Proteomic and Phosphoproteomic Analysis in Tobacco Mosaic Virus-Infected Tobacco (Nicotiana tabacum)

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Growing, Viral Inoculation, and Taking of Samples
2.2. Quantitative Reverse-Transcription Polymerase Chain Reaction (RT-PCR) Analysis
2.3. Sample Preparation and Instrument Settings for GC–MS Analysis
2.4. Protein Extraction and Digestion
2.5. iTRAQ Labeling Quantification and Fractionation
2.6. IMAC Enrichment of Phosphopeptides
2.7. Nano-LC–MS/MS Analysis
2.8. Bioinformatics Analysis
3. Results
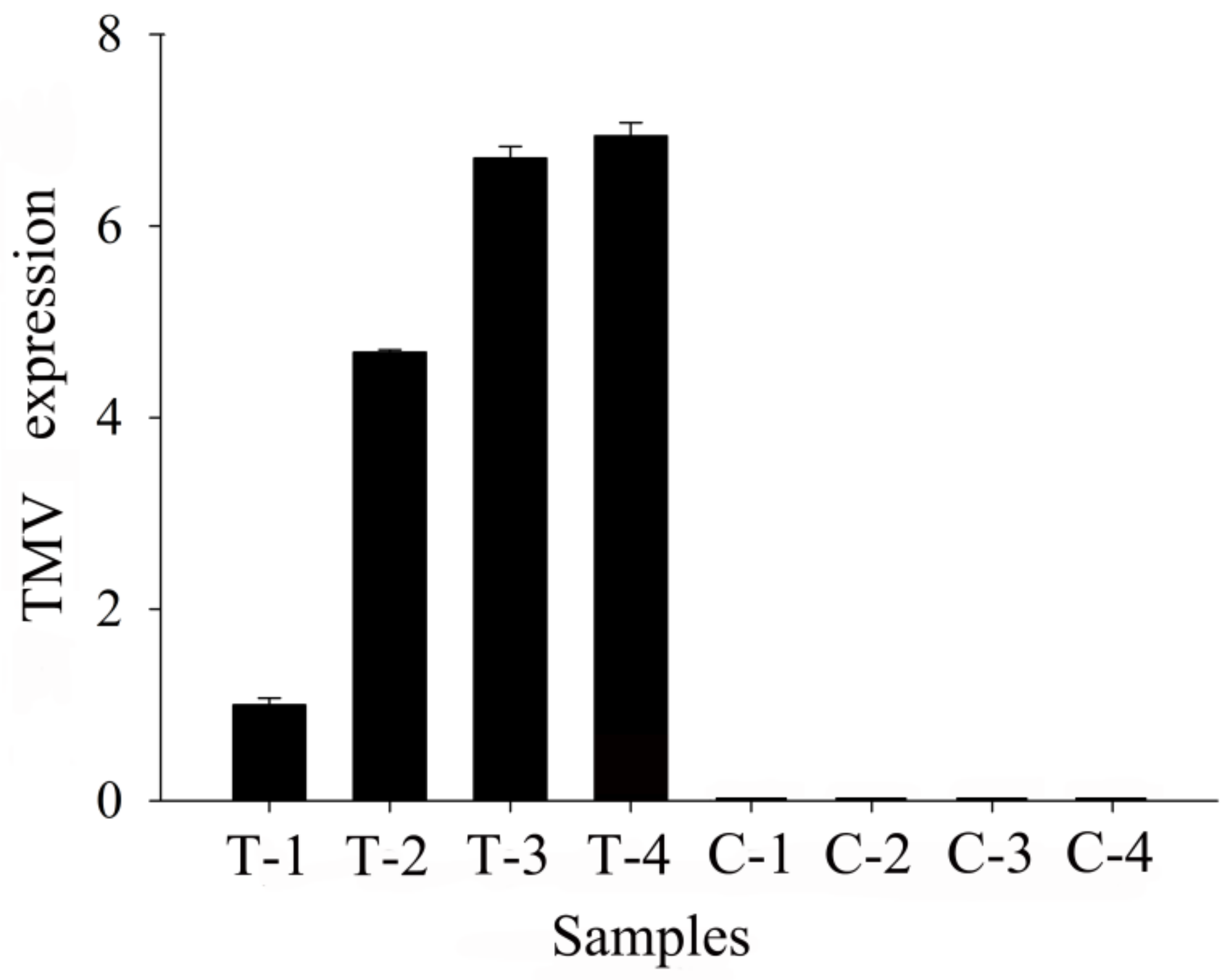
3.1. Real-Time PCR Analysis and Metabolomic Changes in TMV Tobacco
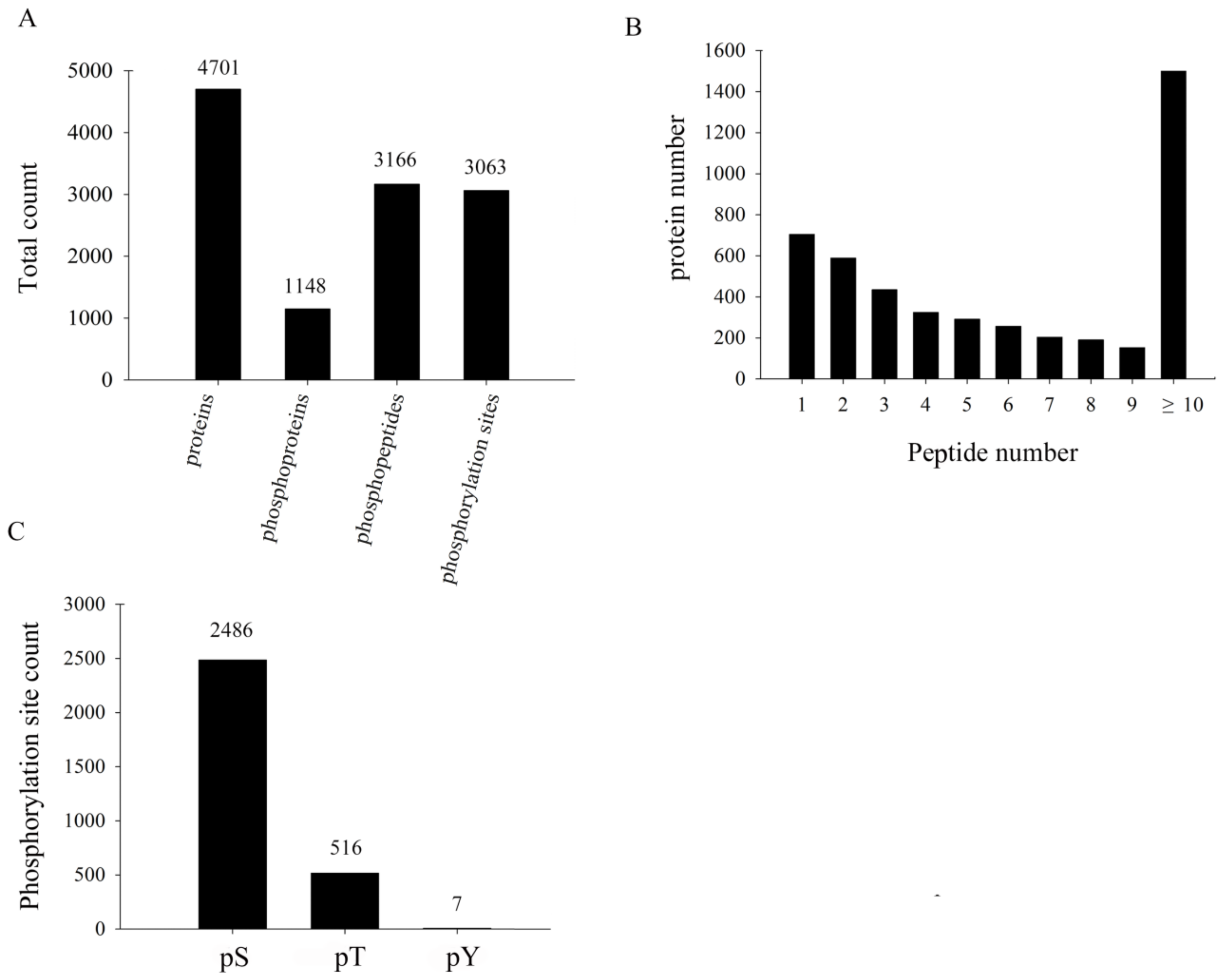
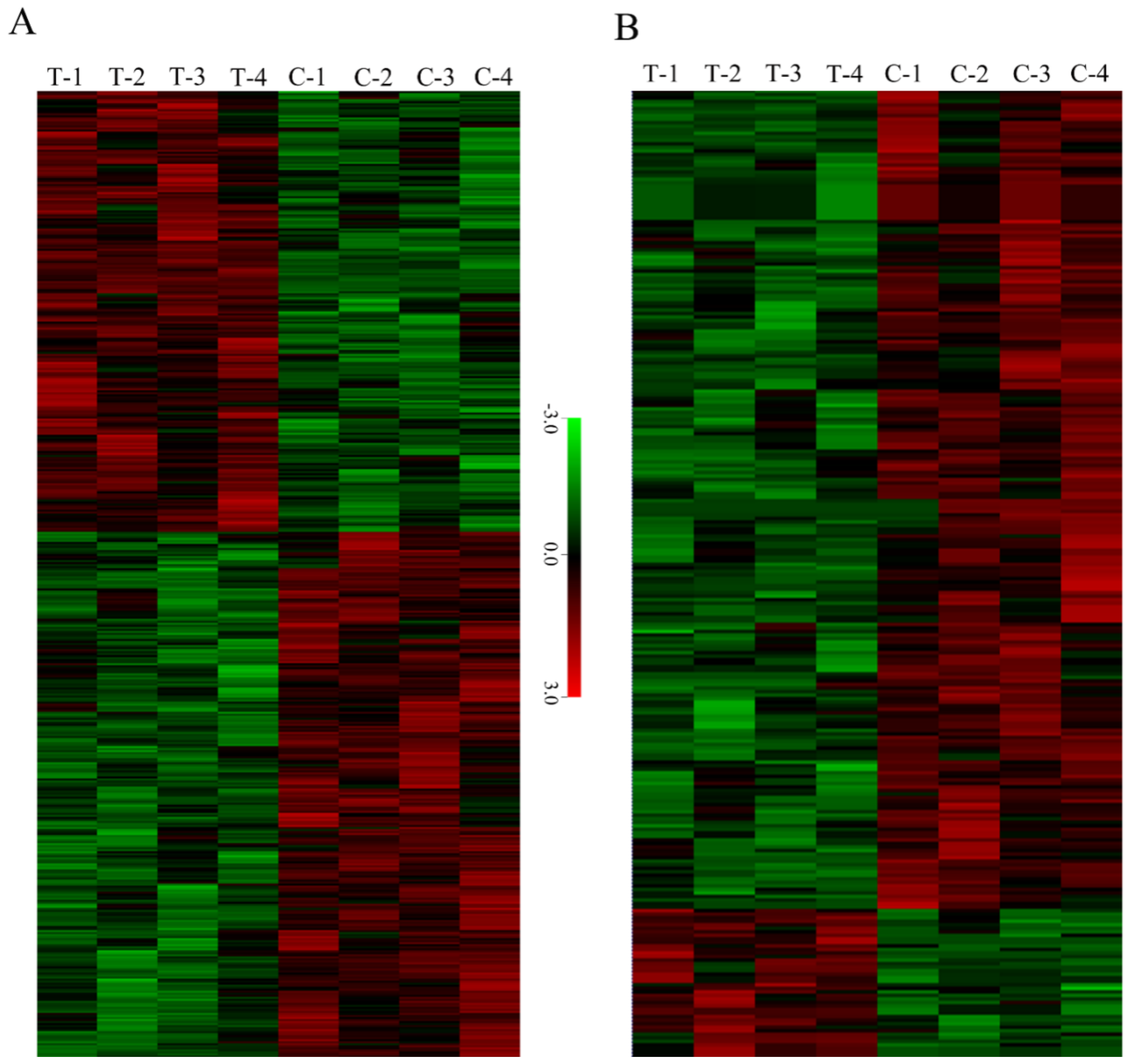
3.2. Primary Proteome and Phosphoproteome Data on TMV-Infected Tobacco
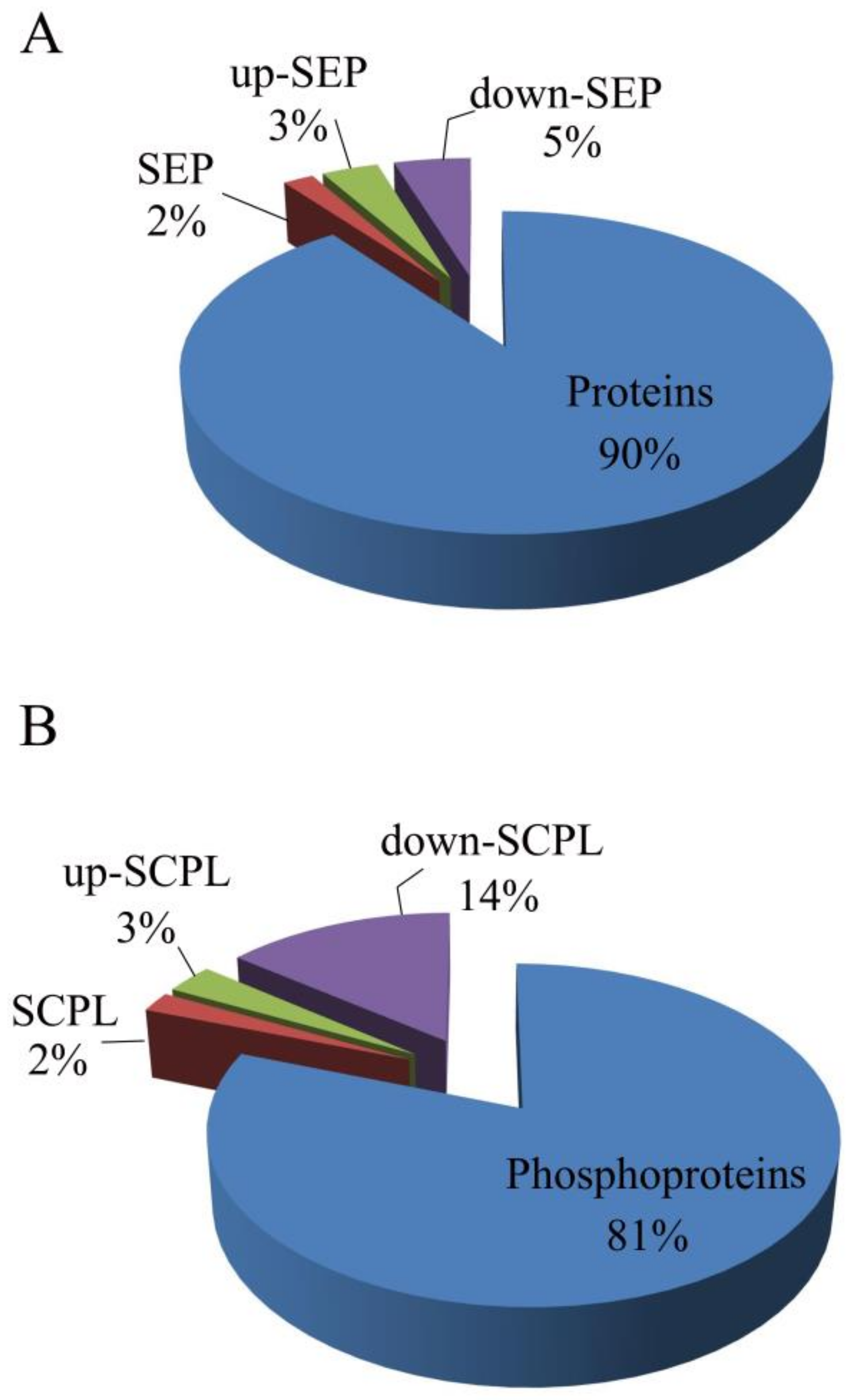
3.3. SEP and SCPL Proteins
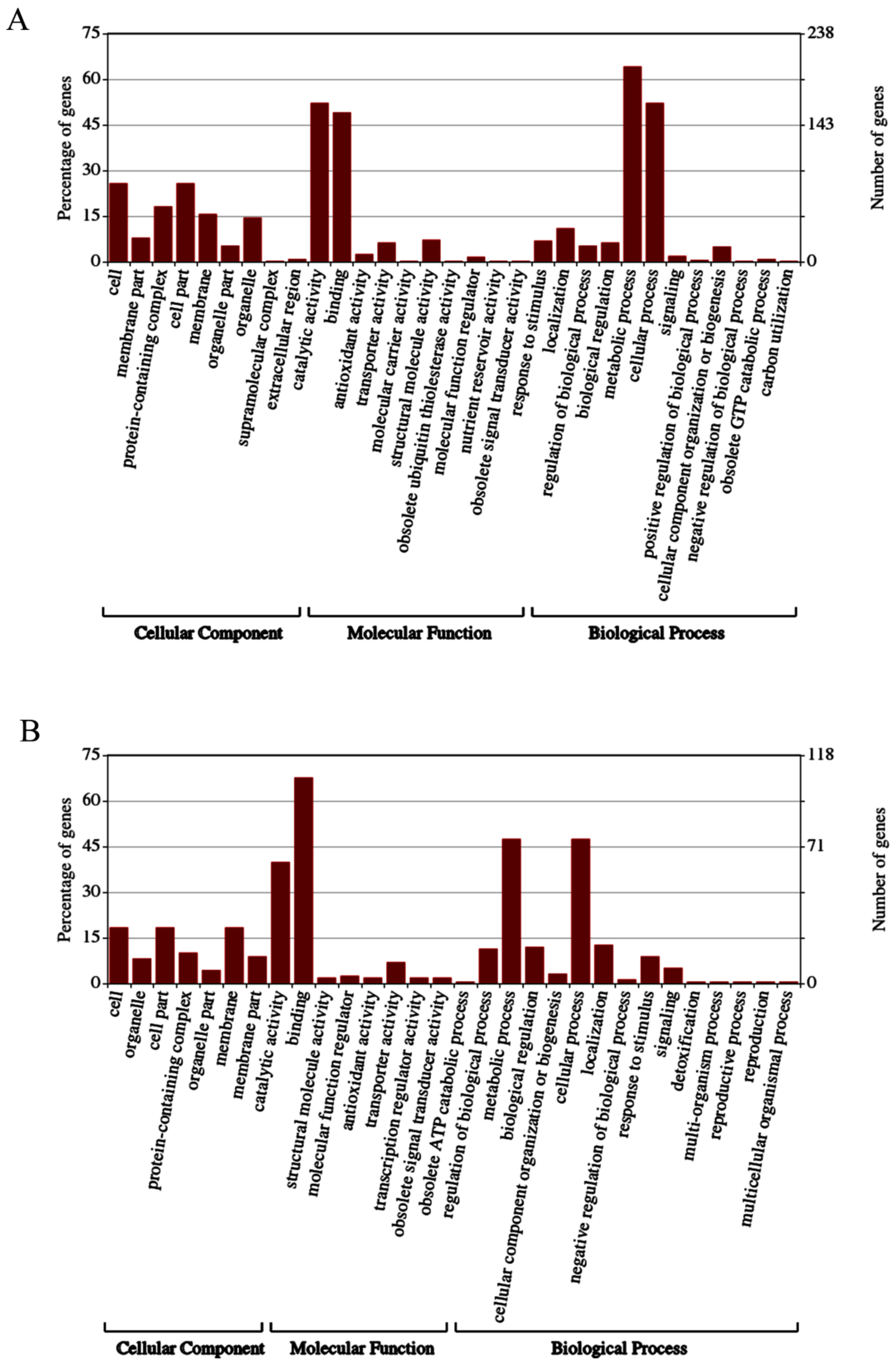
3.4. GO Analysis
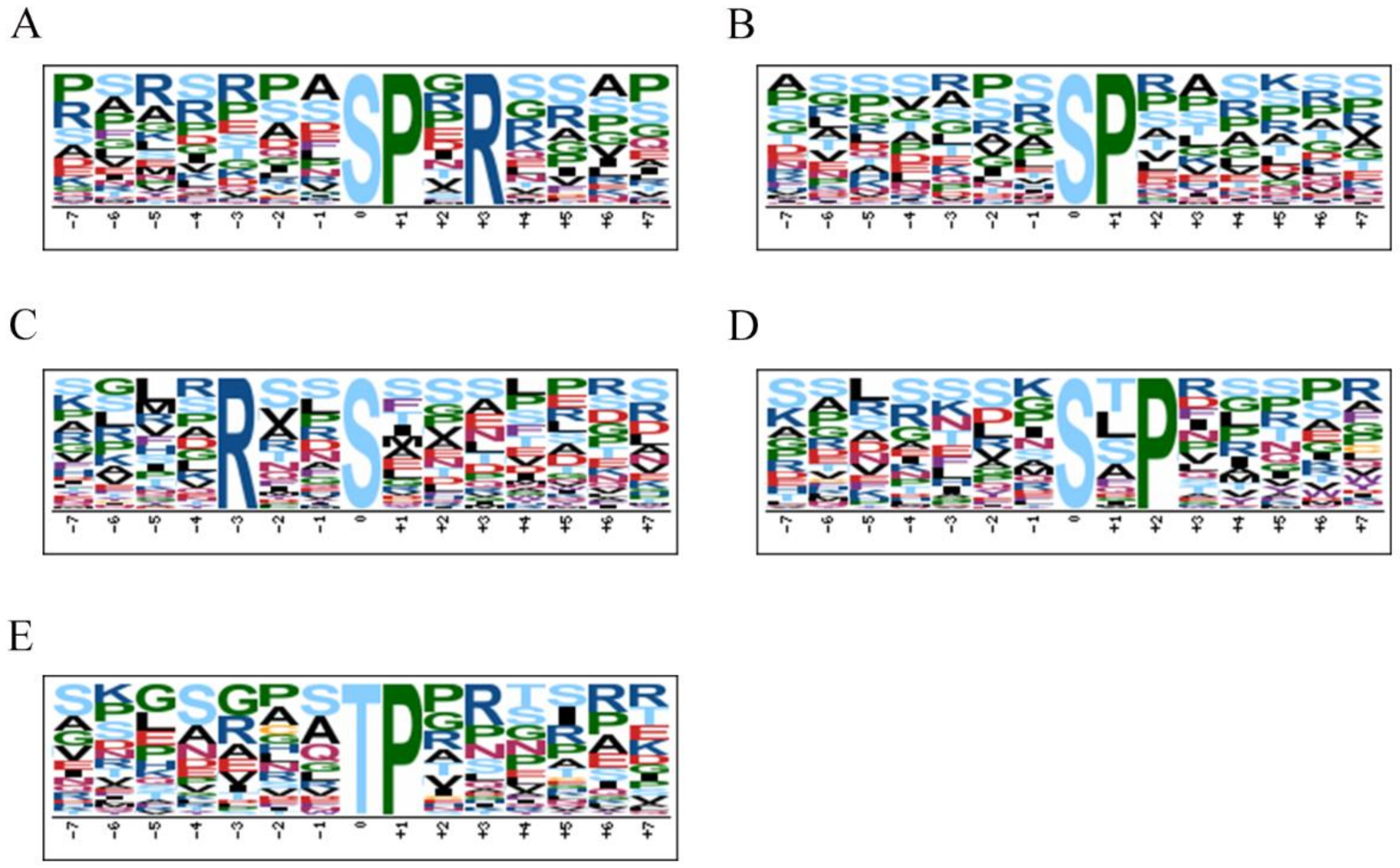
3.5. Motif Analysis
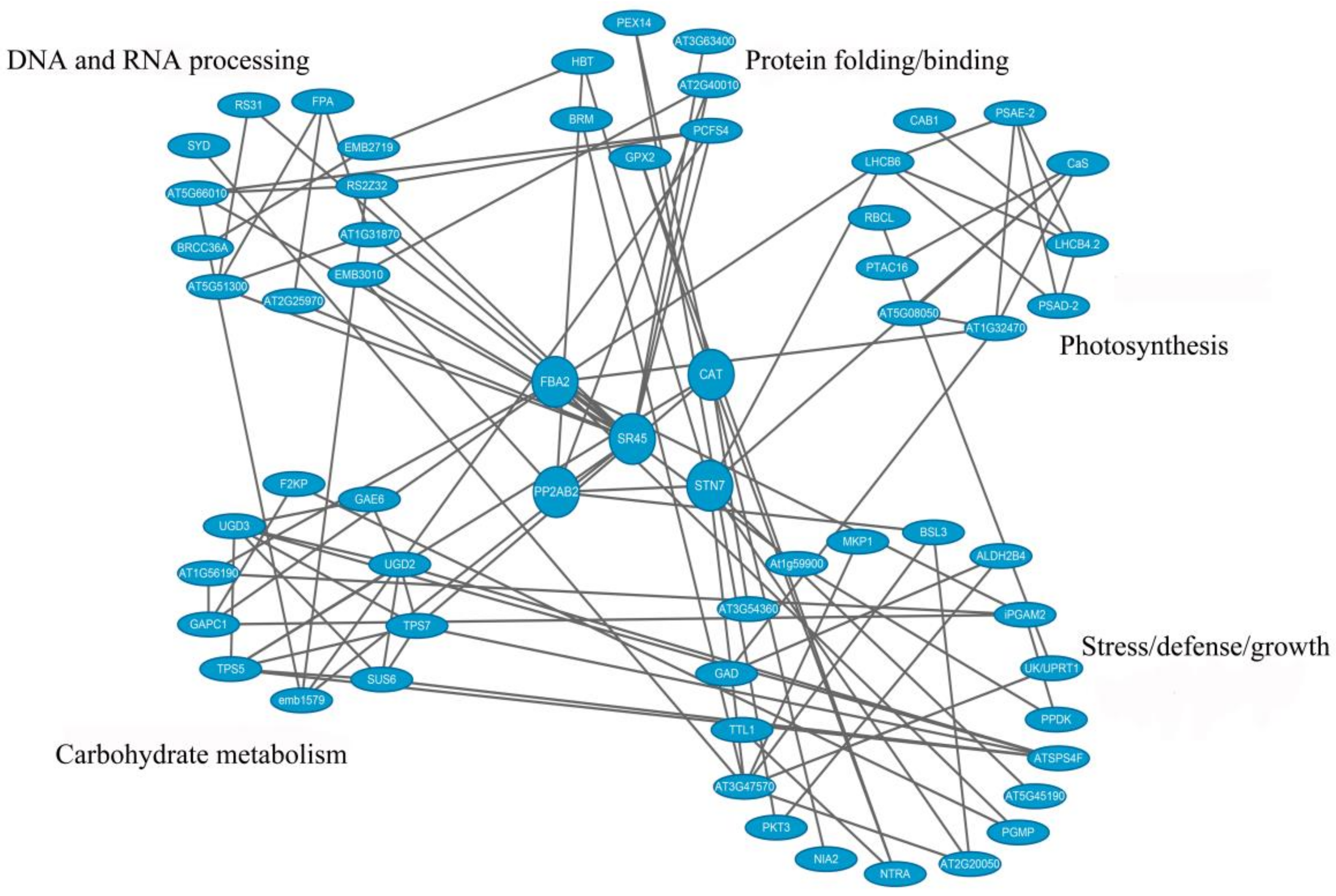
3.6. PPI Analysis of Phosphoproteins
4. Discussion
4.1. Phosphorylated Protein Kinases and Phosphatases Associated with Stress
4.2. Translation Initiation and Transcription Factors
4.3. Tobacco Disease Resistance-Related Phosphoproteins
4.4. Proteins Involved in Disease
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ritzenthaler, C. Resistance to plant viruses: Old issue, news answers? Curr. Opin. Biotechnol. 2005, 16, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wei, P.; Xu, X.; Liu, Y.; Wang, L.; Wang, Q. Design, Synthesis, and Antiviral Activity Evaluation of Phenanthrene-Based Antofine Derivatives. J. Agric. Food Chem. 2012, 60, 8544–8551. [Google Scholar] [CrossRef] [PubMed]
- Wan, Z.; Hu, D.; Li, P.; Xie, D.; Gan, X. Synthesis, Antiviral Bioactivity of Novel 4-Thioquinazoline Derivatives Containing Chalcone Moiety. Molecules 2015, 20, 11861–11874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, X.H.; Ren, Z.L.; Li, D.D.; Ruan, B.F.; Li, Q.S.; Chu, M.J.; Ai, C.-Y.; Liu, D.-H.; Mo, K.; Cao, H.-Q. Discovery of novel double pyrazole Schiff base derivatives as anti-tobacco mosaic virus (TMV) agents. Chin. Chem. Lett. 2017, 28, 377–382. [Google Scholar] [CrossRef]
- Wang, J.; Wang, X.R.; Zhou, Q.; Yang, J.M.; Guo, H.X.; Yang, L.J.; Liu, W.Q. iTRAQ protein profile analysis provides integrated insight into mechanisms of tolerance to TMV in tobacco (Nicotiana tabacum). J. Proteom. 2016, 132, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Bozsó, Z.; Ott, P.G.; Kámán-Tóth, E.; Bognár, G.F.; Pogány, M.; Szatmári, Á. Overlapping Yet Response-Specific Transcriptome Alterations Characterize the Nature of Tobacco–Pseudomonas syringae Interactions. Front. Plant Sci. 2016, 7, 251. [Google Scholar] [CrossRef]
- Dong-Wen, L.; Pei, G.; Ming, Z.; Zhi-Wei, C.; Xiao-Hui, L.; Yue-Ming, Y. Integrative network analysis of the signaling cascades in seedling leaves of bread wheat by large-scale phosphoproteomic profiling. J. Proteome Res. 2014, 13, 2381–2395. [Google Scholar] [CrossRef]
- Cohen, P. The origins of protein phosphorylation. Nat. Cell Biol. 2002, 4, 127–130. [Google Scholar] [CrossRef]
