Endo-β-1,3-glucanase (GH16 Family) from Trichoderma harzianum Participates in Cell Wall Biogenesis but Is Not Essential for Antagonism Against Plant Pathogens

 , , and
, , and 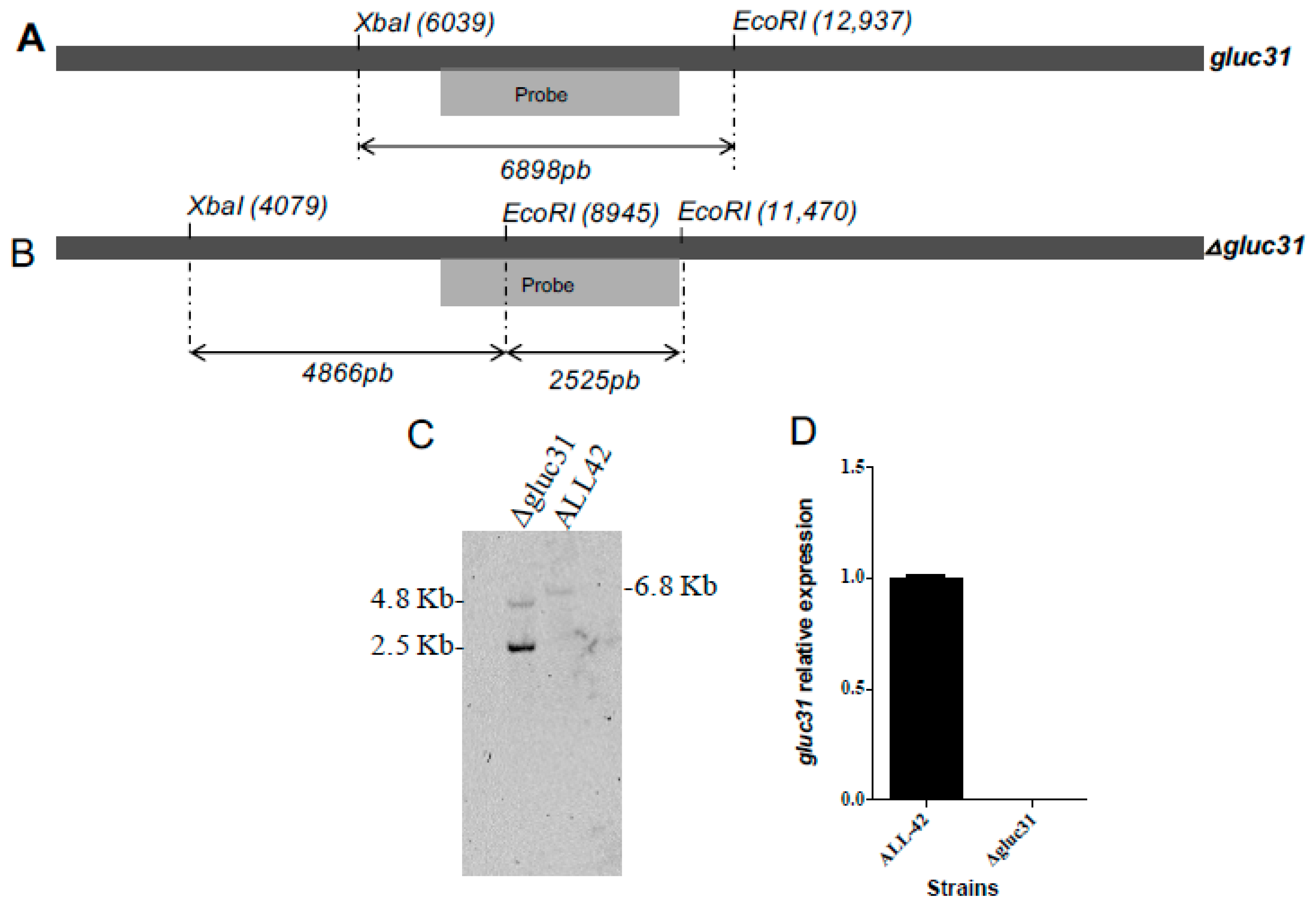{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture Conditions
2.2. Selection of the Gene for Knockout Studies and In Silico Analysis
2.3. Construction of the Δgluc31 T. harzianum Mutant Strain
2.4. Characterization of Transformants
2.5. Gene Expression Analysis
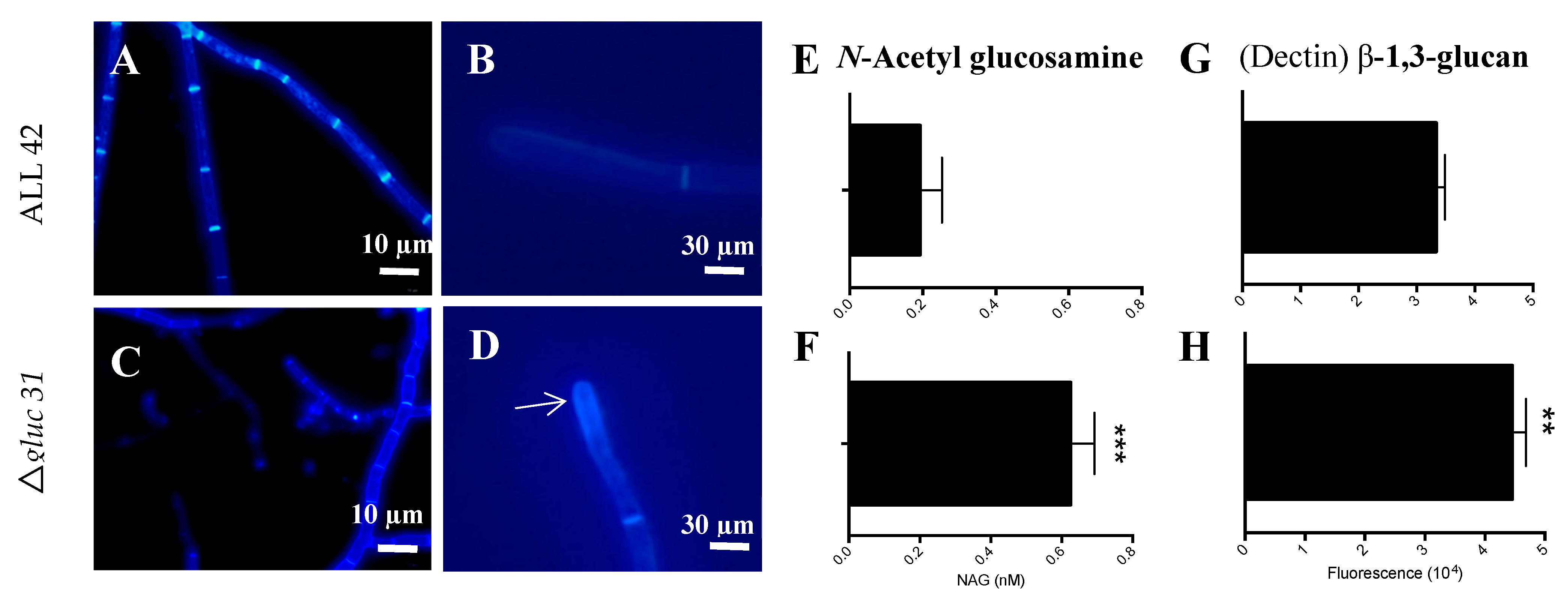
2.6. Extraction of Cell Wall Polysaccharides and Sugar Quantification
2.7. Fluorescence Microscopy
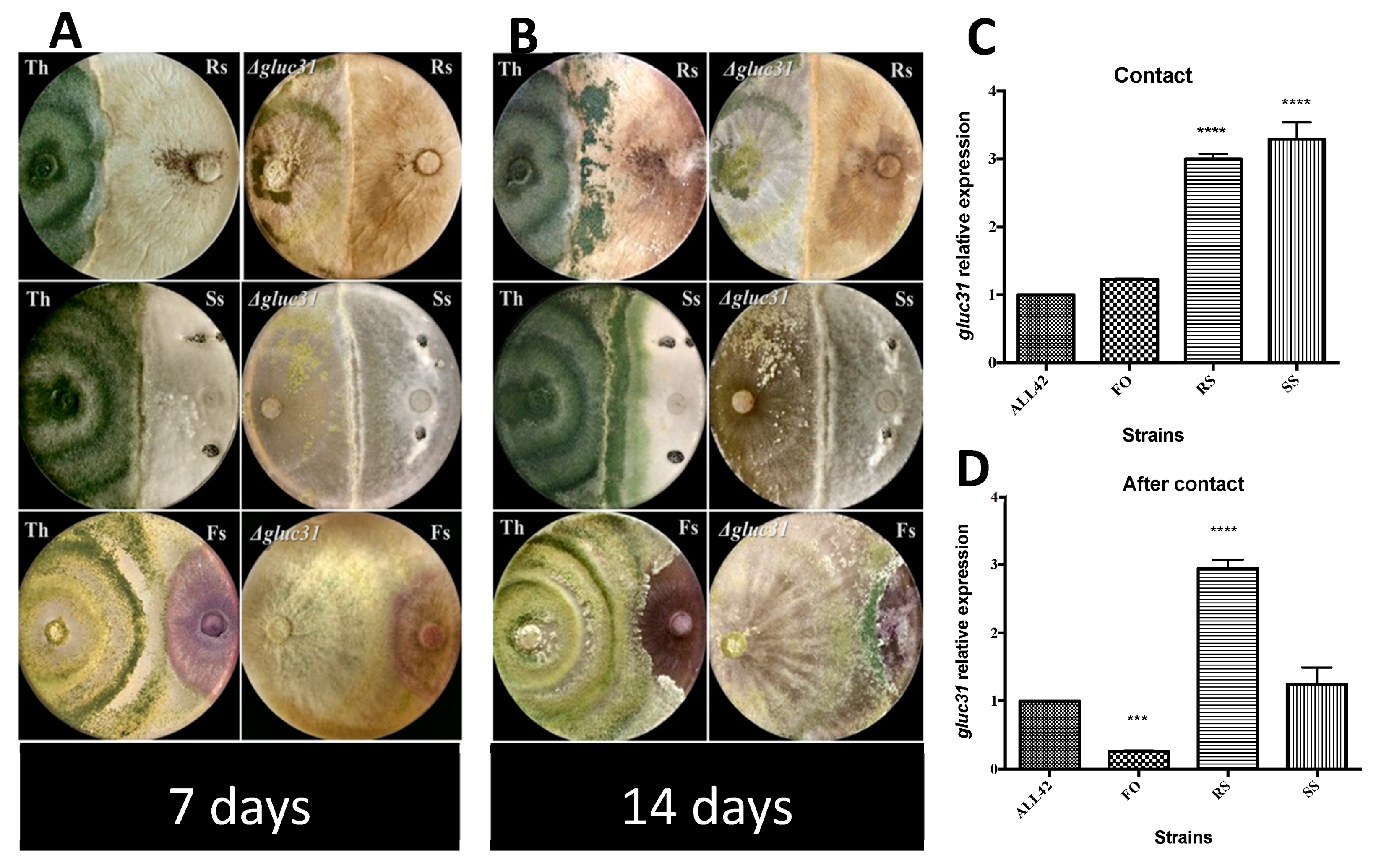
2.8. Direct Confrontation Assays
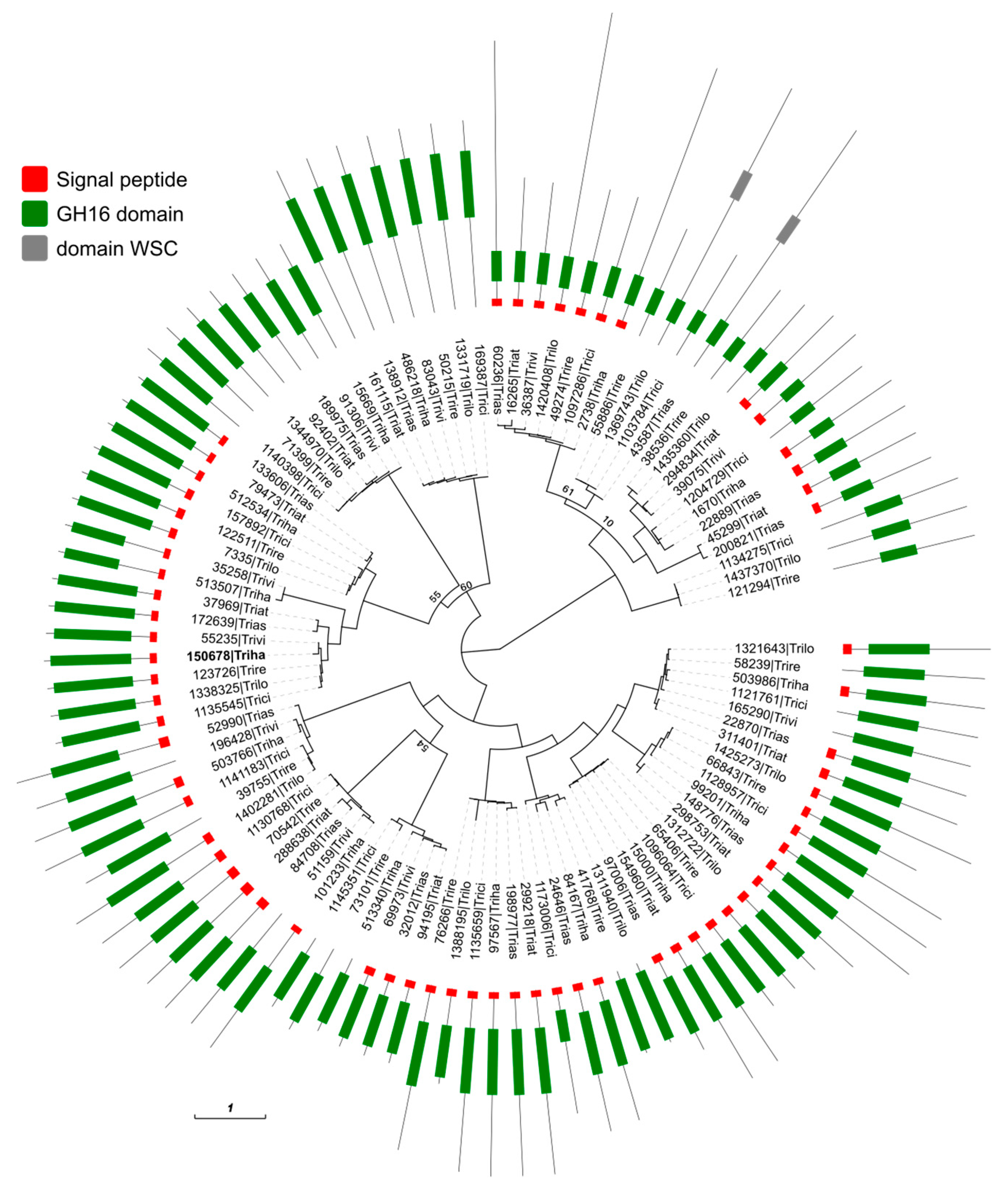
2.9. Phylogeny of the Glycoside Hydrolase Family 16 in the Trichoderma Genus
2.10. Enzyme Activities
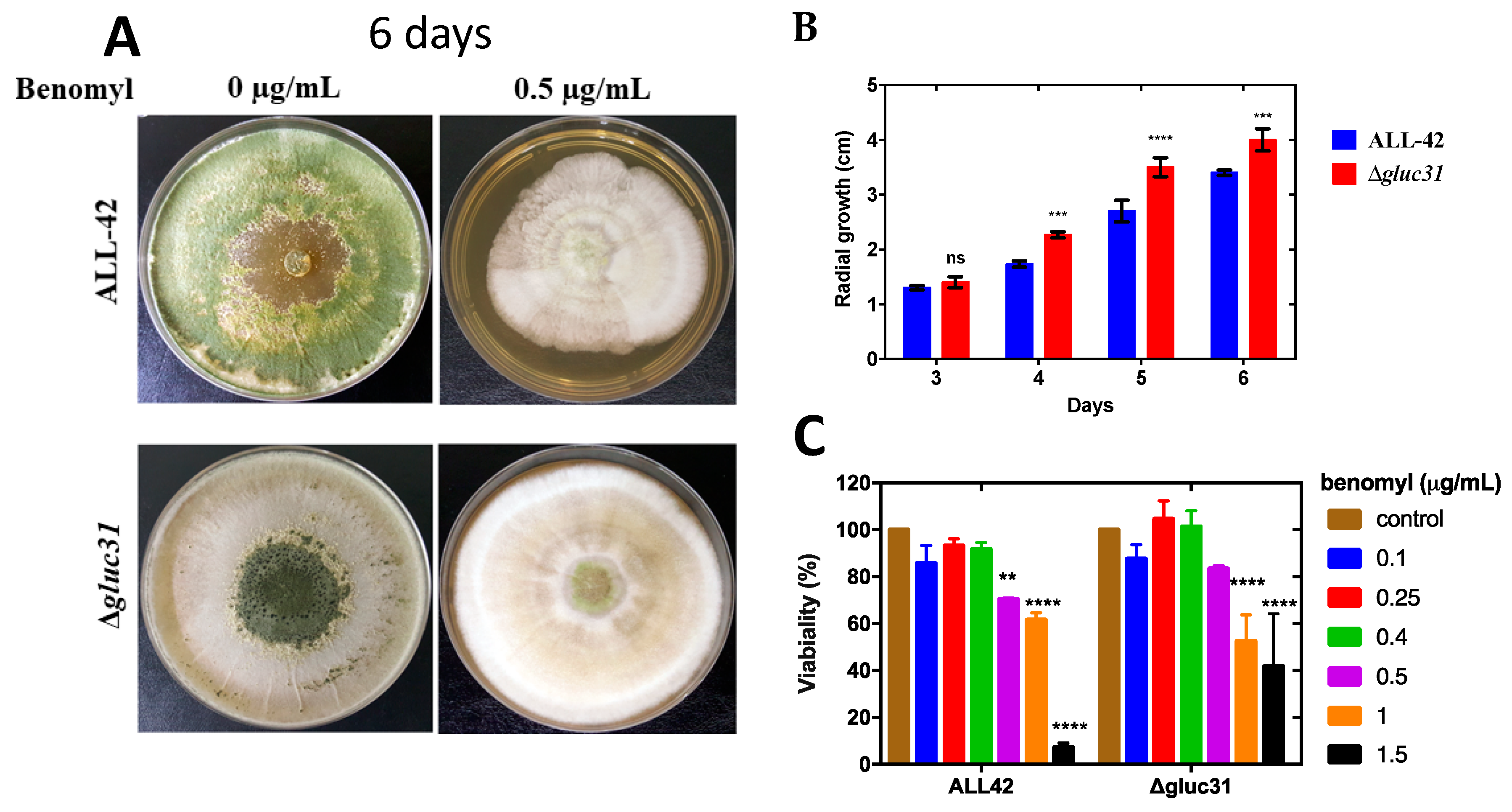
2.11. Alamar Blue Viability Assay
2.12. Statistical Analysis
3. Results
3.1. Sequence Analysis of the gluc31 Gene
3.2. Deletion of the gluc31 Gene from T. harzianum
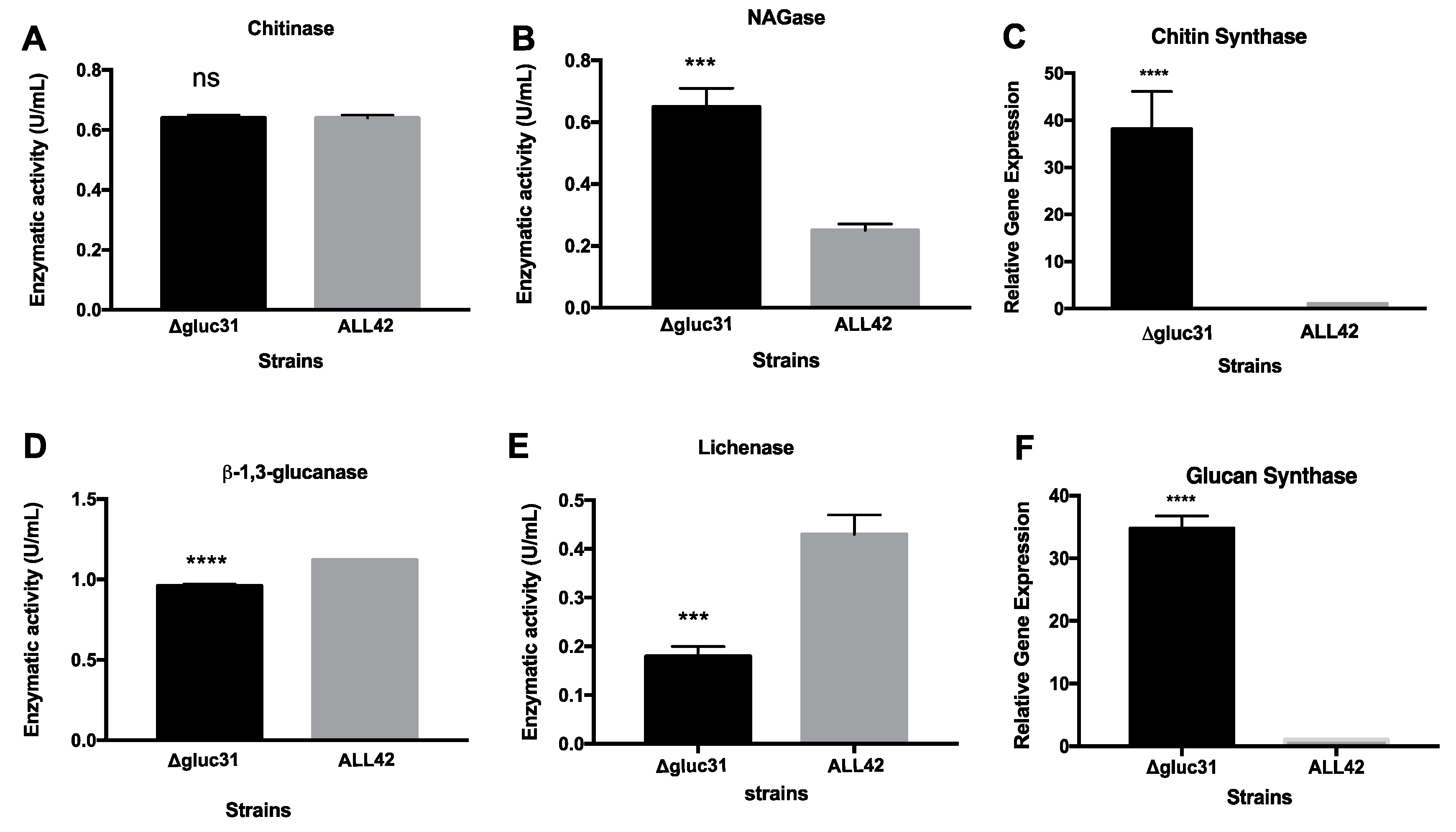
3.3. Gluc31 Is Involved in Cell Wall Remodeling of T. harzianum
3.4. Cell Viability Assay
3.5. The β-Glucanase Encoded by the gluc31 Gene Is Not Required for the Potential Antagonism of T. harzianum
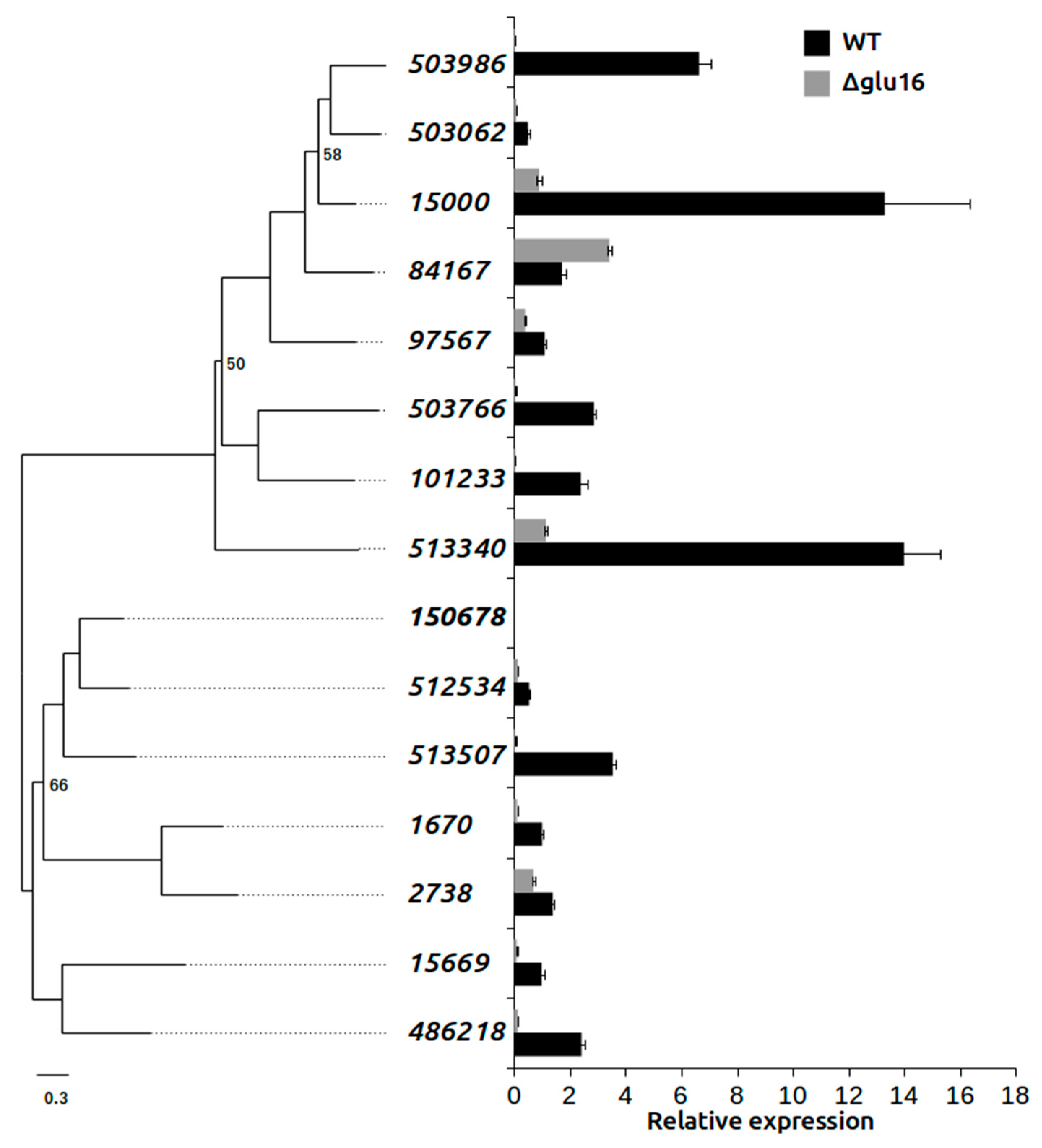
3.6. Effect on GH16 Family Gene Expression in Δgluc31 T. harzianum
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ait-Lahsen, H.; Soler, A.; Rey, M.; De La Cruz, J.; Monte, E.; Llobell, A. An Antifungal Exo-α-1,3-Glucanase (AGN13.1) from the Biocontrol Fungus Trichoderma harzianum. Appl. Environ. Microbiol. 2001, 67, 5833–5839. [Google Scholar] [CrossRef] [PubMed]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Woo, S.L.; Lorito, M. Trichoderma-plant-pathogen interactions. Soil Biol. Biochem. 2008, 40, 1–10. [Google Scholar] [CrossRef]
