Long Exposure to a Diet Supplemented with Antioxidant and Anti-Inflammatory Probiotics Improves Sperm Quality and Progeny Survival in the Zebrafish Model


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Model Organisms
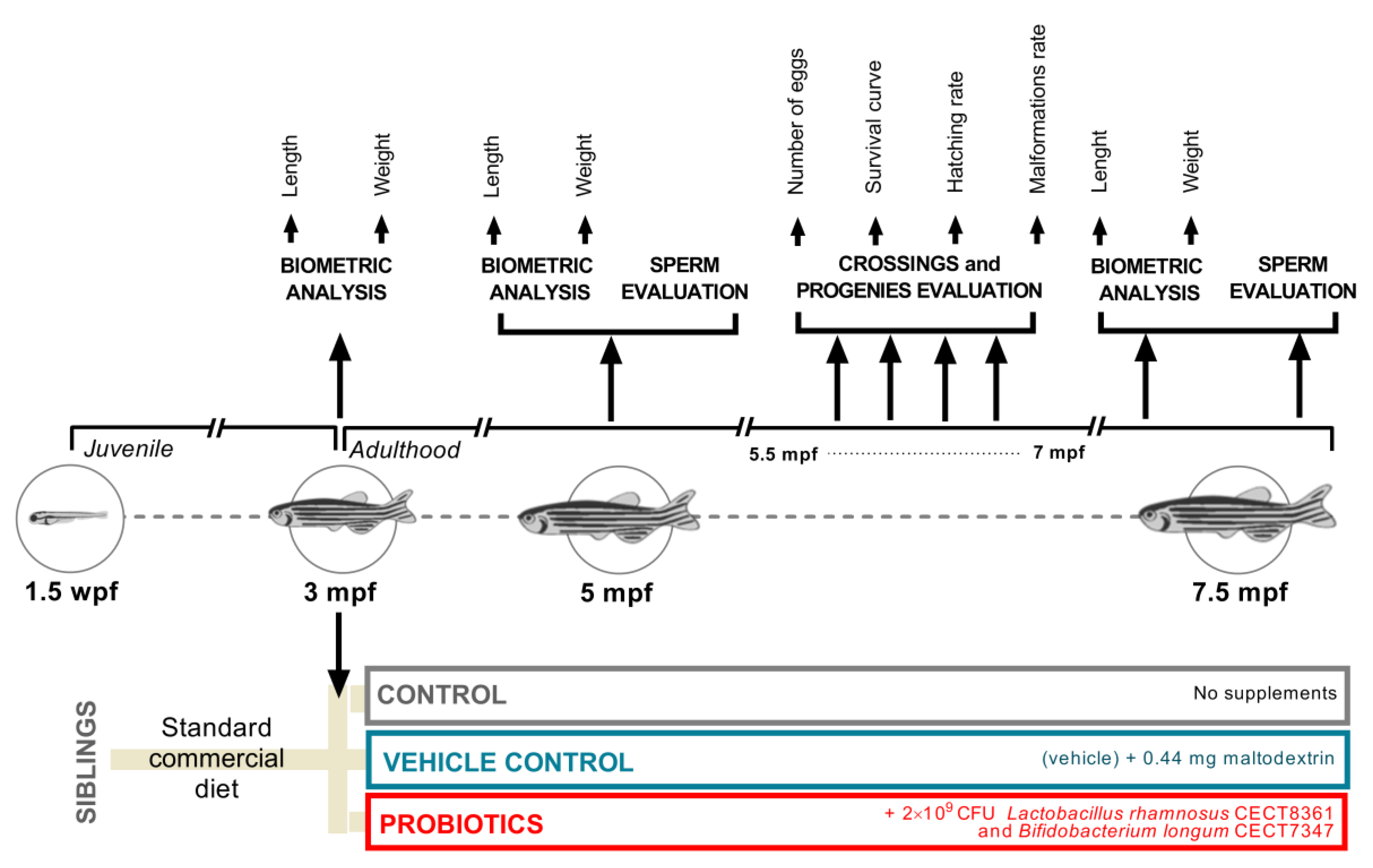
2.3. Study Design, Experimental Diets, and Feeding Regimes
2.4. Biometric Analysis
2.5. Collection of Germplasm
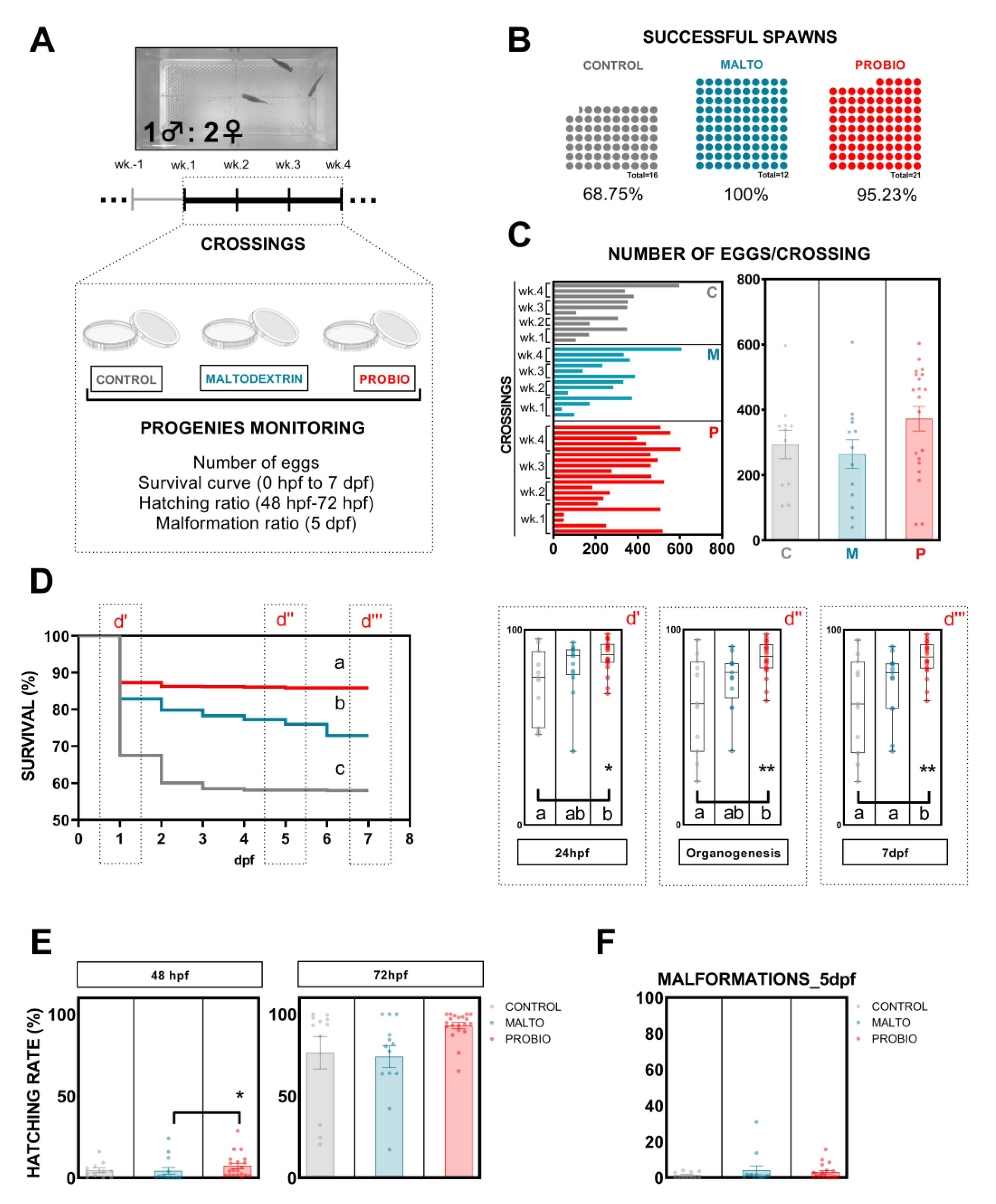
2.6. Progenies
2.7. Progeny Evaluation
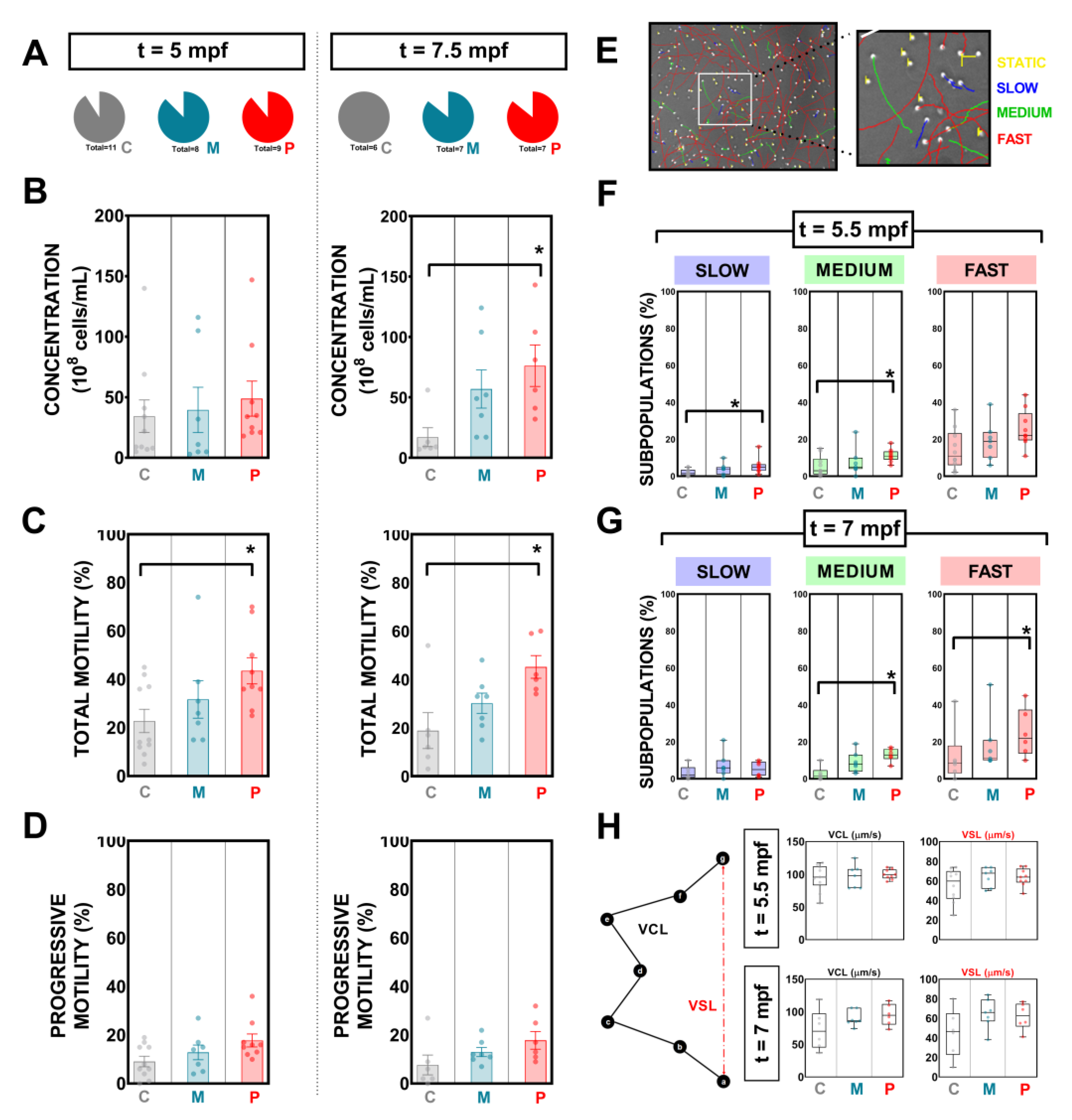
2.8. Sperm Analysis
2.9. Statistical Analysis
3. Results
3.1. Effects of Probiotic Ingestion on Biometric Parameters
3.2. Effects of Probiotic Ingestion on Progenies
3.3. Effects of Probiotic Ingestion on Concentration, Total Sperm Motility, and Progressive Motility
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Broughton, R.E.; Milam, J.E.; Roe, B.A. The Complete Sequence of the Zebrafish (Danio rerio) Mitochondrial Genome and Evolutionary Patterns in Vertebrate Mitochondrial DNA. Genome Res. 2001, 11, 1958–1967. [Google Scholar] [CrossRef] [PubMed]
- Golling, G.; Amsterdam, A.; Sun, Z.; Antonelli, M.; Maldonado, E.; Chen, W.; Burgess, S.; Haldi, M.; Artzt, K.; Farrington, S.; et al. Insertional mutagenesis in zebrafish rapidly identifies genes essential for early vertebrate development. Nat. Genet. 2002, 31, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Grunwald, D.J.; Eisen, J.S. Headwaters of the zebrafish -- emergence of a new model vertebrate. Nat. Rev. Genet. 2002, 3, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Stewart, A.M.; Braubach, O.; Spitsbergen, J.; Gerlai, R.; Kalueff, A.V. Zebrafish models for translational neuroscience research: from tank to bedside. Trends Neurosci. 2014, 37, 264–278. [Google Scholar] [CrossRef] [PubMed]