- Asurmendi, S.; Berg, R.H.; Koo, J.C.; Beachy, R.N. Coat protein regulates formation of replication complexes during tobacco mosaic virus infection. Proc. Natl. Acad. Sci. USA 2004, 101, 1415–1420. [Google Scholar] [CrossRef] [Green Version]
- Vu, L.D.; Stes, E.; Van Bel, M.; Nelissen, H.; Maddelein, D.; Inzé, D.; Coppens, F.; Martens, L.; Gevaert, K.; De Smet, I. An up-to-date workflow for plant (phospho)proteomics identifies differential drought-responsive phosphorylation events in maize leaves. J. Proteome Res. 2016, 15, 4304–4317. [Google Scholar] [CrossRef]
- Ren, J.; Li, X.; Mao, J.; Zuo, C.; Zhao, X.; Chen, B. Physiological and quantitative phosphoproteome analyses of drought stress-induced mechanisms in Malus baccata (L.) Borkh. Biochem. Syst. Ecol. 2017, 72, 47–55. [Google Scholar] [CrossRef]
- Zhao, J.; Xian, Z.; Hong, Y.; Liu, Y. Chloroplast in Plant-Virus Interaction. Front. Microbiol. 2016, 7, 1565. [Google Scholar] [CrossRef] [PubMed]
- Whitham, S.A.; Quan, S.; Chang, H.S.; Cooper, B.; Estes, B.; Zhu, T.; Wang, X.; Hou, Y.M. Diverse RNA viruses elicit the expression of common sets of genes in susceptible Arabidopsis thaliana plants. Plant J. 2010, 33, 271–283. [Google Scholar] [CrossRef]
- Chen, X.; Chan, W.; Zhu, F.; Lo, C. Phosphoproteomic analysis of the non-seed vascular plant model Selaginella moellendorffii. Proteome Sci. 2014, 12, 16. [Google Scholar] [CrossRef]
- Guo, M.; Chen, K.; Zhang, P. Transcriptome profile analysis of resistance induced by burdock fructooligosaccharide in tobacco. J. Plant Physiol. 2012, 169, 1511–1519. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yu, G.; Li, Y.; Shen, L.; Qian, Y.; Yang, J.; Wang, F. Inhibitory effects of sulfated lentinan with different degree of sulfation against tobacco mosaic virus (TMV) in tobacco seedlings. Pestic. Biochem. Physiol. 2015, 122, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Jan, F.; Věra, A.; David, H. Phosphoproteomic studies in Arabidopsis and tobacco male gametophytes. Biochem. Soc. Trans. 2014, 42, 383–387. [Google Scholar] [CrossRef]
- Zhao, X.; She, X.; Du, Y.; Liang, X. Induction of antiviral resistance and stimulary effect by oligochitosan in tobacco. Pestic. Biochem. Physiol. 2007, 87, 78–84. [Google Scholar] [CrossRef]
- Hoeller, K.; Kiraly, L.; Kuenstler, A.; Mueller, M.; Gullner, G.; Fattinger, M.; Zechmann, B. Enhanced Glutathione Metabolism Is Correlated with Sulfur-Induced Resistance in Tobacco mosaic Virus-lnfected Genetically Susceptible Nicotiana tabacum Plants. Mol. Plant-Microbe Interact. 2010, 23, 1448–1459. [Google Scholar] [CrossRef]
- Jia, X.; Meng, Q.; Zeng, H.; Wang, W.; Yin, H. Chitosan oligosaccharide induces resistance to Tobacco mosaic virus in Arabidopsis via the salicylic acid-mediated signalling pathway. Sci. Rep. 2016, 6, 26144. [Google Scholar] [CrossRef]
