Microglia Activated by Excess Cortisol Induce HMGB1 Acetylation and Neuroinflammation in the Hippocampal DG Region of Mice Following Cold Exposure

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Serum and Hippocampus Corticosterone Assay
2.3. Brain Tissue Collection
2.4. Immunofluorescence Analysis
2.5. Immunohistochemistry
2.6. Nissl Staining
2.7. Terminal Deoxynucleotidyl Transferase dUTP Nick End Labeling (TUNEL) Assay
2.8. Cell Culture and Reagents
2.9. CORT Treatment
2.10. Cell Counting Kit (CCK)-8 Assay
2.11. Quantitative Real-Time (qRT-)PCR Analysis
2.12. Hippocampus Tissue and Cell Protein Extraction
2.13. Cytoplasmic and Nuclear Protein Extraction
2.14. Western Blot Analysis
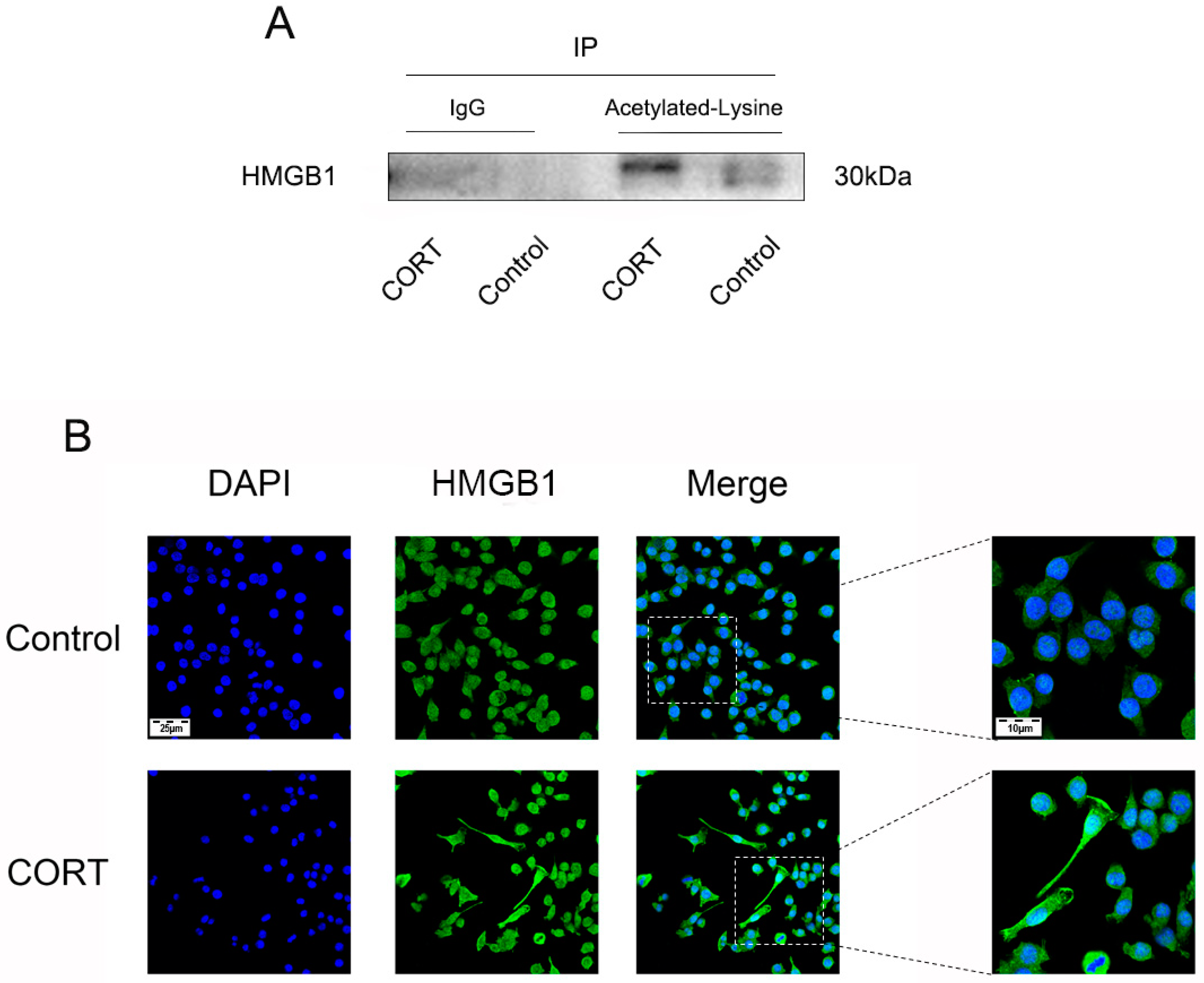
2.15. Immunocytochemistry
2.16. Co-Immunoprecipitation (Co-IP)
2.17. Statistical Analysis
3. Results
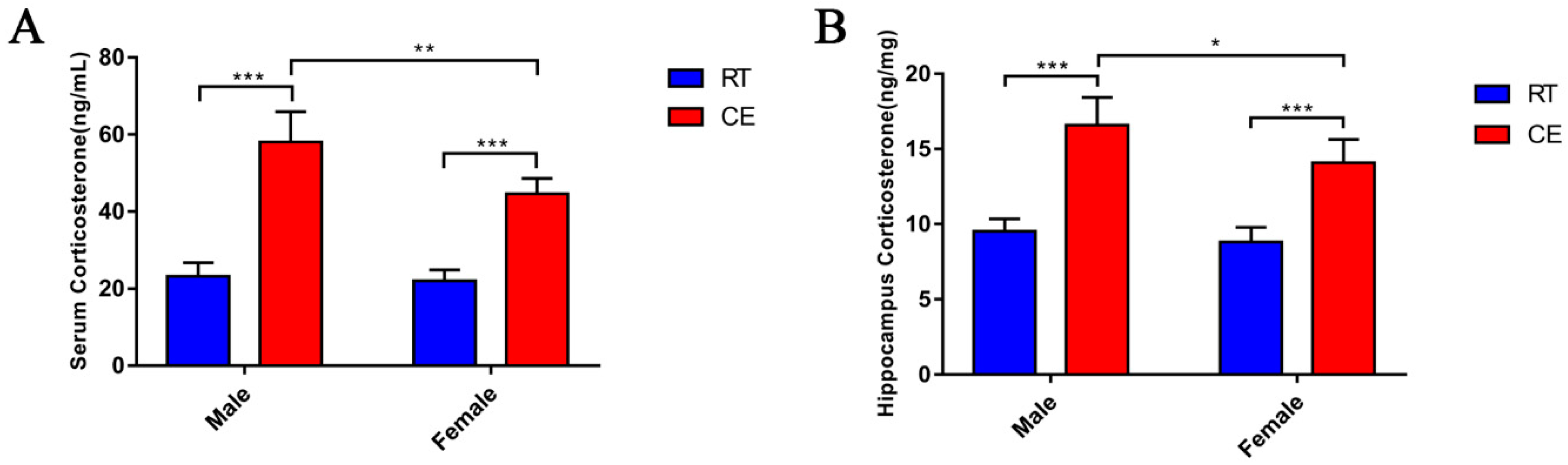
3.1. CORT Levels in the Mouse Serum and Hippocampus Following Cold Exposure
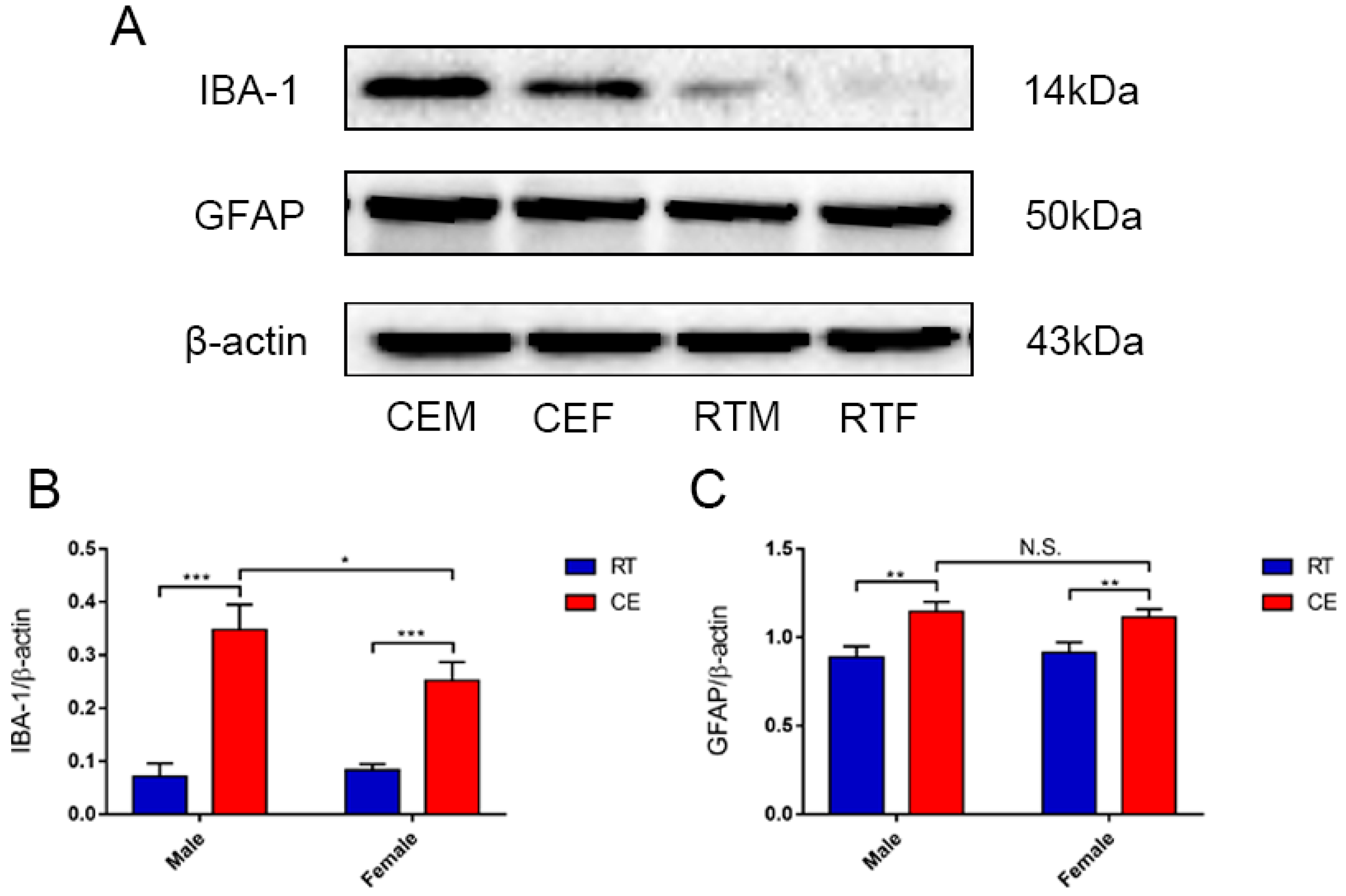
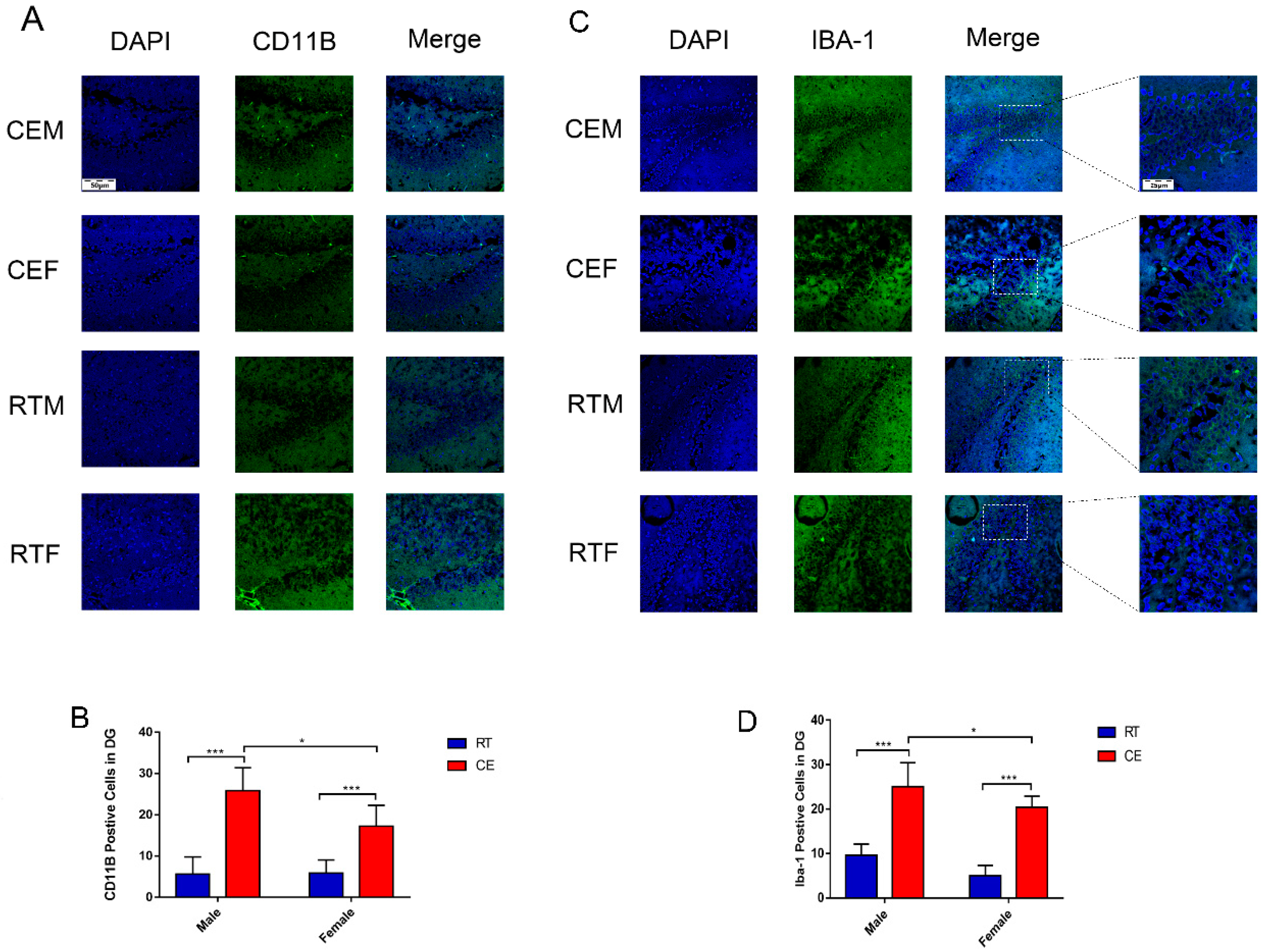
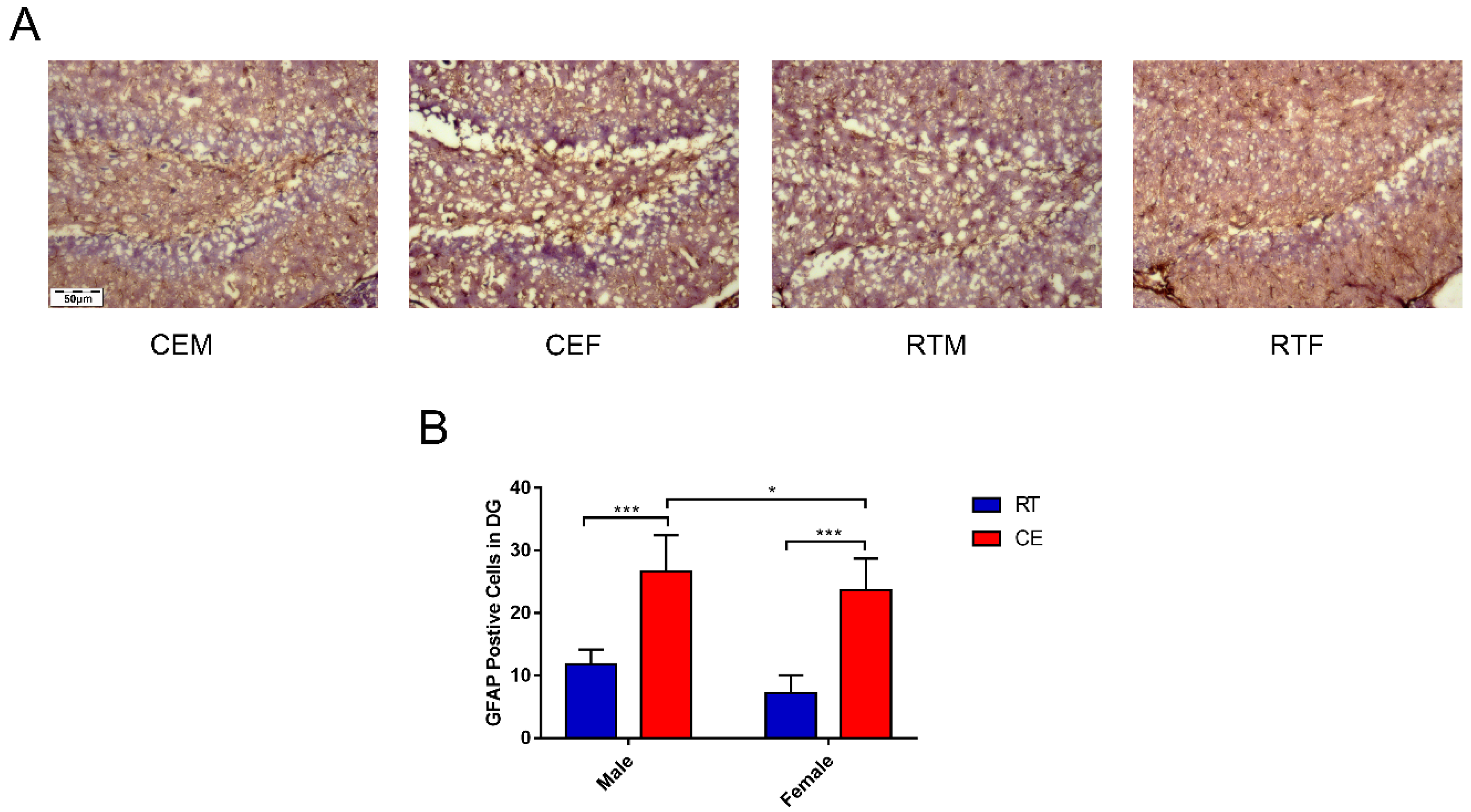
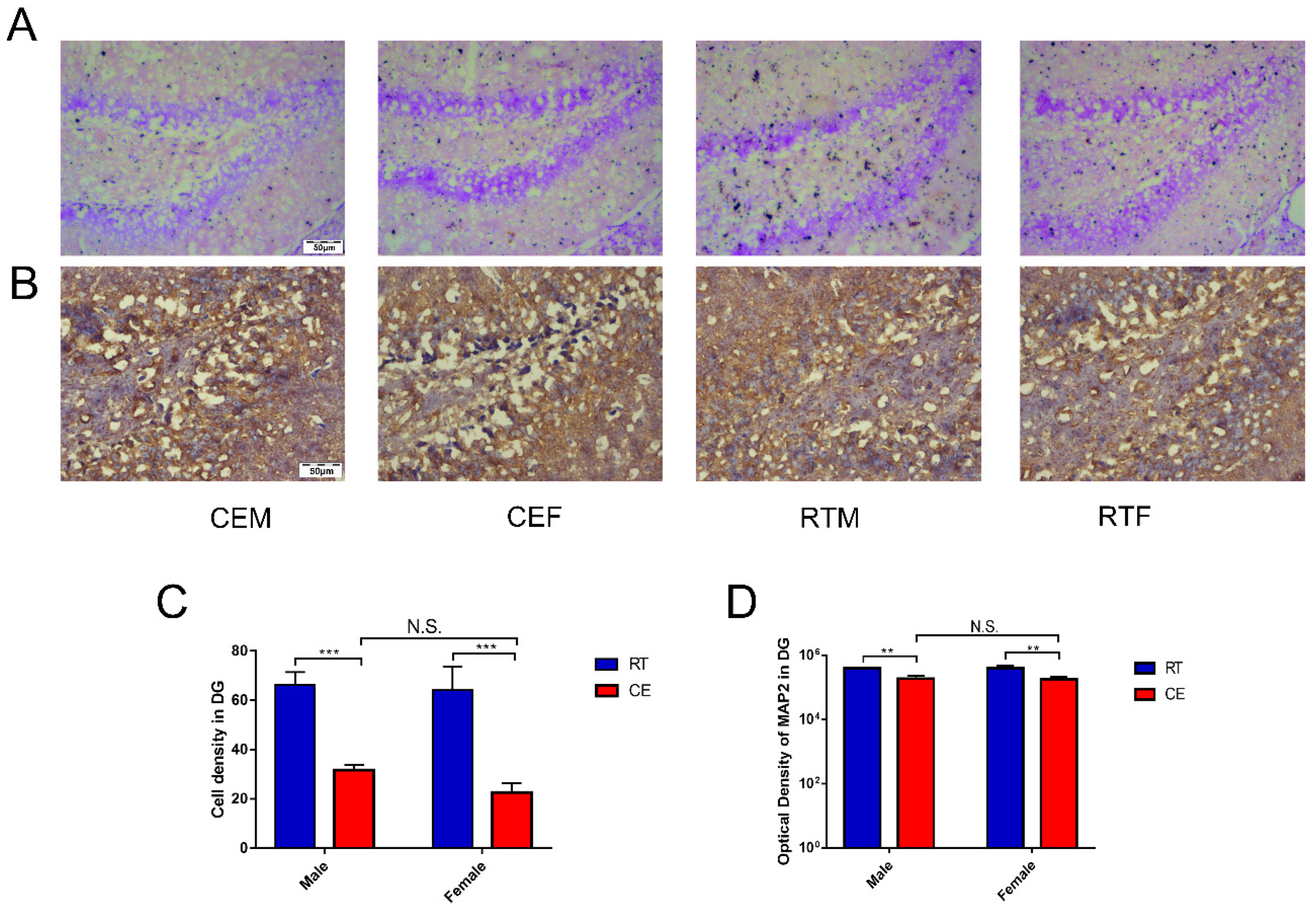
3.2. Cold Exposure Increases the Number of Glial Cells in the Hippocampus
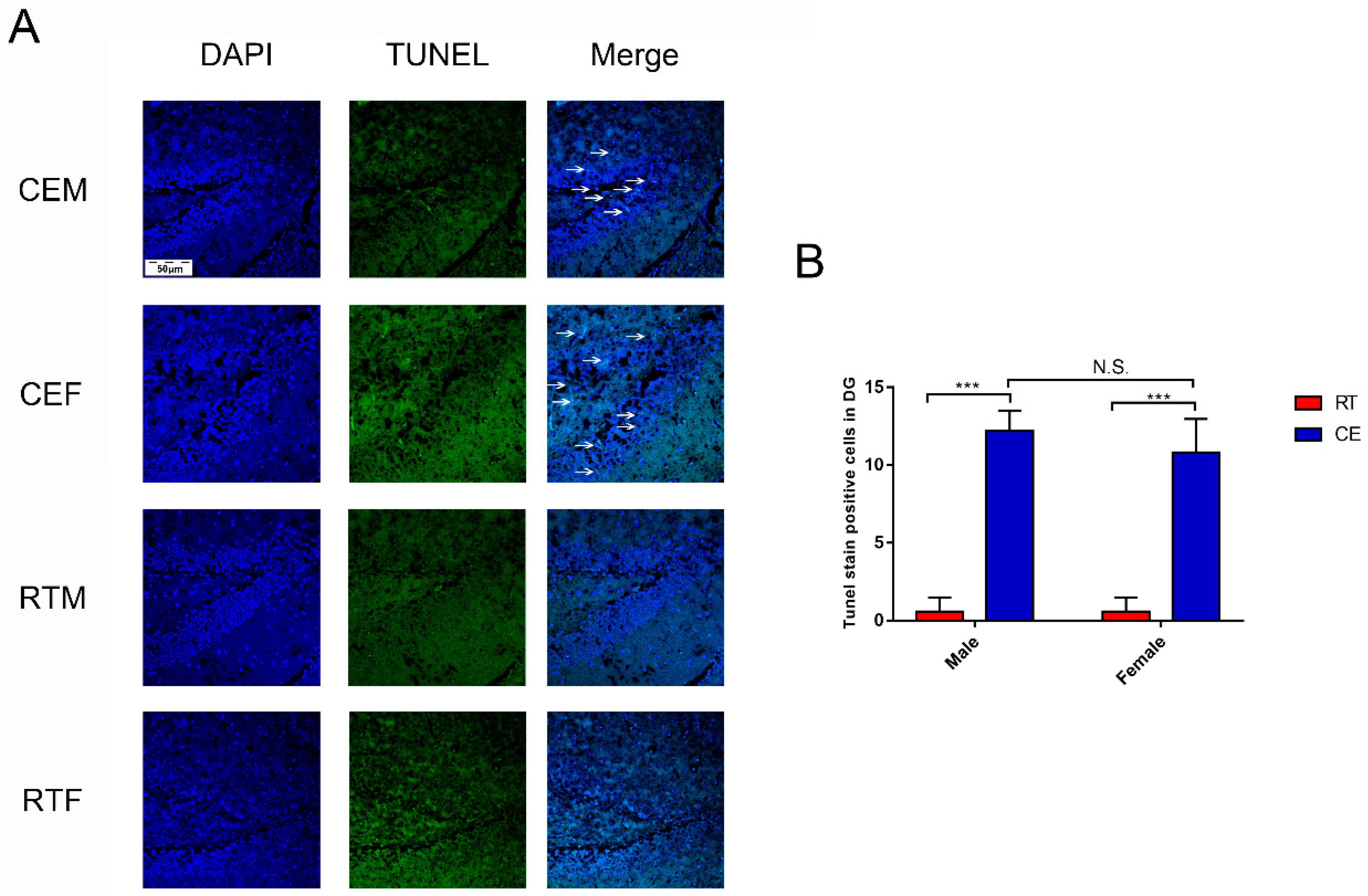
3.3. Cold Exposure Alters Hippocampus Homeostasis
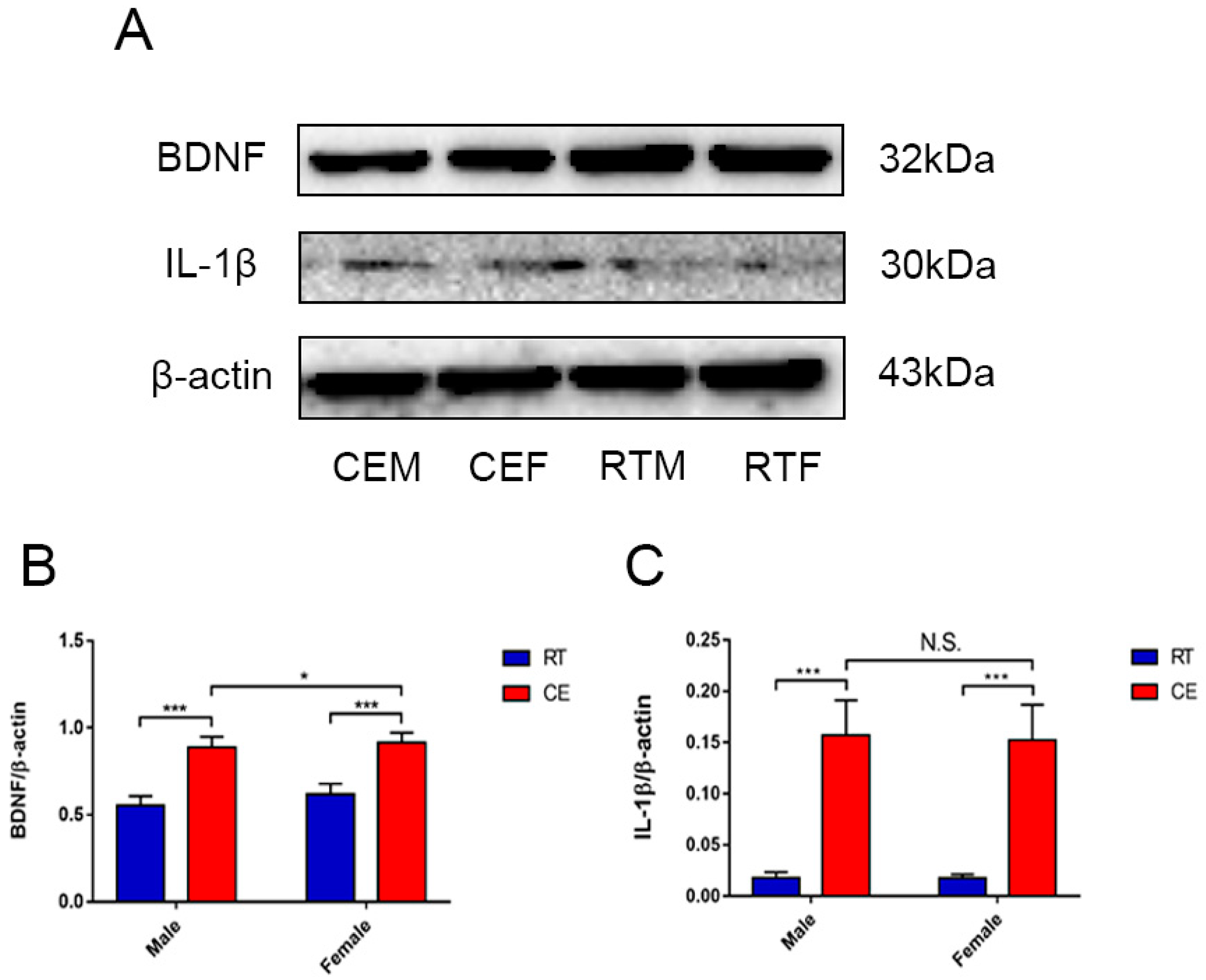
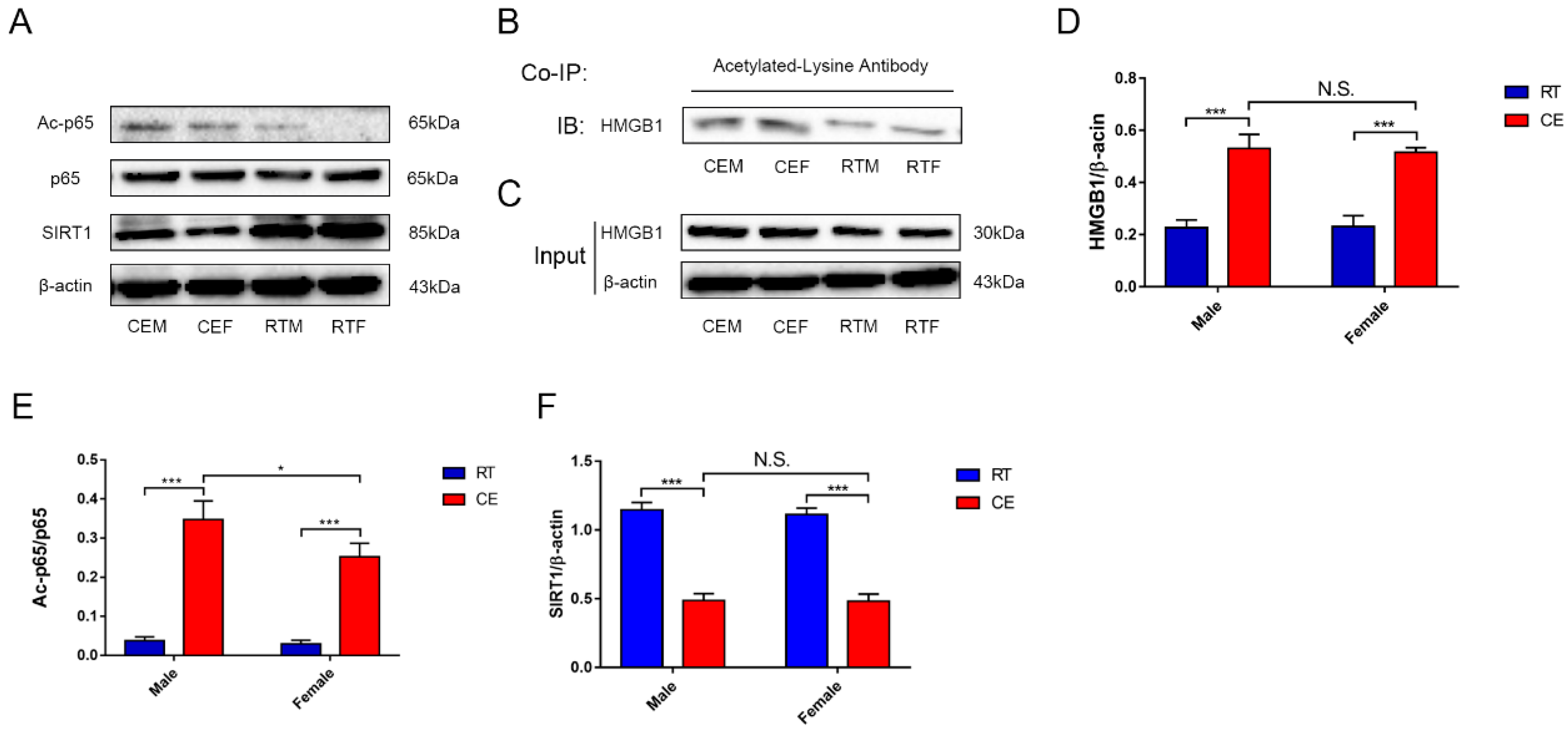
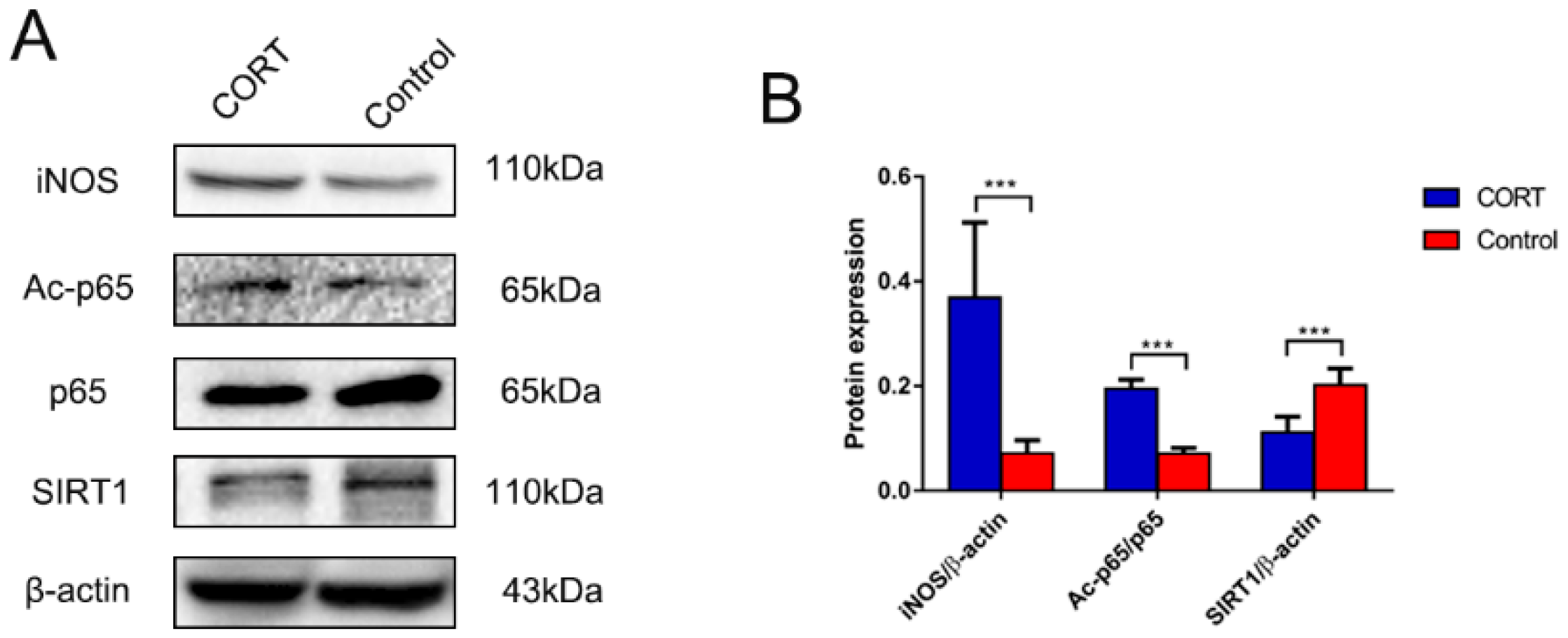
3.4. Cold Exposure Increases HMGB1 and p65 Acetylation in the Hippocampus
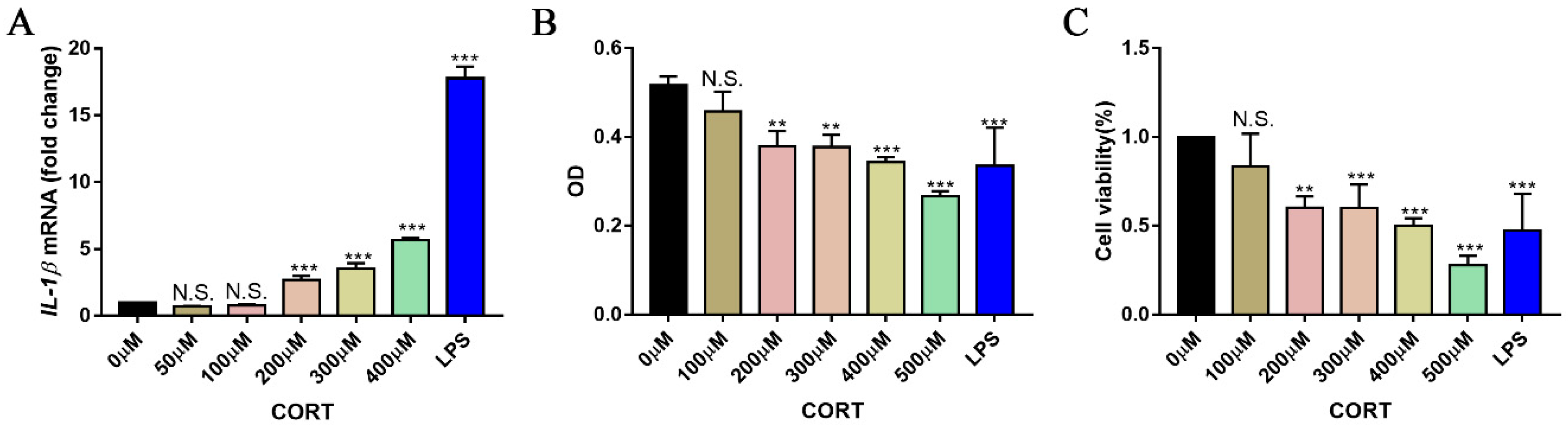
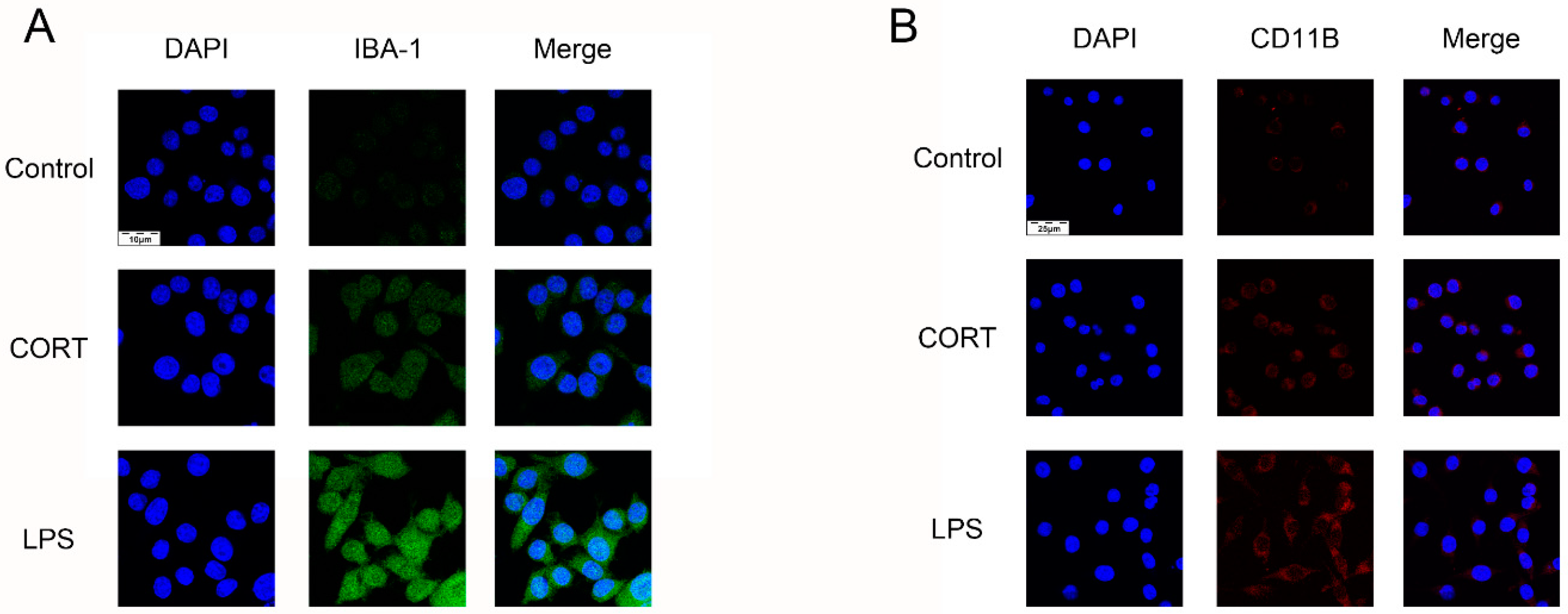
