Detoxification, Apoptosis, and Immune Transcriptomic Responses of the Gill Tissue of Bay Scallop Following Exposure to the Algicide Thiazolidinedione 49


 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Maintenance and TD49 Exposure
2.2. RNA Preparation
2.3. Library Preparation and Illumina Sequencing
2.4. De Novo Transcriptome Assembly
2.5. Gene Annotation and Analysis
2.6. Enrichment Analysis of DEGs
2.7. Quantitative Real-Time PCR Verification
2.8. Statistical Analysis
3. Results
3.1. Analysis of DGE Libraries
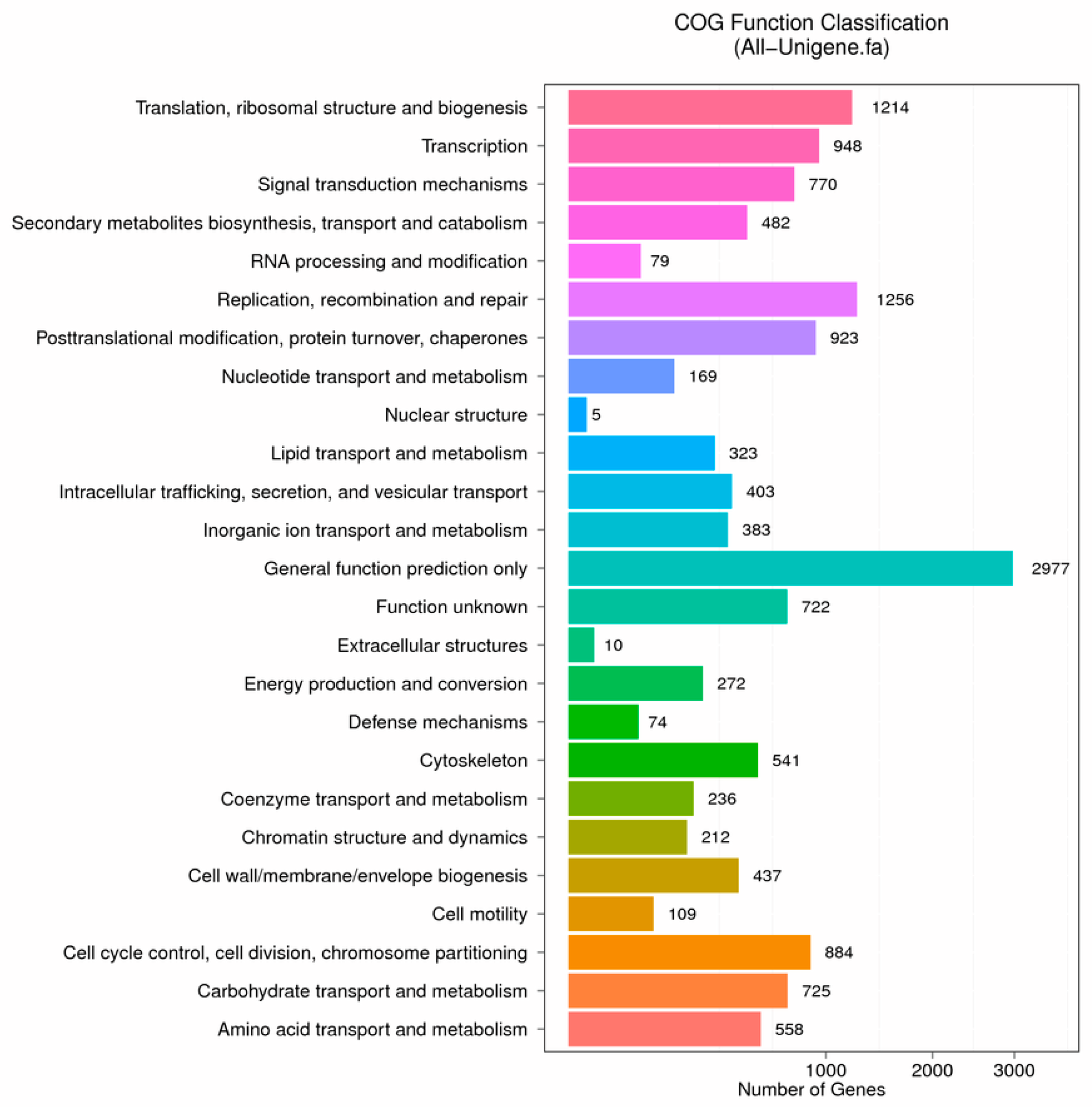
3.2. Functional Annotation and Species Distribution
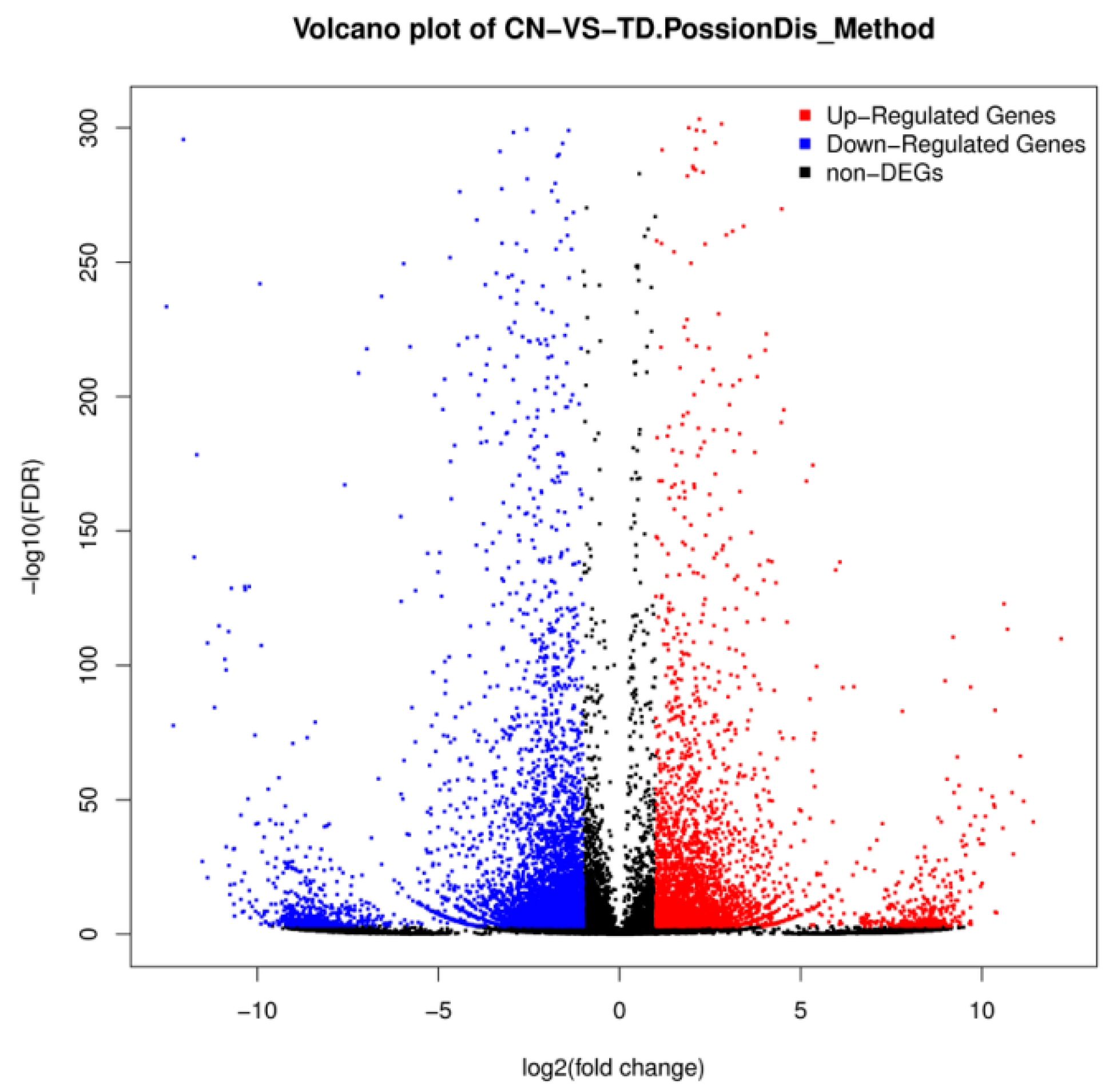
3.3. DEG Analysis
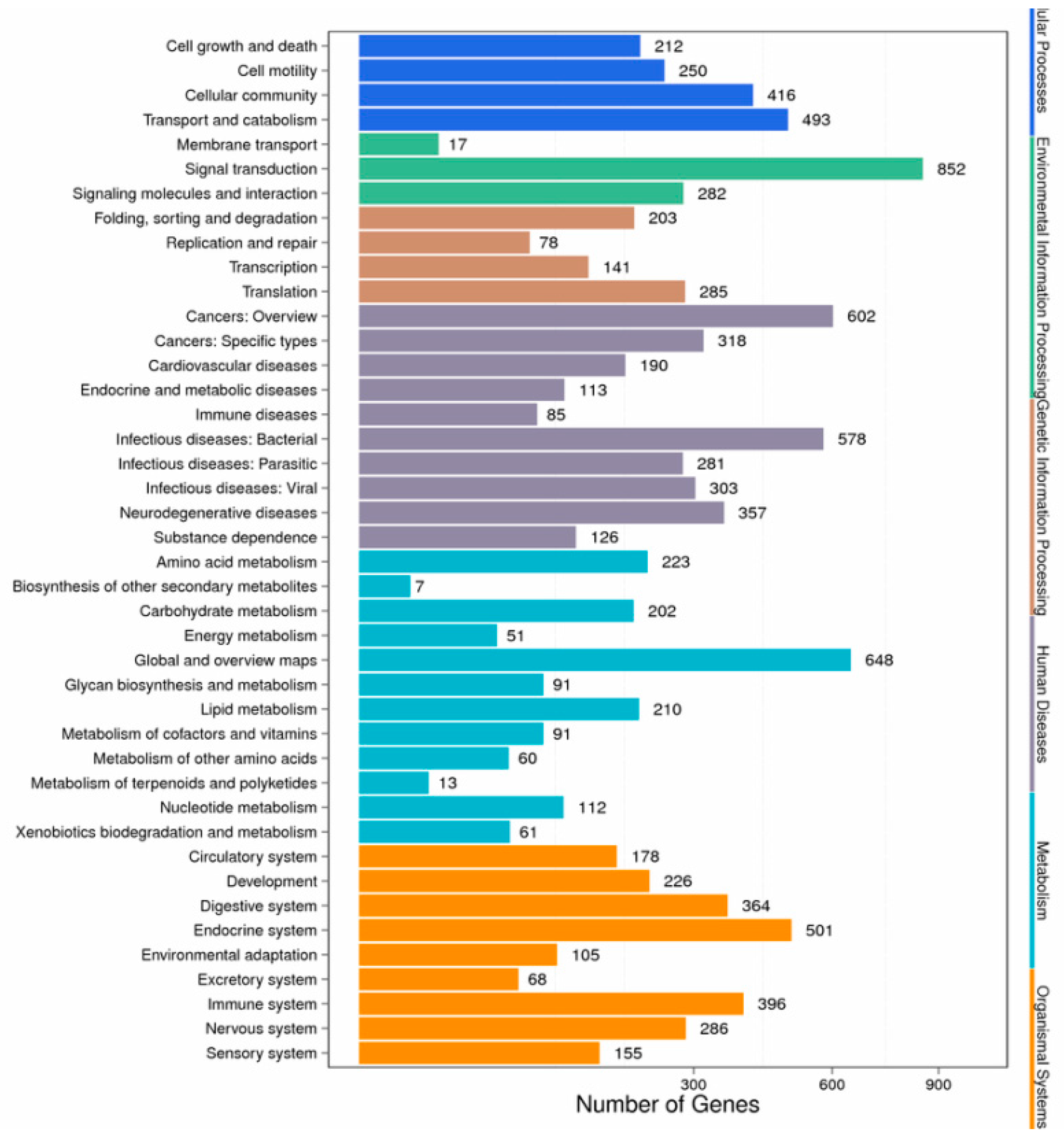
3.4. Enrichment and Pathway Analysis
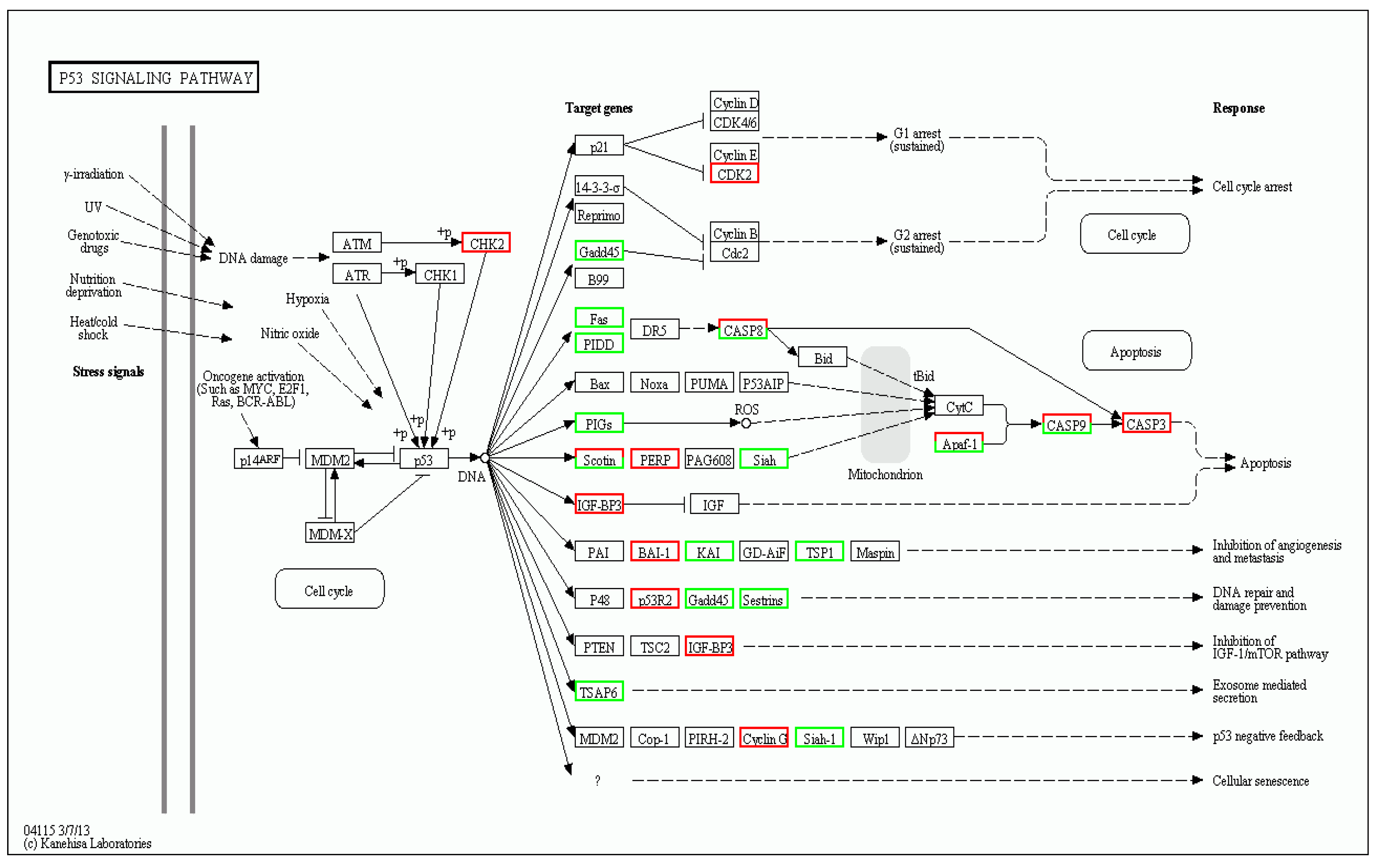
3.5. Genes Related to TD49-Induced Stress Response
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cai, Y.; Pan, L.; Hu, F.; Jin, Q.; Liu, T. Deep sequencing-based transcriptome profiling analysis of Chlamys farreri exposed to benzo pyrene. Gene 2014, 551, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Miao, J.; Wang, J.; Liu, J. AHH activity, tissue dose and DNA damage in different tissues of the scallop Chlamys farreri exposed to benzo pyrene. Environ. Pollut. 2008, 153, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Pan, L.Q.; Zhang, L.; Miao, J.; Wang, J. Immune responses, ROS generation and the haemocyte damage of scallop Chlamys farreri exposed to Aroclor 1254. Fish Shellfish Immunol. 2009, 26, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, E.D.; Bowen, V.T.; Farrington, J.W.; Harvey, G.; Martin, J.H.; Parker, P.L.; Risebrough, R.W.; Robertson, W.; Schneider, E.; Gamble, E. The mussel watch. Environ. Conserv. 1978, 5, 101–125. [Google Scholar] [CrossRef]