- Fan, J.; Chen, C.; Yu, Q.; Brlansky, R.H.; Li, Z.G.; Gmitter, F.G. Comparative iTRAQ proteome and transcriptome analyses of sweet orange infected by “Candidatus Liberibacter asiaticus”. Physiol. Plant. 2011, 143, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Yanni, Z.; Chunxia, Z.; Yanli, L.; Yuwei, C.; Junjie, Z.; Zhongda, Z.; Xin, L.; Guowang, X. Study of metabolite differences of flue-cured tobacco from different regions using a pseudotargeted gas chromatography with mass spectrometry selected-ion monitoring method. J. Sep. Sci. 2014, 37, 2177–2184. [Google Scholar] [CrossRef]
- Wang, W.; Scali, M.; Vignani, R.; Spadafora, A.; Sensi, E.; Mazzuca, S.; Cresti, M. Protein extraction for two-dimensional electrophoresis from olive leaf, a plant tissue containing high levels of interfering compounds. Electrophoresis 2010, 24, 2369–2375. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Low, T.Y.; Hennrich, M.L.; Toorn, H.V.D.; Schwend, T.; Zou, H.; Mohammed, S.; Heck, A.J.R. Enhancing the Identification of Phosphopeptides from Putative Basophilic Kinase Substrates Using Ti (IV) Based IMAC Enrichment. Mol. Cell. Proteom. 2013, 12, 2673. [Google Scholar] [CrossRef]
- Andrews, G.L.; Simons, B.L.; Young, J.B.; Hawkridge, A.M.; Muddiman, D.C. Performance Characteristics of a New Hybrid Triple Quadrupole Time-of-Flight Tandem Mass Spectrometer. Anal. Chem. 2011, 83, 5442–5446. [Google Scholar] [CrossRef] [PubMed]
- Edwards, K.D.; Fernandez-Pozo, N.; Drake-Stowe, K.; Humphry, M.; Evans, A.D.; Bombarely, A.; Allen, F.; Hurst, R.; White, B.; Kernodle, S.P. A reference genome for Nicotiana tabacum enables map-based cloning of homeologous loci implicated in nitrogen utilization efficiency. Bmc Genom. 2017, 18, 448. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Zhang, Y.; Cui, H.; Liu, J.; Wu, Y.; Cheng, Y.; Xu, H.; Huang, X.; Li, S.; Zhou, A. WEGO 2.0: A web tool for analyzing and plotting GO annotations, 2018 update. Nucleic Acids Res. 2018, 46. [Google Scholar] [CrossRef]
- Drago, L.; Bortolin, M.; Taschieri, S.; Vecchi, E.D.; Agrappi, S.; Fabbro, M.D.; Francetti, L.; Mattina, R. Erythritol/chlorhexidine combination reduces microbial biofilm and prevents its formation on titanium surfaces in vitro. J. Oral Pathol. Med. 2017, 46, 625. [Google Scholar] [CrossRef]
- Ozyürek, M.; Güngör, N.; Baki, S.; Güçlü, K.; Apak, R. Development of a silver nanoparticle-based method for the antioxidant capacity measurement of polyphenols. Anal. Chem. 2012, 84, 8052–8059. [Google Scholar] [CrossRef]
- Pratt, H.M. In vivo Conversion of 5-Oxoproline to Glutamate by Higher Plants. Plant Physiol. 1976, 57, 85–87. [Google Scholar] [CrossRef]
- Zhang, M.; Lv, D.; Ge, P.; Bian, Y.; Chen, G.; Zhu, G.; Li, X.; Yan, Y. Phosphoproteome analysis reveals new drought response and defense mechanisms of seedling leaves in bread wheat (Triticum aestivum L.). J. Proteom. 2014, 109, 290–308. [Google Scholar] [CrossRef] [PubMed]
- Normile, D.; Pennisi, E. The rice genome. Rice: Boiled down to bare essentials. Science 2002, 296, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Qingyou, X.; Zeyang, Z.; Cheng, L.; Daojun, C.; Fangyin, D.; Bin, L.; Ping, Z.; Xingfu, Z.; Tingcai, C.; Chunli, C. A draft sequence for the genome of the domesticated silkworm (Bombyx mori). Science 2004, 306, 1937–1940. [Google Scholar] [CrossRef]