- MacRae, C.A.; Peterson, R.T. Zebrafish as tools for drug discovery. Nat. Rev. Drug Discov. 2015, 14, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Bambino, K.; Chu, J. Zebrafish in Toxicology and Environmental Health. Curr. Top. Dev. Biol. 2017, 124, 331–367. [Google Scholar]
- Keller, E.T.; Murtha, J.M. The use of mature zebrafish (Danio rerio) as a model for human aging and disease. Comp. Biochem. Physiol. C. Toxicol. Pharmacol. 2004, 138, 335–341. [Google Scholar] [CrossRef]
- Lieschke, G.J.; Currie, P.D. Animal models of human disease: zebrafish swim into view. Nat. Rev. Genet. 2007, 8, 353–367. [Google Scholar] [CrossRef]
- Riesco, M.F.; Valcarce, D.G.; Alfonso, J.; Herráez, M.P.; Robles, V. In vitro generation of zebrafish PGC-like cells. Biol. Reprod. 2014, 91, 114. [Google Scholar] [CrossRef]
- Lee, O.; Takesono, A.; Tada, M.; Tyler, C.R.; Kudoh, T. Biosensor zebrafish provide new insights into potential health effects of environmental estrogens. Environ. Health Perspect. 2012, 120, 990–996. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Lau, S.-W.; Zhang, L.; Ge, W. Disruption of Zebrafish Follicle-Stimulating Hormone Receptor (fshr) But Not Luteinizing Hormone Receptor (lhcgr) Gene by TALEN Leads to Failed Follicle Activation in Females Followed by Sexual Reversal to Males. Endocrinology 2015, 156, 3747–3762. [Google Scholar] [CrossRef] [PubMed]
- Darrow, K.O.; Harris, W.A. Characterization and development of courtship in zebrafish, Danio rerio. Zebrafish 2004, 1, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Laan, M.; Richmond, H.; He, C.; Campbell, R.K. Zebrafish as a Model for Vertebrate Reproduction: Characterization of the First Functional Zebrafish (Danio rerio) Gonadotropin Receptor. Gen. Comp. Endocrinol. 