- Liu, N.; Pan, L.; Wang, J.; Yang, H.; Liu, D. Application of the biomarker responses in scallop (Chlamys farreri) to assess metals and PAHs pollution in Jiaozhou Bay, China. Mar. Environ. Res. 2012, 80, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Yim, E.C.; Park, I.T.; Kim, S.W.; Cho, H. Comparison of the acute toxicities of novel algicides, thiazolidinedione derivatives TD49 and TD53, to various marine organisms. Environ. Toxicol. Chem. 2011, 30, 2810–2816. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.M.; Wu, Y.; Duong, T.U.; Ghodake, G.S.; Kim, S.W.; Jin, E.; Cho, H. Thiazolidinediones as a novel class of algicides against red tide harmful algal species. Appl. Biochem. Biotechnol. 2010, 162, 2273–2283. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.H.; Son, M.; Kim, Y.O.; Cho, H.; Chung, I.K.; Park, B.S. Can algicide (the thiazolidinedione derivative TD49) truly contribute to the restoration of microbial communities? Environ. Res. 2019, 173, 517–527. [Google Scholar] [CrossRef]
- Baek, S.H.; Son, M.; Kim, Y.O.; Cho, H.; Lee, M.; Na, D.H.; Kim, S.W. Response of Chattonella marina (Raphidophyceae) and marine plankton to yellow clay and thiazolidinedione derivative TD49 in a mesocosm enclosure. J. Appl. Phycol. 2017, 29, 285–296. [Google Scholar] [CrossRef]
