Procyanidin B2 3″-O-gallate Isolated from Reynoutria elliptica Prevents Glutamate-Induced HT22 Cell Death by Blocking the Accumulation of Intracellular Reactive Oxygen Species

 ,
,  and
and 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Extraction and Isolation
2.3. Quantitative Analysis of Procyanidin B2 3″-O-gallate
2.4. Cell Culture and Treatment
2.5. Determination of Cell Viability
2.6. Determination of Antioxidant Activity and Intracellular ROS Levels
2.7. Determination of Apoptotic Cells
2.8. Western Blot Analysis
2.9. Statistical Analysis
3. Results and Discussions
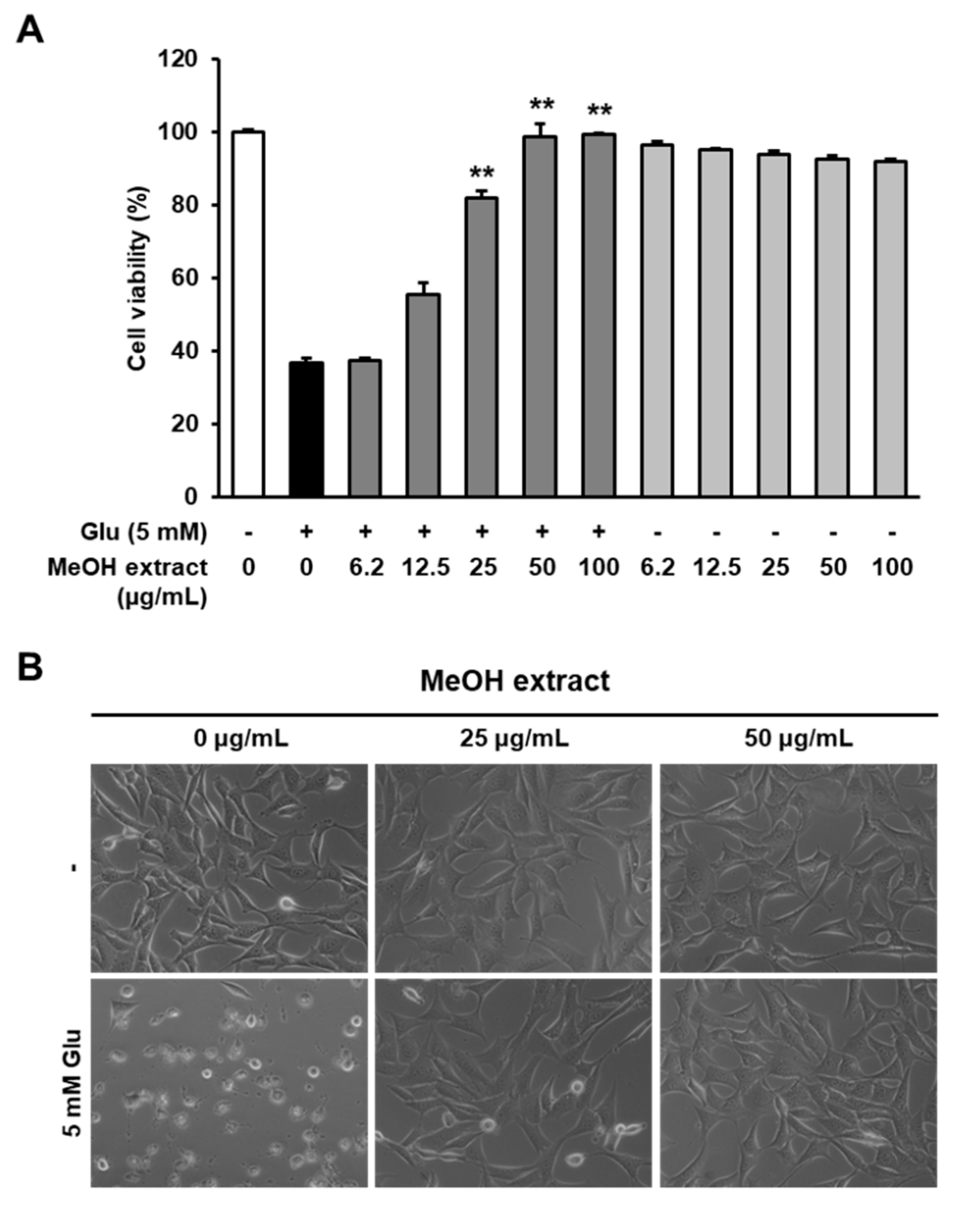
3.1. Neuroprotective Effects of the MeOH Extract from R. elliptica Seeds on Glutamate-Induced Excitotoxicity in an HT22 Cell Line
3.2. LC/MS-Guided Isolation and Identification of the Active Compound from the MeOH Extract
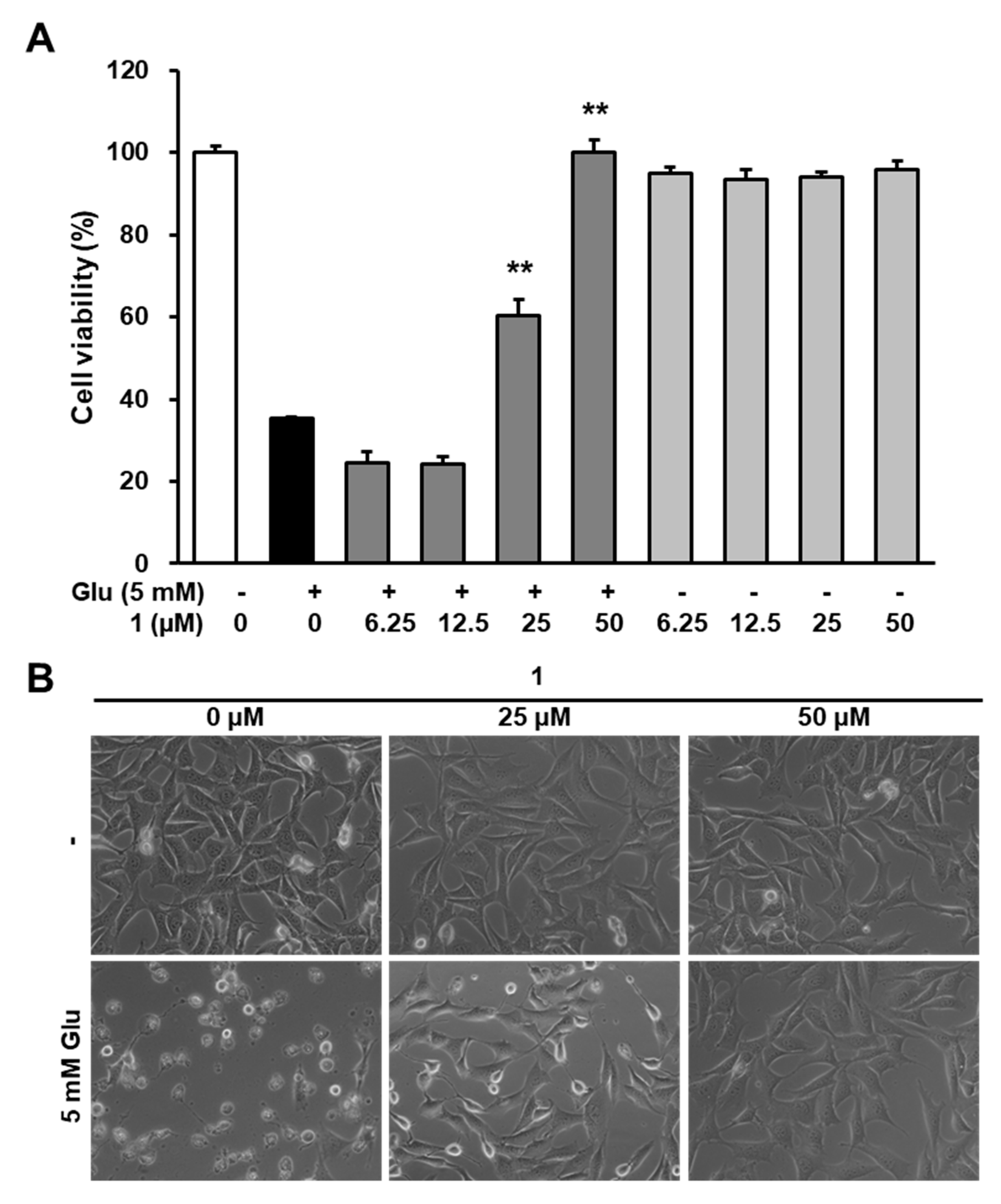
3.3. Neuroprotective Efficacy of 1 Against Glutamate-Induced HT22 Cell Death
3.4. Effects of 1 on Glutamate-Induced Reactive Oxygen Species (ROS) Accumulation
3.5. Inhibitory Effect of 1 on Glutamate-Induced Phosphorylation of Mitogen-Activated Protein Kinases (MAPKs) in HT22 Cells
3.6. Anti-Apoptotic Effects of 1 in HT22 Cells
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yuan, J.; Lipinski, M.; Degterev, A. Diversity in the mechanisms of neuronal cell death. Neuron 2003, 40, 401–413. [Google Scholar] [CrossRef]
- Pankevich, D.E.; Altevogt, B.M.; Dunlop, J.; Gage, F.H.; Hyman, S.E. Improving and accelerating drug development for nervous system disorders. Neuron 2014, 84, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Rehman, M.U.; Wali, A.F.; Ahmad, A.; Shakeel, S.; Rasool, S.; Ali, R.; Rashid, S.M.; Madkhali, H.; Genai, M.A.; Khan, R. Neuroprotective strategies for neurological disorders by natural products: An update. Curr. Neuropharmacol. 2019, 17, 247–267. [Google Scholar] [CrossRef] [PubMed]
- Dadhania, V.P.; Trivedi, P.P.; Vikram, A.; Tripathi, D.N. Nutraceuticals against neurodegeneration: A mechanistic insight. Curr. Neuropharmacol. 2016, 14, 627–640. [Google Scholar] [CrossRef] [PubMed]