2002, 125, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Blanton, M.L.; Specker, J.L. The hypothalamic-pituitary-thyroid (HPT) axis in fish and its role in fish development and reproduction. Crit. Rev. Toxicol. 2007, 37, 97–115. [Google Scholar] [CrossRef] [PubMed]
- Hoo, J.Y.; Kumari, Y.; Shaikh, M.F.; Hue, S.M.; Goh, B.H. Zebrafish: A Versatile Animal Model for Fertility Research. Biomed Res. Int. 2016, 2016, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Schagdarsurengin, U.; Steger, K. Epigenetics in male reproduction: effect of paternal diet on sperm quality and offspring health. Nat. Rev. Urol. 2016, 13, 584–595. [Google Scholar] [CrossRef]
- Salas-Huetos, A.; Bulló, M.; Salas-Salvadó, J. Dietary patterns, foods and nutrients in male fertility parameters and fecundability: a systematic review of observational studies. Hum. Reprod. Update 2017, 23, 371–389. [Google Scholar] [CrossRef]
- Gaskins, A.J.; Colaci, D.S.; Mendiola, J.; Swan, S.H.; Chavarro, J.E. Dietary patterns and semen quality in young men. Hum. Reprod. 2012, 27, 2899–2907. [Google Scholar] [CrossRef]
- Nassan, F.L.; Chavarro, J.E.; Tanrikut, C. Diet and men’s fertility: does diet affect sperm quality? Fertil. Steril. 2018, 110, 570–577. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Valcarce, D.G.; Genovés, S.; Riesco, M.F.; Martorell, P.; Herráez, M.P.; Ramón, D.; Robles, V. Probiotic administration improves sperm quality in asthenozoospermic human donors. Benef. Microbes 2017, 8, 193–206. [Google Scholar] [CrossRef] [PubMed]
- Medina, M.; De Palma, G.; Ribes-Koninckx, C.; Calabuig, M.; Sanz, Y. Bifidobacterium strains suppress in vitro the pro-inflammatory milieu triggered by the large intestinal microbiota of coeliac patients. J. Inflamm. (Lond). 2008, 5, 19. [Google Scholar] [CrossRef] [PubMed]