- Tian, S.; Pan, L.; Sun, X. An investigation of endocrine disrupting effects and toxic mechanisms modulated by benzo [a] pyrene in female scallop Chlamys farreri. Aquat. Toxicol. 2013, 144, 162–171. [Google Scholar] [CrossRef]
- Paules, R.S.; Aubrecht, J.; Corvi, R.; Garthoff, B.; Kleinjans, J.C. Moving forward in human cancer risk assessment. Environ. Health Perspect. 2010, 119, 739–743. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chi, C.; Giri, S.S.; Jun, J.W.; Kim, H.J.; Kim, S.W.; Kang, J.W.; Park, S.C. Detoxification and immune transcriptomic response of the gill tissue of bay scallop (Argopecten irradians) following exposure to the algicide palmitoleic acid. Biomolecules 2018, 8, 139. [Google Scholar] [CrossRef] [PubMed]
- Chi, C.; Giri, S.; Jun, J.; Kim, S.; Kim, H.; Kang, J.; Park, S. Detoxification-and immune-related transcriptomic analysis of gills from bay scallops (Argopecten irradians) in response to algal toxin okadaic acid. Toxins 2018, 10, 308. [Google Scholar] [CrossRef] [PubMed]
- Reuter, J.A.; Spacek, D.V.; Snyder, M.P. High-throughput sequencing technologies. Mol. Cell 2015, 58, 586–597. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Sun, Y.; Wang, J.; Xing, Q.; Zou, J.; Li, R.; Wang, Z.; Wang, S.; Hu, X.; Zhang, L. Sequencing-based gene network analysis provides a core set of gene resource for understanding thermal adaptation in Zhikong scallop Chlamys farreri. Mol. Ecol. Resour. 2014, 14, 184–198. [Google Scholar] [CrossRef] [PubMed]
- Philipp, E.E.; Kraemer, L.; Melzner, F.; Poustka, A.J.; Thieme, S.; Findeisen, U.; Schreiber, S.; Rosenstiel, P. Massively parallel RNA sequencing identifies a complex immune gene repertoire in the lophotrochozoan Mytilus Edulis. PLoS ONE 2012, 7, e33091. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]