- Benítez, T.; Rincón, A.M.; Limón, M.C.; Codón, A.C. Biocontrol mechanisms of Trichoderma strains. Int. Microbiol. 2004, 7, 249–260. [Google Scholar] [PubMed]
- Harman, G.E. Overview of mechanisms and uses of Trichoderma spp. Am. Phytopathol. Soc. 2006, 96, 190–194. [Google Scholar] [CrossRef]
- Abubaker, K.S.; Sjaarda, C.; Castle, A.J. Regulation of three genes encoding cell-wall-degrading enzymes of Trichoderma aggressivum during interaction with Agaricus bisporus. Can. J. Microbiol. 2013, 59, 417–424. [Google Scholar] [CrossRef]
- Farkas, V. Biosynthesis of cell walls of fungi. Microbiol. Rev. 1979, 43, 117–144. [Google Scholar]
- Latgé, J.P. Tasting the fungal cell wall. Cell. Microbiol. 2010, 12, 863–872. [Google Scholar] [CrossRef]
- Martin, K.; McDougall, B.M.; McIlroy, S.; Chen, J.; Seviour, R.J. Biochemistry and molecular biology of exocellular fungal β-(1,3)- and β-(1,6)-glucanases. FEMS Microbiol. Rev. 2007, 31, 168–192. [Google Scholar] [CrossRef]
- Monteiro, V.N.; Ulhoa, C.J. Biochemical characterization of a β-1,3-glucanase from Trichoderma koningii induced by cell wall of Rhizoctonia solani. Curr. Microbiol. 2006, 52, 92–96. [Google Scholar] [CrossRef]
- Kück, U.; Hoff, B. New tools for the genetic manipulation of filamentous fungi. Appl. Microbiol. Biotechnol. 2010, 86, 51–62. [Google Scholar] [CrossRef]
- Bischof, R.; Seiboth, B. Molecular Tools for Strain Improvement of Trichoderma spp. Biotechnol. Biol. Trichoderma 2014, 179–191. [Google Scholar] [CrossRef]
- Seidl, V.; Song, L.; Lindquist, E.; Gruber, S.; Koptchinskiy, A.; Zeilinger, S.; Schmoll, M.; Martínez, P.; Sun, J.; Grigoriev, I.; et al. Transcriptomic response of the mycoparasitic fungus Trichoderma atroviride to the presence of a fungal prey. BMC Genom. 2009, 10, 567. [Google Scholar] [CrossRef] [PubMed]
