showed that AP1, CO, FT, SOC and LFY were upregulated in both PmAGL6-1 and PmAGL6-2 transgenic plants, while FLC was downregulated. These genes whose expression levels were measured are the key genes in the flower induction pathway: they determine a relationship between regulation and being regulated. However, based on the current research, it is impossible to determine the mechanism used by the two PmAGL6 genes to promote flowering. We speculate that PmAGL6-1 and PmAGL6-2 may promote the expression of the flowering-time genes CO, FT and SOC; reduce the expression of FLC; promote the expression of the meristem identity genes AP1 and LFY; and then promote flowering. As for which genes PmAGL6-1 and PmAGL6-2 directly act on to cause early flowering, further research is needed.
CYC was upregulated in the bilaterally symmetrical flowers of 35S::PmAGL6-1 and 35S::PmAGL6-2 transgenic Arabidopsis plants. This shows that the ectopic expression of PmAGL6-1 and PmAGL6-2 promotes the expression of CYC in transgenic plants, which in turn affects the symmetry of flowers. In transgenic Arabidopsis thaliana, the ectopic expression of PmAGL6-1 and PmAGL6-2 changes the number and arrangement of Arabidopsis petals. In addition, some petals of PmAGL6-2 transgenic plants were sharp at the apex. In Arabidopsis flowers, the initiation of the four petals depends on the size of the floral meristem, the establishment of boundaries that demarcate the position of petal primordia on the floral meristem, the transient development of auxin activity maxima at these positions and the general mechanisms of lateral organ outgrowth [38,42,43]. The ectopic expression of PmAGL6-1 and PmAGL6-2 may affect the expression of genes related to petal development and cause the petals of transgenic plants to become abnormal. The ectopic expression of PmAGL6-2 makes Arabidopsis petals change in number, arrangement and shape, but PmAGL6-2 is not expressed in Prunus mume petals, indicating that PmAGL6-2 may not participate in the development regulation of petals, but its transfer into Arabidopsis will cause petal changes. ZAG3 is expressed in floral meristems. OMADS1 is also expressed in floral meristems, and ectopic expression makes transgenic Arabidopsis plants produce terminal flowers [15,16]. Terminal flowers appeared in PmAGL6-1 and PmAGL6-2 transgenic Arabidopsis, indicating that PmAGL6-1 and PmAGL6-2 may be involved in the regulation of floral meristems. Both PmAGL6-1 and PmAGL6-2 can make Arabidopsis fruit bend, and the two genes are highly expressed in fruit, indicating that PmAGL6-1 and PmAGL6-2 may participate in the development of fruit in P. mume.
The roles of AGL6 genes in Oncidium Gower Ramsey and Nymphaea tetragona are considered to be similar to that of AP1 [16,44]. The study of PmAP1 in P. mume showed that PmAP1 is expressed only in sepals; similarly, the two AGL6 genes were also expressed in sepals. The expression pattern of PmAP1 was similar to those of PmAGL6-1 and PmAGL6-2, and both PmAGL6-1 and PmAGL6-2 can interact strongly with PmAP1. PmAP1 promoted early flowering in Arabidopsis and transformed the inflorescence meristem into the floral meristem, which has also been observed in PmAGL6-1 and PmAGL6-2 transgenic Arabidopsis (Xu 2015). From this point of view, the functions of the two PmAGL6 genes and PmAP1 are similar, but the two PmAGL6 genes have their own unique functions.
In many plants, AGL6 genes have experienced gene duplication events, producing several AGL6 genes, but the functional changes in AGL6 genes are different in different species. Although the two AGL6 genes of P. mume differ greatly in their gene sequences, expression patterns and protein–protein interaction patterns, they exhibit important, similar functions in promoting flowering and floral organ development, especially petal development. In Arabidopsis, AGL6 and AGL13 exhibit different expression patterns and functions, showing subfunctionalization [6,17,45,46,47]. There are two AGL6 genes in Magnolia wufengensis, which arose via gene duplication and underwent diversification of their expression and protein–protein interaction patterns thereafter. Functional analysis revealed that, in addition to showing common functions in accelerating flowering, MawuAGL6-1 might be responsible for flower meristem determinacy, while MawuAGL6-2 is preferentially recruited to regulate tepal morphogenesis [25]. Rice contains two AGL6-like genes: OsMADS17 plays minor and redundant roles, whereas OsMADS6 is a key regulator of flower development [26,27,48]. The expression of MapaAGL6-1 and MapaAGL6-2 differs, and the ectopic expression of MapaAGL6-1 and MapaAGL6-2 in Arabidopsis ap1–10 mutants results in different phenotypes [49]. Three TaAGL6 genes exhibit similar expression patterns and protein–protein interaction patterns and show similar functions in transgenic Arabidopsis [28]. CgAGL6-1, CgAGL6-2 and CgAGL6-3 present different expression patterns and protein–protein interaction patterns [29]. Thus, it can be seen that there is species specificity in the differences in the expression patterns, protein–protein interaction patterns and functions of AGL6 genes, without following any obvious rules. Therefore, further research on the AGL6 gene should be conducted.
P. mume has many floral patterns, but the molecular mechanism of floral organ development in P. mume is still unclear. PmAGL6-1 and PmAGL6-2 play important roles in promoting flowering and floral organ development. Gene duplication events cause differences in the expression patterns and protein–protein interaction patterns of PmAGL6-1 and PmAGL6-2, but their functions are similar. Therefore, we believe that the two AGL6 genes should be included in the molecular model of floral organ development in P. mume. The growth period of P. mume is long, and the flowering period is one of the important factors affecting the horticulture characters of P. mume. Additionally, the AGL6 gene of P. mume can promote flowering. By promoting the expression of AGL6 genes, it may be possible to shorten the growth period of P. mume and advance the flowering time, thus improving the ornamental value of P. mume. In conclusion, our research provides valuable information for the study of flower organ development in other plants and as a supplement to the floral organ development model.
4. Materials and Methods
4.1. Plant Material
All samples were collected from P. mume “Sanlun Yudie” growing under natural conditions in the Jiufeng International Plum Blossom Garden in Beijing, China (40°07′ N, 116°11′ E). Samples were collected, and the stages of flower bud development were determined using the method described by Zhou et al. (2019) [50]. All samples were quickly frozen in liquid nitrogen and stored at −80 °C until RNA extraction.
The Arabidopsis Columbia ecotype was used to produce the transgenic material. After 3 days of vernalization at 4 °C in darkness, the seeds were sown in flower pots and placed in a growth chamber at 21 °C under a 16 h light/8 h dark cycle.