- Audic, S.; Claverie, J.M. The significance of digital gene expression profiles. Genome Res. 1997, 7, 986–995. [Google Scholar] [CrossRef] [PubMed]
- Chi, C.; Giri, S.S.; Jun, J.W.; Kim, H.J.; Kim, S.W.; Yun, S.; Park, S.C. Effects of algal toxin okadaic acid on the non-specific immune and antioxidant response of bay scallop (Argopecten irradians). Fish Shellfish Immunol. 2017, 65, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.C.; Bratton, S.B. Regulation of the intrinsic apoptosis pathway by reactive oxygen species. Antioxid. Redox Signal. 2013, 19, 546–558. [Google Scholar] [CrossRef] [PubMed]
- Wong, W.W.L.; Puthalakath, H. Bcl-2 family proteins: The sentinels of the mitochondrial apoptosis pathway. Iubmb Life 2008, 60, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Budihardjo, I.; Oliver, H.; Lutter, M.; Luo, X.; Wang, X. Biochemical pathways of caspase activation during apoptosis. Annu. Rev. Cell Dev. Biol. 1999, 15, 269–290. [Google Scholar] [CrossRef]
- Deveraux, Q.L.; Reed, J.C. IAP family proteins—suppressors of apoptosis. Genes Dev. 1999, 13, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Wu, C.H.; Gordon, J.D.; Zhong, X.; Emami, A.; Safa, A.R. Taxol induces caspase-10-dependent apoptosis. J. Biol. Chem. 2004, 279, 51057–51067. [Google Scholar] [CrossRef]
- Huang, L.; Zou, Y.; Weng, H.W.; Li, H.Y.; Liu, J.S.; Yang, W.D. Proteomic profile in Perna viridis after exposed to Prorocentrum lima, a dinoflagellate producing DSP toxins. Environ. Pollut. 2015, 196, 350–357. [Google Scholar] [CrossRef]