- Wang, K.; Zhao, Y.; Li, M.; Gao, F.; Yang, M.K.; Wang, X.; Li, S.; Yang, P. Analysis of phosphoproteome in rice pistil. Proteomics 2015, 14, 2319–2334. [Google Scholar] [CrossRef] [PubMed]
- Van Wijk, K.J.; Friso, G.; Walther, D.; Schulze, W.X. Meta-Analysis of Arabidopsis thaliana Phospho-Proteomics Data Reveals Compartmentalization of Phosphorylation Motifs. Plant Cell 2014, 26, 2367–2389. [Google Scholar] [CrossRef]
- Hou, Y.; Qiu, J.; Tong, X.; Wei, X.; Nallamilli, B.R.; Wu, W.; Huang, S.; Zhang, J. A comprehensive quantitative phosphoproteome analysis of rice in response to bacterial blight. Bmc Plant Biol. 2015, 15, 163. [Google Scholar] [CrossRef]
- Guo, M.; Huang, B.X. Integration of phosphoproteomic, chemical, and biological strategies for the functional analysis of targeted protein phosphorylation. Proteomics 2013, 13, 424–437. [Google Scholar] [CrossRef]
- Ardito, F.; Giuliani, M.; Perrone, D.; Troiano, G.; Muzio, L.L. The crucial role of protein phosphorylation in cell signaling and its use as targeted therapy (Review). Int. J. Mol. Med. 2017, 40, 271–280. [Google Scholar] [CrossRef] [Green Version]
- Lorenzo, O.; Rodríguez, D.; Rodríguez, P.L. A New Protein Phosphatase 2C (FsPP2C1) Induced by Abscisic Acid Is Specifically Expressed in Dormant Beechnut Seeds. Plant Physiol. 2001, 125, 1949–1956. [Google Scholar] [CrossRef] [Green Version]
- Ton, J.; Flors, V.; Mauch-Mani, B. The multifaceted role of ABA in disease resistance. Trends Plant Sci. 2009, 14, 310–317. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.J.; Shimono, M.; Sugano, S.; Kojima, M.; Yazawa, K.; Yoshida, R.; Inoue, H.; Hayashi, N.; Sakakibara, H.; Takatsuji, H. Abscisic acid interacts antagonistically with salicylic acid signaling pathway in rice-Magnaporthe grisea interaction. Mol. Plant-Microbe Interact. 2010, 23, 791–798. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, D.; Zhang, C.; Xia, X.; Yin, W.; Tian, Q. A Putative PP2C-Encoding Gene Negatively Regulates ABA Signaling in Populus euphratica. PLoS ONE 2015, 10, e0139466. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.M.; Yu, Y.; Lin, C.M.; Cui, N.; Zhao, J.Y.; Song, T.F.; Fan, H.Y. Glucohexaose-induced protein phosphatase 2C regulates cell redox status of cucumber seedling. J. Biosci. 2018, 43, 1–10. [Google Scholar] [CrossRef]
- Takeuchi, J.; Okamoto, M.; Akiyama, T.; Muto, T.; Yajima, S.; Sue, M.; Seo, M.; Kanno, Y.; Kamo, T.; Endo, A. Designed abscisic acid analogs as antagonists of PYL-PP2C receptor interactions. Nat. Chem. Biol. 2014, 10, 477–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosatéllez, S.; Anoman, A.D.; Florestornero, M.; Toujani, W.; Alseekh, S.; Fernie, A.R.; Nebauer, S.G.; Muñozbertomeu, J.; Segura, J.; Ros, R. Phosphoglycerate kinases are co-regulated to adjust metabolism and to optimize growth. Plant Physiol. 2018, 176. [Google Scholar] [CrossRef]
- Cheng, S.F.; Huang, Y.P.; Chen, L.H.; Hsu, Y.H.; Tsai, C.H. Chloroplast phosphoglycerate kinase is involved in the targeting of Bamboo mosaic virus to chloroplasts in Nicotiana benthamiana plants. Plant Physiol. 2013, 163, 1598–1608. [Google Scholar] [CrossRef] [PubMed]
