
Genome-Wide Association of Stem Carbohydrate Accumulation and Remobilization during Grain Growth in Bread Wheat (Triticum aestivum L.) in Mediterranean Environments


Abstract
:1. Introduction
2. Results
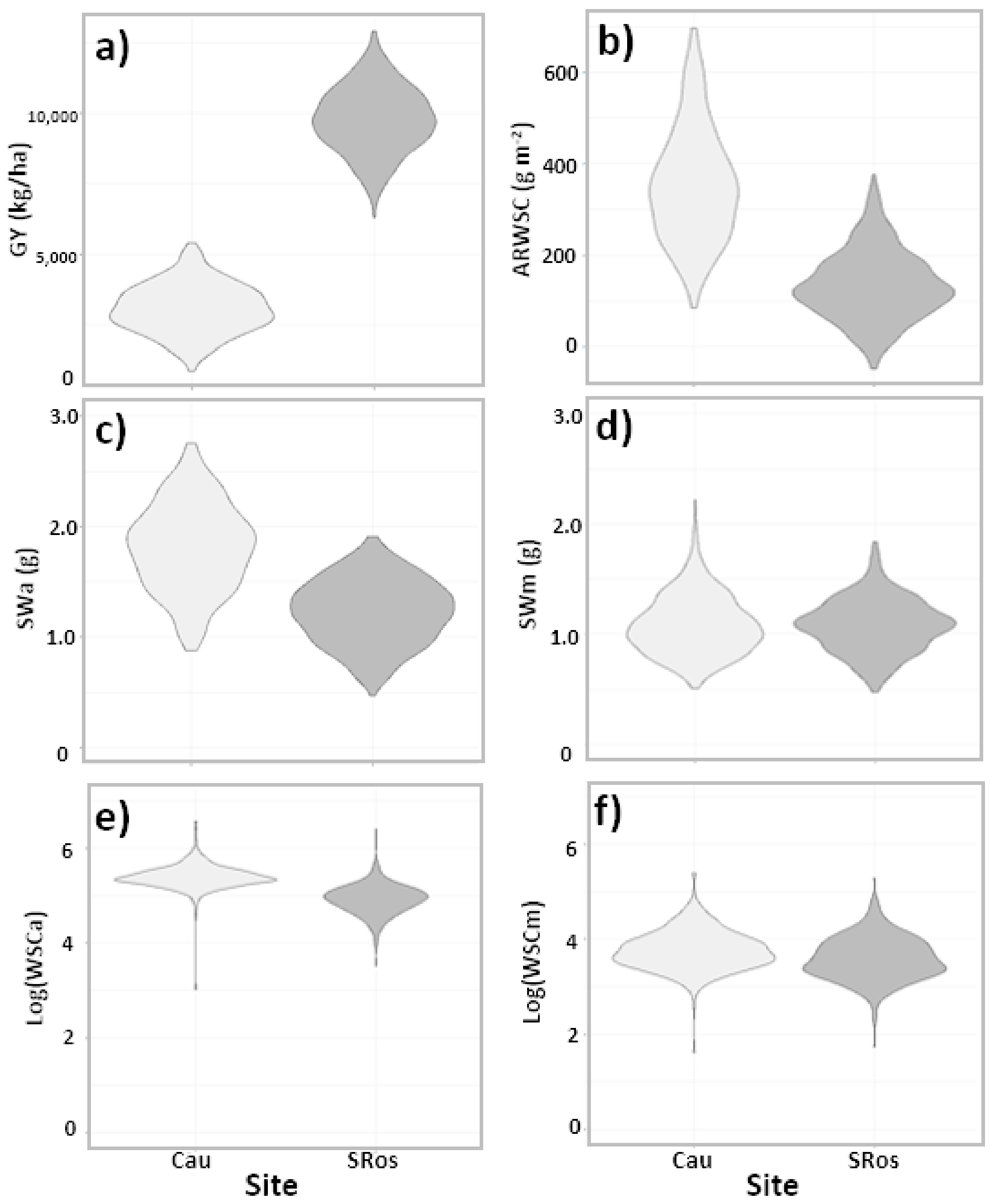
2.1. Grain Yield, Stem Carbohydrate Accumulation, and Remobilization Were Influenced by Genotype and Site
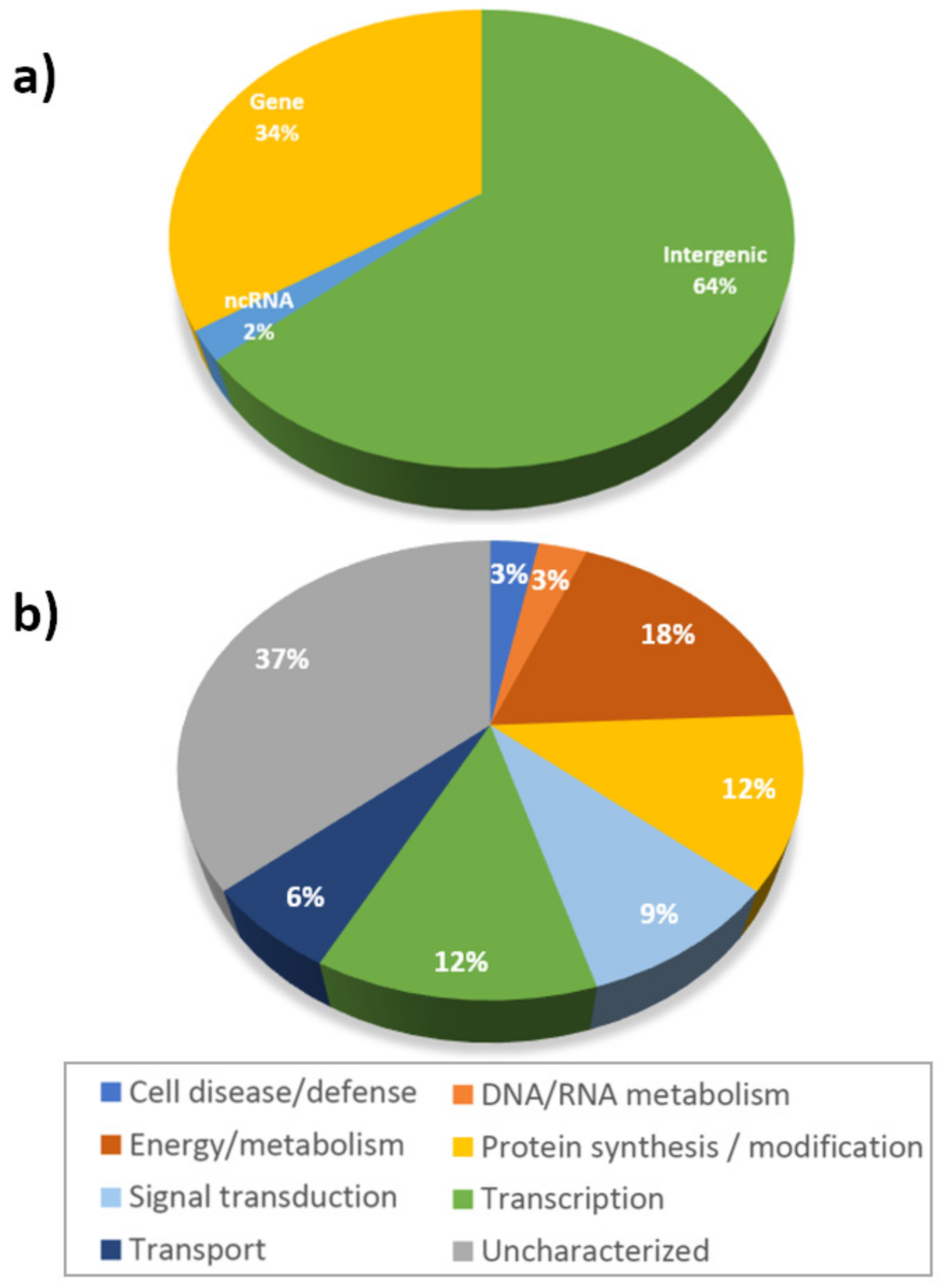
2.2. Significant SNPs Were Detected in Association with Stem Carbohydrate Accumulation and Remobilization
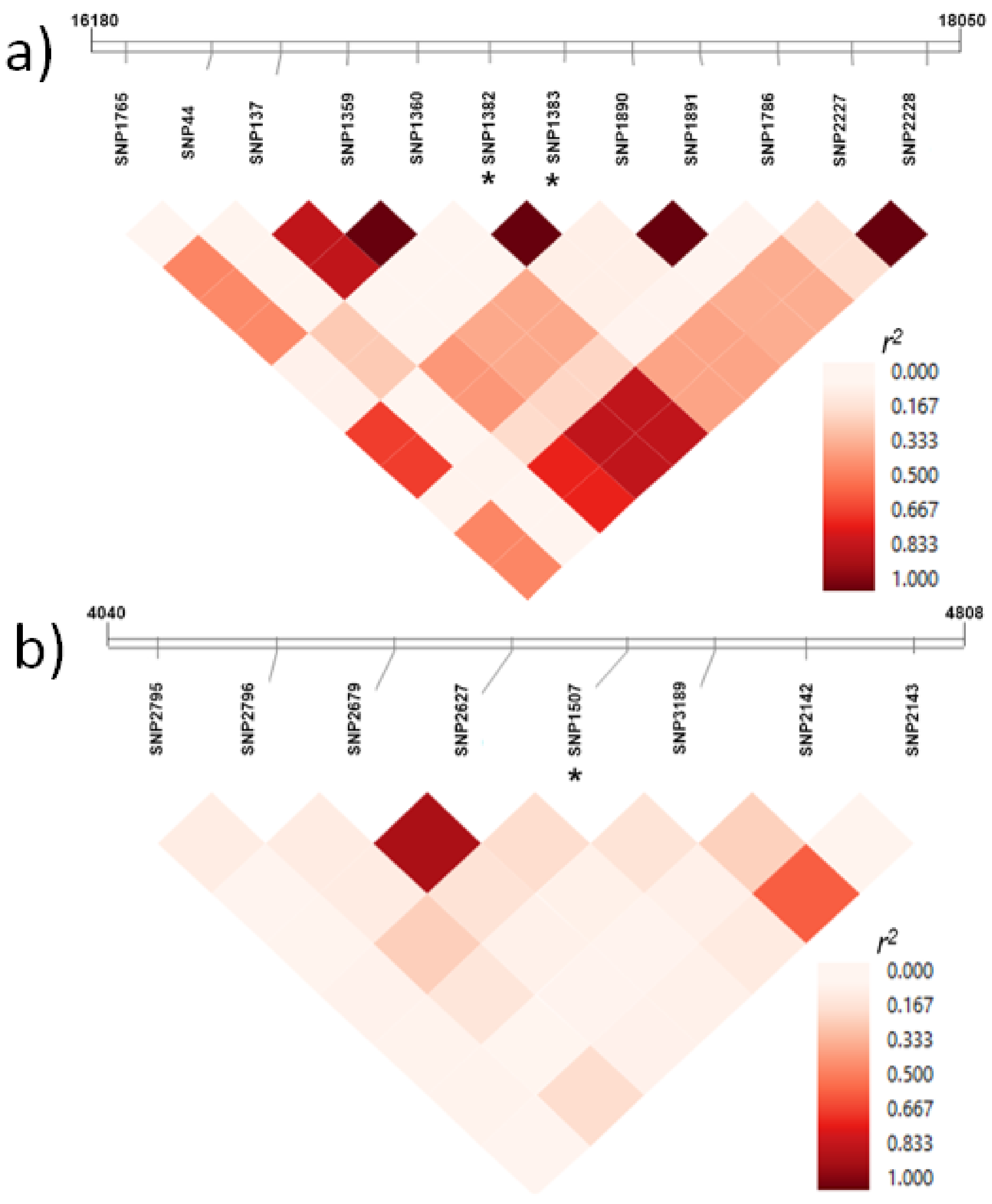
2.3. A Rapid Linkage Disequilibrium (LD) Decay Was Observed across Chromosomes
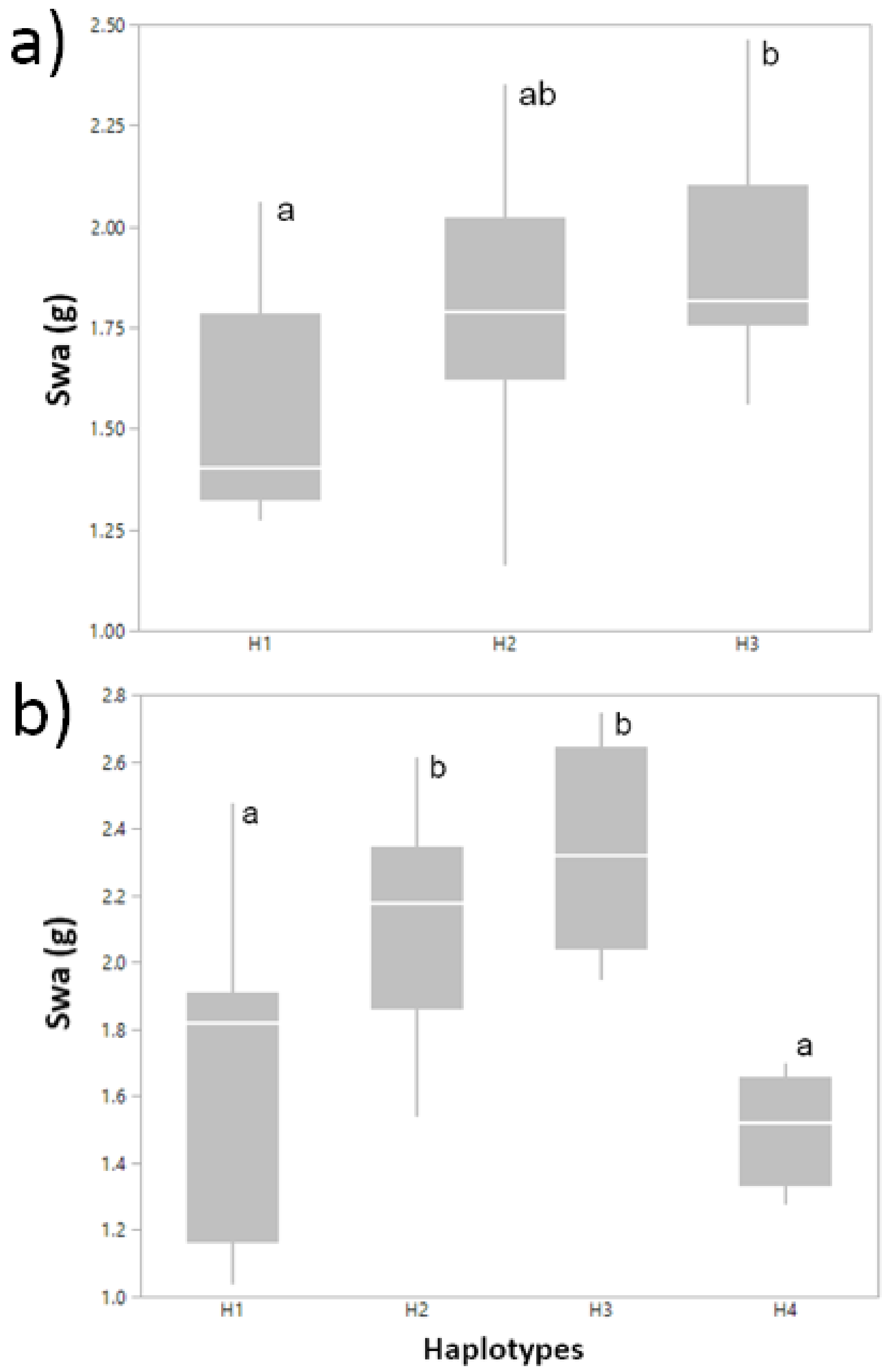
2.4. Single-SNP Markers Are Part of Significant Haplotypes in Association with Traits
3. Discussion
4. Materials and Methods
4.1. Association Population and Growth Conditions
4.2. Agronomic and Physiological Traits
4.3. Statistical and Genetic Analysis
4.4. Genotyping
4.5. Linkage Disequilibrium
4.6. Association Analysis
4.7. Haplotype Analysis
4.8. Gene Models and Annotations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lopes, M.S.; Rebetzke, G.J.; Reynolds, M. Integration of phenotyping and genetic platforms for a better understanding of wheat performance under drought. J. Exp. Bot. 2014, 65, 6167–6177. [Google Scholar] [CrossRef] [Green Version]
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield Trends Are Insufficient to Double Global Crop Production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef] [Green Version]
- Ortiz, R.; Sayre, K.D.; Govaerts, B.; Gupta, R.; Subbarao, G.V.; Ban, T.; Hodson, D.; Dixon, J.M.; Iván Ortiz-Monasterio, J.; Reynolds, M. Climate change: Can wheat beat the heat? Agric. Ecosyst. Environ. 2008, 126, 46–58. [Google Scholar] [CrossRef]
- Porter, J.R.; Semenov, M.A. Crop responses to climatic variation. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2005, 360, 2021–2035. [Google Scholar] [CrossRef]
- Pinto, R.S.; Reynolds, M.P.; Mathews, K.L.; McIntyre, C.L.; Olivares-Villegas, J.J.; Chapman, S.C. Heat and drought adaptive QTL in a wheat population designed to minimize confounding agronomic effects. TAG Theor. Appl. Genet. 2010, 121, 1001–1021. [Google Scholar] [CrossRef] [Green Version]
- Dolferus, R.; Powell, N.; Xuemei, J.I.; Ravash, R.; Edlington, J.; Oliver, S.; Van Dongen, J.; Shiran, B. The physiology of reproductive-stage abiotic stress tolerance in cereals. In Molecular Stress Physiology of Plants; Rout, G.D.A., Ed.; Springer: New Delhi, India, 2013; pp. 193–218. [Google Scholar]
- Monneveux, P.; Rekika, D.; Acevedo, E.; Merah, O. Effect of drought on leaf gas exchange, carbon isotope discrimination, transpiration efficiency and productivity in field grown durum wheat genotypes. Plant Sci. 2006, 170, 867–872. [Google Scholar] [CrossRef]
- Shokat, S.; Großkinsky, D.K.; Liu, F. Impact of elevated CO2 on two contrasting wheat genotypes exposed to intermediate drought stress at anthesis. J. Agron. Crop Sci. 2021, 207, 20–33. [Google Scholar] [CrossRef]