The tobacco (Nicotiana benthamiana) plants used for bimolecular fluorescence complementation (BiFC) were raised in a greenhouse under a light/dark cycle of 16 h/8 h at 25 °C.
4.2. Screening of AGL6 Gene in Prunus mume
The Prunus mume genome data were downloaded from the Prunus mume genome database http://prunusmumegenome.bjfu.edu.cn/ (accessed on 5 December 2022) (Zhang, 2012). Following Xu’s work, two methods, BlastP and HMMER, were used for gene screening (2015).
4.3. Extraction of Total RNA and Synthesis of First-Strand cDNA
Total RNA was extracted from floral buds using an EASYspin Plant RNA Rapid Extraction Kit (Aidlab Biotech, Beijing, China) following the manufacturer’s protocol, and the qualified RNA was preserved at −80 °C. First-strand cDNA was synthesized with a 5× All-In-One RT MasterMix Reverse Transcription Kit (ABM Company, Vancouver, BC, Canada) in accordance with both the manufacturer’s protocol and the requirements of RT-PCR and qRT-PCR.
4.4. Cloning of the Full-Length CDSs of PmAGL6-1 and PmAGL6-2
PmAGL6-1F, PmAGL6-1R, PmAGL6-2F and PmAGL6-2R were designed based on CDSs annotated in the genome database. PCR was performed starting with a 60 s predenaturation step at 94 °C, followed by 30 cycles of 10 s of denaturation at 98 °C, 15 s of annealing at 60 °C and 30 s of extension at 68 °C.
4.5. Sequence Alignment and Phylogenetic Analysis of PmAGL6-1 and PmAGL6-2
ClustalW was used to perform multiple protein sequence alignment of PmAGL6-1, PmAGL6-2 and ten AGL6 genes from other plants. The phylogenetic tree was constructed via the neighbor-joining method using MEGA 5-X software with a bootstrap analysis of 1000 replicates.
4.6. Real-Time Quantitative RT-PCR
We analyzed gene expression via qRT-PCR on a Bio-Rad CFX96™ Real-Time PCR instrument (Bio-Rad, Inc., Hercules, CA, USA). PP2A was used as the reference gene in the analysis of expression patterns of PmAGL6-1 and PmAGL6-2. Actin was used as the reference gene in transgenic Arabidopsis (Table 1). The CYC was extracted from the flower buds of transgenic Arabidopsis, and other genes were extracted from the leaves. The reaction mixture of 20 μL was composed of 1 μL of cDNA, 0.4 μL of each primer, 10 μL of SYBR®Premix Ex Taq™ (TaKaRa, Inc., Tokyo, Japan) and 8.2 μL of ddH2O. The PCR program consisted of an initial step of 95 °C for 30 s, which was followed by 40 cycles of 95 °C for 5 s and 60 °C for 30 s and then a dissociation stage of 95 °C for 10 s, 65 °C for 5 s and 95 °C for 5 s. Each gene was assessed in three biological replicates, and the relative expression levels were calculated using the 2−ΔΔCt method.
4.7. Vector Construction and Arabidopsis Transformation
The full-length CDSs of PmAGL6-1 and PmAGL6-2 were cloned into the pCAMBIA1304 vector under the control of the CaMV35S promoter. These plasmids were then transformed into an Agrobacterium tumefaciens strain for inflorescence infection, which was used to transform Arabidopsis. Mature T0 seeds were collected, disinfected and then sown on 1/2 MS medium that contained 30 mg/mL hygromycin to screen the transformants. The resistant plants grew normally; they were removed and transplanted into small flowerpots and grown at 21 °C under long-day conditions. Screening was repeated according to the above methods until T2 plants were obtained.
4.8. Yeast Two-Hybrid Assay
The Matchmaker Gold Yeast Two-Hybrid System (Clontech, Mountain View, CA, USA) was used for Y2H assays. The full-length CDSs of PmAGL6-1 and PmAGL6-2 were recombined into the pGBKT7 vector to express the bait proteins. The full-length CDSs of PmAGL6-1, PmAGL6-2, PmAP1, PmFUL1, PmFUL2, PmAP3, PmAP3-2, PmPI, PmAG, PmSEP1, PmSEP2, PmSEP3 and PmSEP4 were recombined into pGADT7. These two types of fusion vectors were cotransformed into Y2Hgold cells. The transformed Y2H Gold strain was cultured on SD/-Leu/-Trp and SD/-Trp/-Leu/-Ade/-His media with aureobasidin A (AbA) and X-α-Gal and cultured at 28 °C for three days.
4.9. Bimolecular Fluorescence Complementation Assay
The full-length CDSs of PmAGL6-1 and PmAGL6-2 without termination codons were subcloned into the pSPYCE vector, and the full-length CDSs of PmAGL6-1, PmAGL6-2, PmAP1, PmFUL1, PmFUL2, PmAP3, PmAP3-2, PmPI, PmAG, PmSEP1, PmSEP2, PmSEP3 and PmSEP4 without termination codons were subcloned into the pSPYNE vector.
These fusion vectors were transfected into A. tumefaciens strain GV3101. Agrobacterium cells positive for the two types of fusion vectors were coinfiltrated into N. benthamiana leaves, which was followed by culturing in darkness. After 2–3 days, YFP fluorescence was observed using a confocal laser microscope.
5. Conclusions
The full-length CDS sequences of PmAGL6-1 and PmAGL6-2 were first cloned from the cDNA of P. mume flower buds. PmAGL6-1 and PmAGL6-2 are not expressed in vegetative organs (leaves, roots and stems) but are highly expressed in fruits, and there are differences in expression patterns in floral organs. PmAGL6-1 is expressed in sepals, petals, stamens and pistils and is highly expressed in sepals and petals, while PmAGL6-2 is only expressed in sepals and pistils and is highly expressed in sepals. PmAGL6-1 and PmAGL6-2 increased continuously with the flower bud differentiation, and the expression of PmAGL6-1 reached the highest level at the ovule development stage (S6) and PmAGL6-2 reached the highest level at the pistil initiation (S5) stage, and then the expression of these two genes was downregulated. The expression patterns of PmAGL6-1 and PmAGL6-2 indicated that they may be involved in the development of floral organs and fruits. According to the bioinformatics analysis, expression patterns, protein–protein interaction patterns and phenotypic changes in transgenic Arabidopsis, PmAGL6-1 and PmAGL6-2 play important roles in promoting flowering and flower organ development. In addition, PmAGL6-1 and PmAGL6-2 may also participate in the development of floral meristems and fruits.
