
Management of Post-Harvest Anthracnose: Current Approaches and Future Perspectives


Department of Agricultural and Food Sciences (DISTAL), University of Bologna, Viale Fanin 44, 40126 Bologna, Italy
*
Authors to whom correspondence should be addressed.
Plants 2022, 11(14), 1856; https://doi.org/10.3390/plants11141856
Submission received: 19 June 2022
/
Revised: 8 July 2022
/
Accepted: 11 July 2022
/
Published: 15 July 2022
(This article belongs to the Special Issue Interactions between Colletotrichum Species and Plants Ⅱ)
Abstract
:Anthracnose is a severe disease caused by Colletotrichum spp. on several crop species. Fungal infections can occur both in the field and at the post-harvest stage causing severe lesions on fruits and economic losses. Physical treatments and synthetic fungicides have traditionally been the preferred means to control anthracnose adverse effects; however, the urgent need to decrease the use of toxic chemicals led to the investigation of innovative and sustainable protection techniques. Evidence for the efficacy of biological agents and vegetal derivates has been reported; however, their introduction into actual crop protection strategies requires the solutions of several critical issues. Biotechnology-based approaches have also been explored, revealing the opportunity to develop innovative and safe methods for anthracnose management through genome editing and RNA interference technologies. Nevertheless, besides the number of advantages related to their use, e.g., the putative absence of adverse effects due to their high specificity, a number of aspects remain to be clarified to enable their introduction into Integrated Pest Management (IPM) protocols against Colletotrichum spp. disease.
1. Epidemiology and Pathology of Colletotrichum spp.
1.1. Interaction between Colletotrichum spp. and Their Hosts
The genus Colletotrichum comprises more than 200 fungal species, informally gathered in 15 species complexes [1]. Many of them are pathogens of important crops where they cause anthracnose, a severe disease with great economic impact. For this reason, Colletotrichum spp. have been listed among the top ten most relevant fungal pathogens worldwide [2,3].
Plant infections occur mainly in tropical and sub-tropical regions and less frequently at temperate latitudes since their onset requires warm temperature and high relative humidity [4,5]. Duration and intensity of rainfalls, wetness of the leaf surface and light intensity have also been reported as factors positively correlated with the infective process [6]. For these reasons, crops located in regions with frequent precipitations, never really drying between rainfalls, are particularly affected by Colletotrichum infections.
With respect to fruit infection, the early stages of the interaction with host tissues are similar for all the Colletotrichum species [7]: conidiospores spread from infected vegetal material or through insects, adhere by means of a hemicellulosic mucilage to the external vegetal surface, germinate and infect often by mean of specialized structures, such as appressoria [8,9,10,11]. Infections can take place even by penetration through stomata, lenticels, wounds, or abscission of scar tissue [11,12,13,14,15]. Although specific host–pathogen interaction and infection strategies have been detected, such as in C. acutatum sensu lato that exhibit four different colonization pathways [16], the process proceeds according to two main strategies, depending on the different species, hosts, and tissues: (i) intracellular hemibiotrophy or (ii) subcuticular, intramural necrotrophy [6,7,17,18]. The first one includes an initial short (from 24 h to over 72 h) biotrophic stage, characterized by the formation of intracellular primary hyphae, largely differing among species in their morphology. This stage is followed by a destructive necrotrophic phase when secondary ramified narrow hyphae spread throughout the host tissues [19,20,21,22]. “Signals” from the fruit ripening stage, such as occurrence of disaggregated cell wall components, accumulation of ethylene, changes in the environmental pH and in the content of organic compounds, decrease of both antifungal substances (e.g., polyphenols and phytoalexins) and host defense mechanisms can act as promoters of the pathogen necrotrophic phase [23,24,25,26,27]. On the contrary, subcuticular, intramural necrotrophy, typical of C. capsici (syn. C. truncatum) and C. circinans, consists of an early asymptomatic short (24 h) stage where the pathogen grows within the periclinal and anticlinal walls of the epidermal cells (intramural development). Subsequently, these species rapidly spread throughout the host tissues producing destructive effects [28,29].
1.2. Antracnhose Disease: A Challenge for the Agri-Food Sector
Anthracnose symptoms highly vary depending on the plant tissue and cause huge losses of fruit production [11]. For instance, strawberry plants, which are susceptible to several Colletotrichum species, can exhibit serious anthracnose symptoms in all its parts [30,31]. For this crop, anthracnose can account for up to 80% and more than 50% of plant and fruit losses in nurseries and in field, respectively [31,32,33].
In general, symptoms on stems or leaves range from small greyish-brown sunken spots to darker patches on branches. On the contrary, infected blossoms appear dry and rotten with compromised fruit development, whereas on fruits anthracnose produce dark lesions surrounding pink or orange conidia masses that under suitable conditions can evolve causing tissue necrosis [4,11,34,35,36]. In any case, the highest impact of this disease is due to the fruit damage during postharvest storage (Figure 1).
Here symptoms can cause consistent decays and a reduction in both quality and aesthetic standards, resulting in severe economic losses [23,37]. For instance, banana fruits are highly affected by anthracnose that is responsible for losses up to 30–40% of the marketable products [38]. The damages caused by this disease even grow in other susceptible hosts. In this context, the shelf life of papaya in India, the main producer of this fruit worldwide [5], is seriously compromised by Colletotrichum spp., determining losses up to 93% [39,40]. Similarly, anthracnose represents a severe post-harvest disease for mango fruits, especially in regions where infections are favored by the climatic conditions and its incidence can reach the 100% of the production [23].
Post-harvest anthracnose is mainly due to infections taking place in the field that become quiescent until the occurrence of conditions favorable for pathogen germination and development [37,41,42]. As a result, the disease is particularly severe for climacteric fruits such as banana, guava, avocado, pear, mango, and papaya since their ripening stage includes physiological and biochemical changes, due to the re-modelling and degradation of cell walls, providing suitable conditions for the fungal development [11,43,44]. Nevertheless, anthracnose represents a serious constrain even for non-climateric fruits, such as strawberry, citrus, and dragon fruit [3,45,46,47].
After a brief insight into the methods traditionally applied for the management of anthracnose disease, this review will revise the most important alternative and eco-friendly approaches that have been recently explored to counteract anthracnose, with a special focus at their possible application at post-harvest stage. Particular attention will be given to recently developed biotechnology-based strategies, that offer high specificity and low risk of negative effects on environment and human health (Figure 2).
2. Traditional Approaches
The strategies traditionally applied to counteract anthracnose disease in fruits after harvesting are mainly based on physical treatments and applications of few chemical fungicides.
Physical treatments include environmentally friendly practices finalized to inhibit Colletotrichum development, such as vapor heat, forced-air dry heat and hot water dips of harvested products. However, some of these storage techniques are not suitable for the majority of fruit production, since potentially affecting the fruit quality. For instance, hot water dips have been associated to decays of organoleptic and nutritional features and subsequent reduction of shelf life [3,48,49,50].
On the other hand, chemical-based control of anthracnose (both in the field and in commercial packinghouses after harvest) has been for many years the main tool to prevent the damages. Indeed, synthetic fungicides are effective means to reduce Colletotrichum inoculum in the field and the fungal development during the fruit storage [3,15]. The products traditionally used against Colletotrichum spp. include cupric products, strobirulines, dithiocarbamates, benzimidazole, and triazole compounds, together with other chemicals, such as prochloraz, imazalil, and chorothalonil [3,7]. However, their improper use, as well as consecutive treatments based on substances with the same mechanisms of action, have been related to decreases in the sensitivity of pathogens and the emergence of resistant strains [23,33,47,51,52,53,54,55,56,57,58,59,60]. For instance, impaired sensitivity toward prochloraz and benzimidazole has been revealed in C. gloeosporioides sensu lato isolates from avocado and mango fruits [23,51]. In addition, the use of fungicides in the agri-food sector represents a serious risk for the human health and the environment, given both the presence of chemical residues on fruits and their pollutant effects on soil, water, and not-target organisms [3,55,61,62,63,64,65]. For example, the use of thiophanate-methyl in post-harvest anthracnose control, has been associated with severe toxic effects on the human health [3]. Similarly, prochloraz, effective to prevent the phytopathology on mango and avocado fruits [66,67], has been recently listed as a priority pollutant by the US Environmental Protection Agency (EPA) because of its putative carcinogenic effects.
Recently several Governments introduced regulations on the use of phytosanitary products in the agri-food sector and the presence of chemical residues on final products (Maximum Level Residues, MLR), such as the European directive 2009/128/CE (http://data.europa.eu/eli/dir/2009/128/o, accessed on 21 December 2021) that was deliberated with the aim to mitigate the adverse effects of the agricultural practices and promote the implementation of safer approaches in the European Union. As a consequence, some fungicides used to counteract Colletotrichum spp. have been banned [68].
Restrictions vary among countries in both the categories of active compounds regulated and the MLRs allowed [69]. In this context, farmers of exporter countries importing in countries with higher standards incur in great investments for the adjustment of their productive systems, such as expensive inputs and specialized human capital. According to Fiankor et al. [69], many producers not able to support the expenses for this renovation process are bounded to exit the market.
On the basis of these considerations, it emerges the urgency of a transition toward innovative, effective, and sustainable strategies; indeed, over the years many efforts have been made worldwide to find alternative tools to reduce the impact of the traditional fungicides. In some cases, the research has been oriented toward antifungal active substances with low toxicity profiles, such as fludioxonil [66,67], but scientific investigations have mainly focused on the development of chemical-free managing approaches and technologies.
3. Innovative and Sustainable Approaches
3.1. Biological Strategies
3.1.1. Biological Control
Biocontrol-based approaches can represent a valid strategy to improve sustainability in agriculture. These techniques are based on the antagonistic activity exerted by bacterial, fungal or yeast species that can be used to develop non-polluting commercial formulates. The mechanisms by which they act are various and include space and/or nutrient competition, parasitism, and production of toxic metabolites [70].
Post-harvest management of anthracnose based on biological control strategies are available both in the field, to prevent infections, and after the harvesting, to limit the pathogen development [70,71,72,73].
In recent years the efficacy of this approach has been largely explored, finding several biological agents (or antifungal compounds extracted from them) potentially suitable to counteract Colletotrichum spp. infection or development in susceptible fruits (Table 1).
Although many species are responsible for consistent damages, investigations were focused mainly on C. acutatum and C. gloeosporioides species complexes and C. truncatum [73,85,86,87]. In this regard, some studies revealed the strong efficacy of bacterial species, such as some strains of Bacillus subtilis and Paenibacillus polymyxa, and their derivates to reduce both the incidence of anthracnose caused by C. acutatum and/or C. gloeosporioides species complex and the severity of the post-harvest lesions [73,74,75,76]. In particular, Bacillus spp. were shown as prominent candidates for the biological control of these pathogen species in various host species since capable to highly reduce anthracnose incidence (from 76% to 83%) and severity (from 65% to 85%) [74,75].
Moreover, Streptomyces philanti has been suggested as a promising tool for the biological control of C. gloeosporioides species complex, since treatments with volatile compounds produced by this species resulted in a complete absence of symptoms in chili fruits [77]. Some strains of Burkholderia spp. and Pseudomonas aeruginosa exhibited instead a strong efficacy for the control of C. truncatum in chili, as reported by Sandani et al. [78] that revealed a reduction of anthracnose incidence ranging from 75% to 100%.
Moreover, various filamentous fungi have been suggested as efficient biological tools for post-harvest anthracnose prevention; in this case, the antagonistic activity can be also due to the capability of these microorganism to colonize Colletotrichum hyphae [73]. With respect to this, Trichoderma spp. have been shown very effective [79,80,81]; indeed, investigations conducted on strawberry [79] and banana [81] revealed a great efficacy of these fungi to reduce anthracnose incidence (−51%) and severity (−88%) respectively. Furthermore, Oliveri et al. pointed out also that the transformation of citrus plants with a T. harzianum gene, encoding an antifungal protein, resulted in decreased anthracnose symptoms in fruits inoculated with C. gloeosporioides species complex [80].
Finally, also various yeast species were shown to be excellently efficient Colletotrichum antagonists [73], as in the case of Metchnikowia pulcherrima and Pichia kluyveri, more effective than any other tested bacterial or fungal species in contrasting C. acutatum species complex infections in apple fruits [82,83,84]. Here, the biological control exerted by these species allowed a 100% reduction of anthracnose incidence. The efficacy of yeast species against Colletotrichum infections was even reported in some field trials. For instance, the application of a Rhodotorula minuta suspension of a mango orchard was revealed even more effective of chemicals in controlling anthracnose disease [86].