- Shokat, S.; Großkinsky, D.K.; Roitsch, T.; Liu, F. Activities of leaf and spike carbohydrate-metabolic and antioxidant enzymes are linked with yield performance in three spring wheat genotypes grown under well-watered and drought conditions. BMC Plant Biol. 2020, 20, 400. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhang, B.; Li, R.; Chang, X.; Jing, R. Favorable alleles for stem water-soluble carbohydrates identified by association analysis contribute to grain weight under drought stress conditions in wheat. PLoS ONE 2015, 10, e0119438. [Google Scholar] [CrossRef]
- Blum, A. Improving wheat grain filling under stress by stem reserve mobilisation. Euphytica 1998, 100, 77–83. [Google Scholar] [CrossRef]
- Dreccer, M.F.; van Herwaarden, A.F.; Chapman, S.C. Grain number and grain weight in wheat lines contrasting for stem water soluble carbohydrate concentration. Field Crops Res. 2009, 112, 43–54. [Google Scholar] [CrossRef]
- Ehdaie, B.; Alloush, G.A.; Madore, M.A.; Waines, J.G. Genotypic Variation for Stem Reserves and Mobilization in Wheat: II. Postanthesis Changes in Internode Water-Soluble Carbohydrates. Crop Sci. 2006, 46, 2093–2103. [Google Scholar] [CrossRef]
- Reynolds, M.; Dreccer, F.; Trethowan, R. Drought-adaptive traits derived from wheat wild relatives and landraces. J. Exp. Bot. 2007, 58, 177–186. [Google Scholar] [CrossRef] [Green Version]
- Del Pozo, A.; Yáñez, A.; Matus, I.A.; Tapia, G.; Castillo, D.; Sanchez-Jardón, L.; Araus, J.L. Physiological Traits Associated with Wheat Yield Potential and Performance under Water-Stress in a Mediterranean Environment. Front. Plant Sci. 2016, 7, 987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yáñez, A.; Tapia, G.; Guerra, F.; del Pozo, A. Stem carbohydrate dynamics and expression of genes involved in fructan accumulation and remobilization during grain growth in wheat (Triticum aestivum L.) genotypes with contrasting tolerance to water stress. PLoS ONE 2017, 12, e0177667. [Google Scholar] [CrossRef] [Green Version]
- Foulkes, M.J.; Sylvester-Bradley, R.; Weightman, R.; Snape, J.W. Identifying physiological traits associated with improved drought resistance in winter wheat. Field Crops Res. 2007, 103, 11–24. [Google Scholar] [CrossRef]
- Ruuska, S.A.; Rebetzke, G.J.; van Herwaarden, A.F.; Richards, R.A.; Fettell, N.A.; Tabe, L.; Jenkins, C.L.D. Genotypic variation in water-soluble carbohydrate accumulation in wheat. Funct. Plant Biol. 2006, 33, 799–809. [Google Scholar] [CrossRef]
- Habash, D.Z.; Baudo, M.; Hindle, M.; Powers, S.J.; Defoin-Platel, M.; Mitchell, R.; Saqi, M.; Rawlings, C.; Latiri, K.; Araus, J.L.; et al. Systems responses to progressive water stress in durum wheat. PLoS ONE 2014, 9, e108431. [Google Scholar]
- Li, W.-T.; Liu, C.; Liu, Y.-X.; Pu, Z.-E.; Dai, S.-F.; Wang, J.-R.; Lan, X.-J.; Zheng, Y.-L.; Wei, Y.-M. Meta-analysis of QTL associated with tolerance to abiotic stresses in barley. Euphytica 2013, 189, 31–49. [Google Scholar] [CrossRef]