- Westerfield, M. The Zebrafish Book. A Guide for the Laboratory Use of Zebrafish (Danio rerio), 4th ed.; University of Oregon Press: Eugene, OR, USA, 2000. [Google Scholar]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of Embryonic Development of the Zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Valcarce, D.G.; Pardo, M.Á.; Riesco, M.F.; Cruz, Z.; Robles, V. Effect of diet supplementation with a commercial probiotic containing Pediococcus acidilactici (Lindner, 1887) on the expression of five quality markers in zebrafish (Danio rerio (Hamilton, 1822)) testis. J. Appl. Ichthyol. 2015, 31, 18–21. [Google Scholar] [CrossRef]
- Maretti, C.; Cavallini, G. The association of a probiotic with a prebiotic (Flortec, Bracco) to improve the quality/quantity of spermatozoa in infertile patients with idiopathic oligoasthenoteratospermia: a pilot study. Andrology 2017, 5, 439–444. [Google Scholar] [CrossRef] [PubMed]
- Valcarce, D.G.; Riesco, M.F.; Martínez-Vázquez, J.M.; Robles, V. Diet Supplemented with Antioxidant and Anti-Inflammatory Probiotics Improves Sperm Quality after Only One Spermatogenic Cycle in Zebrafish Model. Nutrients 2019, 11, 843. [Google Scholar] [CrossRef]
- Tremellen, K. Gut Endotoxin Leading to a Decline IN Gonadal function (GELDING) - a novel theory for the development of late onset hypogonadism in obese men. Basic Clin. Androl. 2016, 26, 7. [Google Scholar] [CrossRef]
- Tremellen, K.; Pearce, K. Probiotics to improve testicular function (Andrology 5:439-444, 2017) - a comment on mechanism of action and therapeutic potential of probiotics beyond reproduction. Andrology 2017, 5, 1052–1053. [Google Scholar] [CrossRef] [PubMed]