Despite this scientific evidence, to date, only two microorganism-based biofungicides against Colletotrichum spp. are commercially available, both containing bacterial strains belonging to the genus Bacillus [73]. The first one is Serenade ASO (composed by B. amyloliquefaciens QST713, former classified as B. subtilis QST713), whereas the other is Double Nickel 55 (containing B. amyloliquefaciens D747), commercialized by Bayer and Certis respectively. Notably, whereas Serenade ASO has been registered even in the European Union (EU), Double Nickel 55 is currently not commercially available in the EU (while its commercialization has been authorized in other countries such as Canada and USA). However, as reported above and underlined by Shi et al. [73], other microorganisms perform greatly in anthracnose control, suggesting that more effort is needed to provide farmers with more biological tools.
3.1.2. Plant Derivates
The research of eco-friendly methods to manage anthracnose disease has been often oriented toward plant derivates with direct antifungal activity (e.g., secondary metabolites involved in the plant immunity mechanisms).
Over the years, the potential of several plant-based substances against Colletotrichum spp. has been examined, by research institutions in countries where anthracnose represents a severe economic threat. However, many studies have limited their investigations to the inhibitory activity exerted by some crude vegetal extracts on cultured fungi without exploring their efficacy against anthracnose on fruits i.e., [88,89,90,91,92]. Therefore, their possible application in post-harvest managing strategies is currently scarcely known. Moreover, these substances need to be carefully evaluated not only to define their fungicide potential but also to exclude any undesirable adverse effects of their use. With respect to this, Bordoh et al. [92] pointed out a dose-dependent phytotoxicity and a deterioration of some organoleptic features in dragon fruits following treatments with two crude ginger extracts that were found effective against C. gloeosporioides species complex.
Research conducted on papaya fruits revealed that extracts purified from Vitex mollis pulp can exert enhanced antifungal activity compared to the crude ones suggesting that partitioned fractions of plant derivates can be preferable for pathogen management [93]. A class of purified extracts that is receiving increasing attention as fungicide alternatives is represented by essential oils (EOs) from several aromatic plants [94]. Indeed, EOs exhibit a strong antifungal activity related mainly to two categories of lipid active compounds: terpenoids and phenylpropanoids, capable to interact with hydrophobic components of the pathogen membrane [95,96,97,98]. EOs are known from ancient times for their antimicrobial activity, and in recent years, they have been largely explored to assess their suitability to replace chemicals in agriculture.
The effectiveness of some of these substances for the post-harvest management of anthracnose has been deeply studied and shown effective for different fruits [97] (Table 2).
Investigations carried out on papaya fruits showed that the development of C. gloeosporioides species complex is affected by different EOs such as lemongrass, ginger, savory and thyme oils [99,100,101]; in particular, the latter was shown the most effective to reduce the anthracnose incidence (−26.5%) [101].
Savory and thyme oils have been demonstrated effective in limiting C. gloeosporioides species complex growth also in avocado fruits [102] where both the incidence and the severity of the disease decreased by the application of the EOs; whereas in mango, only thyme oils showed to act against these species [103]. EOs extracted from thyme, as well as cinnamon bark oils, were also efficient in reducing the incidence of anthracnose disease caused by C. acutatum species complex on strawberry fruits [104]. On the contrary, C. musae was shown to be sensitive to the application of EOs extracted from Ocimum basilicum and Ocimum gratissimum with reduced anthracnose severity on banana fruits [105].
Nevertheless, also for this class of compounds, knowledge on undesirable effects of EOs is scarce although fundamental. For example, Ali et al. [99] found a dose-dependent phytotoxicity on papaya fruits treated with lemongrass oils.
On the other hand, the suitability of various formulations to confer stability and durability to EOs, which display high volatility and hydrophobicity has received great attention. Nanotechnology-based methods, such as EO encapsulation, incorporation into edible or biodegradable coatings, and development of microemulsions, have been suggested as promising candidates to achieve stabilized formulates [96,106,107,108,109,110,111,112].
Among the plant active compounds explored as alternative solutions for crop protection, several algal derivates have been found to act as plant resistance inducers against biotic and abiotic stressors. Seaweed-based commercial formulates have been developed for young seedling immersions or foliar (high/low-pressure) spraying treatments. In addition, in vitro studies using cultured fungi revealed that these substances can also display inhibiting effects on the mycelial growth and/or the conidial germination of pathogenic agents [113]; evidence for the efficacy of some algae extracts has been achieved also against Colletotrichum spp. For instance, BDDE [Bis(2,3-dibromo-4,5-dihydroxybenzyl) ether], derived from Leathesia nana, Rhodomela confervoides, and Rhodomela confervoides, was found effective in reducing the mycelial growth of C. gloespoiriodes species complex in vitro [114]; similarly, aqueous and ethanolic fractions extracted from the species Sargassum myricocystum and Gracilaria edulis exhibited a similar inhibitory activity against C. falcatum [115]. Conversely, the germination of C. lindemunthianum was reduced by a protein fraction extracted from Hypnea musciformis [116].
However, as for plant derivates and EOs, in vivo studies are necessary to move from laboratory tests to application in anthracnose management protocols. For example, “Ulvan”, a water-soluble polysaccharidic extract achieved from Ulva spp., was shown very effective as a resistance inducer in bean plants, resulting in a consistent (up to 60%) reduction of severity of C. lindemuthianum anthracnose [117,118]. An ethanolic fraction of the same species applied through foliar spray or infiltration was instead associated to an increased expression of defense gene markers in alfalfa specimens and to a subsequent enhanced resistance against C. trifolii infections [119]. Similarly, Kim et al. [120] pointed out that foliar treatments of cucumber plants with a Chlorella fusca suspension efficiently triggered the endogenous defense mechanisms, leading to a reduced severity of anthracnose caused by C. orbicolare. Finally, the progression of anthracnose caused by C. acutatum species complex on strawberry leaves was consistently limited through spray treatments with an Ascophillum nodosum-based biofungicide [121] (Table 3).
In general, the introduction of plant-based compounds into crop defense strategies, requires the fulfillment of a number of constraints besides the lack of adverse effects and the development of appropriate delivery techniques. Indeed, to obtain the authorization from the regulatory authorities, a complex dossier with data on their stability—presently not available for the majority of these compounds—must be developed [93,97,122]. In addition, the activity of these compounds seems to be not enough reproducible, mostly because of the wide variability of their chemical profile [97,98,123]. Therefore, despite the promising results achieved, the introduction of these substances into the market seems still far off.
3.2. Biotechnology-Based Strategies
In recent years, innovative biotechnology-based methodologies for pathogen and pest management have been considered as tools to increase plant resistance against pathogens or to develop new molecules alternative to pesticides.
One promising approach is genome editing that allows achieving new resistant varieties. This technique relies on the production of breaks in specific sites of the plant genomic DNA that are subsequently restored by different cell repairing mechanisms; through the process, single mutations (insertions, deletions or substitutions) can be introduced into the target loci [124]. Meganucleases (MNs), zinc finger nucleases (ZFNs), transcription activator-like effector nucleases (TALENs), and clustered regularly interspaced short palindrome repeats protein 9 (CRISPR-Cas9) are four types of nucleases that can be used in genome editing for DNA break production at specific locations. The great advantage of this technique, included in the so called “New Breeding Technologies” (NBT), is given by both precise and specific genomic modifications. These can target plant genetic traits associated with susceptibility to pathogens in order to strengthen the plant immune response. Compared to traditional breeding methods, these approaches greatly reduce the time required for developing new varieties with the traditional breeding technologies.
On the other hand, the application of this approach can be limited by the scarce knowledge on the genetic mechanisms regulating plant response to pathogens, which, in case of Colletotrichum are not totally uncovered. To date, the only scientific evidence on the efficacy of NBT application against this disease was reported by Mishra and colleagues [125] who developed CRISPR-Cas9 T-DNA-free homozygous pepper plants (Capsicum annum) harboring a desired mutation in the CaERF28 gene. The expression of this gene leads to the downregulation of the endogenous defense mechanisms [126,127] and transformed plants were significantly less susceptible to C. truncatum.
3.2.1. Bidirectional Cross-Kingdom RNA Interference
Among the biotech-based technologies, RNA interference-based strategies have been more extensively explored than genome editing as putative next-generation tools for a sustainable crop protection.
RNA interference (RNAi) is a regulatory system of post-transcriptional gene silencing widely conserved among Eucaryotes and involved in several processes including host immunity, pathogen virulence, and host–pathogen communication [128,129,130,131,132,133,134]. Besides the pathogenesis processes, RNAi in fungi is engaged in multiple functions such as the control of transposable elements, regulation of endogenous gene transcription, heterochromatin formation, maintenance of genome stability, and adaption to stressful conditions [135].
The RNAi mechanism is mediated by small (21–26 nucleotides) RNA (sRNA) molecules with sequence complementarity to transcripts encoded by the target genes. sRNAs are obtained from the cleavage of longer double-stranded RNA (dsRNA) molecules, performed by Dicer or Dicer-like (DCL) proteins or sRNA-specific RNase III family enzymes. One strand of the cleaved dsRNA, so called the “guide” strand, is then incorporated into the RNA-induced silencing complex (RISC) by binding to the Argonaute (AGO) protein, while the second strand is degraded. The guide strand acts as a probe for the specific recognition of target transcripts enabling their cleavage by the RISC complex and preventing transcription [136,137,138].
In the plant kingdom, the RNAi mechanism plays an important role in defense against biotic stressors, downregulating endogenous genes involved in the host susceptibility [134]. In addition, it has been revealed that during the infection process, plant sRNAs can be transferred to pathogens in order to silence genes critical for their growth or virulence; similarly, pathogens can enhance their virulence by transferring to sRNA targeting host defense genes [133]. Altogether, this sRNA-based communication process, first identified in plant-fungi interactions [139], has been called “Bidirectional cross-kingdom RNAi” [131,132,139,140,141] and its discovery has represented a milestone for inspiring new strategies for crops protection [131,132,140,141]. Recent investigations on the mechanisms of sRNAs uptake by fungal pathogens from plant cells indicate that sRNA molecules are delivered from plants through exosomes or extracellular vesicles that are absorbed by fungal cells through endocytic fusion with the cell membrane [142,143,144,145].
RNAi thus opened up a new promising way to develop efficient, specific, and safe crop protection strategies against pests and pathogens. A fundamental prerequisite for its application is the presence of the RNAi machinery in the target organisms, not found in all the fungal pathogens species [135,145,146,147,148,149].
RNAi-based protection can be conferred in two ways: (i) host-induced gene silencing (HIGS) and (ii) spray-induced gene silencing (SIGS). Both ways can represent a valid alternative to other strategies traditionally adopted in the agri-food sector, however, some issues remain to be addressed for their field application (Figure 3).
3.2.2. Host-Induced Gene Silencing Approaches
The HIGS strategy is based on the development of genetically modified crop plants expressing sRNAs (or longer dsRNAs, processed into sRNAs by the RNAi machinery) [150,151]. These sRNAs specifically target pathogen key genes arresting the infection processes and conferring a stable protection of the host [139,152,153,154]. Alternatively, sRNAs can be designed to silence plant genes encoding susceptibility factors, i.e., proteins preventing pathogen recognition and the host defense response [141,155].
This strategy has been demonstrated to be effective to protect crops from a large variety of pathogens and pests, including viruses, viroids, insect, and nematodes [156,157,158]. In 2017, the U.S. Environmental Protection Agency (EPA) approved transformed corn produced by Bayer by means of the SmartStax® PRO technology which make plants capable to express dsRNA molecules targeting Western corn rootworm transcripts [156,159].
HIGS technologies has been used in different crops to control a variety of fungal species as shown by several scientific reports [151,160,161,162,163,164,165]. Among these, Mahto et al. published in 2020 [166] the first evidence for the control of anthracnose in chilly and tomato. Here, the gene encoding the Conidial Morphology 1 protein (COM1), homolog to a Magnaporthe oryzae gene reported to have a crucial role in the conidium morphology and appressorium formation [167], was chosen as a target to control C. gloeosporioides species complex. Agrobacterium-mediated transformation of tomato and chilli plants with a C. gloeosporioides CgCOM1 sRNA expressing cassette resulted in a significant reduction of anthracnose symptoms in both leaves and fruit. These results were corroborated by microscopy analysis indicating the fungal growth inhibition and impairing the production of functional appressoria.