- Dean, M.; Hamon, Y.; Chimini, G. The human ATP-binding cassette (ABC) transporter superfamily. J. Lipid Res. 2001, 42, 1007–1017. [Google Scholar] [CrossRef]
- Schinkel, A.H.; Jonker, J.W. Mammalian drug efflux transporters of the ATP binding cassette (ABC) family: An overview. Adv. Drug Deliv. Rev. 2012, 64, 138–153. [Google Scholar] [CrossRef]
- Nebert, D.W.; Russell, D.W. Clinical importance of the cytochromes P450. Lancet 2002, 360, 1155–1162. [Google Scholar] [CrossRef]
- Lambeth, J.D. NOX enzymes and the biology of reactive oxygen. Nat. Rev. Immunol. 2004, 4, 181. [Google Scholar] [CrossRef] [PubMed]
- Guengerich, F.P. Cytochrome p450 and chemical toxicology. Chem. Res. Toxicol. 2007, 21, 70–83. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.D.; Pulford, D.J. The glut athione S-transferase supergene family: Regulation of GST and the contribution of the lsoenzymes to cancer chemoprotection and drug resistance part I. Crit. Rev. Biochem. Mol. Biol. 1995, 30, 445–520. [Google Scholar] [CrossRef] [PubMed]
- Fattman, C.L.; Schaefer, L.M.; Oury, T.D. Extracellular superoxide dismutase in biology and medicine. Free Radic. Biol. Med. 2003, 35, 236–256. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Bauché, F.; Fouchard, M.H.; Jégou, B. Antioxidant system in rat testicular cells. Febs Lett. 1994, 349, 392–396. [Google Scholar] [CrossRef]
- Dinkova-Kostova, A.T.; Talalay, P. NAD (P) H: Quinone acceptor oxidoreductase 1 (NQO1), a multifunctional antioxidant enzyme and exceptionally versatile cytoprotector. Arch. Biochem. Biophys. 2010, 501, 116–123. [Google Scholar] [CrossRef]