- Cowan, G.H.; Roberts, A.G.; Chapman, S.N.; Ziegler, A.; Savenkov, E.I.; Torrance, L. The potato mop-top virus TGB2 protein and viral RNA associate with chloroplasts and viral infection induces inclusions in the plastids. Front. Plant Sci. 2012, 3, 290. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.W.; Ding, M.P.; Hsu, Y.H.; Tsai, C.H. Chloroplast phosphoglycerate kinase, a gluconeogenetic enzyme, is required for efficient accumulation of Bamboo mosaic virus. Nucleic Acids Res. 2007, 35, 424–432. [Google Scholar] [CrossRef]
- Prem, P.D.; Lin, Q.; Sek-Man, W. A study of certain aspects of sulfer metabolism of tobacco mosaic virus infected Nicotiana tabacum L. J. Proteom. 1965, in press. [Google Scholar] [CrossRef]
- Wang, Y.M.; Xu, W.G.; Hu, L.; Zhang, L.; Li, Y.; Du, X.H. Expression of Maize Gene Encoding C 4 -Pyruvate Orthophosphate Dikinase (PPDK) and C 4 -Phosphoenolpyruvate Carboxylase (PEPC) in Transgenic Arabidopsis. Plant Mol. Biol. Reporter 2012, 30, 1367–1374. [Google Scholar] [CrossRef]
- Chastain, C.J.; Failing, C.J.; Manandhar, L.; Zimmerman, M.A.; Lakner, M.M.; Nguyen, T.H.T. Functional evolution of C4 pyruvate, orthophosphate dikinase. J. Exp. Bot. 2011, 62, 3083–3091. [Google Scholar] [CrossRef] [Green Version]
- Du, X.; Xu, W.; Hu, L.; Zhang, L.; Li, Y.; Qi, X.; Wang, H.; Wang, Y. Response of Maize C_4-type PEPC and PPDK Transgenic Arabidopsis Plants to Drought-Stress. Mol. Plant Breed. 2013, 11, 477–484. [Google Scholar] [CrossRef]
- Moons, A.; Valcke, R.; Van, M. Low-oxygen stress and water deficit induce cytosolic pyruvate orthophosphate dikinase (PPDK) expression in roots of rice, a C3 plant. Plant J. 2010, 15, 89–98. [Google Scholar] [CrossRef]
- Dong, X.M.; Li, Y.; Chao, Q.; Shen, J.; Gong, X.J.; Zhao, B.G.; Wang, B.C. Analysis of gene expression and histone modification between C4 and non-C4 homologous genes of PPDK and PCK in maize. Photosynth. Res. 2016, 129, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Moldenhauer, K.A.K. Rice Biotechnology: Improving Yield, Stress Tolerance and Grain Quality. Novartis Found. Symp. 2008, 236, 100–111. [Google Scholar] [CrossRef]
- Howard, A.; Rogers, A.N. Role of translation initiation factor 4G in lifespan regulation and age-related health. Ageing Res. Rev. 2014, 13, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Piron, F.; Nicolaï, M.; Minoïa, S.; Piednoir, E.; Moretti, A.; Salgues, A.; Zamir, D.; Caranta, C.; Bendahmane, A. An Induced Mutation in Tomato eIF4E Leads to Immunity to Two Potyviruses. PLoS ONE 2010, 5, e11313. [Google Scholar] [CrossRef]
- Yoshii, M.; Nishikiori, M.; Tomita, K.; Yoshioka, N.; Kozuka, R.; Naito, S.; Ishikawa, M. The Arabidopsis cucumovirus multiplication 1 and 2 loci encode translation initiation factors 4E and 4G. J. Virol. 2004, 78, 6102. [Google Scholar] [CrossRef]
- Roberts, R.; Mayberry, L.K.; Browning, K.S.; Rakotondrafara, A.M. TheTriticum Mosaic Virus5’ Leader Binds to Both eIF4G and eIFiso4G for Translation. PLoS ONE 2017, 12, e0169602. [Google Scholar] [CrossRef]