Despite the promising results achieved, the development of a HIGS strategy requires high investments in terms of time and cost necessary to generate stably transformed plants. Moreover, HIGS implies the generation of GMO plants, the use of which is limited by social concerns and national regulations.
3.2.3. Spray-Induced Gene Silencing Approaches
SIGS technique is based on the exogenous application of dsRNAs targeting key transcripts of pathogens, and for this probably represents a more promising candidate to replace agrochemicals [141,153,156,168]. In recent years, several reports showed the efficacy of SIGS technologies for controlling fungal diseases, paving the way for the launch of new GMO-free RNAi-based crop protection strategies [129,163,167,168,169,170].
The efficiency of these techniques relies on dsRNA uptake by the pathogen, which can occur through direct absorption from the environment (the so-called environmental RNAi), or indirectly through the host (cross-kingdom RNAi) [131,132,171,172,173,174,175].
Despite many fungal species have been shown to efficiently uptake RNAs molecules from 21 up to 800 bp, SIGS through environmental RNAi do not work for all fungi, suggesting that some species are reluctant to absorb RNA, possibly due to biochemical feature of cell or membrane wall or the lack of essential RNAi machinery gene or protein component [131,134,169,171]. Thus, the application of this approach requires careful preliminary analysis of the target species.
The development of methodologies suited to increase the stability and persistence of dsRNA molecules in the field is another important requirement for SIGS application, since it is well-known that naked RNA undergoes rapid degradation in the environment. The use of carrier molecules, such as double-layered hydroxide or chitosan/carbon dot-based nanoparticles can be used to stabilize, increase durability, and help plant absorption of RNA [176,177,178,179,180,181,182,183,184].
Despite these aspects, the SIGS strategy remains a promising technology to substitute chemical pesticide application, given both the low toxicological profile of RNA and its low environmental persistency [185,186]. In addition, since RNAi silencing effect is based on the recognition of specific transcript sequences, the risk of the off-target effects is putatively very low. Contrary to the HIGS, the SIGS approach, besides being GMO-free, can be applied also on fruit at post-harvest stage [11,141].
Evidence of successful applications of SIGS against Colletotrichum spp. were first reported by Gu et al. in 2019 [187]. The silencing effect induced by dsRNAs recognizing a fragment of the Fusarium asiaticum gene β2-tubulin (Faβ2tub-3), highly conserved among fungi, was evaluated in different pathogenic species, including C. truncatum, the causal agent of anthracnose on soybean plants, where it efficiently inhibited the spore germination. Cross-kingdom RNAi was the mechanism suggested to be involved there, since the preliminary spray of dsRNA on susceptible host tissues, followed by later pathogen inoculation, led to conidial germination and mycelial growth inhibition.
A functional RNAi machinery was recently reported for C. abscissum species [149], the causal agent of anthracnose on citrus fruits, whose growth was heavily impaired when a fungal strain was transformed so to express dsRNA targeting succinate dehydrogenase transcription. Conidial suspensions of mutant fungi were highly affected in germination and appressorium formation. Furthermore, the fungal inoculum did not determine relevant symptoms when infected into susceptible citrus tissues.
As stated above, SIGS strategies for crop protection can exploit but also the environmental RNAi process that implies the pathogen capability of external RNA uptake. On the other hand, when approached as environmental RNAi, SIGS was not an effective strategy to control C. gloeosporioides species complex, since fungal cells were not capable to uptake fluorescein-labelled dsRNA even after 20 h from the treatments [171]. Consistently, the topical application of dsRNA targeting the DCL gene of this species did not reduce the symptom occurrence in susceptible plant tissues, supporting the lack of dsRNA uptake ability in this species [153].
4. Concluding Remarks
The increased social concern about food safety and the environmental impact of the agri-food system, together with the forthcoming restrictions on the use of phytosanitary products, encouraged the research of innovative strategies alternative to toxic chemicals for crop protection. In this review the most promising strategies under investigation to control anthracnose in postharvest disease management are described. A number of issues must be addressed to allow their application in large-scale production systems. With respect to biocontrol agents, for instance, several investigations showed that a multitude of microorganisms are potentially suitable for anthracnose control; however, for the majority of cases, the antagonistic activity was explored only on few fruit systems and more studies are needed for developing commercial preparations [73]. In addition, their safety, absence of toxic effects and stability (i.e., capability of adhering and colonizing plant tissues, survival under adverse conditions, broad spectrum of action) should also be carefully addressed [70,73,188,189,190].
As about plant derivates, their antifungal activity was tested mostly on cultured fungi, whereas tests in relevant applicative environment are missing. Several EOs showed good efficacy to control anthracnose; however, further research is necessary to exclude phytotoxic effect on produces, to fulfil the requirements needed, for authorization and develop appropriate methodologies for their delivery. Nanotechnology is currently the more promising strategy to enhance their persistence and bioavailability in the environment taking into account also the economic and environmental sustainability of delivery formulations [97,181].
New strategies for Colletotrichum spp. management have been proposed also through biotechnology-based approaches, such as those exploiting the RNA interference mechanism (Table 4).
Despite some promising results recently achieved [166] and the putative environmental safety of these methodologies, RNAi-based approaches still need to meet a number of technological, legal, and social issues. For instance, HIGS strategy is achieved through the development of GMO crops expressing interfering RNAs with all the regulatory and social concerns that this implies, although here plant transformation does not lead to synthesis of new proteins but to sRNA molecules acting in a sequence-specific manner [190,191,192,193,194,195]. Furthermore, for many susceptible crops fast and cost-effective transformation protocols are not available yet, making it difficult to reach large-scale production of sRNA expressing plants. On the other hand, once these limitations are overcome, this technique can provide stable protection allowing a substantial decrease in the use of toxic agrochemicals, a great advantage especially for fruit producers of developing countries, where productions are often seriously affected by anthracnose and postharvest technologies are not advanced.
GMO-linked constraints do not affect the SIGS strategy, where interfering RNAs are exogenously applied. However, more studies are needed to consolidate knowledge on the RNAi functioning and siRNA uptake mechanisms by Colletotrichum spp., mostly with respect to the different behavior so far reported for different species [149,171,187]. These considerations become particularly important for the postharvest management of fruit crops, where restrictions in the use of agrochemicals and consumers’ attention on fruit safety raise particular concern.
Owing to the sequence recognition mechanism, both RNAi-based approaches are characterized by high specificity toward target pathogens; however, the lack of a multi-spectrum efficacy can represent a limiting factor, especially for protection of crops susceptible to multiple Colletotrichum species. For this reason, the target gene/sequence selection process should be focused on the detection of regions efficient for the silencing of discrete related pathogen species.
Author Contributions
Writing—original draft preparation, A.C. and F.N.; writing—review and editing, all authors; supervision, E.B. and R.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Talhinhas, P.; Baroncelli, R. Colletotrichum species and complexes: Geographic distribution, host range and conservation status. Fungal Divers. 2021, 110, 109–198. [Google Scholar] [CrossRef]
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [Green Version]
- Bordoh, P.K.; Ali, A.; Dickinson, M.; Siddiqui, Y.; Romanazzi, G. A review on the management of postharvest anthracnose in dragon fruits caused by Colletotrichum spp. Crop Prot. 2020, 130, 105067. [Google Scholar] [CrossRef]
- Freeman, S.; Katan, T.; Shabi, E. Characterization of Colletotrichum species responsible for anthracnose in various fruits. Plant Dis. 1998, 82, 596–605. [Google Scholar] [CrossRef] [Green Version]
- Tripathy, A.P.; Satapathy, R.R.; Rout, M. Management of Colletotrichum gloeosporioides causing papaya anthracnose with best resulting plant extract and bio-agents. Pharma Innov. J. 2021, 10, 629–630. [Google Scholar]
- Banya, M.; Garg, S.; Lal Meena, N. A review: Chilli anthracnose, its spread and management. J. Pharmacogn. Phytochem. 2020, 9, 1432–1438. [Google Scholar]
- Wharton, P.S.; Diéguez-Uribeondo, J. The biology of Colletotrichum acutatum. An. Jard. Bot. Madr. 2004, 61, 3–22. [Google Scholar] [CrossRef]
- Jeffries, P.; Dodd, J.C.; Jeger, M.J.; Plumbey, R.A. The biology and control of Colletotrichum species on tropical fruit crops. Plant Pathol. 1990, 39, 343–366. [Google Scholar] [CrossRef]
- Prusky, D.; Kobiler, I.; Ardi, R.; Beno-Moalem, D.; Yakoby, N.; Keen, N.T. Resistance mechanisms of subtropical fruits to Colletotrichum gloeosporioides. In Colletotrichum: Host Specificity, Pathology, and Host-Pathogen Interaction; Prusky, D., Freeman, S., Dickman, M.B., Eds.; The American Phytopathological Society Press: St. Paul, MN, USA, 2000; pp. 232–244. [Google Scholar]
- Nazzaro, F.; Fratianni, F.; Coppola, R.; De Feo, V. Essential oils and antifungal activity. Pharmaceuticals 2017, 10, 86. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira Filho, J.G.; da Cruz Silva, G.; Cipriano, L.; Gomes, M.; Buranelo Egea, M. Control of postharvest fungal diseases in fruits using external application of RNAi. J. Food Sci. 2021, 86, 3341–3348. [Google Scholar] [CrossRef]
- Van der Bruggen, P.; Maraite, H. Histopathology of cassava anthracnose disease caused by Colletotrichum gloeosporioides f. sp. manihotis. Parasitica 1987, 43, 3–21. [Google Scholar]
- Zulfiqar, M.; Brlansky, R.H.; Timmer, L.W. Infection of flower and vegetative tissues of citrus by Colletotrichum acutatum and C. gloeosporioides. Mycologia 1996, 88, 121–128. [Google Scholar] [CrossRef]
- Latunde-Dada, A.O.; O’Connell, R.J.; Nash, C.; Lucas, J.A. Stomatal penetration of cowpea (Vigna unguiculata) leaves by a Colletotrichum species causing latent anthracnose. Plant Pathol. 1999, 48, 777–784. [Google Scholar] [CrossRef]
- Singh, D.; Sharma, R.R. Postharvest disinfection of fruits and vegetables and their management. In Postharvest Disinfection of Fruits and Vegetables, 1st ed.; Siddiqui, M.W., Ed.; Elsevier: Oxford, UK; pp. 1–52.