- Kaspar, J.W.; Niture, S.K.; Jaiswal, A.K. Nrf2: INrf2 (Keap1) signaling in oxidative stress. Free Radic. Biol. Med. 2009, 47, 1304–1309. [Google Scholar] [CrossRef]
- Li, H.; Yang, L. Molecular regulatory mechanism of Nrf2 antioxidant. Chin. J. Bioinform. 2018, 16, 1–6. [Google Scholar]
- Chen, M.S.; Yang, H.S.; Delaporte, M.; Zhao, S.J.; Xing, K. Immune responses of the scallop Chlamys farreri after air exposure to different temperatures. J. Exp. Mar. Biol. Ecol. 2007, 345, 52–60. [Google Scholar] [CrossRef]
- Suresh, K.; Mohandas, A. Hemolymph acid phosphatase activity pattern in copper-stressed bivalves. J. Invertebr. Pathol. 1990, 55, 118–125. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Total Raw Reads (Mb) | Total Clean Reads (Mb) | Total Clean Bases (Gb) | Clean Reads Q20 (%) | Clean Reads Q30 (%) | Clean Reads Ratio (%) |
|---|---|---|---|---|---|---|
| CN | 45.92 | 45.76 | 4.58 | 98.21 | 95.30 | 99.65 |
| TD | 45.92 | 45.85 | 4.58 | 98.30 | 95.52 | 99.84 |
| Sample | Total Number | Total Length | Mean Length | N50 | N70 | N90 | GC (%) |
|---|---|---|---|---|---|---|---|
| CN | 78,376 | 52,817,747 | 673 | 1231 | 568 | 252 | 39.12 |
| TD | 71,899 | 45,492,111 | 632 | 1093 | 518 | 245 | 39.23 |
| Sample | Total Number | Total Length | Mean Length (bp) | N50 | N70 | N90 | GC (%) |
|---|---|---|---|---|---|---|---|
| CN | 51,394 | 40,985,988 | 797 | 1410 | 702 | 302 | 39.48 |
| TD | 48,654 | 36,217,952 | 744 | 1266 | 637 | 288 | 39.54 |