- Jo, H.; Lee, H.J.; Kim, C.Y.; Son, J.K.; Jung, S.H. 8-Hydroxycalamenene isolated from the rhizomes of Reynoutria elliptica exerts neuroprotective effects both in vitro and in vivo. Food Chem. Toxicol. 2013, 51, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.; Choi, T.W.; Kim, C.; Nam, D.; Lee, S.G.; Jang, H.J.; Lee, J.H.; Um, J.Y.; Jung, S.H.; Shim, B.S.; et al. Anti-inflammatory activities of Reynoutria elliptica through suppression of mitogen-activated protein kinases and nuclear factor-kappa B activation pathways. Immunopharmacol. Immunotoxicol. 2012, 34, 454–464. [Google Scholar] [CrossRef] [PubMed]
- Nam, D.; Song, J.; Kim, S.M.; Chiang, S.Y.; Kim, J.S.; Chung, W.S.; Jang, H.J.; Jung, S.H.; Na, Y.S.; Kim, S.H.; et al. 8-Hydrocalamenene, Derived from Reynoutria elliptica, suppresses constitutive STAT3 activation, inhibiting proliferation and enhancing chemosensitization of human multiple myeloma cells. J. Med. Food 2014, 17, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Kim, S.I.; Lee, K.B.; Yoo, Y.C.; Ryu, S.Y.; Song, K.S. Neuraminidase inhibitors from Reynoutria elliptica. Arch. Pharmacal. Res. 2003, 26, 367–374. [Google Scholar] [CrossRef]
- Zhao, Y.; Chen, M.X.; Kongstad, K.T.; Jager, A.K.; Staerk, D. Potential of Polygonum cuspidatum root as an antidiabetic food: Dual high-resolution alpha-glucosidase and PTP1B inhibition profiling combined with HPLC-HRMS and NMR for identification of antidiabetic constituents. J. Agric. Food Chem. 2017, 65, 4421–4427. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Qin, R.X.; Li, X.L.; Zhou, H. Botany, phytochemistry, pharmacology, and potential application of Polygonum cuspidatum Sieb.et Zucc.: A review. J. Ethnopharmacol. 2013, 148, 729–745. [Google Scholar]
- Fan, P.H.; Hostettmann, K.; Lou, H.X. Allelochemicals of the invasive neophyte Polygonum cuspidatum Sieb. & Zucc. (Polygonaceae). Chemoecology 2010, 20, 223–227. [Google Scholar]
- Shan, B.; Cai, Y.Z.; Brooks, J.D.; Corke, H. Antibacterial properties of Polygonum cuspidatum roots and their major bioactive constituents. Food Chem. 2008, 109, 530–537. [Google Scholar] [CrossRef]