- Tripathi, P.; Rabara, R.C.; Rushton, P.J. A systems biology perspective on the role of WRKY transcription factors in drought responses in plants. Planta 2014, 239, 255–266. [Google Scholar] [CrossRef]
- Snape, J.W.; Foulkes, M.J.; Simmonds, J.; Leverington, M.; Fish, L.J.; Wang, Y.; Ciavarrella, M. Dissecting gene × environmental effects on wheat yields via QTL and physiological analysis. Euphytica 2007, 154, 401–408. [Google Scholar] [CrossRef]
- Rebetzke, G.J.; van Herwaarden, A.F.; Jenkins, C.; Weiss, M.; Lewis, D.; Ruuska, S.; Tabe, L.; Fettell, N.A.; Richards, R.A. Quantitative trait loci for water-soluble carbohydrates and associations with agronomic traits in wheat. Aust. J. Agric. Res. 2008, 59, 891–905. [Google Scholar] [CrossRef]
- McIntyre, C.L.; Seung, D.; Casu, R.E.; Rebetzke, G.J.; Shorter, R.; Xue, G.P. Genotypic variation in the accumulation of water soluble carbohydrates in wheat. Funct. Plant Biol. 2012, 39, 560–568. [Google Scholar] [CrossRef]
- Lai, K.; Lorenc, M.T.; Lee, H.C.; Berkman, P.J.; Bayer, P.E.; Visendi, P.; Ruperao, P.; Fitzgerald, T.L.; Zander, M.; Chan, C.K.; et al. Identification and characterization of more than 4 million intervarietal SNPs across the group 7 chromosomes of bread wheat. Plant Biotechnol. J. 2015, 13, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Garg, B.; Lata, C.; Prasad, M. A study of the role of gene TaMYB2 and an associated SNP in dehydration tolerance in common wheat. Mol. Biol. Rep. 2012, 39, 10865–10871. [Google Scholar] [CrossRef] [PubMed]
- Shokat, S.; Sehgal, D.; Vikram, P.; Liu, F.; Singh, S. Molecular Markers Associated with Agro-Physiological Traits under Terminal Drought Conditions in Bread Wheat. Int. J. Mol. Sci. 2020, 21, 3156. [Google Scholar] [CrossRef] [PubMed]
- Kassa, M.T.; You, F.M.; Hiebert, C.W.; Pozniak, C.J.; Fobert, P.R.; Sharpe, A.G.; Menzies, J.G.; Humphreys, D.G.; Harrison, N.R.; Fellers, J.P.; et al. Highly predictive SNP markers for efficient selection of the wheat leaf rust resistance gene Lr16. BMC Plant Biol. 2017, 17, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.-X.; Zhu, Y.-L.; Zhang, D.-X.; Shao, H.; Liu, P.; Hu, J.-B.; Zhang, H.; Zhang, H.-P.; Chang, C.; Lu, J.; et al. Genome-wide association study for grain yield and related traits in elite wheat varieties and advanced lines using SNP markers. PLoS ONE 2017, 12, e0188662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mora, F.; Castillo, D.; Lado, B.; Matus, I.; Poland, J.; Belzile, F.; von Zitzewitz, J.; del Pozo, A. Genome-wide association mapping of agronomic traits and carbon isotope discrimination in a worldwide germplasm collection of spring wheat using SNP markers. Mol. Breed. 2015, 35, 69. [Google Scholar] [CrossRef]
- Kumar, A.; Mantovani, E.E.; Seetan, R.; Soltani, A.; Echeverry-Solarte, M.; Jain, S.; Simsek, S.; Doehlert, D.; Alamri, M.S.; Elias, E.M.; et al. Dissection of Genetic Factors underlying Wheat Kernel Shape and Size in an Elite x Nonadapted Cross using a High Density SNP Linkage Map. Plant Genome 2016, 9. [Google Scholar] [CrossRef] [Green Version]