- Reithner, B.; Brunner, K.; Schuhmacher, R.; Peissl, I.; Seidl, V.; Krska, R.; Zeilinger, S. The G protein α subunit Tga1 of Trichoderma atroviride is involved in chitinase formation and differential production of antifungal metabolites. Fungal Genet. Biol. 2005, 42, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Brunner, K.; Omann, M.; Pucher, M.E.; Delic, M.; Lehner, S.M.; Domnanich, P.; Kratochwill, K.; Druzhinina, I.; Denk, D.; Zeilinger, S. Trichoderma G protein-coupled receptors: Functional characterisation of a cAMP receptor-like protein from Trichoderma atroviride. Curr. Genet. 2008, 54, 283–299. [Google Scholar] [CrossRef]
- Gomes, E.V.; Costa, M.D.N.; De Paula, R.G.; Ricci De Azevedo, R.; Da Silva, F.L.; Noronha, E.F.; José Ulhoa, C.; Neves Monteiro, V.; Elena Cardoza, R.; Gutiérrez, S.; et al. The Cerato-Platanin protein Epl-1 from Trichoderma harzianum is involved in mycoparasitism, plant resistance induction and self cell wall protection. Sci. Rep. 2015, 5, 17998. [Google Scholar] [CrossRef]
- Carsolio, C.; Gutiérrez, A.; Jiménez, B.; Van Montagu, M.; Herrera-Estrella, A. Characterization of ech-42, a Trichoderma harzianum endochitinase gene expressed during mycoparasitism. Proc. Natl. Acad. Sci. USA 1994, 91, 10903–10907. [Google Scholar] [CrossRef]
- López-Mondéjar, R.; Catalano, V.; Kubicek, C.P.; Seidl, V. The β-N-acetylglucosaminidases NAG1 and NAG2 are essential for growth of Trichoderma atroviride on chitin. FEBS J. 2009, 276, 5137–5148. [Google Scholar] [CrossRef]
- Djonović, S.; Pozo, M.J.; Kenerley, C.M. Tvbgn3, a β-1,6-glucanase from the biocontrol fungus Trichoderma virens, is involved in mycoparasitism and control of Pythium ultimum. Appl. Environ. Microbiol. 2006, 72, 7661–7670. [Google Scholar] [CrossRef]
- Migheli, Q.; González-Candelas, L.; Dealessi, L.; Camponogara, A.; Ramón-Vidal, D. Transformants of Trichoderma longibrachiatum overexpressing the β-1,4-endoglucanase gene egl1 show enhanced biocontrol of Pythium ultimum on cucumber. Phytopathology 1998, 88, 673–677. [Google Scholar] [CrossRef]