- Peres, N.A.; Timmer, L.W.; Adaskaveg, J.E.; Correl, J.C. Lifestyle of Colletotrichum acutatum. Plant Dis. 2005, 89, 784–796. [Google Scholar] [CrossRef] [Green Version]
- Bailey, J.A.; O’Connell, R.J.; Pring, R.J.; Nash, C. Infection strategies of Colletotrichum species. In Colletotrichum: Biology, Pathology and Control; Bailey, J.A., Jeger, M.J., Eds.; CAB International: Wallingford, UK, 1992; pp. 88–120. [Google Scholar]
- Skipp, R.A.; Beever, R.E.; Sharrock, K.R.; Rikkerink, E.H.A.; Templeton, M.D. Histopathological, genetic, Biochemical and molecular basis in Colletotrichum. In Pathogens and Host Parasite Specificity in Plant Disease; Singh, U.S., Kohmoto, K., Singh, R.P., Eds.; Elsevier Science Ltd.: Oxford, UK, 1995; pp. 119–143. [Google Scholar]
- O’Connell, R.J.; Bailey, J.A.; Richmond, D.V. Cytology and physiology of infection of Phaseolus vulgaris by Colletotrichum lindemuthianum. Physiol. Mol. Plant Pathol. 1985, 27, 75–98. [Google Scholar] [CrossRef]
- Latunde-Dada, A.O.; O’Connell, R.J.; Nash, C.; Pring, R.J.; Lucas, J.A.; Bailey, J.A. Infection process and identity of the hemibiotrophic anthracnose fungus (Colletotrichum destructivum) from cowpea (Vigna unguiculata). Mycol. Res. 1996, 100, 1133–1141. [Google Scholar] [CrossRef]
- Wharton, P.S.; Julian, A.M. A cytological study of compatible and incompatible interactions between Sorghum bicolor and Colletotrichum sublineolum. N. Phytol. 1996, 134, 25–34. [Google Scholar] [CrossRef]
- Wharton, P.S.; Julian, A.M.; O’Connell, R.J. Ultrastructure of the infection of Sorghum bicolor by Colletotrichum sublineolum. Phytopathology 2001, 91, 149–158. [Google Scholar] [CrossRef] [Green Version]
- Arauz, L.F. Mango anthracnose: Economic impact and current option for integrated management. Plant Dis. 2000, 84, 600–611. [Google Scholar] [CrossRef] [Green Version]
- O’Connell, R.J.; Thon, M.R.; Hacquard, S.; Amyotte, S.G.; Kleemann, J.; Torres, M.F.; Damm, U.; Buiate, E.; Epstein, L.; Alkan, N.; et al. Lifestyle transitions in plant pathogenic Colletotrichum fungi deciphered by genome and transcriptome analyses. Nat. Genet. 2012, 44, 1060–1065. [Google Scholar] [CrossRef]
- Prusky, D.; Barad, S.; Ment, D.; Bi, F. The pH modulation by fungal secreted molecules: A mechanism affecting pathogenicity by postharvest pathogens. Isr. J. Plant Sci. 2016, 63, 22–30. [Google Scholar] [CrossRef]
- Alkan, N.; Friedlander, G.; Ment, D.; Prusky, D.; Fluhr, R. Simultaneous transcriptome analysis of Colletotrichum gloeosporioides and tomato fruit pathosystem reveals novel fungal pathogenicity and fruit defense strategies. N. Phytol. 2015, 205, 801–815. [Google Scholar] [CrossRef]
- Md Nor, S.; Ding, P. Trends and advances in edible biopolymer coating for tropical fruit: A review. Food Res. Int. 2020, 134, 109208. [Google Scholar] [CrossRef]
- Roberts, R.G.; Snow, J.P. Histopathology of cotton boll rot caused by Colletotrichum capsici. Phytopathology 1984, 74, 390–397. [Google Scholar] [CrossRef]
- Pring, R.J.; Nash, C.; Zakaria, M.; Bailey, J.A. Infection process and host range of Colletotrichum capsici. Physiol. Mol. Plant Pathol. 1995, 46, 137–152. [Google Scholar] [CrossRef]
- Smith, B.J. Epidemiology and pathology of strawberry anthracnose: A North American perspective. Hortscience 2008, 43, 69–73. [Google Scholar] [CrossRef] [Green Version]
- Higuera, J.J.; Garrido-Gala, J.; Lekhbou, A.; Arjona-Girona, I.; Amil-Ruiz, F.; Mercado, J.A.; Pliego-Alfaro, F.; Muñoz-Blanco, J.; López-Herrera, C.J.; Caballero, J.L. The strawberry FaWRKY1 transcription factor negatively regulates resistance to Colletotrichum acutatum in fruit upon infection. Front. Plant Sci. 2019, 10, 480. [Google Scholar] [CrossRef]
- Sreenivasaprasad, S.; Talhinhas, P. Genotypic and phenotypic diversity in Colletotrichum acutatum, a cosmopolitan pathogen causing anthracnose on a wide range of hosts. Mol. Plant Pathol. 2005, 6, 361–378. [Google Scholar] [CrossRef]
- Marian, M.; Ohno, T.; Suzuki, H.; Kitamura, H.; Kuroda, K.; Shimizu, M. A novel strain of endophytic Streptomyces for the biocontrol of strawberry anthracnose caused by Glomerella cingulata. Microbiol. Res. 2020, 234, 126428. [Google Scholar] [CrossRef]
- Freeman, S.; Katan, T.; Shabi, E. Characterization of Colletotrichum gloeosporioides isolates from avocado and almond fruits with molecular and pathogenicity tests. Appl. Environ. Microbiol. 1996, 62, 1014–1020. [Google Scholar] [CrossRef] [Green Version]
- Saxena, A.; Raghuwanshi, R.; Gupta, V.K.; Singh, H.B. Chilli anthracnose: The epidemiology and management. Front. Microbiol. 2016, 7, 1527. [Google Scholar] [CrossRef] [Green Version]
- Fischer, I.; Moraes, M.; Palharini, M.; Cruz, J.; Firmino, A. Ocorrência de antracnose em abacate, agressividade e sensibilidade de Colletotrichum gloeosporioides a fungicidas. Agropecu. Cient. Semiárido 2019, 13, 130–137. [Google Scholar]
- Lakshmi, B.K.M.; Reddy, P.N.; Prasad, R.D. Cross-infection potential of Colletotrichum gloeosporioides Penz. isolates causing anthracnose in subtropical fruit crops. Trop. Agric. Res. 2011, 22, 183–193. [Google Scholar] [CrossRef] [Green Version]
- Maqbool, M.; Ali, A.; Ramachandran, S.; Smith, D.R.; Alderson, P.G. Control of postharvest anthracnose of banana using a new edible composite coating. Crop Prot. 2010, 29, 1136–1141. [Google Scholar] [CrossRef]
- Paull, R.E.; Nishijima, W.; Reyes, M.; Cavaletto, C. Postharvest handling and losses during marketing of papaya (Carica papaya). Postharvest Biol. Technol. 1997, 11, 165–179. [Google Scholar] [CrossRef]
- Darshan, K.; Vanitha, S.; Venugopala, K.M.; Parthasarathy, S. Strategic eco-friendly management of post-harvest fruit rot in papaya caused by Colletotrichum gloeosporioides. J. Biol. Control 2019, 33, 225–235. [Google Scholar] [CrossRef]
- Freeman, S.; Shabi, E. Cross-infection of subtropical and temperature fruits by Colletotrichum species from various hosts. Physiol. Mol. Plant Path. 1996, 49, 395–404. [Google Scholar] [CrossRef]
- Nelson, S.C. Mango anthracnose (Colletotrichum gloeosporioides). Plant Dis. 2008, PD-48. [Google Scholar]
- Botelho, L.N.S.; Rocha, D.A.; Braga, M.A.; Silva, A.; de Abreu, C.M.P. Quality of guava cv. ‘Pedro Sato’ treated with cassava starch and cinnamon essential oil. Sci. Hortic. 2016, 209, 214–220. [Google Scholar] [CrossRef]
- Uluisik, S.; Seymour, G.B. Pectate lyases: Their role in plants and importance in fruit ripening. Food Chem. 2020, 309, 125559. [Google Scholar] [CrossRef]
- Masyahit, M.; Kamaruzaman, S.; Yahya, A.; Ghazali, M. First report of the occurrence of the anthracnose disease caused by Colletotrichum gloeosporioides (Penz.) Penz. & Sacc. on dragon fruit (Hylocereus spp.) in Peninsular Malaysia. Am. J. Appl. Sci. 2009, 6, 902–912. [Google Scholar]
- Deng, L.; Zeng, K.; Zhou, Y.; Huang, Y. Effects of postharvest oligochitosan treatment on anthracnose disease in citrus (Citrus sinensis L. Osbeck) fruit. Eur. Food Res. Technol. 2015, 240, 795–804. [Google Scholar] [CrossRef]
- Martínez-Blay, V.; Pérez-Gago, M.B.; de la Fuente, B.; Carbó, R.; Palou, L. Edible coatings formulated with antifungal GRAS salts to control citrus anthracnose caused by Colletotrichum gloeosporioides and preserve postharvest fruit quality. Coatings 2020, 10, 730. [Google Scholar] [CrossRef]
- Lum, M.S.; Norazira, M.A. Effects of hot water, submergence time and storage duration on quality of dragon fruit (Hylocereus polyrhizus). J. Agric. Sci. 2011, 3, 146. [Google Scholar] [CrossRef] [Green Version]
- Maringgal, B.; Hashim, N.; Tawakkal, I.S.M.A.; Mohamed, M.T.M.; Hamzah, M.H.; Shukor, N.I.A. The causal agent of anthracnose in papaya fruit and control by three different Malaysian stingless bee honeys, and the chemical profile. Sci. Hortic. 2019, 257, 108590. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, W.; Li, C.; Shao, T.; Jiang, X.; Zhao, H.; Ai, W. Postharvest hot water dipping and hot water forced convection treatments alleviate chilling injury for zucchini fruit during cold storage. Scientia Hortic. 2019, 249, 219–227. [Google Scholar] [CrossRef]
- Sanders, G.M.; Korsten, L.; Wehner, F.C. Survey of fungicide sensitivity in Colletotrichum gloeosporioides from different avocado and mango production areas in South Africa. Eur. J. Plant Pathol. 2000, 106, 745–752. [Google Scholar] [CrossRef]
- Chung, W.H.; Ishii, H.; Nishimura, K.; Fukaya, M.; Yano, K.; Kajitani, Y. Fungicide sensitivity and phylogenetic relationship of anthracnose fungi isolated from various fruit crops in Japan. Plant Dis. 2006, 90, 506–512. [Google Scholar] [CrossRef] [Green Version]
- Inada, M.; Ishii, H.; Chung, W.H.; Yamada, T.; Yamaguchi, J.; Furuta, A. Occurrence of strobilurin-resistant strains of Colletotrichum gloeosporioides (Glomerella cingulata), the causal fungus of strawberry anthracnose. Jpn. J. Phytopathol. 2008, 74, 114–117. [Google Scholar] [CrossRef] [Green Version]
- Kikuchi, M.; Ogawara, T.; Hashimoto, Y.; Miyamoto, T.; Kaneda, M.; Tomita, Y. Identification of Colletotrichum species causing strawberry anthracnose and distribution of fungal strains resistant to some fungicides in Ibaraki prefecture. Bull. Hortic. Inst. Ibaraki Agric. Cent. 2010, 17, 35–42. [Google Scholar]
- Cannon, P.F.; Damm, U.; Johnston, P.R.; Weir, B.S. Colletotrichum—Current status and future directions. Stud. Mycol. 2012, 73, 181–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamichhane, J.R.; Dachbrodt-Saaydeh, S.; Kudsk, P.; Messean, A. Conventional pesticides in agriculture: Benefits versus risks. Plant Dis. 2016, 100, 10–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.; Wu, D.; Zhang, Q.; Chen, H.; Li, H.; Han, Q.; Lai, X.; Wang, H.; Wu, Y.; Yuan, J.; et al. Efficacy and mechanism of cinnamon essential oil on inhibition of Colletotrichum acutatum isolated from “Hongyang” kiwifruit. Front. Microbiol. 2018, 9, 1288. [Google Scholar] [CrossRef] [Green Version]
- Chechi, A.; Stahlecker, J.; Dowling, M.E.; Schnabel, G. Diversity in species composition and fungicide resistance profiles in Colletotrichum isolates from apples. Pestic. Biochem. Phys. 2019, 158, 18–24. [Google Scholar] [CrossRef]
- Kimaru, K.S.; Muchemi, K.P.; Mwangi, J.W. Effects of anthracnose disease on avocado production in Kenya. Cogent Food Agric. 2020, 6, 1799531. [Google Scholar] [CrossRef]
- Zhang, L.; Song, L.; Xu, X.; Zou, X.; Duan, K.; Gao, Q. Characterization and fungicide sensitivity of Colletotrichum species causing strawberry anthracnose in Eastern China. Plant Dis. 2020, 104, 1960–1968. [Google Scholar] [CrossRef] [PubMed]
- Yuan, G.; Chen, X.; Li, D. Chitosan films and coatings containing essential oils: The antioxidant and antimicrobial activity, and application in food systems. Food Res. Int. 2016, 89, 117–128. [Google Scholar] [CrossRef]
- Yokosawa, S.; Eguchi, N.; Kondo, K.I.; Sato, T. Phylogenetic relationship and fungicide sensitivity of members of the Colletotrichum gloeosporioides species complex from apple. J. Gen. Plant Pathol. 2017, 83, 291–298. [Google Scholar] [CrossRef]