Author Contributions
Z.X. and X.H. designed the experiments. L.W. and J.S. conducted the experiments. Q.W., Y.Y. and S.Q. analyzed the data. L.W. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
National Natural Science Foundation of China Sub-Class of Project 31700622.
Data Availability Statement
All data in this study can be found in the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Smaczniak, C.; Immink, R.G.; Angenent, G.C.; Kaufmann, K. Developmental and evolutionary diversity of plant MADS-domain factors: Insights from recent studies. Development 2012, 139, 3081–3098. [Google Scholar] [CrossRef] [Green Version]
- Theißen, G.; Saedler, H. Floral quartets. Nature 2001, 409, 469–471. [Google Scholar] [CrossRef] [PubMed]
- Carlsbecker, A.; Tandre, K.; Johanson, U.; Englund, M.; Engström, P. The MADS-box gene DAL1 is a potential mediator of the juvenile-to-adult transition in Norway spruce (Picea abies). Plant J. 2004, 40, 546–557. [Google Scholar] [CrossRef] [PubMed]
- Katahata, S.-I.; Futamura, N.; Igasaki, T.; Shinohara, K. Functional analysis of SOC1-like and AGL6-like MADS-box genes of the gymnosperm Cryptomeria japonica. Tree Genet. Genomes 2014, 10, 317–327. [Google Scholar] [CrossRef]
- Koo, S.C.; Bracko, O.; Park, M.S.; Schwab, R.; Chun, H.J.; Park, K.M.; Seo, J.S.; Grbic, V.; Balasubramanian, S.; Schmid, M.; et al. Control of lateral organ development and flowering time by the Arabidopsis thaliana MADS-box Gene AGAMOUS-LIKE6. Plant J. 2010, 62, 807–816. [Google Scholar] [CrossRef] [PubMed]
- Hsu, W.-H.; Yeh, T.-J.; Huang, K.-Y.; Li, J.-Y.; Chen, H.-Y.; Yang, C.-H. AGAMOUS-LIKE13, a putative ancestor for the E functional genes, specifies male and female gametophyte morphogenesis. Plant J. 2014, 77, 1–15. [Google Scholar] [CrossRef]
- Hatsuda, Y.; Nishio, S.; Komori, S.; Nishiyama, M.; Kanahama, K.; Kanayama, Y. Relationship between MdMADS11 Gene Expression and Juvenility in Apple. J. Jpn. Soc. Hortic. Sci. 2011, 80, 396–403. [Google Scholar] [CrossRef] [Green Version]
- Viaene, T.; Vekemans, D.; Becker, A.; Melzer, S.; Geuten, K. Expression divergence of the AGL6 MADS domain transcription factor lineage after a core eudicot duplication suggests functional diversification. BMC Plant Biol. 2010, 10, 148. [Google Scholar] [CrossRef] [Green Version]
- Su, C.L.; Chen, W.C.; Lee, A.Y.; Chen, C.Y.; Chang, Y.C.; Chao, Y.T.; Shih, M.C. A modified ABCDE model of flowering in orchids based on gene expression profiling studies of the moth orchid Phalaenopsis aphrodite. PLoS ONE 2013, 8, e80462. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Fan, T.; Song, J.; Sun, W.; Xia, K.; Liao, J.; Zhang, M. Functional conservation and divergence of four ginger AP1/AGL9 MADS-box genes revealed by analysis of their expression and protein-protein interaction, and ectopic expression of AhFUL gene in Arabidopsis. PLoS ONE 2014, 9, e114134. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Li, W.; Dong, X.; Guo, W.; Shu, H. Ectopic expression of a hyacinth AGL6 homolog caused earlier flowering and homeotic conversion in Arabidopsis. Sci. China C Life Sci. 2007, 50, 676–689. [Google Scholar] [CrossRef] [PubMed]
- Rijpkema, A.S.; Zethof, J.; Gerats, T.; Vandenbussche, M. The petunia AGL6 gene has a SEPALLATA-like function in floral patterning. Plant J. 2009, 60, 1–9. [Google Scholar] [CrossRef]
- Wang, B.-G.; Zhang, Q.; Wang, L.-G.; Duan, K.; Pan, A.-H.; Tang, X.-M.; Sui, S.-Z.; Li, M.-Y. The AGL6-like Gene CpAGL6, a Potential Regulator of Floral Time and Organ Identity in Wintersweet (Chimonanthus praecox). J. Plant Growth Regul. 2011, 30, 343–352. [Google Scholar] [CrossRef]
- Duan, Y.; Xing, Z.; Diao, Z.; Xu, W.; Li, S.; Du, X.; Wu, G.; Wang, C.; Lan, T.; Meng, Z.; et al. Characterization of Osmads6-5, a null allele, reveals that OsMADS6 is a critical regulator for early flower development in rice (Oryza sativa L.). Plant Mol. Biol. 2012, 80, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Thompson, B.E.; Bartling, L.; Whipple, C.; Hall, D.H.; Sakai, H.; Schmidt, R.; Hake, S. bearded-ear encodes a MADS box transcription factor critical for maize floral development. Plant Cell 2009, 21, 2578–2590. [Google Scholar] [CrossRef] [Green Version]
- Hsu, H.F.; Huang, C.H.; Chou, L.T.; Yang, C.H. Ectopic expression of an orchid (Oncidium Gower Ramsey) AGL6-like gene promotes flowering by activating flowering time genes in Arabidopsis thaliana. Plant Cell Physiol. 2003, 44, 783–794. [Google Scholar] [CrossRef] [Green Version]
- Yoo, S.K.; Wu, X.; Lee, J.S.; Ahn, J.H. AGAMOUS-LIKE 6 is a floral promoter that negatively regulates the FLC/MAF clade genes and positively regulates FT in Arabidopsis. Plant J. 2011, 65, 62–76. [Google Scholar] [CrossRef]
- Duarte, J.M.; Cui, L.; Wall, P.K.; Zhang, Q.; Zhang, X.; Leebens-Mack, J.; Ma, H.; Altman, N.; de Pamphilis, C.W. Expression Pattern Shifts Following Duplication Indicative of Subfunctionalization and Neofunctionalization in Regulatory Genes of Arabidopsis. Mol. Biol. Evol. 2005, 23, 469–478. [Google Scholar] [CrossRef]
- Airoldi, C.; Davies, B. Gene Duplication and the Evolution of Plant MADS-box Transcription Factors. J. Genet. Genom. 2012, 39, 157–165. [Google Scholar] [CrossRef]
- Panchy, N.; Lehti-Shiu, M.; Shiu, S.H. Evolution of Gene Duplication in Plants. Plant Physiol. 2016, 171, 2294–2316. [Google Scholar] [CrossRef]
- Ali, Z.; Raza, Q.; Atif, R.M.; Aslam, U.; Ajmal, M.; Chung, G. Genetic and Molecular Control of Floral Organ Identity in Cereals. Int. J. Mol. Sci. 2019, 20, 2743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuchimoto, S.; Mayama, T.; van der Krol, A.; Ohtsubo, E. The whorl-specific action of a petunia class B floral homeotic gene. Genes Cells 2000, 5, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Zhang, R.Z.; Guo, J.J.; Liu, D.M.; Li, A.L.; Fan, R.C.; Mao, L.; Zhang, X.Q. Genome-wide analysis of the MADS-box gene family in Brachypodium distachyon. PLoS ONE 2014, 9, e84781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yockteng, R.; Almeida, A.M.; Morioka, K.; Alvarez-Buylla, E.R.; Specht, C.D. Molecular evolution and patterns of duplication in the SEP/AGL6-like lineage of the Zingiberales: A proposed mechanism for floral diversification. Mol. Biol. Evol. 2013, 30, 2401–2422. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Deng, S.; Chen, L.; Jia, Z.; Sang, Z.; Zhu, Z.; Ma, L.; Chen, F. Gene duplication led to divergence of expression patterns, protein-protein interaction patterns and floral development functions of AGL6-like genes in the basal angiosperm Magnolia wufengensis (Magnoliaceae). Tree Physiol. 2019, 39, 861–876. [Google Scholar] [CrossRef]