- Steindorff, A.S.; do Nascimento Silva, R.; Coelho, A.S.G.; Nagata, T.; Noronha, E.F.; Ulhoa, C.J. Trichoderma harzianum expressed sequence tags for identification of genes with putative roles in mycoparasitism against Fusarium solani. Biol. Control 2012, 61, 134–140. [Google Scholar] [CrossRef]
- Vieira, P.M.; Coelho, A.S.G.; Steindorff, A.S.; de Siqueira, S.J.L.; do Nascimento Silva, R.; Ulhoa, C.J. Identification of differentially expressed genes from Trichoderma harzianum during growth on cell wall of Fusarium solani as a tool for biotechnological application. BMC Genom. 2013, 14, 177. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.H.; Hamari, Z.; Han, K.H.; Seo, J.A.; Reyes-Domínguez, Y.; Scazzocchio, C. Double-joint PCR: A PCR-based molecular tool for gene manipulations in filamentous fungi. Fungal Genet. Biol. 2004, 41, 973–981. [Google Scholar] [CrossRef] [PubMed]
- Mach, R.L.; Schindler, M.; Kubicek, C.P. Transformation of Trichoderma reesei based on hygromycin B resistance using homologous expression signals. Curr. Genet. 1994, 25, 567–570. [Google Scholar] [CrossRef] [PubMed]
- Gruber, F.; Visser, J.; Kubicek, C.P.; de Graaff, L.H. The development of a heterologous transformation system for the cellulolytic fungus Trichoderma reesei based on a pyrG-negative mutant strain. Curr. Genet. 1990, 18, 71–76. [Google Scholar] [CrossRef]
- Cardoza, R.E.; Malmierca, M.G.; Hermosa, M.R.; Alexander, N.J.; McCormick, S.P.; Proctor, R.H.; Tijerino, A.M.; Rumbero, A.; Monte, E.; Gutiérrez, S. Identification of loci and functional characterization of trichothecene biosynthesis genes in filamentous fungi of the genus Trichoderma. Appl. Environ. Microbiol. 2011, 77, 4867–4877. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- François, J.; Parrou, J.L. Reserve carbohydrates metabolism in the yeast Saccharomyces cerevisiae. FEMS Microbiol. Rev. 2001, 25, 125–145. [Google Scholar] [CrossRef]