- Lucas, S.J.; Salantur, A.; Yazar, S.; Budak, H. High-throughput SNP genotyping of modern and wild emmer wheat for yield and root morphology using a combined association and linkage analysis. Funct. Integr. Genom. 2017, 17, 667–685. [Google Scholar] [CrossRef]
- Xiong, H.; Guo, H.; Zhou, C.; Guo, X.; Xie, Y.; Zhao, L.; Gu, J.; Zhao, S.; Ding, Y.; Liu, L. A combined association mapping and t-test analysis of SNP loci and candidate genes involving in resistance to low nitrogen traits by a wheat mutant population. PLoS ONE 2019, 14, e0211492. [Google Scholar] [CrossRef]
- Huang, S.; Sun, L.; Hu, X.; Wang, Y.; Zhang, Y.; Nevo, E.; Peng, J.; Sun, D. Associations of canopy leaf traits with SNP markers in durum wheat (Triticum turgidum L. durum (Desf.)). PLoS ONE 2018, 13, e0206226. [Google Scholar] [CrossRef]
- Dong, Y.; Liu, J.; Zhang, Y.; Geng, H.; Rasheed, A.; Xiao, Y.; Cao, S.; Fu, L.; Yan, J.; Wen, W.; et al. Genome-Wide Association of Stem Water Soluble Carbohydrates in Bread Wheat. PLoS ONE 2016, 11, e0164293. [Google Scholar] [CrossRef]
- Ovenden, B.; Milgate, A.; Wade, L.J.; Rebetzke, G.J.; Holland, J.B. Genome-Wide Associations for Water-Soluble Carbohydrate Concentration and Relative Maturity in Wheat Using SNP and DArT Marker Arrays. G3 Genes Genomes Genet. 2017, 7, 2821–2830. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.; Wu, J.; Yang, S.; Jin, Y.; Liu, J.; Yang, M.; Rasheed, A.; Zhang, Y.; Xia, X.; Jing, R.; et al. Genome-wide association analysis of stem water-soluble carbohydrate content in bread wheat. Theor. Appl. Genet. 2020, 133, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Ovenden, B.; Milgate, A.; Lisle, C.; Wade, L.J.; Rebetzke, G.J.; Holland, J.B. Selection for water-soluble carbohydrate accumulation and investigation of genetic x environment interactions in an elite wheat breeding population. Theor. Appl. Genet. 2017, 130, 2445–2461. [Google Scholar] [CrossRef] [PubMed]
- Sela, H.; Loutre, C.; Keller, B.; Schulman, A.; Nevo, E.; Korol, A.; Fahima, T. Rapid Linkage Disequilibrium Decay in the Lr10 Gene in Wild Emmer Wheat (Triticum dicoccoides) Populations. Theor. Appl. Genet. 2011, 122, 175–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oraguzie, N.; Rikkerink, E.; Gardiner, S.; Nihal De Silva, H. Association Mapping in Plants; Springer: New York, NY, USA, 2007; p. 278. [Google Scholar]
- The International Wheat Genome Sequencing Consortium (IWGSC); Appels, R.; Eversole, K.; Stein, N.; Feuillet, C.; Keller, B.; Rogers, J.; Pozniak, C.J.; Choulet, F.; Distelfeld, A.; et al. Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 2018, 361, eaar7191. [Google Scholar]
- Chen, H.; Semagn, K.; Iqbal, M.; Pazooki Moakhar, N.; Haile, T.; N’Diaye, A.; Yang, R.-C.; Hucl, P.; Pozniak, C.; Spaner, D. Genome-Wide Association Mapping of Genomic Regions Associated with Phenotypic Traits in Canadian Western Spring Wheat. Mol. Breed. 2017, 37, 1–17. [Google Scholar] [CrossRef]