- Ohmori, S.; Kimizu, M.; Sugita, M.; Miyao, A.; Hirochika, H.; Uchida, E.; Nagato, Y.; Yoshida, H. MOSAIC FLORAL ORGANS1, an AGL6-Like MADS Box Gene, Regulates Floral Organ Identity and Meristem Fate in Rice. Plant Cell 2009, 21, 3008–3025. [Google Scholar] [CrossRef] [Green Version]
- Tao, J.; Liang, W.; An, G.; Zhang, D. OsMADS6 Controls Flower Development by Activating Rice FACTOR OF DNA METHYLATION LIKE1. Plant Physiol. 2018, 177, 713–727. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.; Liu, J.; Liang, W.; Dou, Y.; Fu, R.; Li, W.; Feng, C.; Gao, C.; Zhang, D.; Kang, Z.; et al. Wheat AGAMOUS LIKE 6 transcription factors function in stamen development by regulating the expression of Ta APETALA3. Development 2019, 146, dev177527. [Google Scholar] [CrossRef] [Green Version]
- Xiang, L.; Chen, Y.; Chen, L.; Fu, X.; Zhao, K.; Zhang, J.; Sun, C. B and E MADS-box genes determine the perianth formation in Cymbidium goeringii Rchb.f. Physiol. Plant 2018, 162, 353–369. [Google Scholar] [CrossRef]
- Liu, J.; Fu, X.; Dong, Y.; Lu, J.; Ren, M.; Zhou, N.; Wang, C. MIKC(C)-type MADS-box genes in Rosa chinensis: The remarkable expansion of ABCDE model genes and their roles in floral organogenesis. Hortic. Res. 2018, 5, 25. [Google Scholar] [CrossRef]
- Busch, A.W.U.; Zachgo, S. Flower symmetry evolution: Towards understanding the abominable mystery of angiosperm radiation. BioEssays 2009, 31, 1181–1190. [Google Scholar] [CrossRef] [PubMed]
- Hileman, L.C. Trends in flower symmetry evolution revealed through phylogenetic and developmental genetic advances. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.D. Functional Analysis of MADS-box Genes Related to Floral Organ Development in Prunus Mume; Beijing Forestry University: Beijing, China, 2015. [Google Scholar]
- Song, G.Q.; Chen, Q. Overexpression of the MADS-box gene K-domain increases the yield potential of blueberry. Plant Sci. 2018, 276, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Jack, T. Defining subdomains of the K domain important for protein–protein interactions of plant MADS proteins. Plant Mol. Biol. 2004, 55, 45–59. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.-Y.; Chiu, Y.-F.; Wu, J.-W.; Yang, C.-H. Four Orchid (Oncidium Gower Ramsey) AP1/AGL9-like MADS Box Genes Show Novel Expression Patterns and Cause Different Effects on Floral Transition and Formation in Arabidopsis thaliana. Plant Cell Physiol. 2009, 50, 1425–1438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbesier, L.; Coupland, G. The quest for florigen: A review of recent progress. J. Exp. Bot. 2006, 57, 3395–3403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, T.; Irish, V.F. Gene networks controlling petal organogenesis. J. Exp. Bot. 2015, 67, 61–68. [Google Scholar] [CrossRef] [Green Version]
- Kaufmann, K.; Melzer, R.; Theissen, G. MIKC-type MADS-domain proteins: Structural modularity, protein interactions and network evolution in land plants. Gene 2005, 347, 183–198. [Google Scholar] [CrossRef]
- Valverde, F.; Mouradov, A.; Soppe, W.; Ravenscroft, D.; Samach, A.; Coupland, G. Photoreceptor regulation of CONSTANS protein in photoperiodic flowering. Science 2004, 303, 1003–1006. [Google Scholar] [CrossRef] [Green Version]
- Searle, I.; He, Y.; Turck, F.; Vincent, C.; Fornara, F.; Kröber, S.; Amasino, R.A.; Coupland, G. The transcription factor FLC confers a flowering response to vernalization by repressing meristem competence and systemic signaling in Arabidopsis. Genes Dev. 2006, 20, 898–912. [Google Scholar] [CrossRef]
- Irish, V.F. The Arabidopsis petal: A model for plant organogenesis. Trends Plant Sci. 2008, 13, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Monniaux, M.; Pieper, B.; McKim, S.M.; Routier-Kierzkowska, A.L.; Kierzkowski, D.; Smith, R.S.; Hay, A. The role of APETALA1 in petal number robustness. eLife 2018, 7, e39399. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Chen, S.; Jiang, J.; Chen, Y.; Chen, F.; Teng, N.; Yin, D.; Huang, C. The expression of floral organ identity genes in contrasting water lily cultivars. Plant Cell. Rep. 2011, 30, 1909–1918. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Effgen, S.; Meyer, R.C.; Theres, K.; Koornneef, M. Epistatic Natural Allelic Variation Reveals a Function of AGAMOUS-LIKE6 in Axillary Bud Formation in Arabidopsis. Plant Cell 2012, 24, 2364–2379. [Google Scholar] [CrossRef] [Green Version]
- Schauer, S.E.; Schlüter, P.M.; Baskar, R.; Gheyselinck, J.; Bolaños, A.; Curtis, M.D.; Grossniklaus, U. Intronic regulatory elements determine the divergent expression patterns of AGAMOUS-LIKE6 subfamily members in Arabidopsis. Plant J. 2009, 59, 987–1000. [Google Scholar] [CrossRef]
- Yoo, S.K.; Hong, S.M.; Lee, J.S.; Ahn, J.H. A genetic screen for leaf movement mutants identifies a potential role for AGAMOUS-LIKE 6 (AGL6) in circadian-clock control. Mol. Cells 2011, 31, 281–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Liang, W.; Jia, R.; Yin, C.; Zong, J.; Kong, H.; Zhang, D. The AGL6-like gene OsMADS6 regulates floral organ and meristem identities in rice. Cell Res. 2010, 20, 299–313. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.-x.; Zhang, K.; Li, L.; Fei, Y.; Chen, F. Characterization of Two AGL6–Like Genes from a Chinese Endemic Woody Tree, Manglietia patungensis (Magnoliaceae) Provides Insight into Perianth Development and Evolution in Basal Angiosperms. Forests 2019, 10, 669. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Xu, Z.; Yong, X.; Ahmad, S.; Yang, W.; Cheng, T.; Wang, J.; Zhang, Q. SEP-class genes in Prunus mume and their likely role in floral organ development. BMC Plant Biol. 2017, 17, 10. [Google Scholar] [CrossRef]
Figure 1.