3.5. Excess CORT Activates BV2 Cells
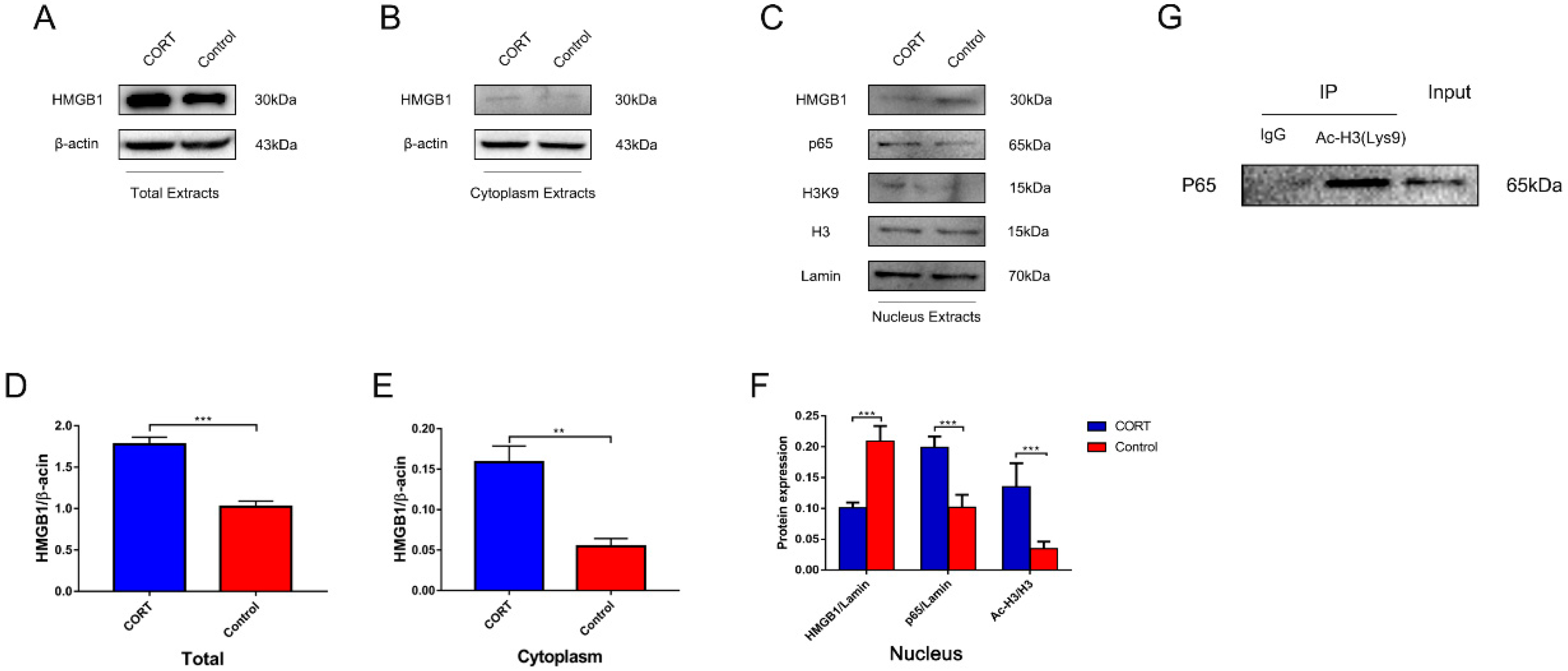
3.6. Excess CORT Promotes HMGB1 Activation and Secretion in BV2 Cells
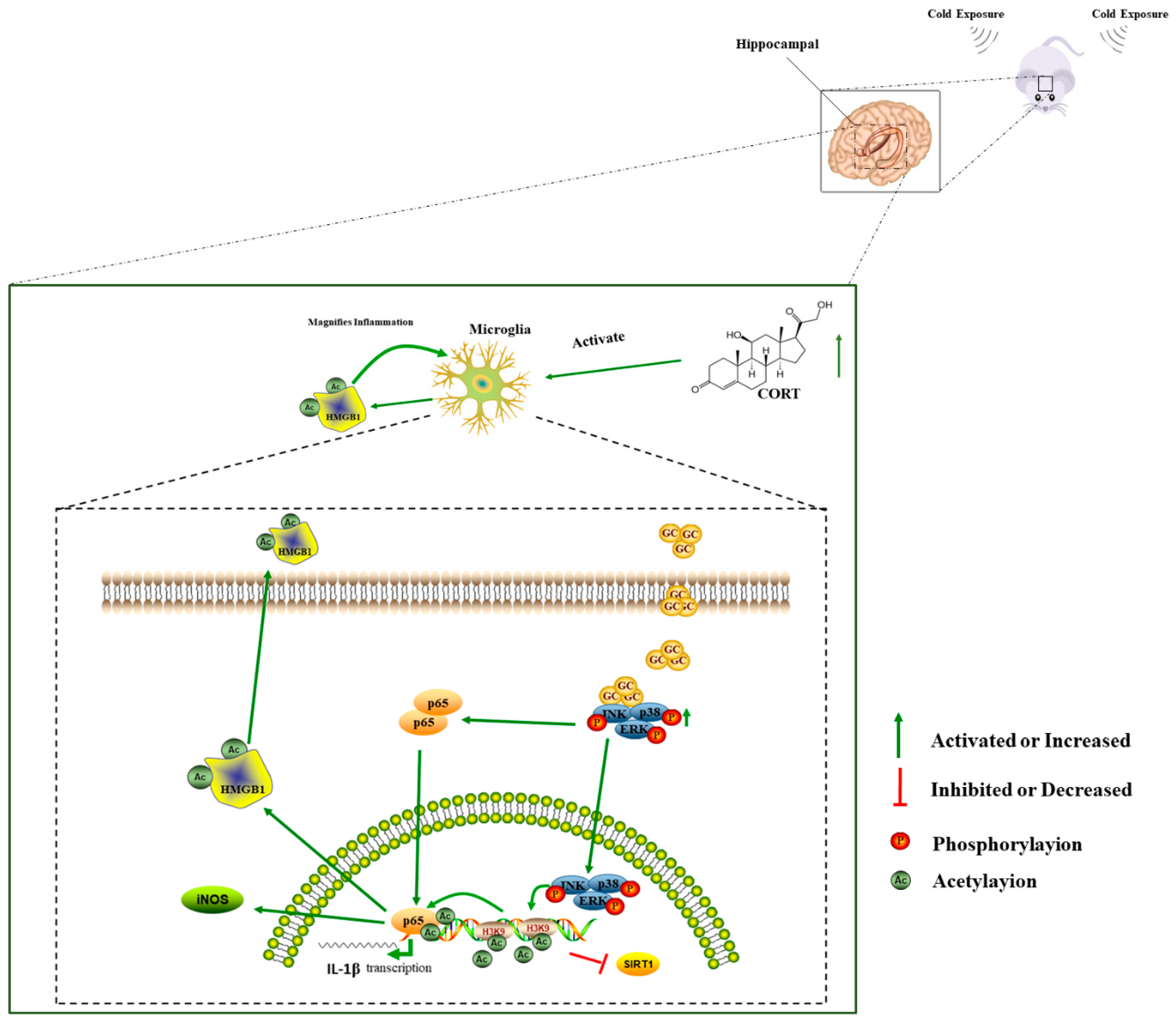
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bhat, A.H.; Dar, K.B.; Anees, S.; Zargar, M.A.; Masood, A.; Sofi, M.A.; Ganie, S.A. Oxidative stress, mitochondrial dysfunction and neurodegenerative diseases; a mechanistic insight. Biomed. Pharmacother. 2015, 74, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Marseglia, L.; Manti, S.; D’Angelo, G.; Nicotera, A.; Parisi, E.; Di Rosa, G.; Gitto, E.; Arrigo, T. Oxidative stress in obesity: A critical component in human diseases. Int. J. Mol. Sci. 2015, 16, 378–400. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.D.; Kogan, J.F.; Marrocco, J.; McEwen, B.S. Genomic and epigenomic mechanisms of glucocorticoids in the brain. Nat. Rev. Endocrinol. 2017, 13, 661. [Google Scholar] [CrossRef] [PubMed]
- Witte, A.V.; Köbe, T.; Kerti, L.; Rujescu, D.; Flöel, A. Impact of KIBRA polymorphism on memory function and the hippocampus in older adults. Neuropsychopharmacology 2016, 41, 781. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S.; Nasca, C.; Gray, J.D. Stress effects on neuronal structure: Hippocampus, amygdala, and prefrontal cortex. Neuropsychopharmacology 2016, 41, 3. [Google Scholar] [CrossRef] [PubMed]