- Ogbonnaya, F.C.; Rasheed, A.; Okechukwu, E.C.; Jighly, A.; Makdis, F.; Wuletaw, T.; Hagras, A.; Uguru, M.I.; Agbo, C.U. Genome-wide association study for agronomic and physiological traits in spring wheat evaluated in a range of heat prone environments. TAG Theor. Appl. Genet. 2017, 130, 1819–1835. [Google Scholar] [CrossRef]
- Komar, A. Single Nucleotide Polymorphisms; Humana Press: New York, NY, USA, 2009; p. 464. [Google Scholar]
- Yue, A.-Q.; Li, A.; Mao, X.-G.; Chang, X.-P.; Li, R.-Z.; Jing, R.-L. Single-nucleotide polymorphisms, mapping and association analysis of 1-FFT-A1 gene in wheat. J. Integr. Agric. 2017, 16, 789–799. [Google Scholar] [CrossRef]
- Zhang, J.; Huang, S.; Fosu-Nyarko, J.; Dell, B.; McNeil, M.; Waters, I.; Moolhuijzen, P.; Conocono, E.; Appels, R. The genome structure of the 1-FEH genes in wheat (Triticum aestivum L.): New markers to track stem carbohydrates and grain filling QTLs in breeding. Mol. Breed. 2008, 22, 339–351. [Google Scholar] [CrossRef]
- Dong, Y.; Zhang, Y.; Xiao, Y.; Yan, J.; Liu, J.; Wen, W.; Zhang, Y.; Jing, R.; Xia, X.; He, Z. Cloning of TaSST genes associated with water soluble carbohydrate content in bread wheat stems and development of a functional marker. TAG Theor. Appl. Genet. 2016, 129, 1061–1070. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, S.; Kamal, A.H.M.; Hossain, Z. Wheat proteomics: Proteome modulation and abiotic stress acclimation. Front. Plant Sci. 2014, 5, 684. [Google Scholar] [CrossRef] [Green Version]
- Rampino, P.; De Pascali, M.; De Caroli, M.; Luvisi, A.; De Bellis, L.; Piro, G.; Perrotta, C. Td4IN2: A drought-responsive durum wheat (Triticum durum Desf.) gene coding for a resistance like protein with serine/threonine protein kinase, nucleotide binding site and leucine rich domains. Plant Physiol. Biochem. 2017, 120, 223–231. [Google Scholar] [CrossRef]
- Chang, X.; Mao, X.; Jing, R.; Zhang, H.; Tian, S. TaSnRK2.4, an SNF1-type serine/threonine protein kinase of wheat (Triticum aestivum L.), confers enhanced multistress tolerance in Arabidopsis. J. Exp. Bot. 2009, 61, 683–696. [Google Scholar]
- Luan, P.; Balch, W.E.; Emr, S.D.; Burd, C.G. Molecular Dissection of Guanine Nucleotide Dissociation Inhibitor Function In Vivo. Rab-Independent Binding to Membranes and Role of Rab Recycling Factors. J. Biol. Chem. 1999, 274, 14806–14817. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; McCormick, S. A distinct mechanism regulating a pollen-specific guanine nucleotide exchange factor for the small GTPase Rop in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2007, 104, 18830–18835. [Google Scholar] [CrossRef] [Green Version]
- Zou, T.; Li, S.; Liu, M.; Wang, T.; Xiao, Q.; Chen, D.; Li, Q.; Liang, Y.; Zhu, J.; Liang, Y.; et al. An atypical strictosidine synthase, OsSTRL2, plays key roles in anther development and pollen wall formation in rice. Sci. Rep. 2017, 7, 6863. [Google Scholar] [CrossRef] [Green Version]
- Dutta, A.; Sen, J.; Deswal, R. New Evidences about Strictosidine Synthase (Str) Regulation by Salinity, Cold Stress and Nitric Oxide in Catharanthus roseus. J. Plant. Biochem. Biot. 2013, 22, 124–131. [Google Scholar] [CrossRef]