- Ries, L.N.A.; Rocha, M.C.; de Castro, P.A.; Silva-Rocha, R.; Silva, R.N.; Freitas, F.Z.; de Assis, L.J.; Bertolini, M.C.; Malavazi, I.; Goldman, G.H. The Aspergillus fumigatus CrzA Transcription Factor Activates Chitin Synthase Gene Expression during the Caspofungin Paradoxical Effect. MBio 2017, 8, 2–15. [Google Scholar] [CrossRef]
- Riddell, R.W. Permanent Stained Mycological Preparations Obtained by Slide Culture. Mycologia 1950, 42, 265–270. [Google Scholar] [CrossRef]
- Bell, D.K. In Vitro Antagonism of Trichoderma species Against Six Fungal Plant Pathogens. Phytopathology 1982, 72, 28–35. [Google Scholar]
- Grigoriev, I.V.; Nikitin, R.; Haridas, S.; Kuo, A.; Ohm, R.; Otillar, R.; Riley, R.; Salamov, A.; Zhao, X.; Korzeniewski, F.; et al. MycoCosm portal: Gearing up for 1000 fungal genomes. Nucleic Acids Res. 2014, 42, D699–D704. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Zhang, H.; Gao, S.; Lercher, M.J.; Chen, W.H.; Hu, S. Evolview v2: An online visualization and management tool for customized and annotated phylogenetic trees. Nucleic Acids Res. 2016, 44, W236–W241. [Google Scholar] [CrossRef]
- Yang, S.; Qiaojuan, Y.; Jiang, Z.; Fan, G.; Wang, L. Biochemical characterization of a novel thermostable β-1,3-1,4- glucanase (Lichenase) from Paecilomyces thermophila. J. Agric. Food Chem. 2008, 56, 5345–5351. [Google Scholar] [CrossRef]
- Almeida, F.B.D.R.; Cerqueira, F.M.; Silva, R.D.N.; Ulhoa, C.J.; Lima, A.L. Mycoparasitism studies of Trichoderma harzianum strains against Rhizoctonia solani: Evaluation of coiling and hydrolytic enzyme production. Biotechnol. Lett. 2007, 29, 1189–1193. [Google Scholar] [CrossRef] [Green Version]