- Mandairon, N.; Kuczewski, N.; Kermen, F.; Forest, J.; Midroit, M.; Richard, M.; Thevenet, M.; Sacquet, J.; Linster, C.; Didier, A. Opposite regulation of inhibition by adult-born granule cells during implicit versus explicit olfactory learning. Elife 2018, 7, e34976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brocca, M.E.; Pietranera, L.; Meyer, M.; Lima, A.; Roig, P.; De Kloet, E.R.; De Nicola, A.F. Mineralocorticoid receptor associates with pro-inflammatory bias in the hippocampus of spontaneously hypertensive rats. J. Neuroendocrinol. 2017, 29. [Google Scholar] [CrossRef]
- Gado, M.; Noll, M.; Heinrich, A.; Rauner, M.; Hofbauer, L.; Henneicke, H. Glucocorticoid-induced loss of cortical bone is exacerbated during cold-exposure in male mice. Osteologie 2019, 28, 16. [Google Scholar]
- Ziętak, M.; Kovatcheva-Datchary, P.; Markiewicz, L.H.; Ståhlman, M.; Kozak, L.P.; Bäckhed, F. Altered microbiota contributes to reduced diet-induced obesity upon cold exposure. Cell Metab. 2016, 23, 1216–1223. [Google Scholar] [CrossRef]
- Morton, G.J.; Muta, K.; Kaiyala, K.J.; Rojas, J.M.; Scarlett, J.M.; Matsen, M.E.; Nelson, J.T.; Acharya, N.K.; Piccinini, F.; Stefanovski, D. Evidence that the sympathetic nervous system elicits rapid, coordinated, and reciprocal adjustments of insulin secretion and insulin sensitivity during cold exposure. Diabetes 2017, 66, 823–834. [Google Scholar] [CrossRef]
- Tournissac, M.; Bourassa, P.; Martinez-Cano, R.D.; Vu, T.M.; Hebert, S.S.; Planel, E.; Calon, F. Repeated cold exposures protect a mouse model of Alzheimer’s disease against cold-induced tau phosphorylation. Mol. Metab. 