- Liu, Y.; Meng, Q.; Duan, X.; Zhang, Z.; Li, D. Effects of PEG-induced drought stress on regulation of indole alkaloid biosynthesis in Catharanthus roseus. J. Plant Interact. 2017, 12, 87–91. [Google Scholar] [CrossRef] [Green Version]
- Walter, S.; Kahla, A.; Arunachalam, C.; Perochon, A.; Khan, M.R.; Scofield, S.R.; Doohan, F.M. A wheat ABC transporter contributes to both grain formation and mycotoxin tolerance. J. Exp. Bot. 2015, 66, 2583–2593. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Cheung, M.-Y.; Li, M.-W.; Fu, Y.; Sun, Z.; Sun, S.-M.; Lam, H.-M. Rice hypersensitive induced reaction protein 1 (OsHIR1) associates with plasma membrane and triggers hypersensitive cell death. BMC Plant Biol. 2010, 10, 290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregorová, Z.; Kováčik, J.; Klejdus, B.; Maglovski, M.; Kuna, R.; Hauptvogel, P.; Matušíková, I. Drought-Induced Responses of Physiology, Metabolites, and PR Proteins in Triticum aestivum. J. Agric. Food Chem. 2015, 63, 8125–8133. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.; Hickey, L.T.; Stahl, A.; Werner, C.R.; Hayes, B.; Snowdon, R.J.; Voss-Fels, K.P. Exploring and Harnessing Haplotype Diversity to Improve Yield Stability in Crops. Front. Plant Sci. 2017, 8, 1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- N’Diaye, A.; Haile, J.K.; Cory, A.T.; Clarke, F.R.; Clarke, J.M.; Knox, R.E.; Pozniak, C.J. Single Marker and Haplotype-Based Association Analysis of Semolina and Pasta Colour in Elite Durum Wheat Breeding Lines Using a High-Density Consensus Map. PLoS ONE 2017, 12, e0170941. [Google Scholar]
- Yemm, E.W.; Willis, A.J. The estimation of carbohydrates in plant extracts by anthrone. Biochem. J. 1954, 57, 508–514. [Google Scholar] [CrossRef] [Green Version]
- SAS_Institute. SAS 9.4; SAS Institute: Cary, NC, USA, 2013. [Google Scholar]
- Lado, B.; Matus, I.; Rodríguez, A.; Inostroza, L.; Poland, J.; Belzile, F.; del Pozo, A.; Quincke, M.; Castro, M.; von Zitzewitz, J. Increased genomic prediction accuracy in wheat breeding through spatial adjustment of field trial data. G3 Genes Genomes Genet. 2013, 3, 2105–2114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef]
- Remington, D.L.; Thornsberry, J.M.; Matsuoka, Y.; Wilson, L.M.; Whitt, S.R.; Doebley, J.; Kresovich, S.; Goodman, M.M.; Buckler, E.S.T. Structure of linkage disequilibrium and phenotypic associations in the maize genome. Proc. Natl. Acad. Sci. USA 2001, 98, 11479–11484. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y. Discovering the false discovery rate. J. R. Stat. Soc. Ser. B Stat. Methodol. 2010, 72, 405–416. [Google Scholar] [CrossRef]
- Storey, J.D.; Tibshirani, R. Statistical significance for genomewide studies. Proc. Natl. Acad. Sci. USA 2003, 100, 9440–9445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinnwell, J.; Schaid, D. Haplo.Stats: Statistical Analysis of Haplotypes with Traits and Covariates when Linkage Phase Is Ambiguous. Available online: https://cran.r-project.org/web/packages/haplo.stats/index.html (accessed on 11 March 2021).