- Yan, X.; Li, H.; Su, X. Review of optical sensors for pesticides. Trends Analyt. Chem. 2018, 103, 1–20. [Google Scholar] [CrossRef]
- Craddock, H.A.; Huang, D.; Turner, P.C.; Quiros-Alcala, L.; Payne-Sturges, D.C. Trends in neonicotinoid pesticide residues in food and water in the United States, 1999–2015. Environ. Health 2019, 18, 7. [Google Scholar] [CrossRef] [Green Version]
- Ncama, K.; Mditshwa, A.; Tesfay, S.Z.; Mbili, N.C.; Magwaza, L.S. Topical procedures adopted in testing and application of plant-based extracts as bio-fungicides in controlling postharvest decay of fresh produce. Crop Prot. 2019, 115, 142–151. [Google Scholar] [CrossRef]
- Diskin, S.; Sharir, T.; Feygenberg, O.; Maurer, D.; Alkan, N. Fludioxonil—A potential alternative for postharvest disease control in mango fruit. Crop Prot. 2019, 124, 104855. [Google Scholar] [CrossRef]
- Shimshoni, J.A.; Bommuraj, V.; Chen, Y.; Sperling, R.; Barel, S.; Feygenberg, O.; Maurer, D.; Alkan, N. Postharvest fungicide for avocado fruits: Antifungal efficacy and peel to pulp distribution kinetics. Foods 2020, 9, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lokare, P.; Fatima, S.; Jagdale, P.E. A review on the management practices of Colletotrichum gloeosporioides (Penz.) Penz. & Sacc. causes anthracnose disease of mango. Int. J. Botany Stud. 2021, 6, 742–746. [Google Scholar]
- Fiankor, D.-D.D.; Curzi, D.; Olper, A. Trade, price and quality upgrading effects of agri-food standards Eur. Rev. Agric. Econ. 2021, 48, 835–877. [Google Scholar] [CrossRef]
- Carmona-Hernandez, S.; Reyes-Pérez, J.J.; Chiquito-Contreras, R.G.; Rincon-Enriquez, G.; Cerdan-Cabrera, C.R.; Hernandez-Montiel, L.G. Biocontrol of postharvest fruit fungal diseases by bacterial antagonists: A review. Agronomy 2019, 9, 121. [Google Scholar] [CrossRef] [Green Version]
- Droby, S.; Wisniewski, M.; Teixido, N.; Spadaro, D.; Jijakli, M.H. The science, development, and commercialization of postharvest biocontrol products. Postharvest Biol. Technol. 2016, 122, 22–29. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Mahunu, G.K.; Castoria, R.; Yang, Q.Y.; Apaliya, M.T. Recent developments in the enhancement of some postharvest biocontrol agents with unconventional chemicals compounds. Trends Food Sci. Technol. 2018, 78, 180–187. [Google Scholar] [CrossRef]
- Shi, X.-C.; Wang, S.-Y.; Duan, X.C.; Wang, Y.-Z.; Liu, F.-Q.; Laborda, P. Biocontrol strategies for the management of Colletotrichum species in postharvest fruits. Crop Prot. 2021, 141, 105454. [Google Scholar] [CrossRef]
- Arroyave-Toro, J.J.; Mosquera, S.; Villegas-Escobar, V. Biocontrol activity of Bacillus subtilis EA-CB0015 cells and lipopeptides against postharvest fungal pathogens. Biol. Control 2017, 114, 195–200. [Google Scholar] [CrossRef]
- Lee, G.-W.; Ko, J.-A.; Oh, B.-T.; Choi, J.-R.; Lee, K.-J.; Chae, J.-C.; Kamala-Kannan, S. Biological control of postharvest diseases of apples, peaches and nectarines by Bacillus subtilis S16 isolated from halophytes rhizosphere. Biocontrol Sci. Technol. 2012, 22, 351–361. [Google Scholar] [CrossRef]
- Kim, Y.S.; Balaraju, K.; Jeon, Y. Biological control of apple anthracnose by Paenibacillus polymyxa APEC128, an antagonistic rhizobacterium. Plant Pathol. J. 2016, 32, 251–259. [Google Scholar] [CrossRef] [Green Version]
- Boukaew, S.; Petlamul, W.; Bunkrongcheap, R.; Chookaew, T.; Kabbua, T.; Thippated, A.; Prasertsan, P. Fumigant activity of volatile compounds of Streptomyces philanthi RM-1-138 and pure chemicals (acetophenone and phenylethyl alcohol) against anthracnose pathogen in postharvest chili fruit. Crop Protect. 2018, 103, 1–8. [Google Scholar] [CrossRef]
- Sandani, H.B.P.; Ranathunge, N.P.; Lakshman, P.L.N.; Weerakoon, W.M.W. Biocontrol potential of five Burkholderia and Pseudomonas strains against Colletotrichum truncatum infecting chilli pepper. Biocontrol Sci. Technol. 2019, 29, 727–745. [Google Scholar] [CrossRef]
- Freeman, S.; Minz, D.; Kolesnik, I.; Barbul, O.; Zveibil, A.; Maymon, M.; Nitzani, Y.; Kirshner, B.; Rav-David, D.; Bilu, A.; et al. Trichoderma biocontrol of Colletotrichum acutatum and Botrytis cinerea and survival in strawberry. Eur. J. Plant Pathol. 2004, 110, 361–370. [Google Scholar] [CrossRef]
- Oliveri, C.; Distefano, G.; La Malfa, S.; La Rosa, R.; Deng, Z.N.; Gentile, A. Lemon fruits from endochitinase transgenic plants exhibit resistance against postharvest fungal pathogens. Acta Hortic. 2015, 1065, 1639–1645. [Google Scholar] [CrossRef]
- Sangeetha, G.; Usharani, S.; Muthukumar, A. Biocontrol with Trichoderma species for the management of postharvest crown rot of banana. Phytopathol. Mediterr. 2009, 48, 214–225. [Google Scholar]
- Conway, W.S.; Leverentz, B.; Janisiewicz, W.J.; Saftner, R.A.; Camp, M.J. Improving biocontrol using antagonist mixtures with heat and/or sodium bicarbonate to control postharvest decay of apple fruit. Postharvest Biol. Technol. 2005, 36, 235–244. [Google Scholar] [CrossRef]
- Janisiewicz, W.J.; Leverentz, B.; Conway, W.S.; Saftner, R.A.; Reed, A.N.; Camp, M.J. Control of bitter rot and blue mold of apples by integrating heat and antagonist treatments on 1-MCP treated fruit stored under controlled atmosphere conditions. Postharvest Biol. Technol. 2003, 29, 129–143. [Google Scholar] [CrossRef]
- Mewa-Ngongang, M.; du Plessis, H.W.; Ntwampe, S.K.O.; Chidi, B.S.; Hutchinson, U.F.; Mekuto, L.; Jolly, N.P. The use of Candida pyralidae and Pichia kluyveri to control spoilage microorganisms of raw fruits used for beverage production. Foods 2019, 8, 454. [Google Scholar] [CrossRef] [Green Version]
- Damm, U.; Sato, T.; Alizadeh, A.; Groenewald, J.Z.; Crous, P.W. The Colletotrichum dracaenophilum, C. magnum and C. orchidearum species complexes. Stud. Mycol. 2019, 90, 71–118. [Google Scholar] [CrossRef] [PubMed]
- Patiño-Vera, M.; Jimenez, B.; Balderas, K.; Ortiz, M.; Allende, R.; Carrillo, A.; Galindo, E. Pilot-scale production and liquid formulation of Rhodotorula minuta, a potential biocontrol agent of mango anthracnose. J. Appl. Microbiol. 2005, 99, 540–550. [Google Scholar] [CrossRef] [PubMed]
- Chung, P.C.; Wu, H.Y.; Wang, Y.W.; Ariyawansa, H.A.; Hu, H.P.; Hung, T.H.; Tzean, S.S.; Chung, C.L. Diversity and pathogenecity of Colletotrichum species causing strawberry anthracnose in Taiwan and description of a new species, Colletotrichum miaoliense sp. nov. Sci. Rep. 2020, 10, 14664. [Google Scholar] [CrossRef] [PubMed]
- Begum, J.; Yusuf, M.; Chowdhury, J.U.; Khan, S.; Anwar, M.N. Antifungal activity of forty higher plants against phytopathogenic fungi. Bangl. J. Microbiol. 2008, 24, 76–78. [Google Scholar] [CrossRef] [Green Version]
- Nduagu, C.; Ekefan, E.J.; Nwankiti, A.O. Effect of some crude plant extracts on growth of Colletotrichum capsici (Synd) & Bisby, causal agent of pepper anthracnose. J. Appl. Biosci. 2008, 6, 184–190. [Google Scholar]
- Johnny, L.; Yusuf, U.; Nulit, R. Antifungal activity of selected plant leaves crude extracts against a pepper anthracnose fungus, Colletotrichum capsici (Sydow) butler and bisby (Ascomycota: Phyllachorales). Afr. J. Biotechnol. 2011, 10, 4157–4165. [Google Scholar]
- Saravanakumar, P.; Karthikeyan, V.; Patharajan, S.; Kannabiran, B. Antifungal activity of Plumbago species against anthracnose fungus Colletotrichum gloeosporioides (Penz.) of chilli. Arch. Phytopathol. Plant Prot. 2011, 44, 287–297. [Google Scholar] [CrossRef]
- Bordoh, P.K.; Ali, A.; Dickinson, M.; Siddiqui, Y. Antimicrobial effect of rhizome and medicinal herb extract in controlling postharvest anthracnose of dragon fruit and their possible phytotoxicity. Sci. Hortic. 2020, 265, 109249. [Google Scholar] [CrossRef]
- López-Velázquez, J.G.; Delgado-Vargas, F.; Ayón-Reyna, L.E.; López-Angulo, G.; Bautista-Baños, S.; Uriarte-Gastelum, Y.G.; López-López, M.E.; Vega-García, M.O. Postharvest application of partitioned plant extracts from Sinaloa, Mexico for controlling papaya pathogenic fungus Colletotrichum gloeosporioides. J. Plant Pathol. 2021, 103, 831–842. [Google Scholar] [CrossRef]
- Raveau, R.; Fontaine, J.; Lounès-Hadj Sahraoui, A. Essential oils as potential alternative biocontrol products against plant pathogens and weeds: A review. Foods 2020, 9, 365. [Google Scholar] [CrossRef] [Green Version]
- Freiesleben, S.; Jager, A.K. Correlation between plant secondary metabolites and their antifungal mechanisms—A review. J. Med. Aromat. Plants 2014, 3, 2167-0412. [Google Scholar]
- Gutierrez-del-Rio, I.; Fernandez, J.; Lombo, F. Plant nutraceuticals as antimicrobial agents in food preservation: Terpenoids, polyphenols and thiols. Int. J. Antimicrob. Agents 2018, 52, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Gonҫalves, D.C.; Rodrigues Ribeiro, W.; Gonҫalves, D.C.; Menini, L.; Costa, H. Recent advances and future perspective of essential oils in control Colletotrichum spp.: A sustainable alternative in postharvest treatment of fruits. Food Res. Int. 2021, 150, 110758. [Google Scholar] [CrossRef]
- Vianna, T.C.; Marinho, C.O.; Junior, L.M.; Ibrahim, S.A.; Vieira, R.P. Essential oils as additives in active starch-based food packaging films: A review. Int. J. Biol. Macromol. 2021, 182, 1803–1819. [Google Scholar] [CrossRef]
- Ali, A.; Wee Pheng, T.; Mustafa, M.A. Application of lemongrass oil in vapour phase for the effective control of anthracnose of “Sekaki” papaya. J. Appl. Microbiol. 2015, 118, 1456–1464. [Google Scholar] [CrossRef]
- Ali, A.; Hei, G.K.; Keat, Y.W. Efficacy of ginger oil and extract combined with gum arabic on anthracnose and quality of papaya fruit during cold storage. J. Food Sci. Technol. 2016, 53, 1435–1444. [Google Scholar] [CrossRef] [Green Version]
- Sarkhosh, A.; Schaffer, B.; Vargas, A.I.; Palmateer, A.J.; Lopez, P.; Soleymani, A.; Farzaneh, M. Antifungal activity of five plant-extracted essential oils against anthracnose in papaya fruit. Biol. Agric. Hort. 2017, 34, 18–26. [Google Scholar] [CrossRef]
- Sarkhosh, A.; Vargas, A.I.; Schaffer, B.; Palmateer, A.J.; Lopez, P.; Soleymani, A.; Farzaneh, M. Postharvest management of anthracnose in avocado (Persea americana Mill.) fruit with plant extracted oils. Food Packag. 2017, 12, 16–22. [Google Scholar] [CrossRef]
- Perumal, A.B.; Sellamuthu, P.S.; Nambiar, R.B.; Sadiku, E.R. Antifungal activity of five different essential oils in vapour phase for the control of Colletotrichum gloeosporioides and Lasiodiplodia theobromae in vitro and on mango. Inter. J. Food Sci. Technol. 2016, 51, 411–418. [Google Scholar] [CrossRef]
- Duduk, N.; Markovic, T.; Vasic, M.; Duduk, B.; Vico, I.; Obradovic, A. Antifungal activity of three essential oils against Colletotrichum acutatum, the causal agent of strawberry anthracnose. J. Essent. Oil Bear. Plants 2015, 18, 529–537. [Google Scholar] [CrossRef]
- Madjouko, M.A.; Tchameni, S.N.; Tchinda, E.S.; Dongmo Jazet, P.M.; Kamsu, P.N.; Medzue Souop Kamga, V.A.; Sameza, M.S.; Tchoumbougnang, F.; Menut, C. Inhibitory effects of essential oils from Ocimum basilicum and Ocimum gratissimum on Colletotrichum musae: The causal agent of bananas anthracnose. J. Phytopath. 2019, 167, 257–264. [Google Scholar] [CrossRef]