2019, 22, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Lian, S.; Li, S.Z.; Guo, J.R.; Wang, J.F.; Wang, D.; Zhang, L.P.; Yang, H.M. GABAB receptor mediate hippocampal neuroinflammation in adolescent male and female mice after cold expose. Brain Res. Bull. 2018, 142, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Frank, M.G.; Weber, M.D.; Watkins, L.R.; Maier, S.F. Stress sounds the alarmin: The role of the danger-associated molecular pattern HMGB1 in stress-induced neuroinflammatory priming. Brain Behav. Immun. 2015, 48, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agresti, A.; Scaffidi, P.; Riva, A.; Caiolfa, V.R.; Bianchi, M.E. GR and HMGB1 interact only within chromatin and influence each other’s residence time. Mol. Cell 2005, 18, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Liu, H.; Zeng, Q.; Imperato, G.H.; Addorisio, M.E.; Li, J.; He, M.; Cheng, K.F.; Al-Abed, Y.; Harris, H.E. Inhibition of HMGB1/RAGE-mediated endocytosis by HMGB1 antagonist box A, anti-HMGB1 antibodies, and cholinergic agonists suppresses inflammation. Mol. Med. 2019, 25, 13. [Google Scholar] [CrossRef] [PubMed]
- Frank, M.G.; Weber, M.D.; Fonken, L.K.; Hershman, S.A.; Watkins, L.R.; Maier, S.F. The redox state of the alarmin HMGB1 is a pivotal factor in neuroinflammatory and microglial priming: A role for the NLRP3 inflammasome. Brain Behav. Immun. 2016, 55, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Zeng, H.; Wang, Q.; Yu, Q.; Wu, J.; Feng, Y.; Deng, P.; Zhang, H. Glycyrrhizin ameliorates inflammatory pain by inhibiting microglial activation-mediated inflammatory response via blockage of the HMGB1-TLR4-NF-kB pathway. Exp. Cell Res. 2018, 369, 112–119. [Google Scholar] [CrossRef]