- Schaid, D.J.; Rowland, C.M.; Tines, D.E.; Jacobson, R.M.; Poland, G.A. Score tests for association between traits and haplotypes when linkage phase is ambiguous. Am. J. Hum. Genet. 2002, 70, 425–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howe, K.L.; Contreras-Moreira, B.; De Silva, N.; Maslen, G.; Akanni, W.; Allen, J.; Alvarez-Jarreta, J.; Barba, M.; Bolser, D.M.; Cambell, L.; et al. Ensembl Genomes 2020—Enabling non-vertebrate genomic research. Nucleic Acids Res. 2019, 48, D689–D695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binns, D.; Dimmer, E.; Huntley, R.; Barrell, D.; O’Donovan, C.; Apweiler, R. QuickGO: A web-based tool for Gene Ontology searching. Bioinformatics 2009, 25, 3045–3046. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | ANOVA (p-Value) | Variance Components (% σ2P) | Ratio | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Genotype | Site | Rep(Site) | Genotype X Site | σ2G | σ2GxS | σ2e | σ2G/σ2P | ||
| GY | <0.0001 | <0.0001 | <0.0001 | 0.0002 | 16.1 | 15.3 | 68.6 | 0.16 | |
| SWa | <0.0001 | <0.0001 | 0.147 | 0.2303 | 51.6 | 1.9 | 46.5 | 0.52 | |
| SWm | <0.0001 | 0.88 | 0.2302 | <0.0001 | 46.6 | 12.4 | 41.0 | 0.47 | |
| WSCa | 0.0115 | <0.0001 | 0.1025 | 0.3557 | 5.8 | 1.9 | 92.3 | 0.06 | |
| WSCm | 0.0003 | <0.0001 | 0.0032 | 0.0279 | 5.0 | 9.4 | 85.7 | 0.05 | |
| WSCCa | <0.0001 | <0.0001 | 0.0527 | 0.2968 | 29.0 | 2.0 | 69.1 | 0.29 | |
| WSCCm | <0.0001 | <0.0001 | 0.012 | 0.0003 | 17.9 | 15.3 | 66.8 | 0.18 | |
| WSCCa/m2 | <0.0001 | <0.0001 | 0.0006 | 0.0001 | 5.8 | 18.3 | 75.9 | 0.06 | |
| WSCCm/m2 | <0.0001 | <0.0001 | 0.0943 | 0.3662 | 13.5 | 1.5 | 84.9 | 0.14 | |
| ARWSC | <0.0001 | <0.0001 | 0.0874 | 0.9364 | 19.5 | 0.0 | 80.5 | 0.20 | |
| Trait a | Site/Condition b | |||||
|---|---|---|---|---|---|---|
| Cau 2011 | Cau 2012 | SRosFI2012 | SRosFI2011 | SRosMWS2011 | Total | |
| GY | 4 | 4 | ||||
| SWa | 2 | 2 | ||||
| SWm | 1 | 1 | 2 | |||
| WSCa | 22 | 1 | 23 | |||
| WSCm | 2 | 3 | 5 | |||
| WSCCa | 1 | 1 | 5 | 7 | ||
| WSCCm | 3 | 5 | 3 | 11 | ||
| WSCCa_m2 | 1 | 7 | 7 | 15 | ||
| WSCCm_m2 | 2 | 1 | 6 | 9 | ||
| ARWSC | 18 | 18 | ||||
| Total | 5 | 26 | 12 | 36 | 17 | 96 |
| Trait a | Site/Condition b | Marker | SNP | Chr. | p-Value | q-Value | R2 | Position | Protein | Functional Category | Ensembl Gene ID |
|---|---|---|---|---|---|---|---|---|---|---|---|
| WSCa | Cau 2012 | iniaGBS56801 | C/T | 3A | 2.5 × 10−5 | 0.0818 | 0.1232 | Gene | serine/threonine-protein kinase | Protein synthesis/modification | TRIAE_CS42_3AS_TGACv1_212383_AA0700440 |
| Cau 2012 | iniaGBS12206 | C/T | 4B | 0.0002 | 0.1136 | 0.1014 | Gene | Uncharacterized | TRIAE_CS42_4BL_TGACv1_320632_AA1045170 | ||
| Cau 2012 | iniaGBS21514 | A/G | 5D | 0.0003 | 0.1136 | 0.0981 | Gene | Guanosine nucleotide diphosphate dissociation inhibitor | Energy/metabolism | TRIAE_CS42_5DS_TGACv1_456722_AA1476920 | |
| Cau 2012 | iniaGBS13140 | A/T | 5B | 0.0004 | 0.1247 | 0.0908 | Gene | Strictosidine synthase | Energy/metabolism | TRIAE_CS42_5BL_TGACv1_404735_AA1309530 | |