- Grüner, S.; Peter, D.; Weber, R.; Wohlbold, L.; Chung, M.Y.; Weichenrieder, O.; Valkov, E.; Igreja, C.; Izaurralde, E. The Structures of eIF4E-eIF4G Complexes Reveal an Extended Interface to Regulate Translation Initiation. Mol. Cell 2016, 64, 467–479. [Google Scholar] [CrossRef]
- Nicaise, V.; Gallois, J.L.; Chafiai, F.; Allen, L.M.; Schurdi-Levraud, V.; Browning, K.S.; Candresse, T.; Caranta, C.; Gall, O.L.; German-Retana, S. Coordinated and selective recruitment of eIF4E and eIF4G factors for potyvirus infection in Arabidopsis thaliana. Febs Lett. 2007, 581, 1041–1046. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Ziegler, J.; Zeier, J.; Birkenbihl, R.P.; Somssich, I.E. Botrytis cinerea B05.10 promotes disease development in Arabidopsis by suppressing WRKY33-mediated host immunity. Plant Cell Environ. 2017, 40. [Google Scholar] [CrossRef] [PubMed]
- Zuyu, Z.; Synan Abu, Q.; Zhixiang, C.; Tesfaye, M. Arabidopsis WRKY33 transcription factor is required for resistance to necrotrophic fungal pathogens. Plant J. 2010, 48, 592–605. [Google Scholar] [CrossRef]
- Dai, S.; Zhang, Z.; Chen, S.; Beachy, R.N. RF2b, a rice bZIP transcription activator, interacts with RF2a and is involved in symptom development of rice tungro disease. Proc. Natl. Acad. Sci. USA 2004, 101, 687–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, S.; Zhang, Z.; Bick, J.; Beachy, R. Essential role of the Box II cis element and cognate host factors in regulating the promoter of Rice tungro bacilliform virus. J. Gen. Virol. 2006, 87, 715. [Google Scholar] [CrossRef] [PubMed]
- Dai, S.; Wei, X.; Alfonso, A.A.; Pei, L.; Duque, U.G.; Zhang, Z.; Babb, G.M.; Beachy, R.N. Transgenic rice plants that overexpress transcription factors RF2a and RF2b are tolerant to rice tungro virus replication and disease. Proc. Natl. Acad. Sci. USA 2008, 105, 21012–21016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, D.; Wang, Y.; Hamilton, M.; Ben-Hur, A.; Reddy, A.S. Transcriptome-Wide Identification of RNA Targets of Arabidopsis SERINE/ARGININE-RICH45 Uncovers the Unexpected Roles of This RNA Binding Protein in RNA Processing. Plant Cell 2015, 27, 3294–3308. [Google Scholar] [CrossRef]
- Manley, J.L.; Tacke, R.; Hogan, B.L.; Friedman, K.L.; Diller, J.D.; Ferguson, B.M. SR proteins and splicing control. Genes Dev. 1996, 10, 1569–1579. [Google Scholar] [CrossRef]
- Graveley, B.R. Sorting out the complexity of SR protein functions. Rna-a Publ. Rna Soc. 2000, 6, 1197–1211. [Google Scholar] [CrossRef] [Green Version]
- Paz, S.; Lu, M.L.; Takata, H.; Trautmann, L.; Caputi, M. SRSF1 RNA recognition motifs are strong inhibitors of HIV-1 replication. J. Virol. 2015, 89, 6275–6286. [Google Scholar] [CrossRef]
- Ajiro, M.; Shuang, T.; Doorbar, J.; Zheng, Z.M. Serine/Arginine-Rich Splicing Factor 3 and Heterogeneous Nuclear Ribonucleoprotein A1 Regulate Alternative RNA Splicing and Gene Expression of Human Papillomavirus 18 through Two Functionally Distinguishable cis Elements. J. Virol. 2016, 90, 9138. [Google Scholar] [CrossRef] [PubMed]