- Pardini, G.; De Groot, P.W.J.; Coste, A.T.; Karababa, M.; Klis, F.M.; De Koster, C.G.; Sanglard, D. The CRH family coding for cell wall glycosylphosphatidylinositol proteins with a predicted transglycosidase domain affects cell wall organization and virulence of Candida albicans. J. Biol. Chem. 2006, 281, 40399–40411. [Google Scholar] [CrossRef] [Green Version]
- Goto, M. Protein O-glycosylation in fungi: Diverse structures and multiple functions. Biosci. Biotechnol. Biochem. 2007, 71, 1415–1427. [Google Scholar] [CrossRef] [Green Version]
- Jiang, D.; Zhu, W.; Wang, Y.; Sun, C.; Zhang, K.Q.; Yang, J. Molecular tools for functional genomics in filamentous fungi: Recent advances and new strategies. Biotechnol. Adv. 2013, 31, 1562–1574. [Google Scholar] [CrossRef]
- Kalebina, T.S.; Farkaš, V.; Laurinavichiute, D.K.; Gorlovoy, P.M.; Fominov, G.V.; Bartek, P.; Kulaev, I.S. Deletion of BGL2 results in an increased chitin level in the cell wall of Saccharomyces cerevisiae. Antonie van Leeuwenhoek. Int. J. Gen. Mol. Microbiol. 2003, 84, 179–184. [Google Scholar]
- Dichtl, K.; Samantaray, S.; Aimanianda, V.; Zhu, Z.; Prévost, M.C.; Latgé, J.P.; Ebel, F.; Wagener, J. Aspergillus fumigatus devoid of cell wall β-1,3-glucan is viable, massively sheds galactomannan and is killed by septum formation inhibitors. Mol. Microbiol. 2015, 95, 458–471. [Google Scholar] [CrossRef] [PubMed]
- García-Rodriguez, L.J.; Trilla, J.A.; Castro, C.; Valdivieso, M.H.; Durán, A.; Roncero, C. Characterization of the chitin biosynthesis process as a compensatory mechanism in the fks1 mutant of Saccharomyces cerevisiae. FEBS Lett. 2000, 478, 84–88. [Google Scholar] [CrossRef] [Green Version]