Sequence alignment of PmAGL6-1, PmAGL6-2 and AGL6 proteins from other plants. The MADS domain and K domain regions are underlined, and AGL6 motifs I and II are boxed. Accession numbers for the aligned sequences are as follows: MdAGL6 (Malus domestica, NP_001280892), ZmAGL6 (Zea mays, NP_001105332.1), PbAGL6 (Pyrus×bretschneideri, XP_009375842.1), PdAGL6 (Prunus dulcis, BBG97451), PpAGL6 (Pyrus pyrifolia var. culta, BAK18784.2), PyAGL6 (Prunus yedoensis var. nudiflora, PQQ17269.1), HoAGL6 (Hyacinthus orientalis, AAT88088.1), CpAGL6 (Chimonanthus praecox, ACN88212.1), AGL6 (Arabidopsis thaliana, NP_182089) and AGL13 (Arabidopsis thaliana, NP_191671).
Figure 1.
Sequence alignment of PmAGL6-1, PmAGL6-2 and AGL6 proteins from other plants. The MADS domain and K domain regions are underlined, and AGL6 motifs I and II are boxed. Accession numbers for the aligned sequences are as follows: MdAGL6 (Malus domestica, NP_001280892), ZmAGL6 (Zea mays, NP_001105332.1), PbAGL6 (Pyrus×bretschneideri, XP_009375842.1), PdAGL6 (Prunus dulcis, BBG97451), PpAGL6 (Pyrus pyrifolia var. culta, BAK18784.2), PyAGL6 (Prunus yedoensis var. nudiflora, PQQ17269.1), HoAGL6 (Hyacinthus orientalis, AAT88088.1), CpAGL6 (Chimonanthus praecox, ACN88212.1), AGL6 (Arabidopsis thaliana, NP_182089) and AGL13 (Arabidopsis thaliana, NP_191671).

Figure 2.
Phylogenetic analysis of AGL6 proteins. The tree was constructed via the neighbor-joining method using MEGA5-X software with a bootstrap analysis of 1000 replicates. The accession numbers of the other proteins included in the phylogenetic analysis are as follows: PpAGL6-1 (Prunus persica, XP_020412713.1), PpAGL6-2 (Prunus persica, XP_007220094), PdAGL6 (Prunus dulcis, BBG97451), MdAGL6 (Malus domestica, NP_001280892), PavAGL6-1 (Prunus avium, XP_021825726.1), PavAGL6-2 (Prunus avium, XP_021825734.1), PyAGL6 (Prunus yedoensis var. nudiflora, PQQ17269.1), PbAGL6 (Pyrus×bretschneideri, XP_009375842.1), PamAGL6-1 (Persea Americana, ABG49493.1), PamAGL6-2 (Persea Americana, ABG49494.1), MapaAGL6-1 (Magnolia patungensis, ATB53133.1), MapaAGL6-2 (Magnolia patungensis, ATB53135.1), MawuAGL6-1 (Magnolia wufengensis, AOZ15901.1), MawuAGL6-2 (Magnolia wufengensis, AOZ15902.1), TaAGL6-A (Triticum aestivum, AYP71082.1), TaAGL6-B (Triticum aestivum, AYP71084.1), TaAGL6-D (Triticum aestivum, AYP71083.1), AGL6 (Arabidopsis thaliana, NP_182089), AGL13 (Arabidopsis thaliana, NP_191671), CgAGL6-1 (Cymbidium goeringii, ACT66279.1), CgAGL6-2 (Cymbidium goeringii, APY18455.1), CgAGL6-3 (Cymbidium goeringii, AMW17745.1), OsMADS6 (Oryza sativa, Q6EU39.1), OsMADS17 (Oryza sativa, ACX35552.1), GgAGL6-1 (Gnetum gnemon, CAB44455.1), GgAGL6-2 (Gnetum gnemon, CAB44457.1), GbAGL6-1 (Ginkgo biloba, BAD93172.1) and GbAGL6-2 (Ginkgo biloba, AIC79629.1).
Figure 2.