- Fíla, J.; Radau, S.; Matros, A.; Hartmann, A.; Scholz, U.; Feciková, J.; Mock, H.P.; Čapková, V.; Zahedi, R.P.; Honys, D. Phosphoproteomics Profiling of Tobacco Mature Pollen and Pollen Activated in vitro. Mol. Cell. Proteom. 2016, 15, 1338–1350. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.R.R.; Kirti, P.B. Novel role for a serine/arginine-rich splicing factor, AdRSZ21 in plant defense and HR-like cell death. Plant Mol. Biol. 2012, 80, 461. [Google Scholar] [CrossRef] [PubMed]
- Craigie, M.; Regan, P.; Otalora, Y.L.; Sariyer, I.K. Molecular interplay between T-Antigen and splicing factor, arginine/serine-rich 1 (SRSF1) controls JC virus gene expression in glial cells. Virol. J. 2015, 12, 196. [Google Scholar] [CrossRef] [PubMed]
- Mandadi, K.K.; Scholthof, K.B. Genome-wide analysis of alternative splicing landscapes modulated during plant-virus interactions in Brachypodium distachyon. Plant Cell 2015, 27, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Castillo, A.G.; Collinet, D.; Deret, S.; Kashoggi, A.; Bejarano, E.R. Dual interaction of plant PCNA with geminivirus replication accessory protein (Ren) and viral replication protein (Rep). Virology 2003, 312, 381–394. [Google Scholar] [CrossRef]
- Kushwaha, N.K.; Bhardwaj, M.; Chakraborty, S. The replication initiator protein of a geminivirus interacts with host monoubiquitination machinery and stimulates transcription of the viral genome. PLoS Pathog. 2017, 13, e1006587. [Google Scholar] [CrossRef]
- Castillo, A.G.; Kong, L.J.; Hanleybowdoin, L.; Bejarano, E.R. Interaction between a Geminivirus Replication Protein and the Plant Sumoylation System. J. Virol. 2004, 78, 2758. [Google Scholar] [CrossRef]
- Reyes, M.I.; Nash, T.E.; Dallas, M.M.; Ascencioibáñez, J.T.; Hanleybowdoin, L. Peptide aptamers that bind to geminivirus replication proteins confer a resistance phenotype to TYLCV and ToMoV infection in tomato. J. Virol. 2013, 87. [Google Scholar] [CrossRef]
- Liu, J.; Wang, H.; Gu, J.; Deng, T.; Yuan, Z.; Hu, B.; Xu, Y.; Yan, Y.; Zan, J.; Liao, M. BECN1-dependent CASP2 incomplete autophagy induction by binding to rabies virus phosphoprotein. Autophagy 2017, 13, 739–753. [Google Scholar] [CrossRef] [Green Version]
- Mauthe, M.; Reggiori, F. ATG proteins: Are we always looking at autophagy? Autophagy 2016, 12, 2502–2503. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wang, L.; Xiao, R.; Zhu, G.; Li, Y.; Liu, C.; Yang, R.; Tang, Z.; Li, J.; Huang, W. The invasion of tobacco mosaic virus RNA induces endoplasmic reticulum stress-related autophagy in HeLa cells. Biosci. Rep. 2012, 32, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Fujioka, Y.; Suzuki, S.W.; Noshiro, D.; Suzuki, H.; Kondokakuta, C.; Kimura, Y.; Hirano, H.; Ando, T.; Noda, N.N. The Intrinsically Disordered Protein Atg13 Mediates Supramolecular Assembly of Autophagy Initiation Complexes. Dev. Cell 2016, 38, 86–99. [Google Scholar] [CrossRef] [PubMed]
- An, C.; Mou, Z. Salicylic Acid and its Function in Plant Immunity. J. Integr. Plant Biol. 2011, 53, 412–428. [Google Scholar] [CrossRef] [PubMed]
- Kazan, K.; Manners, J.M. Linking development to defense: Auxin in plant-pathogen interactions. Trends Plant Sci. 2009, 14, 373–382. [Google Scholar] [CrossRef]