| All-unigene | 57,882 | 50,329,144 | 869 | 1594 | 827 | 321 | 39.36 |
| Values | Total | Nr- Annotated | Nt- Annotated | SwissProt- Annotated | KEGG- Annotated | COG- Annotated | Interpro- Annotated | GO- Annotated | Overall |
|---|---|---|---|---|---|---|---|---|---|
| Number | 57,882 | 25,267 | 9983 | 19,555 | 18,931 | 8729 | 19,017 | 4032 | 28,172 |
| Percentage | 100% | 43.65% | 17.25% | 33.78% | 32.71% | 15.08% | 32.85% | 6.97% | 48.67% |
| Description | Transcript | Log2 Fold Change (RNAseq) | Regulation |
|---|---|---|---|
| Downregulated | |||
| Immune system | CLEC 4M | −1.62 | Down |
| CLEC 4F | −2.69 | Down | |
| FIBCD 1 | −2.00 | Down | |
| ACP 5 | −2.94 | Down | |
| ACP 6 | −2.40 | Down | |
| PAPL | −1.27 | Down | |
| HSP70 | −2.00 | Down | |
| Apoptosis | Bcl-2 | −1.90 | Down |
| BIRC 2 | −2.51 | Down | |
| BIRC 3 | −4.97 | Down | |
| BIRC 6 | −4.25 | Down | |
| BIRC 7A | −1.33 | Down | |
| BIRC 7B | −1.61 | Down | |
| Transmembrane proteins | ABC A3 | −5.01 | Down |
| ABC A5 | −2.80 | Down | |
| ABC B1 | −2.11 | Down | |
| ABC B10 | −4.27 | Down | |
| ABC C1 | −1.16 | Down | |
| ABC D3 | −1.32 | Down | |
| ABC G2 | −1.95 | Down | |
| Antioxidant system | NQO1 | −5.10 | Down |
| CAT | −3.06 | Down | |
| Mn SOD | −1.30 | Down | |
| Cu/Zn SOD | −1.56 | Down | |
| GR | −5.08 | Down | |
| GPx | −2.55 | Down | |
| Nrf2 | −1.61 | Down | |
| Metabolism of xenobiotics | CREBP 3 | −1.39 | Down |
| MOA | −1.79 | Down | |
| NOX3 | −1.55 | Down | |
| GSTω | −11.18 | Down | |
| Upregulated | |||
| Apoptosis | Cas3 | 2.29 | Up |
| Cas6 | 2.79 | Up | |
| FADD | 1.68 | Up | |