Phylogenetic analysis of AGL6 proteins. The tree was constructed via the neighbor-joining method using MEGA5-X software with a bootstrap analysis of 1000 replicates. The accession numbers of the other proteins included in the phylogenetic analysis are as follows: PpAGL6-1 (Prunus persica, XP_020412713.1), PpAGL6-2 (Prunus persica, XP_007220094), PdAGL6 (Prunus dulcis, BBG97451), MdAGL6 (Malus domestica, NP_001280892), PavAGL6-1 (Prunus avium, XP_021825726.1), PavAGL6-2 (Prunus avium, XP_021825734.1), PyAGL6 (Prunus yedoensis var. nudiflora, PQQ17269.1), PbAGL6 (Pyrus×bretschneideri, XP_009375842.1), PamAGL6-1 (Persea Americana, ABG49493.1), PamAGL6-2 (Persea Americana, ABG49494.1), MapaAGL6-1 (Magnolia patungensis, ATB53133.1), MapaAGL6-2 (Magnolia patungensis, ATB53135.1), MawuAGL6-1 (Magnolia wufengensis, AOZ15901.1), MawuAGL6-2 (Magnolia wufengensis, AOZ15902.1), TaAGL6-A (Triticum aestivum, AYP71082.1), TaAGL6-B (Triticum aestivum, AYP71084.1), TaAGL6-D (Triticum aestivum, AYP71083.1), AGL6 (Arabidopsis thaliana, NP_182089), AGL13 (Arabidopsis thaliana, NP_191671), CgAGL6-1 (Cymbidium goeringii, ACT66279.1), CgAGL6-2 (Cymbidium goeringii, APY18455.1), CgAGL6-3 (Cymbidium goeringii, AMW17745.1), OsMADS6 (Oryza sativa, Q6EU39.1), OsMADS17 (Oryza sativa, ACX35552.1), GgAGL6-1 (Gnetum gnemon, CAB44455.1), GgAGL6-2 (Gnetum gnemon, CAB44457.1), GbAGL6-1 (Ginkgo biloba, BAD93172.1) and GbAGL6-2 (Ginkgo biloba, AIC79629.1).
Figure 3.
Expression patterns of PmAGL6-1 and PmAGL6-2. (a) Expression patterns of PmAGL6-1 and PmAGL6-2 in different organs of P. mume. R: root, Ste: stem, L: leaf, Se: sepal, Pe: petal, Sta: stamen, Pis: pistile, Fr1–3: fruit development stages 1–3. (b) Expression patterns of PmAGL6-1 and PmAGL6-2 during P. mume floral bud differentiation. S1: flower primordium formation, S2: sepal initiation, S3: petal initiation, S4: stamen initiation, S5: pistil initiation, S6: ovule development, S7: anther development. Significant differences are identified by SPSS with Duncan’s test (p < 0.05) and are represented by different letters above the error bars.
Figure 3.
Expression patterns of PmAGL6-1 and PmAGL6-2. (a) Expression patterns of PmAGL6-1 and PmAGL6-2 in different organs of P. mume. R: root, Ste: stem, L: leaf, Se: sepal, Pe: petal, Sta: stamen, Pis: pistile, Fr1–3: fruit development stages 1–3. (b) Expression patterns of PmAGL6-1 and PmAGL6-2 during P. mume floral bud differentiation. S1: flower primordium formation, S2: sepal initiation, S3: petal initiation, S4: stamen initiation, S5: pistil initiation, S6: ovule development, S7: anther development. Significant differences are identified by SPSS with Duncan’s test (p < 0.05) and are represented by different letters above the error bars.
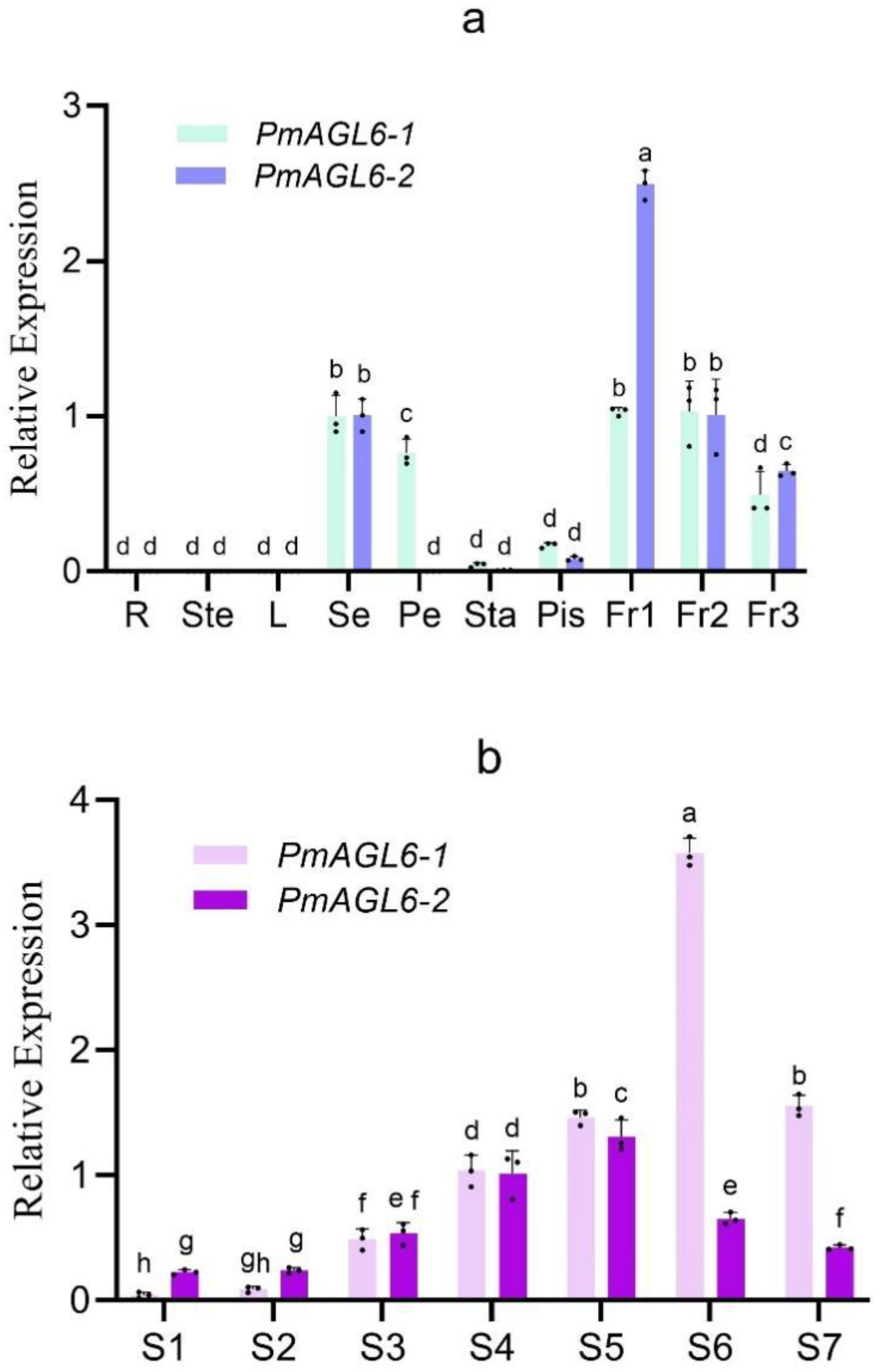
Figure 4.