- Ghanashyam, C.; Jain, M. Role of auxin-responsive genes in biotic stress responses. Plant Signal. Behav. 2009, 4, 846–848. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Li, C.; Ge, J.; Xu, M.; Zhu, Q.; Wu, T.; Guo, A.; Xie, J.; Dong, H. Recessive mutation identifies auxin-repressed protein ARP1, which regulates growth and disease resistance in tobacco. Mol. Plant Microbe Interact. 2014, 27, 638–654. [Google Scholar] [CrossRef]
- Shi, H.Y.; Zhang, Y.X.; Chen, L. Two pear auxin-repressed protein genes, PpARP1 and PpARP2, are predominantly expressed in fruit and involved in response to salicylic acid signaling. Plant Cell Tissue Organ Culture 2013, 114, 279–286. [Google Scholar] [CrossRef]
- Su, J.; Jia, B.; Jia, S.; Ye, Z.F.; Wei, H.; Zhu, L.W. Effect of plant growth regulators on calyx abscission, fruit quality, and auxin-repressed protein (ARP) gene expression in fruitlets of ‘Dangshansuli’ pear (Pyrus bretschneideri Rehd.). J. Pomol. Hortic. Sci. 2015, 90, 135–142. [Google Scholar] [CrossRef]
- Ruiz-Medrano, R.; Jimenez-Moraila, B.; Herrera-Estrella, L.; Rivera-Bustamante, R.F. Nucleotide sequence of an osmotin-like cDNA induced in tomato during viroid infection. Plant Mol. Biol. 1992, 20, 1199–1202. [Google Scholar] [CrossRef]
- Singh, N.K.; Bracker, C.A.; Hasegawa, P.M.; Handa, A.K.; Buckel, S.; Hermodson, M.A.; Pfankoch, E.; Regnier, F.E.; Bressan, R.A. Characterization of osmotin: A thaumatin-like protein associated with osmotic adaptation in plant cells. Plant Physiol. 1987, 85, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Chen, T.H.; Li, P.H. Analysis of late-blight disease resistance and freezing tolerance in transgenic potato plants expressing sense and antisense genes for an osmotin-like protein. Planta 1996, 198, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Larosa, P.C.; Chen, Z.; Nelson, D.E.; Singh, N.K.; Hasegawa, P.M.; Bressan, R.A. Osmotin Gene Expression Is Posttranscriptionally Regulated. Plant Physiol. 1992, 100, 409–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhury, S.; Basu, A.; Kundu, S. Overexpression of a New Osmotin-Like Protein Gene (SindOLP) Confers Tolerance against Biotic and Abiotic Stresses in Sesame. Front. Plant Sci. 2017, 8, 410. [Google Scholar] [CrossRef] [PubMed]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, Z.-S.; Chen, Q.-S.; Zheng, Q.-X.; Shen, J.-J.; Luo, Z.-P.; Fan, K.; Xu, S.-H.; Shen, Q.; Liu, P.-P. Proteomic and Phosphoproteomic Analysis in Tobacco Mosaic Virus-Infected Tobacco (Nicotiana tabacum). Biomolecules 2019, 9, 39. https://doi.org/10.3390/biom9020039
Lu Z-S, Chen Q-S, Zheng Q-X, Shen J-J, Luo Z-P, Fan K, Xu S-H, Shen Q, Liu P-P. Proteomic and Phosphoproteomic Analysis in Tobacco Mosaic Virus-Infected Tobacco (Nicotiana tabacum). Biomolecules. 2019; 9(2):39. https://doi.org/10.3390/biom9020039
Chicago/Turabian StyleLu, Zi-Shu, Qian-Si Chen, Qing-Xia Zheng, Juan-Juan Shen, Zhao-Peng Luo, Kai Fan, Sheng-Hao Xu, Qi Shen, and Ping-Ping Liu. 2019. "Proteomic and Phosphoproteomic Analysis in Tobacco Mosaic Virus-Infected Tobacco (Nicotiana tabacum)" Biomolecules 9, no. 2: 39. https://doi.org/10.3390/biom9020039