- Galus, S.; Kadzińska, J. Food applications of emulsion-based edible films and coatings. Trends Food Sci. Technol. 2015, 45, 273–283. [Google Scholar] [CrossRef]
- Valencia-Sullca, C.; Jiménez, M.; Jiménez, A.; Atarés, L.; Vargas, M.; Chiralt, A. Influence of liposome encapsulated essential oils on properties of chitosan films. Polym. Int. 2016, 65, 979–987. [Google Scholar] [CrossRef]
- Scremin, F.R.; Veiga, R.S.; Silva-Buzanello, R.A.; Becker-Algeri, T.A.; Corso, M.P.; Torquato, A.S.; Bittencourt, P.R.S.; Flores, E.L.M.; Canan, C. Synthesis and characterization of protein microcapsules for eugenol storage. J. Therm. Anal. Calorim. 2016, 131, 653–660. [Google Scholar] [CrossRef]
- Shin, J.; Na, K.; Shin, S.; Seo, S.-M.; Youn, H.J.; Park, I.-K.; Hyun, J. Biological activity of thyme white essential oil stabilized by cellulose nanocrystals. Biomolecules 2019, 9, 799. [Google Scholar] [CrossRef] [Green Version]
- Talón, E.; Vargas, M.; Chiralt, A.; González-Martínez, C. Antioxidant starch-based films with encapsulated eugenol. Application to sunflower oil preservation. LWT 2019, 113, 108290. [Google Scholar] [CrossRef]
- Weisany, W.; Samadi, S.; Amini, J.; Hossaini, S.; Yousefi, S.; Maggi, F. Enhancement of the antifungal activity of thyme and dill essential oils against Colletotrichum nymphaeae by nano-encapsulation with copper NPs. Ind. Crops Prod. 2019, 132, 213–225. [Google Scholar] [CrossRef]
- Wińska, K.; Mączka, W.; Łyczko, J.; Grabarczyk, M.; Czubaszek, A.; Szumny, A. Essential oils as antimicrobial agents—Myth or real alternative? Molecules 2019, 24, 2130. [Google Scholar] [CrossRef] [Green Version]
- Righini, H.; Roberti, R.; Baraldi, E. Use of algae in strawberry management. J. Appl. Phycol. 2018, 30, 3551–3564. [Google Scholar] [CrossRef]
- Liu, M.; Wang, G.; Xiao, L.; Xu, X.; Liu, X.; Xu, P.; Lin, X. Bis (2,3-dibromo-4,5-dihydroxybenzyl) ether, a marine algae derived bromophenol, inhibits the growth of Botrytis cinerea and interacts with DNA molecules. Mar. Drugs 2014, 12, 3838–3851. [Google Scholar] [CrossRef]
- Ambika, S.; Sujatha, K. Antifungal activity of aqueous and ethanol extracts of seaweeds against sugarcane red rot pathogen (Colletotrichum falcatum). Sci. Res. Essays 2015, 10, 232–235. [Google Scholar]
- Melo, V.M.M.; Medeiros, D.A.; Rios, F.J.B.; Castelar, L.I.M.; de, F.F.U. Carvalho, A. Antifungal Properties of Proteins (Agglutinins) from the Red Alga Hypnea musciformis (Wulfen) Lamouroux. Bot. Mar. 1997, 40, 281–284. [Google Scholar] [CrossRef]
- de Freitas, M.B.; Stadnik, M.J. Race-specific and ulvan-induced defense responses in bean (Phaseolus vulgaris) against Colletotrichum lindemuthianum. Physiol. Mol. Plant Pathol. 2012, 78, 8–13. [Google Scholar] [CrossRef]
- Paulert, R.; Talamini, V.; Cassolato, J.E.F.; Duarte, M.E.R.; Noseda, M.D.; Smania, A.; Stadnik, M.J. Effects of sulfated polysaccharide and alcoholic extracts from green seaweed Ulva fasciata on anthracnose severity and growth of common bean (Phaseolus vulgaris L.). J. Plant Dis. Prot. 2009, 116, 263–270. [Google Scholar] [CrossRef]
- Cluzet, S.; Torregrosa, C.; Jacquet, C.; Lafitte, C.; Fournier, J.; Mercier, L.; Salamagne, S.; Briand, X.; Esquerré-Tugaye, M.-T.; Dumas, B. Gene expression profiling and protection of Medicago truncatula against a fungal infection in response to an elicitor from green algae Ulva spp. Plant Cell Environ. 2004, 27, 917–928. [Google Scholar] [CrossRef]
- Kim, S.J.; Ko, E.J.; Hong, J.K.; Jeun, Y.C. Ultrastructures of Colletotrichum orbiculare in cucumber leaves expressing systemic acquired resistance mediated by Chlorella fusca. Plant Pathol. J. 2018, 34, 113–120. [Google Scholar] [CrossRef]
- Aguado, A.; Pastrana, A.M.; de los Santos, B.; Romero, F.; Sánchez, M.C.; Capotea, N. Efficiency of natural products in the control of Colletotrichum acutatum monitored by real-time PCR. Acta Hortic. 2014, 1049, 329–334. [Google Scholar] [CrossRef]
- Pavela, R.; Benelli, G. Essential Oils as Ecofriendly Biopesticides? Challenges and Constraints. Trends Plant Sci. 2016, 21, 1000–1007. [Google Scholar] [CrossRef]
- Jugreet, B.S.; Suroowan, S.; Rengasamy, R.R.K.; Mahomoodally, M.F. Chemistry, bioactivities, mode of action and industrial applications of essential oils. Trends Food Sci. Technol. 2020, 101, 89–105. [Google Scholar] [CrossRef]
- Borrelli, V.M.G.; Brambilla, V.; Rogowsky, P.; Marocco, A.; Lanubile, A. The enhancement of plant disease resistance using CRISPR/Cas9 technology. Front. Plant Sci. 2018, 9, 1245. [Google Scholar] [CrossRef]
- Mishra, R.; Mohanty, J.N.; Mahanty, B.; Joshi, R.K. A single transcript CRISPR/Cas9 mediated mutagenesis of CaERF28 confers anthracnose resistance in chilli pepper (Capsicum annuum L.). Planta 2021, 254, 5. [Google Scholar] [CrossRef] [PubMed]
- Mishra, R.; Joshi, R.K.; Zhao, K. Genome editing in rice: Recent advances, challenges, and future implications. Front. Plant Sci. 2018, 9, 1361. [Google Scholar] [CrossRef] [PubMed]
- Mishra, R.; Mohanty, J.N.; Chand, S.K.; Joshi, R.K. Can-miRn37a mediated suppression of ethylene response factors enhances the resistance of chilli against anthracnose pathogen Colletotrichum truncatum L. Plant Sci. 2018, 267, 135–147. [Google Scholar] [CrossRef] [PubMed]
- Baulcombe, D. RNA silencing in plants. Nature 2004, 431, 356–363. [Google Scholar] [CrossRef]
- Ketting, R.F. The many faces of RNAi. Dev. Cell 2011, 20, 148–161. [Google Scholar] [CrossRef] [Green Version]
- Castel, S.E.; Martienssen, R.A. RNA interference in the nucleus: Roles for small RNAs in transcription, epigenetics and beyond. Nat. Rev. Genet. 2013, 14, 100–112. [Google Scholar] [CrossRef]
- Wang, M.; Weiberg, A.; Lin, F.M.; Thomma, B.P.H.J.; Huang, H.-D.; Jin, H. Bidirectional cross-kingdom RNAi and fungal uptake of external RNAs confer plant protection. Nat. Plants 2016, 2, 16151. [Google Scholar] [CrossRef]
- Cai, Q.; He, B.; Kogel, K.-H.; Jin, H. Cross-kingdom RNA trafficking and environmental RNAi—Nature’s blueprint for modern crop protection strategies. Curr. Opin. Microbiol. 2018, 46, 58–64. [Google Scholar] [CrossRef]
- Huang, C.-Y.; Wang, H.; Hu, P.; Hamby, R.; Jin, H. Small RNAs—Big players in plant-microbe interactions. Cell Host Microbe 2019, 26, 173–182. [Google Scholar] [CrossRef] [Green Version]
- Gebremichael, D.E.; Haile, Z.M.; Negrini, F.; Sabbadini, S.; Capriotti, L.; Mezzetti, B.; Baraldi, E. RNA interference strategies for future management of plant pathogenic fungi: Prospects and challenges. Plants 2021, 10, 650. [Google Scholar] [CrossRef]
- Lax, C.; Tahiri, G.; Patiño-Medina, J.A.; Cánovas-Márquez, J.T.; Pérez-Ruiz, J.A.; Osorio-Concepción, M.; Navarro, E.; Calo, S. The evolutionary significance of RNAi in the fungal kingdom. Int. J. Mol. Sci. 2020, 21, 9348. [Google Scholar] [CrossRef] [PubMed]
- Senthil-Kumar, M.; Mysore, K.S. RNAi in plants: Recent developments and applications in agriculture. In Gene Silencing: Theory Techniques and Applications; Catalano, A.J., Ed.; Nova Science Publishers: New York, NY, USA, 2010; pp. 183–199. [Google Scholar]
- Nunes, C.C.; Dean, R.A. Host-induced gene silencing: A tool for understanding fungal host interaction and for developing novel disease control strategies. Mol. Plant Pathol. 2012, 13, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Capriotti, L.; Baraldi, E.; Mezzetti, B.; Limera, C.; Sabbadini, S. Biotechnological approaches: Gene overexpression, gene silencing and genome editing to control fungal and oomycete diseases in grapevine. Int. J. Mol. Sci. 2020, 21, 5701. [Google Scholar] [CrossRef]
- Weiberg, A.; Wang, M.; Lin, F.-M.; Zhao, H.; Zhang, Z.; Kaloshian, I.; Huang, H.-D.; Jin, H. Fungal small RNAs suppress plant immunity by hijacking host RNA interference pathways. Science 2013, 342, 118–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosa, C.; Kuo, Y.-W.; Wuriyanghan, H.; Falk, B.W. RNA interference mechanisms and applications in plant pathology. Annu. Rev. Phytopathol. 2018, 56, 581–610. [Google Scholar] [CrossRef] [PubMed]
- Vetukuri, R.R.; Dubey, M.; Kalyandurg, P.B.; Carlsson, A.S.; Whisson, S.C.; Ortiz, R. Spray-induced gene silencing: An innovative strategy for plant trait improvement and disease control. Crop Breed. Appl. Biotechnol. 2021, 21, e387921S11. [Google Scholar] [CrossRef]
- Rutter, B.D.; Innes, R.W. Extracellular vesicles as key mediators of plant–microbe interactions. Curr. Opin. Plant Biol. 2018, 44, 16–22. [Google Scholar] [CrossRef]
- Bielska, E.; Birch, P.R.J.; Buck, A.H.; Abreu-Goodger, C.; Innes, R.W.; Jin, H.; Pfaffl, M.W.; Robatzek, S.; Regev-Rudzki, N.; Tisserant, C.; et al. Highlights of the mini-symposium on extracellular vesicles in inter-organismal communication, held in Munich, Germany, August 2018. J. Extracell. Vesicles 2019, 8, 1590116. [Google Scholar] [CrossRef] [Green Version]
- Cai, Q.; He, B.; Weiberg, A.; Buck, A.H.; Jin, H. Small RNAs and extracellular vesicles: New mechanisms of cross-species communication and innovative tools for disease control. PLoS Pathog. 2019, 15, e1008090. [Google Scholar] [CrossRef] [Green Version]
- Šečić, E.; Kogel, K.H. Requirements for fungal uptake of dsRNA and gene silencing in RNAi-based crop protection strategies. Curr. Opin. Biotechnol. 2021, 70, 136–142. [Google Scholar] [CrossRef]
- Laurie, J.D.; Linning, R.; Bakkeren, G. Hallmarks of RNA silencing are found in the smut fungus Ustilago hordei but not in its close relative Ustilago maydis. Curr. Genet. 2008, 53, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-C.; Li, L.; Gu, W.; Xue, Z.; Crosthwaite, S.K.; Pertsemlidis, A.; Lewis, Z.; Freitag, M.; Selker, E.U.; Mello, C.C.; et al. Diverse pathways generate microRNA-like RNAs and Dicer-independent small interfering RNAs in fungi. Mol. Cell. 2010, 38, 803–814. [Google Scholar] [CrossRef] [Green Version]
- Trieu, T.A.; Calo, S.; Nicolás, F.E.; Vila, A.; Moxon, S.; Dalmay, T.; Torres-Martínez, S.; Garre, V.; Ruiz Vázquez, R.M. A non-canonical RNA silencing pathway promotes mRNA degradation in basal fungi. PLoS Genet. 2015, 11, e1005168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goulin, E.H.; de Lima, T.A.; dos Santos, P.J.C.; Machado, M.A. RNAi-induced silencing of the succinate dehydrogenase subunits gene in Colletotrichum abscissum, the causal agent of postbloom fruit drop (PFD) in citrus. Microbiol. Res. 2022, 260, 126938. [Google Scholar] [CrossRef]
- Baum, J.A.; Bogaert, T.; Clinton, W.; Heck, G.R.; Feldmann, P.; Ilagan, O.; Johnson, S.; Plaetinck, G.; Munyikwa, T.; Pleau, M.; et al. Control of coleopteran insect pests through RNA interference. Nat. Biotechnol. 2007, 25, 1322–1326. [Google Scholar] [CrossRef] [PubMed]
- Nowara, D.; Gay, A.; Lacomme, C.; Shaw, J.; Ridout, C.; Douchkov, D.; Hensel, G.; Kumlehn, J.; Schweizer, P. HIGS: Host-induced gene silencing in the obligate biotrophic fungal pathogen Blumeria graminis. Plant Cell 2010, 22, 3130–3141. [Google Scholar] [CrossRef] [Green Version]