| Cau 2012 | iniaGBS61502 | A/G | 1A | 0.0007 | 0.1247 | 0.0837 | Gene | ABC transporter C family member 10 | Transport | TRIAE_CS42_1AL_TGACv1_000028_AA0000820 | |
| Cau 2012 | iniaGBS86062 | C/G | 6B | 0.0007 | 0.1247 | 0.0861 | Gene | Hypersensitive induced response protein 3 | Cell disease/defense | TRIAE_CS42_5BL_TGACv1_404707_AA1308960 | |
| ARWSC | SRosFI2011 | iniaGBS83719 | G/T | 6D | 2.3 × 10−6 | 0.0017 | 0.1491 | Intergenic | Non-coding | ||
| SRosFI2011 | iniaGBS76557 | G/T | 6D | 6.8 × 10−6 | 0.0034 | 0.1373 | Gene | Uncharacterized | TRIAE_CS42_U_TGACv1_644569_AA2140450 | ||
| WSCCa/m2 | SRosFI2011 | iniaGBS52443 | A/G | 6B | 0.0001 | 0.1000 | 0.1248 | Intergenic | Non-coding | ||
| SRosFI2011 | iniaGBS52444 | G/T | 6B | 6.4 × 10−5 | 0.1000 | 0.1248 | Intergenic | Non-coding | |||
| SRosMWS201 | iniaGBS24966 | A/C | 7B | 0.0002 | 0.3181 | 0.0993 | Gene | Endoglucanase | Energy/metabolism | TRIAE_CS42_7BS_TGACv1_592132_AA1931340 |
| Site/Condition a | Trait | Position/Gene | GSSb | p-Value | q-Value | Significant Haplotypes | Freq. | Code c |
|---|---|---|---|---|---|---|---|---|
| SRosMWS2011 | WSCa | endoglucanase gene | 20.4 | 0.0024 | 0.0210 | AGACCCC | 0.09 | |
| WSCCa/m2 | endoglucanase gene | 18.9 | 0.0043 | 0.0220 | CACGTTG | 0.03 | ||
| Cau2012 | SWa | serine/threonine | 30.3 | 0.0043 | 0.0205 | GTGTTCAT | 0.07 | H1 |
| GTAGCTAC | 0.07 | H2 | ||||||
| TTAGCCAT | 0.03 | H3 | ||||||
| GTAGCTGT | 0.03 | H4 | ||||||
| SWm | serine/threonine | 23.2 | 0.0395 | 0.0403 | GCGTCCAT | 0.03 | ||
| WSCCm | serine/threonine | 13.0 | 0.0403 | 0.0403 | GCGTCCAT | 0.03 | ||
| WSCCa/m2 | serine/threonine | 16.5 | 0.0102 | 0.0205 | GTAGCTGT | 0.03 | ||
| SRosFI2011 | SWa | intergenic region | 20.4 | 0.0024 | 0.0215 | ACACCAGGGCCT | 0.04 | H1 |
| ACACCAGAACCT | 0.36 | H2 | ||||||
| CCGTTAGGGTTC | 0.11 | H3 | ||||||
| SWm | intergenic region | 18.9 | 0.0043 | 0.0210 | ACACCAGGGCCT | 0.04 | ||
| ACACCAGAATCT | 0.28 | |||||||
| CCGTTAGGGTTC | 0.11 | |||||||
| WSCCm/m2 | intergenic region | 12.9 | 0.0444 | 0.0494 | ACACCAGGGCCT | 0.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guerra, F.P.; Yáñez, A.; Matus, I.; del Pozo, A. Genome-Wide Association of Stem Carbohydrate Accumulation and Remobilization during Grain Growth in Bread Wheat (Triticum aestivum L.) in Mediterranean Environments. Plants 2021, 10, 539. https://doi.org/10.3390/plants10030539
Guerra FP, Yáñez A, Matus I, del Pozo A. Genome-Wide Association of Stem Carbohydrate Accumulation and Remobilization during Grain Growth in Bread Wheat (Triticum aestivum L.) in Mediterranean Environments. Plants. 2021; 10(3):539. https://doi.org/10.3390/plants10030539
Chicago/Turabian StyleGuerra, Fernando P., Alejandra Yáñez, Iván Matus, and Alejandro del Pozo. 2021. "Genome-Wide Association of Stem Carbohydrate Accumulation and Remobilization during Grain Growth in Bread Wheat (Triticum aestivum L.) in Mediterranean Environments" Plants 10, no. 3: 539. https://doi.org/10.3390/plants10030539