- Whiteside, S.A.; Razvi, H.; Dave, S.; Reid, G.; Burton, J.P. The microbiome of the urinary tract—a role beyond infection. Nat. Rev. Urol. 2015, 12, 81–90. [Google Scholar] [CrossRef]
- Rautava, S.; Luoto, R.; Salminen, S.; Isolauri, E. Microbial contact during pregnancy, intestinal colonization and human disease. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 565–576. [Google Scholar] [CrossRef]
- Gensollen, T.; Iyer, S.S.; Kasper, D.L.; Blumberg, R.S. How colonization by microbiota in early life shapes the immune system. Science (80-. ) 2016, 352, 539–544. [Google Scholar] [CrossRef]
- Selma-Royo, M.; Tarrazó, M.; García-Mantrana, I.; Gómez-Gallego, C.; Salminen, S.; Collado, M.C. Shaping Microbiota During the First 1000 Days of Life. Adv. Exp. Med. Biol. 2019, 1125, 3–24. [Google Scholar]
- De Palma, G.; Kamanova, J.; Cinova, J.; Olivares, M.; Drasarova, H.; Tuckova, L.; Sanz, Y. Modulation of phenotypic and functional maturation of dendritic cells by intestinal bacteria and gliadin: relevance for celiac disease. J. Leukoc. Biol. 2012, 92, 1043–1054. [Google Scholar] [CrossRef]
- Fijan, S. Microorganisms with claimed probiotic properties: an overview of recent literature. Int. J. Environ. Res. Public Health 2014, 11, 4745–4767. [Google Scholar] [CrossRef]
- Sinclair, K.D.; Watkins, A.J. Parental diet, pregnancy outcomes and offspring health: metabolic determinants in developing oocytes and embryos. Reprod. Fertil. Dev. 2014, 26, 99. [Google Scholar] [CrossRef]
- Carnevali, O.; Avella, M.A.; Gioacchini, G. Effects of probiotic administration on zebrafish development and reproduction. Gen. Comp. Endocrinol. 2013, 188, 297–302. [Google Scholar] [CrossRef]