- Mouyna, I.; Fontaine, T.; Vai, M.; Monod, M.; Fonzi, W.A.; Diaquin, M.; Popolo, L.; Hartlandt, R.P.; Latgé, J.P. Glycosylphosphatidylinositol-anchored glucanosyltransferases play an active role in the biosynthesis of the fungal cell wall. J. Biol. Chem. 2000, 275, 14882–14889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshimi, A.; Miyazawa, K.; Abe, K. Cell wall structure and biogenesis in aspergillus species. Biosci. Biotechnol. Biochem. 2016, 80, 1700–1711. [Google Scholar] [CrossRef] [Green Version]
- Kruszewska, J.S.; Piłsyk, S. Altering the fungal cell wall integrity and practical aspects of these modifications. In The Fungal Cell Wall; Mora-Montes, H.M., Ed.; Nova Science Publishers: New York, NY, USA, 2013; ISBN 978-1-62618-229-5. [Google Scholar]
- Mouyna, I.; Aimanianda, V.; Hartl, L.; Prevost, M.C.; Sismeiro, O.; Dillies, M.A.; Jagla, B.; Legendre, R.; Coppee, J.Y.; Latgé, J.P. GH16 and GH81 family β-(1,3)-glucanases in Aspergillus fumigatus are essential for conidial cell wall morphogenesis. Cell. Microbiol. 2016, 18, 1285–1293. [Google Scholar] [CrossRef] [Green Version]
- Gozalbo, D.; Roig, P.; Villamón, E.; Gil, M.L. Candida and candidiasis: The cell wall as a potential molecular target for antifungal therapy. Curr. Drug Targets Infect. Disord. 2004, 4, 117–135. [Google Scholar] [CrossRef]
- Lupetti, A.; Danesi, R.; Campa, M.; Del Tacca, M.; Kelly, S. Molecular basis of resistance to azole antifungals. Trends Mol. Med. 2002, 8, 76–81. [Google Scholar] [CrossRef]
- Kapteyn, J.C.; Van Den Ende, H.; Klis, F.M. The contribution of cell wall proteins to the organization of the yeast cell wall. Biochim. Biophys. Acta Gen. Subj. 1999, 1426, 373–383. [Google Scholar] [CrossRef]
- Osmond, B.C.; Specht, C.A.; Robbins, P.W. Chitin synthase III: Synthetic lethal mutants and “stress related” chitin synthesis that bypasses the CSD3/CHS6 localization pathway. Proc. Natl. Acad. Sci. USA 1999, 96, 11206–11210. [Google Scholar] [CrossRef] [Green Version]