- Alejo-Armijo, A.; Glibota, N.; Frias, M.P.; Altarejos, J.; Galvez, A.; Salido, S.; Ortega-Morente, E. Synthesis and evaluation of antimicrobial and antibiofilm properties of A-type procyanidin analogues against resistant bacteria in food. J. Agric. Food Chem. 2018, 66, 2151–2158. [Google Scholar] [CrossRef] [PubMed]
- Rong, S.; Zhao, S.Q.; Xu, K.; Zhang, L.; Zhao, Y.T.; Xiao, X.; Bao, W.; Liu, L.G. Procyanidins extracted from the litchi pericarp attenuate atherosclerosis and hyperlipidemia associated with consumption of a high fat diet in apolipoprotein-E knockout mice. Biomed. Pharmacoether. 2018, 97, 1639–1644. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Deep, G.; Wempe, M.F.; Surek, J.; Kumar, A.; Agarwal, R.; Agarwal, C. Procyanidin B2 3,3″-di-O-gallate induces oxidative stress-mediated cell death in prostate cancer cells via inhibiting MAP kinase phosphatase activity and activating ERK1/2 and AMPK. Mol. Carcinog. 2018, 57, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, M.; Nakanishi, I.; Matsumoto, K.; Fukuhara, K. Enhanced radical scavenging activity of a procyanidin B3 analogue comprised of a dimer of planar catechin. Bioorg. Med. Chem. Lett. 2017, 27, 5010–5013. [Google Scholar] [CrossRef] [PubMed]
- Dudek, M.K.; Gliński, V.B.; Davey, M.H.; Sliva, D.; Kaźmierski, S.; Gliński, J.A. Trimeric and tetrameric A-type procyanidins from peanut skins. J. Nat. Prod. 2017, 80, 415–426. [Google Scholar] [CrossRef] [PubMed]
- Czerwinska, M.E.; Dudek, M.K.; Pawlowska, K.A.; Prus, A.; Ziaja, M.; Granica, S. The influence of procyanidins isolated from small-leaved lime flowers (Tilia cordata Mill.) on human neutrophils. Fitoterapia 2018, 127, 115–122. [Google Scholar] [CrossRef]
- Yin, C.C.; Luo, X.P.; Duan, Y.Q.; Duan, W.Y.; Zhang, H.H.; He, Y.Q.; Sun, G.B.; Sun, X.B. Neuroprotective effects of lotus seedpod procyanidins on extremely low frequency electromagnetic field-induced neurotoxicity in primary cultured hippocampal neurons. Biomed. Pharmacother. 2016, 82, 628–639. [Google Scholar] [CrossRef]