- Waterhouse, P.M.; Graham, M.W.; Wang, M.-B. Virus resistance and gene silencing in plants can be induced by simultaneous expression of sense and antisense RNA. Proc. Natl. Acad. Sci. USA 1998, 95, 13959–13964. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Jin, H. Spray-induced gene silencing: A powerful innovative strategy for crop protection. Trends Microbiol. 2017, 25, 4–6. [Google Scholar] [CrossRef] [Green Version]
- Koch, A.; Wassenegger, M. Host-induced gene silencing—Mechanisms and applications. New Phytol. 2021, 231, 54–59. [Google Scholar] [CrossRef]
- Van Schie, C.C.N.; Takken, F.L.W. Susceptibility genes 101: How to be a good host. Annu. Rev. Phytopathol. 2014, 52, 551–581. [Google Scholar] [CrossRef]
- Niu, D.; Hamby, R.; Sanchez, J.N.; Cai, Q.; Yan, Q.; Jin, H. RNAs—A new frontier in crop protection. Curr. Opin. Biotechnol. 2021, 70, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Yadav, B.C.; Veluthambi, K.; Subramaniam, K. Host-generated double stranded RNA induces RNAi in plant-parasitic nematodes and protects the host from infection. Mol. Biochem. Parasitol. 2006, 148, 219–222. [Google Scholar] [CrossRef] [PubMed]
- Koch, A.; Kogel, K.-H. New wind in the sails: Improving the agronomic value of crop plants through RNAi-mediated gene silencing. Plant Biotechnol. J. 2014, 12, 821–831. [Google Scholar] [CrossRef] [PubMed]
- Head, G.P.; Carroll, M.W.; Evans, S.P.; Rule, D.M.; Willse, A.R.; Clark, T.L.; Storer, N.P.; Flannagan, R.D.; Samuel, L.W.; Meinke, L.J. Evaluation of SmartStax and SmartStax PRO maize against western corn rootworm and northern corn rootworm: Efficacy and resistance management. Pest Manag. Sci. 2017, 73, 1883–1899. [Google Scholar] [CrossRef] [Green Version]
- Cheng, W.; Song, X.-S.; Li, H.-P.; Cao, L.-H.; Sun, K.; Qiu, X.-L.; Xu, Y.-B.; Yang, P.; Huang, T.; Zhang, J.-B.; et al. Host-induced gene silencing of an essential chitin synthase gene confers durable resistance to Fusarium head blight and seedling blight in wheat. Plant Biotechnol. J. 2015, 13, 1335–1345. [Google Scholar] [CrossRef]
- Zhang, T.; Jin, Y.; Zhao, J.-H.; Gao, F.; Zhou, B.-J.; Fang, Y.-Y.; Guo, H.-S. Host-Induced Gene Silencing of the target gene in fungal cells confers effective resistance to the cotton wilt disease pathogen Verticillium dahliae. Mol. Plant 2016, 9, 939–942. [Google Scholar] [CrossRef] [Green Version]
- Panwar, V.; Jordan, M.; McCallum, B.; Bakkeren, G. Host-induced silencing of essential genes in Puccinia triticina through transgenic expression of RNAi sequences reduces severity of leaf rust infection in wheat. Plant Biotechnol. J. 2018, 16, 1013–1023. [Google Scholar] [CrossRef] [Green Version]
- Ghag, S.B.; Shekhawat, U.K.S.; Ganapathi, T.R. Host-induced post-transcriptional hairpin RNA-mediated gene silencing of vital fungal genes confers efficient resistance against Fusarium wilt in banana. Biotechnol. J. 2014, 12, 541–553. [Google Scholar] [CrossRef]
- He, F.; Zhang, R.; Zhao, J.; Qi, T.; Kang, Z.; Guo, J. Host-induced silencing of Fusarium graminearum genes enhances the resistance of Brachypodium distachyon to Fusarium head blight. Front. Plant. Sci. 2019, 10, 1362. [Google Scholar] [CrossRef]
- Hu, D.; Chen, Z.-Y.; Zhang, C.; Ganiger, M. Reduction of Phakopsora pachyrhizi infection on soybean through host- and spray-induced gene silencing. Mol. Plant Pathol. 2020, 21, 794–807. [Google Scholar] [CrossRef] [Green Version]
- Mahto, B.K.; Singh, A.; Pareek, M.; Rajam, M.V.; Dhar-Ray, S.; Reddy, P.M. Host-induced silencing of the Colletotrichum gloeosporioides conidial morphology 1 gene (CgCOM1) confers resistance against Anthracnose disease in chilli and tomato. Plant Mol. Biol. 2020, 104, 381–395. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhao, X.; Sun, J.; Kang, Z.; Ding, S.; Xu, J.-R.; Peng, Y.-L. A novel protein Com1 is required for normal conidium morphology and full virulence in Magnaporthe oryzae. Mol. Plant Microbe Interact. 2010, 23, 112–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, A.; Höfle, L.; Werner, B.T.; Imani, J.; Schmidt, A.; Jelonek, L.; Kogel, K.H. SIGS vs. HIGS: A study on the efficacy of two dsRNA delivery strategies to silence Fusarium FgCYP51 genes in infected host and non-host plants. Mol. Plant Pathol. 2019, 20, 1636–1644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, A.; Biedenkopf, D.; Furch, A.; Weber, L.; Rossbach, O.; Abdellatef, E.; Linicus, L.; Johannsmeier, J.; Jelonek, L.; Goesmann, A.; et al. An RNAi-based control of Fusarium graminearum infections through spraying of long dsRNAs involves a plant passage and is controlled by the fungal silencing machinery. PLoS Pathog. 2016, 12, e1005901. [Google Scholar] [CrossRef]
- McLoughlin, A.G.; Wytinck, N.; Walker, P.L.; Girard, I.J.; Rashid, K.Y.; de Kievit, T.; Fernando, W.G.D.; Whyard, S.; Belmonte, M.F. Identification and application of exogenous dsRNA confers plant protection against Sclerotinia sclerotiorum and Botrytis cinerea. Sci. Rep. 2018, 8, 7320. [Google Scholar] [CrossRef]
- Qiao, L.; Lan, C.; Capriotti, L.; Ah-Fong, A.; Nino Sanchez, J.; Hamby, R.; Heller, J.; Zhao, H.; Glass, N.L.; Judelson, H.S.; et al. Spray-induced gene silencing for disease control is dependent on the efficiency of pathogen RNA uptake. Plant Biotechnol. J. 2021, 19, 1756–1768. [Google Scholar] [CrossRef]
- Rosa, S.; Pesaresi, P.; Mizzotti, C.; Bulone, V.; Mezzetti, B.; Baraldi, E.; Masiero, S. Game-changing alternatives to conventional fungicides: Small RNAs and short peptides. Trends Biotechnol. 2022, 40, 320–337. [Google Scholar] [CrossRef]
- Whangbo, J.S.; Hunter, C.P. Environmental RNA interference. Trends Genet. 2008, 24, 297–305. [Google Scholar] [CrossRef]
- Chaloner, T.; van Kan, J.A.L.; Grant-Downton, R.T. RNA ‘Information Warfare’ in pathogenic and mutualistic interactions. Trends Plant Sci. 2016, 21, 738–748. [Google Scholar] [CrossRef]
- Wytinck, N.; Manchur, C.L.; Li, V.H.; Whyard, S.; Belmonte, M.F. dsRNA uptake in plant pests and pathogens: Insights into RNAi-based insect and fungal control technology. Plants 2020, 9, 1780. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, J.; Zhu, K.Y. Chitosan/double-stranded RNA nanoparticle-mediated RNA interference to silence chitin synthase genes through larval feeding in the African malaria mosquito (Anopheles gambiae). Insect Mol. Biol. 2010, 19, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Numata, K.; Ohtani, M.; Yoshizumi, T.; Demura, T.; Kodama, Y. Local gene silencing in plants via synthetic dsRNA and carrier peptide. Plant Biotechnol. J. 2014, 12, 1027–1034. [Google Scholar] [CrossRef] [PubMed]
- Mitter, N.; Worrall, E.A.; Robinson, K.E.; Li, P.; Jain, R.G.; Taochy, C.; Fletcher, S.J.; Carroll, B.J.; Lu, G.Q.; Xu, Z.P. Clay nanosheets for topical delivery of RNAi for sustained protection against plant viruses. Nat. Plants 2017, 3, 16207. [Google Scholar] [CrossRef] [PubMed]
- Mitter, N.; Worrall, E.A.; Robinson, K.E.; Xu, Z.P.; Carroll, B.J. Induction of virus resistance by exogenous application of double-stranded RNA. Curr. Opin. Virol. 2017, 26, 49–55. [Google Scholar] [CrossRef]
- Lichtenberg, S.S.; Tsyusko, O.V.; Palli, S.R.; Unrine, J.M. Uptake and bioactivity of chitosan/double-stranded RNA polyplex nanoparticles in Caenorhabditis elegans. Environ. Sci. Technol. 2019, 53, 3832–3840. [Google Scholar] [CrossRef] [PubMed]
- Vurro, M.; Miguel-Rojas, C.; Perez-de-Luque, A. Safe nanotechnologies for increasing the effectiveness of environmentally friendly natural agrochemicals. Pest Manag. Sci. 2019, 75, 2403–2412. [Google Scholar] [CrossRef]
- Christiaens, O.; Whyard, S.; Vélez, A.M.; Smagghe, G. Double-stranded RNA technology to control insect pests: Current status and challenges. Front. Plant Sci. 2020, 11, 451. [Google Scholar] [CrossRef]
- Schwartz, S.H.; Hendrix, B.; Hoffer, P.; Sanders, R.A.; Zheng, W. Carbon dots for efficient small interfering RNA delivery and gene silencing in plants. Plant Physol. 2020, 184, 647–657. [Google Scholar] [CrossRef]
- Wang, K.; Peng, Y.; Chen, J.; Peng, Y.; Wang, X.; Shen, Z.; Han, Z. Comparison of efficacy of RNAi mediated by various nanoparticles in the rice striped stem borer (Chilo suppressalis). Pestic. Biochem. Physiol. 2020, 165, 104467. [Google Scholar] [CrossRef]
- Kupferschmidt, K. Lethal dose of RNA. Science 2013, 341, 732–733. [Google Scholar] [CrossRef]
- Taning, C.N.T.; Mezzetti, B.; Kleter, G.; Smagghe, G.; Baraldi, E. Does RNAi-based technology fit within EU sustainability goals? Trends Biotechnol. 2021, 31, 664–667. [Google Scholar] [CrossRef] [PubMed]
- Gu, K.-X.; Song, X.-S.; Xiao, X.-M.; Duan, X.-X.; Wang, J.-X.; Duan, Y.-B.; Hou, Y.-P.; Zhou, M.-G. A β2-tubulin dsRNA derived from Fusarium asiaticum confers plant resistance to multiple phytopathogens and reduces fungicide resistance. Pestic. Biochem. Phys. 2019, 153, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Sui, Y.; Wisniewski, M.; Droby, S.; Liu, Y. Utilization of antagonistic yeasts to manage postharvest fungal diseases of fruit. Int. J. Food Microbiol. 2013, 167, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Ocampo-Suarez, I.B.; Sanchez-Salas, J.L.; Ragazzo-Sanzhez, J.A.; Calder’on-Santoyo, M. Evaluation of the toxicity and pathogenecity of biocontrol agents in murine models, chicken embryos and dermal irritation in rabbits. Toxicol. Res. 2017, 6, 188–198. [Google Scholar]
- Bahadou, S.A.; Ouijja, A.; Karfach, A.; Tahiri, A.; Lahlali, R. New potential bacterial antagonists for the biocontrol of fire blight disease (Erwinia amylovora) in Morocco. Microb. Pathog. 2018, 117, 7–15. [Google Scholar] [CrossRef]
- Bachman, P.M.; Bolognesi, R.; Moar, W.J.; Mueller, G.M.; Paradise, M.S.; Ramaseshadri, P.; Tan, J.; Uffman, J.P.; Warren, J.; Wiggins, B.E.; et al. Characterization of the spectrum of insecticidal activity of a double-stranded RNA with targeted activity against Western Corn Rootworm (Diabrotica virgifera virgifera LeConte). Transgenic Res. 2013, 22, 1207–1222. [Google Scholar] [CrossRef] [Green Version]
- Ramon, M.; Devos, Y.; Lanzoni, A.; Liu, Y.; Gomes, A.; Gennaro, A.; Waigmann, E. RNAi-based GM plants: Food for thought for risk assessors. Plant Biotechnol. J. 2014, 12, 1271–1273. [Google Scholar] [CrossRef]
- Casacuberta, J.M.; Devos, Y.; Du Jardin, P.; Ramon, M.; Vaucheret, H.; Noguè, F. Biotechnological uses of RNAi in plants: Risk assessment considerations. Trends Biotechnol. 2015, 33, 145–147. [Google Scholar] [CrossRef] [Green Version]
- Tan, J.; Levine, S.L.; Bachman, P.M.; Jensen, P.D.; Mueller, G.M.; Uffman, J.P.; Meng, C.; Song, Z.; Richards, K.B.; Beevers, M.H. No impact of DvSnf7 RNA on honey bee (Apis mellifera L.) adults and larvae in dietary feeding tests. Environ. Toxicol. Chem. 2016, 35, 287–294. [Google Scholar] [CrossRef] [Green Version]
- Mezzetti, B.; Smagghe, G.; Arpaia, S.; Christiaens, O.; Pfeilstetter, A.D.; Jones, H.; Kostov, K.; Sabbadini, S.; Opsahl-Sorteberg, H.-G.; Ventura, V.; et al. RNAi: What is its position in agriculture? J. Pest Sci. 2020, 93, 1125–1130. [Google Scholar] [CrossRef]
Figure 1.