Functional analysis of PmAGL6-1 and PmAGL6-2 by transforming Arabidopsis plants. (IA–IK) Constitutive expression in 35S::PmAGL6-1 transgenic Arabidopsis plants. (IIA–IIJ) Constitutive expression in 35S:: PmAGL6-2 transgenic Arabidopsis plants. (IA) Early-flowering phenotype (A4 line) following the ectopic expression of PmAGL6-1 (left) and normal vegetative growth in the wild-type (right). (IIA) Early-flowering phenotype (B4 line) following the ectopic expression of PmAGL6-2 (left) and normal vegetative growth in the wild-type (right). (IB) Extremely early-flowering phenotype (A1 line) following the ectopic expression of PmAGL6-1. (IC) Flower from wild-type Arabidopsis. (ID,IIB) Flower with three petals. (IE,IIC) Flower with five petals. (IF,IID) Bilaterally symmetrical flower. (IG,IIE) The top two flowers of the inflorescence combine to form a single flower. (IH,IIF) The top three flowers of the inflorescence combine to form a single flower. (II,IIG) The top four flowers of the inflorescence combine to form a single flower. (IJ,IIH) Flower with overlapping petals. (IK,III) Curved fruit pod. (IIJ) Flower with a sharp petal apex.
Figure 4.
Functional analysis of PmAGL6-1 and PmAGL6-2 by transforming Arabidopsis plants. (IA–IK) Constitutive expression in 35S::PmAGL6-1 transgenic Arabidopsis plants. (IIA–IIJ) Constitutive expression in 35S:: PmAGL6-2 transgenic Arabidopsis plants. (IA) Early-flowering phenotype (A4 line) following the ectopic expression of PmAGL6-1 (left) and normal vegetative growth in the wild-type (right). (IIA) Early-flowering phenotype (B4 line) following the ectopic expression of PmAGL6-2 (left) and normal vegetative growth in the wild-type (right). (IB) Extremely early-flowering phenotype (A1 line) following the ectopic expression of PmAGL6-1. (IC) Flower from wild-type Arabidopsis. (ID,IIB) Flower with three petals. (IE,IIC) Flower with five petals. (IF,IID) Bilaterally symmetrical flower. (IG,IIE) The top two flowers of the inflorescence combine to form a single flower. (IH,IIF) The top three flowers of the inflorescence combine to form a single flower. (II,IIG) The top four flowers of the inflorescence combine to form a single flower. (IJ,IIH) Flower with overlapping petals. (IK,III) Curved fruit pod. (IIJ) Flower with a sharp petal apex.

Figure 5.
Analysis of related gene expression in transformants. (a) Expression of PmAGL6-1 in five transgenic Arabidopsis plants. (b) Expression of PmAGL6-2 in five transgenic Arabidopsis plants. (c) Expression of CYC in transgenic plants. (d) Expression of flowering regulators in the 35S::PmAGL6-1 transformant A1 line and the 35S::PmAGL6-2 transformant B4 line. Significant differences are identified by SPSS with Duncan’s test (p < 0.05) and are represented by different letters above the error bars.
Figure 5.
Analysis of related gene expression in transformants. (a) Expression of PmAGL6-1 in five transgenic Arabidopsis plants. (b) Expression of PmAGL6-2 in five transgenic Arabidopsis plants. (c) Expression of CYC in transgenic plants. (d) Expression of flowering regulators in the 35S::PmAGL6-1 transformant A1 line and the 35S::PmAGL6-2 transformant B4 line. Significant differences are identified by SPSS with Duncan’s test (p < 0.05) and are represented by different letters above the error bars.
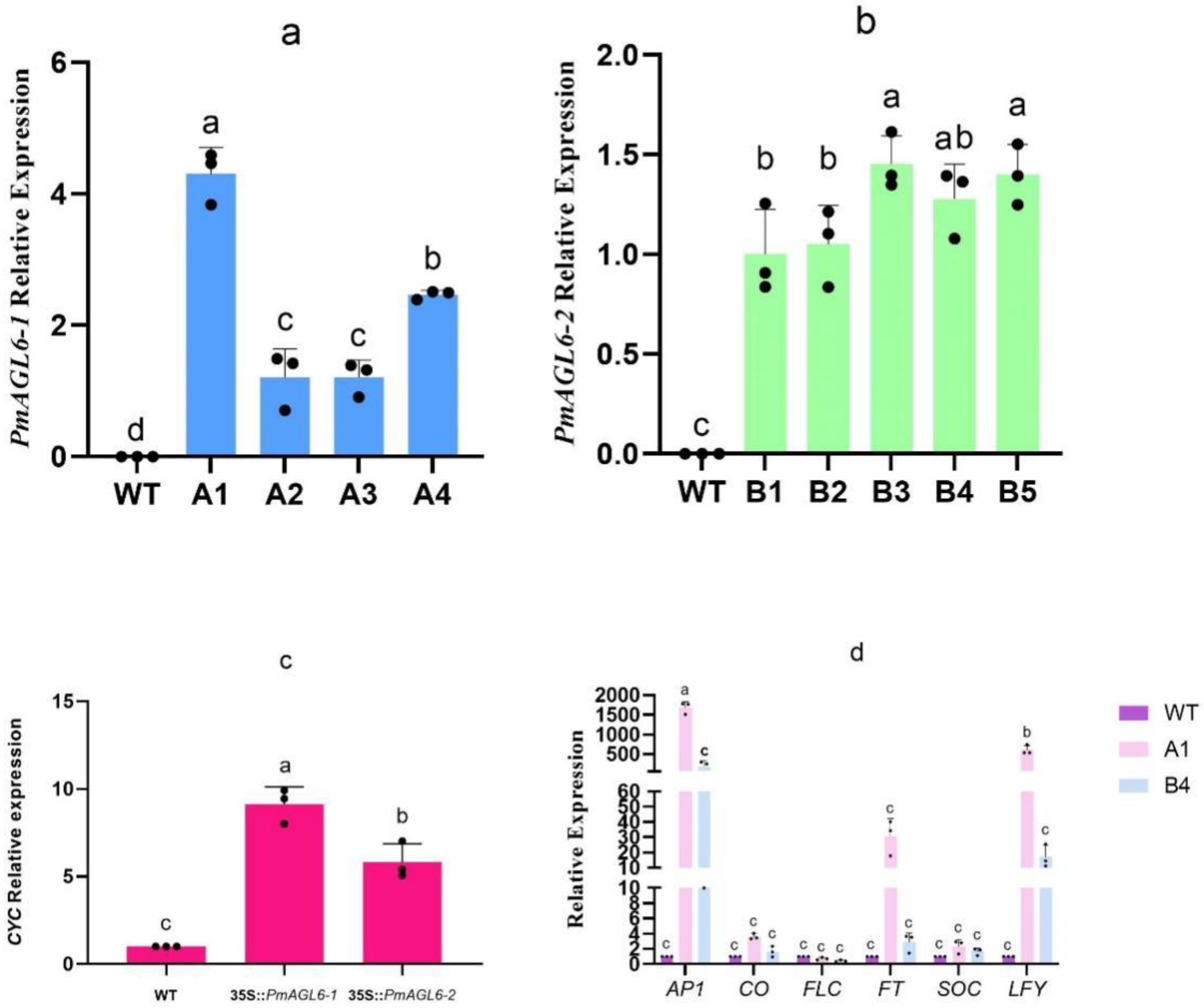
Figure 6.