- Baek, S.C.; Choi, E.; Eom, H.J.; Jo, M.S.; Kim, S.; So, H.M.; Kim, S.H.; Kang, K.S.; Kim, K.H. LC/MS-based analysis of bioactive compounds from the bark of Betula platyphylla var. japonica and their effects on regulation of adipocyte and osteoblast differentiation. Nat. Prod. Sci. 2018, 24, 235–240. [Google Scholar]
- So, H.M.; Eom, H.J.; Lee, D.; Kim, S.; Kang, K.S.; Lee, I.K.; Baek, K.H.; Park, J.Y.; Kim, K.H. Bioactivity evaluations of betulin identified from the bark of Betula platyphylla var. japonica for cancer therapy. Arch. Pharmacal. Res. 2018, 41, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Kim, T.; Kim, K.H.; Ham, J.; Jang, T.S.; Kang, K.S.; Lee, J.W. Evaluation of guggulsterone derivatives as novel kidney cell protective agents against cisplatin-induced nephrotoxicity. Bioorg. Med. Chem. Lett. 2017, 27, 3156–3161. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Lee, S.; Roh, H.; Song, S.; Ryoo, R.; Pang, C.; Baek, K.; Kim, K.H. Cytotoxic constituents from the sclerotia of Poria cocos against human lung adenocarcinoma cells by inducing mitochondrial apoptosis. Cells 2018, 7, 116. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.K.; Roh, H.S.; Yu, J.S.; Kwon, D.J.; Kim, S.Y.; Baek, K.H.; Kim, K.H. A novel cytotoxic activity of the fruit of Sorbus commixta against human lung cancer cells and isolation of the major constituents. J. Funct. Food 2017, 30, 1–7. [Google Scholar] [CrossRef]
- Park, J.Y.; Lee, Y.K.; Lee, D.S.; Yoo, J.E.; Shin, M.S.; Yamabe, N.; Kim, S.N.; Lee, S.; Kim, K.H.; Lee, H.J.; et al. Abietic acid isolated from pine resin (Resina Pini) enhances angiogenesis in HUVECs and accelerates cutaneous wound healing in mice. J. Ethnopharmacol. 2017, 203, 279–287. [Google Scholar] [CrossRef]
- Shin, S.H.; Lee, S.R.; Lee, E.; Kim, K.H.; Byun, S. Caffeic acid phenethyl ester from the twigs of Cinnamomum cassia inhibits malignant cell transformation by inducing c-Fos degradation. J. Nat. Prod. 2017, 80, 2124–2130. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.S.; Kim, J.H.; Lee, S.; Jung, K.; Kim, K.H.; Cho, J.Y. Src/Syk-targeted anti-inflammatory actions of triterpenoidal saponins from Gac (Momordica cochinchinensis) seeds. Am. J. Chin. Med. 2017, 45, 459–473. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.S.; Moon, E.; Kim, K.H. A new cerebroside from the twigs of Lindera glauca (Sieb. et Zucc.) Blume. Bioorg. Chem. 2017, 74, 122–125. [Google Scholar] [CrossRef]
- Yu, J.S.; Roh, H.S.; Baek, K.H.; Lee, S.; Kim, S.; So, H.M.; Moon, E.; Pang, C.; Jang, T.S.; Kim, K.H. Bioactivity-guided isolation of ginsenosides from Korean Red Ginseng with cytotoxic activity against human lung adenocarcinoma cells. J. Ginseng Res. 2018, 42, 562–570. [Google Scholar] [CrossRef]
- Tarascou, I.; Barathieu, K.; Simon, C.; Ducasse, M.A.; Andre, Y.; Fouquet, E.; Dufourc, E.J.; de Freitas, V.; Lqguerre, M.; Pianet, I. A 3D structural and conformational study of procyanidin dimers in water and hydro-alcoholic media as viewed by NMR and molecular modeling. Magn. Reson. Chem. 2006, 44, 868–880. [Google Scholar] [CrossRef]
- Kong, X.; Guan, J.; Gong, S.; Wang, R. Neuroprotective effects of grape seed procyanidin extract on ischemia-reperfusion brain injury. Chin. Med. Sci. J. 2017, 32, 92–99. [Google Scholar] [PubMed]
- Sutcliffe, T.C.; Winter, A.N.; Punessen, N.C.; Linseman, D.A. Procyanidin B2 Protects Neurons from Oxidative, Nitrosative, and Excitotoxic Stress. Antioxidants 2017, 6, 77. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Yue, Y.; Li, J.; Li, Z.; Li, X.; Niu, Y.; Xiang, J.; Ding, H. Procyanidin B2 attenuates neurological deficits and blood-brain barrier disruption in a rat model of cerebral ischemia. Mol. Nutr. Food Res. 2015, 59, 1930–1941. [Google Scholar] [CrossRef] [PubMed]
- Sayre, L.M.; Perry, G.; Smith, M.A. Oxidative stress and neurotoxicity. Chem. Res. Toxicol. 2008, 21, 172–188. [Google Scholar] [CrossRef] [PubMed]
- Ankarcrona, M.; Dypbukt, J.M.; Bonfoco, E.; Zhivotovsky, B.; Orrenius, S.; Lipton, S.A.; Nicotera, P. Glutamate-induced neuronal death: A succession of necrosis or apoptosis depending on mitochondrial function. Neuron 1995, 15, 961–973. [Google Scholar] [CrossRef] [Green Version]