- Gioacchini, G.; Maradonna, F.; Lombardo, F.; Bizzaro, D.; Olivotto, I.; Carnevali, O. Increase of fecundity by probiotic administration in zebrafish (Danio rerio). Reproduction 2010, 140, 953–959. [Google Scholar] [CrossRef]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen; World Health Organization: Geneva, Switzerland, 2010; ISBN 9789241547789.
- Wang, C.; Swerdloff, R.S. Limitations of semen analysis as a test of male fertility and anticipated needs from newer tests. Fertil. Steril. 2014, 102, 1502–1507. [Google Scholar] [CrossRef]
- Kowalski, R.K.; Cejko, B.I. Sperm quality in fish: Determinants and affecting factors. Theriogenology 2019, 135, 94–108. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valcarce, D.G.; Riesco, M.F.; Martínez-Vázquez, J.M.; Robles, V. Long Exposure to a Diet Supplemented with Antioxidant and Anti-Inflammatory Probiotics Improves Sperm Quality and Progeny Survival in the Zebrafish Model. Biomolecules 2019, 9, 338. https://doi.org/10.3390/biom9080338
Valcarce DG, Riesco MF, Martínez-Vázquez JM, Robles V. Long Exposure to a Diet Supplemented with Antioxidant and Anti-Inflammatory Probiotics Improves Sperm Quality and Progeny Survival in the Zebrafish Model. Biomolecules. 2019; 9(8):338. https://doi.org/10.3390/biom9080338
Chicago/Turabian StyleValcarce, David G., Marta F. Riesco, Juan M. Martínez-Vázquez, and Vanesa Robles. 2019. "Long Exposure to a Diet Supplemented with Antioxidant and Anti-Inflammatory Probiotics Improves Sperm Quality and Progeny Survival in the Zebrafish Model" Biomolecules 9, no. 8: 338. https://doi.org/10.3390/biom9080338
APA StyleValcarce, D. G., Riesco, M. F., Martínez-Vázquez, J. M., & Robles, V. (2019). Long Exposure to a Diet Supplemented with Antioxidant and Anti-Inflammatory Probiotics Improves Sperm Quality and Progeny Survival in the Zebrafish Model. Biomolecules, 9(8), 338. https://doi.org/10.3390/biom9080338