- Chen, X.; Wu, S.; Chen, C.; Xie, B.; Fang, Z.; Hu, W.; Chen, J.; Fu, H.; He, H. Omega-3 polyunsaturated fatty acid supplementation attenuates microglial-induced inflammation by inhibiting the HMGB1/TLR4/NF-κB pathway following experimental traumatic brain injury. J. Neuroinflamm. 2017, 14, 143. [Google Scholar] [CrossRef]
- Xu, B.; Zang, S.C.; Li, S.Z.; Guo, J.R.; Wang, J.F.; Wang, D.; Zhang, L.P.; Yang, H.M.; Lian, S. HMGB1-mediated differential response on hippocampal neurotransmitter disorder and neuroinflammation in adolescent male and female mice following cold exposure. Brain Behav. Immun. 2019, 76, 223–235. [Google Scholar] [CrossRef]
- Cichon, M.; Chadzinska, M.; Ksiazek, A.; Konarzewski, M. Delayed effects of cold stress on immune response in laboratory mice. Proc. Biol. Sci. 2002, 269, 1493–1497. [Google Scholar] [CrossRef]
- Xu, B.; Lang, L.-m.; Li, S.-Z.; Guo, J.-R.; Wang, J.-F.; Wang, D.; Zhang, L.-P.; Yang, H.-M.; Lian, S. Cortisol Excess-Mediated Mitochondrial Damage Induced Hippocampal Neuronal Apoptosis in Mice Following Cold Exposure. Cells 2019, 8, 612. [Google Scholar] [CrossRef] [PubMed]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Münch, A.E.; Chung, W.-S.; Peterson, T.C. Neurotoxic reactive astrocytes are induced by activated microglia. Nature 2017, 541, 481. [Google Scholar] [CrossRef] [PubMed]