Positive results of the Y2H analysis of protein–protein interactions among the products of the two AGL6 genes and other floral organ identity-determining genes in P. mume. T7-53/T7-T and T7-Lam/T7-T were the positive and negative controls, respectively. + represents the intensity of the interaction and − indicates no interaction.
Figure 6.
Positive results of the Y2H analysis of protein–protein interactions among the products of the two AGL6 genes and other floral organ identity-determining genes in P. mume. T7-53/T7-T and T7-Lam/T7-T were the positive and negative controls, respectively. + represents the intensity of the interaction and − indicates no interaction.


Figure 7.
Positive results of the BiFC analysis of protein–protein interactions among the products of the two AGL6 genes and other floral organ identity-determining genes in P. mume. YFP: yellow fluorescent protein, BF: bright-field image, merged: merged YFP and BF images.
Figure 7.
Positive results of the BiFC analysis of protein–protein interactions among the products of the two AGL6 genes and other floral organ identity-determining genes in P. mume. YFP: yellow fluorescent protein, BF: bright-field image, merged: merged YFP and BF images.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Primers for qPCR.
| Primers | Primers Sequence |
|---|---|
| qPmAGL6-1-F | TCTCGGACCTCTGAATGTGA |
| qPmAGL6-1-R | TTCTGTCTCCAGCTTGACC |
| qPmAGL6-2-F | ACCGTCAATGTTGCTACTCC |
| qPmAGL6-2-R | TCACCAGTTCTTTCAGGCGAA |
| qPP2A-F | ATATAGCTGCTCAGTTCAACC |
| qPP2A-R | AAAAACAGTCACCACATTCTT |
| qActin-F | TCTCTATGCCAGTGGTCGTA |
| qActin-R | CCTCAGGACAACGGAATC |
| qAP1-F | CAGATCAAGGAGAGGGAA |
| qAP1-R | TTGATACAGACCACCCAT |
| qSOC-F | CTCCAATATGCAAGATACCA |
| qSOC-R | TATGCCTTCTCCCAAGAGTT |
| qLFY-F | CCCAAGAAGGGTTATCTGA |
| qLFY-R | AAACGGATGCTCCCTCTG |
| qCO-F | CCATTAACCATAACGCATAC |
| qCO-R | GTCAGGTTGTTGCTCTACT |
| qFT-F | TGGAACAACCTTTGGCAATG |
| qFT-R | GTCTTCTTCCTCCGCAGC |
| qCYC-F | AAGGCTTTGAGTTTCCTGAGGTG |
| qCYC-R | TGAGCAGCCAGTCTAACGTTTTAC |
| qFLC-F | CGAAGCTGATAATATGGAGATGT |
| qFLC-R | AGATATACAACGTGCACCCTTCA |
| PmAGL6-1F | ATGGGGAGAAGGAAAGTGGTG |
| PmAGL6-1R | TCAAAGAACCCATCCCTG |
| PmAGL6-2F | ATGGGGAGAAGGAAAGTGGTG |
| PmAGL6-2R | TTAAAGAAGCCAACCCCGC |
Table 2.
Days needed for flowering and number of rosette leaves.
| Transgenic Line/WT | Days Needed for Flowering | Number of Rosette Leaves | Transgenic Line/WT | Days Needed for Flowering | Number of Rosette Leaves |
|---|---|---|---|---|---|
| WT | 42.20 ± 1.66a | 10.30 ± 1.37a | B1 | 31.80 ± 0.51bc | 7.50 ± 0.40c |
| A1 | 26.80 ± 0.80d | 7.40 ± 0.68c | B2 | 33.40 ± 1.27b | 7.90 ± 1.38bc |
| A2 | 30.20 ± 0.74c | 7.80 ± 0.49bc | B3 | 31.60 ± 0.52bc | 8.20 ± 0.36bc |
| A3 | 32.10 ± 1.74bc | 8.40 ± 1.68bc | B4 | 30.40 ± 0.69bc | 7.20 ± 0.73bc |
| A4 | 29.20 ± 0.75c | 70.60 ± 0.98bc | B5 | 33.01 ± 1.17b | 7.40 ± 0.48c |
Significant differences are identified by SPSS with Duncan’s test (p < 0.05) and are represented by different letters above the error bars.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, L.; Song, J.; Han, X.; Yu, Y.; Wu, Q.; Qi, S.; Xu, Z. Functional Divergence Analysis of AGL6 Genes in Prunus mume. Plants 2023, 12, 158. https://doi.org/10.3390/plants12010158
AMA Style
Wang L, Song J, Han X, Yu Y, Wu Q, Qi S, Xu Z. Functional Divergence Analysis of AGL6 Genes in Prunus mume. Plants. 2023; 12(1):158. https://doi.org/10.3390/plants12010158
Chicago/Turabian StyleWang, Lei, Jinhai Song, Xu Han, Yunyan Yu, Qikui Wu, Shuai Qi, and Zongda Xu. 2023. "Functional Divergence Analysis of AGL6 Genes in Prunus mume" Plants 12, no. 1: 158. https://doi.org/10.3390/plants12010158
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.