Anthracnose symptoms on harvested fruits. (a) Sunken brown lesions on strawberry fruits; (b) dark lesions surrounding orange conidia masses on pear fruits; (c) dark lesions on banana fruits.
Figure 1.
Anthracnose symptoms on harvested fruits. (a) Sunken brown lesions on strawberry fruits; (b) dark lesions surrounding orange conidia masses on pear fruits; (c) dark lesions on banana fruits.

Figure 2.
Approaches for post-harvest anthracnose management. Traditional strategies (yellow panels) and innovative methodologies explored for the improvement of the sustainability in the agri-food sector (green panels).
Figure 2.
Approaches for post-harvest anthracnose management. Traditional strategies (yellow panels) and innovative methodologies explored for the improvement of the sustainability in the agri-food sector (green panels).
Figure 3.
RNAi-based approaches for plant protection: host-induced gene silencing, HIGS (left panel) and spray-induced gene silencing, SIGS (right panel). The main advantages (in green) and disadvantages (in red) related to each strategy are reported. See text for further information.
Figure 3.
RNAi-based approaches for plant protection: host-induced gene silencing, HIGS (left panel) and spray-induced gene silencing, SIGS (right panel). The main advantages (in green) and disadvantages (in red) related to each strategy are reported. See text for further information.
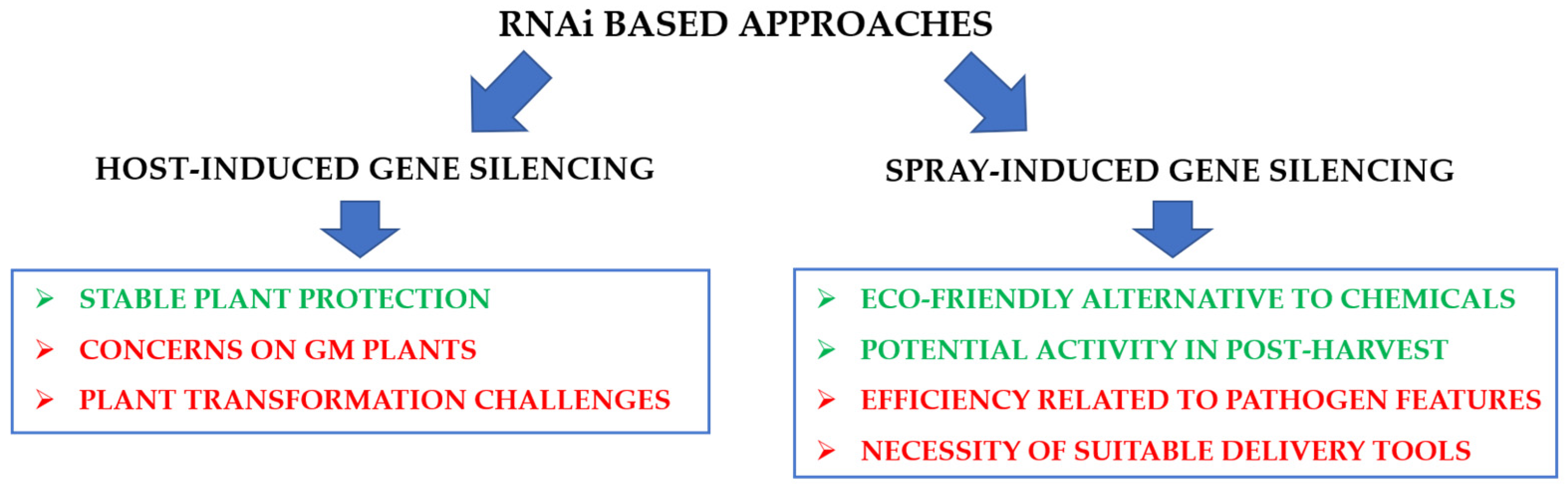

{kind=link}
{kind=link}
{kind=link}
Table 1.
Representative biological agents for Colletotrichum spp. management in post-harvest.
| Biological Agent (Species and Strain/Isolate) | Colletotrichum Species | Host Species | Reference |
|---|---|---|---|
| Bacillus subtilis EA-CB0015 | C. acutatum species complex | Cyphomandra betacea | [74] |
| Bacillus subtilis S16 | C. acutatum species complex | Malus pumila | [75] |
| Prunus persica cv. Chunjungdo | |||
| P. persica cv. Sunfre | |||
| Paenibacillus polymyxa APEC128 | C. acutatum species complex | M. pumila | [76] |
| C. gloeosporioides species complex | |||
| Streptomyces philanthi RM-1-138 | C. gloeosporioides species complex | Capsicum annuum | [77] |
| Burkholderia rinojensis F2 | C. truncatum | C. annuum | [78] |
| B. rinojensis F80 | |||
| Burkholderia gladioli F79 | |||
| Burkholderia arboris F35 | |||
| Pseudomonas aeruginosa F65 | |||
| Trichoderma harzianum T-39 | C. acutatum sensu lato | Fragaria × ananassa | [79] |
| Trichoderma hamatum T-105 | |||
| Trichoderma atroviride T-161 | |||
| Trichoderma longibrachiatum T-166 | |||
| T. harzanium1 | C. gloeosporioides species complex | Citrus limon L. | [80] |
| T. harzanium TH-1 | C. musae | Musa acuminata | [81] |
| T. viridae TV-3 | |||
| T. viridae TV-4 | |||
| Trichoderma pseudokomngii2 | |||
| Metchnikowia pulcherrima FMB-24H-2 | C. acutatum sensu lato | Malus domestica | [82] |
| M. pulcherrima T5-A2 | C. acutatum sensu lato | M. domestica | [83] |
| Pichia kluyveri Y1125 | C. acutatum species complex | M. domestica | [84] |
1 Authors investigated pathogen inhibition in fruits from transgenic plants expressing a T. harzanium antifungal protein (see text for further information). 2 Strain/isolate not specified.
Table 2.
Representative essential oils agents for Colletotrichum spp. management in post-harvest.
| Essential Oil | Colletotrichum Species | Host Species | Reference |
|---|---|---|---|
| Lemongrass oil | C. gloeosporioides species complex | Carica papaya L. | [99] |
| Ginger oil | C. gloeosporioides species complex | C. papaya L. | [100] |
| Savory oil | C. gloeosporioides species complex | C. papaya L. | [101] |
| C. gloeosporioides species complex | Persea americana | [102] | |
| Thyme oil | C. gloeosporioides species complex | C. papaya | [101] |
| C. gloeosporioides species complex | P. americana | [101] | |
| C. gloeosporioides species complex | Mangifera indica L. | [103] | |
| C. acutatum species complex | Fragaria x ananassa | [104] | |
| Cinnamon bark oil | C. acutatum species complex | Fragaria x ananassa | [104] |
| Ocinum basilicum oil | C. musae | Musa spp. | [105] |
| Ocinum gratissimus oil | C. musae | Musa spp. | [105] |
Table 3.
Representative seaweed derivates for Colletotrichum spp. management.
| Seaweed Derivate-Species | Colletotrichum Species | Host Species | Reference |
|---|---|---|---|
| “Ulvan”-Ulva spp. | C. lindemuthianum | Phaseolus vulgaris L. | [117] |
| “Ulvan”-Ulva fasciata | C. lindemuthianum | P. vulgaris L. | [118] |
| Ethanolic fraction-Ulva spp. | C. trifolii | Medicago truncatula | [119] |
| Algal suspension-Chlorella fusca | C. orbiculare | Cucumis sativus | [120] |
| Seaweed-based biofungicide-Ascophyllum nodosum | C. acutatum species complex | Fragaria × ananassa | [121] |
Table 4.
Current investigations on the RNAi-based approaches for Colletotrichum management.
| Approach | Outcome | Reference |
|---|---|---|
| HIGS | C. gloeosporioides species complex inhibition in GM plants carrying a CgCOM1 RNA expressing cassette | [166] |
| SIGS | C. truncatum silencing by means of interfering dsRNAs | [187] |
| Existence of the RNAi machinery in C. abscissum | [149] | |
| Absence of dsRNA uptake efficiency in C. gloeosporioides species complex | [171] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ciofini, A.; Negrini, F.; Baroncelli, R.; Baraldi, E. Management of Post-Harvest Anthracnose: Current Approaches and Future Perspectives. Plants 2022, 11, 1856. https://doi.org/10.3390/plants11141856
AMA Style
Ciofini A, Negrini F, Baroncelli R, Baraldi E. Management of Post-Harvest Anthracnose: Current Approaches and Future Perspectives. Plants. 2022; 11(14):1856. https://doi.org/10.3390/plants11141856
Chicago/Turabian StyleCiofini, Alice, Francesca Negrini, Riccardo Baroncelli, and Elena Baraldi. 2022. "Management of Post-Harvest Anthracnose: Current Approaches and Future Perspectives" Plants 11, no. 14: 1856. https://doi.org/10.3390/plants11141856
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.