- Knierim, J.J. The hippocampus. Curr. Biol. 2015, 25, R1116–R1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gobinath, A.R.; Workman, J.L.; Chow, C.; Lieblich, S.E.; Galea, L.A. Maternal postpartum corticosterone and fluoxetine differentially affect adult male and female offspring on anxiety-like behavior, stress reactivity, and hippocampal neurogenesis. Neuropharmacology 2016, 101, 165–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derks, N.A.; Krugers, H.J.; Hoogenraad, C.C.; Joëls, M.; Sarabdjitsingh, R.A. Effects of early life stress on synaptic plasticity in the developing hippocampus of male and female rats. PLoS ONE 2016, 11, e0164551. [Google Scholar] [CrossRef] [PubMed]
- Santiago, A.R.; Bernardino, L.; Agudo-Barriuso, M.; Gonçalves, J. Microglia in health and disease: A double-edged sword. Mediat. Inflamm. 2017, 2017. [Google Scholar] [CrossRef] [PubMed]
- Gavrieli, Y.; Sherman, Y.; Ben-Sasson, S.A. Identification of programmed cell death in situ via specific labeling of nuclear DNA fragmentation. J. Cell Biol. 1992, 119, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Parent, J.M.; Vexler, Z.S.; Gong, C.; Derugin, N.; Ferriero, D.M. Rat forebrain neurogenesis and striatal neuron replacement after focal stroke. Ann. Neurol. 2002, 52, 802–813. [Google Scholar] [CrossRef]
- Cheng, X.; Yang, Y.-L.; Yang, H.; Wang, Y.-H.; Du, G.-H. Kaempferol alleviates LPS-induced neuroinflammation and BBB dysfunction in mice via inhibiting HMGB1 release and down-regulating TLR4/MyD88 pathway. Int. Immunopharmacol. 2018, 56, 29–35. [Google Scholar] [CrossRef]
- Tang, B.L. Sirt1 and the mitochondria. Mol. Cells 2016, 39, 87. [Google Scholar]