| Immune system | TLR 2 | 4.18 | Up |
| TRAF 6 | 1.17 | Up | |
| Metabolism of xenobiotics | CYP 1A1 | 2.45 | Up |
| CYP 2C8 | 8.52 | Up | |
| GST ζ1 | 8.40 | Up | |
| GST 1 | 3.08 | Up | |
| GSTσ3 | 3.20 | Up | |
| GST A | 3.41 | Up | |
| GSTκ1 | 1.74 | Up |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chi, C.; Giri, S.S.; Jun, J.W.; Kim, H.J.; Yun, S.; Kim, S.W.; Kang, J.W.; Park, S.C. Detoxification, Apoptosis, and Immune Transcriptomic Responses of the Gill Tissue of Bay Scallop Following Exposure to the Algicide Thiazolidinedione 49. Biomolecules 2019, 9, 310. https://doi.org/10.3390/biom9080310
Chi C, Giri SS, Jun JW, Kim HJ, Yun S, Kim SW, Kang JW, Park SC. Detoxification, Apoptosis, and Immune Transcriptomic Responses of the Gill Tissue of Bay Scallop Following Exposure to the Algicide Thiazolidinedione 49. Biomolecules. 2019; 9(8):310. https://doi.org/10.3390/biom9080310
Chicago/Turabian StyleChi, Cheng, Sib Sankar Giri, Jin Woo Jun, Hyoun Joong Kim, Saekil Yun, Sang Wha Kim, Jeong Woo Kang, and Se Chang Park. 2019. "Detoxification, Apoptosis, and Immune Transcriptomic Responses of the Gill Tissue of Bay Scallop Following Exposure to the Algicide Thiazolidinedione 49" Biomolecules 9, no. 8: 310. https://doi.org/10.3390/biom9080310
APA StyleChi, C., Giri, S. S., Jun, J. W., Kim, H. J., Yun, S., Kim, S. W., Kang, J. W., & Park, S. C. (2019). Detoxification, Apoptosis, and Immune Transcriptomic Responses of the Gill Tissue of Bay Scallop Following Exposure to the Algicide Thiazolidinedione 49. Biomolecules, 9(8), 310. https://doi.org/10.3390/biom9080310