- Tan, S.; Wood, M.; Maher, P. Oxidative stress induces a form of programmed cell death with characteristics of both apoptosis and necrosis in neuronal cells. J. Neurochem. 1998, 71, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Cargnello, M.; Roux, P.P. Activation and function of the MAPKs and their substrates, the MAPK-activated protein kinases. Microbiol Mol. Biol. Rev. 2011, 75, 50–83. [Google Scholar] [CrossRef]
- Stanciu, M.; Wang, Y.; Kentor, R.; Burke, N.; Watkins, S.; Kress, G.; Reynolds, I.; Klann, E.; Angiolieri, M.R.; Johnson, J.W.; et al. Persistent activation of ERK contributes to glutamate-induced oxidative toxicity in a neuronal cell line and primary cortical neuron cultures. J. Biol. Chem. 2000, 275, 12200–12206. [Google Scholar] [CrossRef]
- Bratton, D.L.; Fadok, V.A.; Richter, D.A.; Kailey, J.M.; Guthrie, L.A.; Henson, P.M. Appearance of phosphatidylserine on apoptotic cells requires calcium-mediated nonspecific flip-flop and is enhanced by loss of the aminophospholipid translocase. J. Biol. Chem. 1997, 272, 26159–26165. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, J.H.; Kim, S.; Yu, J.S.; Park, D.H.; Kim, S.-Y.; Kang, K.S.; Lee, S.; Kim, K.H. Procyanidin B2 3″-O-gallate Isolated from Reynoutria elliptica Prevents Glutamate-Induced HT22 Cell Death by Blocking the Accumulation of Intracellular Reactive Oxygen Species. Biomolecules 2019, 9, 412. https://doi.org/10.3390/biom9090412
Song JH, Kim S, Yu JS, Park DH, Kim S-Y, Kang KS, Lee S, Kim KH. Procyanidin B2 3″-O-gallate Isolated from Reynoutria elliptica Prevents Glutamate-Induced HT22 Cell Death by Blocking the Accumulation of Intracellular Reactive Oxygen Species. Biomolecules. 2019; 9(9):412. https://doi.org/10.3390/biom9090412
Chicago/Turabian StyleSong, Ji Hoon, Sil Kim, Jae Sik Yu, Do Hwi Park, Song-Yi Kim, Ki Sung Kang, Sullim Lee, and Ki Hyun Kim. 2019. "Procyanidin B2 3″-O-gallate Isolated from Reynoutria elliptica Prevents Glutamate-Induced HT22 Cell Death by Blocking the Accumulation of Intracellular Reactive Oxygen Species" Biomolecules 9, no. 9: 412. https://doi.org/10.3390/biom9090412