- Nakazawa, H.; Chang, K.; Shinozaki, S.; Yasukawa, T.; Ishimaru, K.; Yasuhara, S.; Yu, Y.-M.; Martyn, J.J.; Tompkins, R.G.; Shimokado, K. iNOS as a driver of inflammation and apoptosis in mouse skeletal muscle after burn injury: Possible involvement of Sirt1 S-nitrosylation-mediated acetylation of p65 NF-κB and p53. PLoS ONE 2017, 12, e0170391. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Leak, R.K.; Shi, Y.; Suenaga, J.; Gao, Y.; Zheng, P.; Chen, J. Microglial and macrophage polarization—New prospects for brain repair. Nat. Rev. Neurol. 2015, 11, 56. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.-W.; Lee, S.-A.; Shin, J.-M.; Park, I.-H.; Lee, H.-M. Glucocorticoids ameliorate TGF-β1-mediated epithelial-to-mesenchymal transition of airway epithelium through MAPK and Snail/Slug signaling pathways. Sci. Rep. 2017, 7, 3486. [Google Scholar] [CrossRef] [PubMed]
- Boncompagni, S.; Arthurton, L.; Akujuru, E.; Pearson, T.; Steverding, D.; Protasi, F.; Mutungi, G. Membrane glucocorticoid receptors are localised in the extracellular matrix and signal through the MAPK pathway in mammalian skeletal muscle fibres. J. Physiol. 2015, 593, 2679–2692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.; Min, J.-S.; Kim, B.; Chae, U.-B.; Yun, J.W.; Choi, M.-S.; Kong, I.-K.; Chang, K.-T.; Lee, D.-S. Mitochondrial ROS govern the LPS-induced pro-inflammatory response in microglia cells by regulating MAPK and NF-κB pathways. Neurosci. Lett. 2015, 584, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Li, S.; Wang, Y.; Shen, J.; Zhou, Y.; Li, H.; Yu, X.; Mao, H. Acetylation of HMGB1 by JNK1 Signaling Promotes LPS-Induced Peritoneal Mesothelial Cells Apoptosis. Biomed. Res. Int. 2018, 2018. [Google Scholar] [CrossRef] [PubMed]
- Khavinson, V.K.; Kuznik, B.; Tarnovskaya, S.; Linkova, N. Peptides and CCL11 and HMGB1 as molecular markers of aging: Literature review and own data. Adv. Gerontol. 2015, 5, 133–140. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Primer | Lengh (bp) |
|---|---|---|
| IL-1β (Forward) | 5’-GCTGCTTCCAAACCTTTGAC-3’ | 121 |
| IL-1β (Reverse) | 5’-AGCTTCTCCACAGCCACAAT-3’ | |
| β-actin (Forward) | 5’-GTCAGGTCATCACTATCGGCAAT-3’ | 147 |
| β-actin (Reverse) | 5’-AGAGGTCTTTACGGATGTCAACGT-3’ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, B.; Lang, L.-M.; Li, S.-Z.; Guo, J.-R.; Wang, J.-F.; Yang, H.-M.; Lian, S. Microglia Activated by Excess Cortisol Induce HMGB1 Acetylation and Neuroinflammation in the Hippocampal DG Region of Mice Following Cold Exposure. Biomolecules 2019, 9, 426. https://doi.org/10.3390/biom9090426
Xu B, Lang L-M, Li S-Z, Guo J-R, Wang J-F, Yang H-M, Lian S. Microglia Activated by Excess Cortisol Induce HMGB1 Acetylation and Neuroinflammation in the Hippocampal DG Region of Mice Following Cold Exposure. Biomolecules. 2019; 9(9):426. https://doi.org/10.3390/biom9090426
Chicago/Turabian StyleXu, Bin, Li-Min Lang, Shi-Ze Li, Jing-Ru Guo, Jian-Fa Wang, Huan-Min Yang, and Shuai Lian. 2019. "Microglia Activated by Excess Cortisol Induce HMGB1 Acetylation and Neuroinflammation in the Hippocampal DG Region of Mice Following Cold Exposure" Biomolecules 9, no. 9: 426. https://doi.org/10.3390/biom9090426