- Popolo, L.; Gilardelli, D.; Bonfante, P.; Vai, M. Increase in chitin as an essential response to defects in assembly of cell wall polymers in the ggp1Δ mutant of Saccharomyces cerevisiae. J. Bacteriol. 1997, 179, 463–469. [Google Scholar] [CrossRef] [Green Version]
- Li, R.K.; Rinaldi, M.G. In vitro antifungal activity of nikkomycin Z in combination with fluconazole or itraconazole. Antimicrob. Agents Chemother. 1999, 43, 1401–1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atanasova, L.; Le Crom, S.; Gruber, S.; Coulpier, F.; Seidl-Seiboth, V.; Kubicek, C.P.; Druzhinina, I.S. Comparative transcriptomics reveals different strategies of trichoderma mycoparasitism. BMC Genom. 2013, 14, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrera-Estrella, A. Genome-Wide Approaches toward Understanding Mycotrophic Trichoderma Species. Biotechnol. Biol. Trichoderma 2014, 455–464. [Google Scholar] [CrossRef]
- Schmoll, M.; Dattenböck, C.; Carreras-Villaseñor, N.; Mendoza-Mendoza, A.; Tisch, D.; Alemán, M.I.; Baker, S.E.; Brown, C.; Cervantes-Badillo, M.G.; Cetz-Chel, J.; et al. The genomes of three uneven siblings: Footprints of the lifestyles of three trichoderma species. Microbiol. Mol. Biol. Rev. 2016, 80, 205–327. [Google Scholar] [CrossRef] [Green Version]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suriani Ribeiro, M.; Graciano de Paula, R.; Raquel Voltan, A.; de Castro, R.G.; Carraro, C.B.; José de Assis, L.; Stecca Steindorff, A.; Goldman, G.H.; Silva, R.N.; Ulhoa, C.J.; et al. Endo-β-1,3-glucanase (GH16 Family) from Trichoderma harzianum Participates in Cell Wall Biogenesis but Is Not Essential for Antagonism Against Plant Pathogens. Biomolecules 2019, 9, 781. https://doi.org/10.3390/biom9120781
Suriani Ribeiro M, Graciano de Paula R, Raquel Voltan A, de Castro RG, Carraro CB, José de Assis L, Stecca Steindorff A, Goldman GH, Silva RN, Ulhoa CJ, et al. Endo-β-1,3-glucanase (GH16 Family) from Trichoderma harzianum Participates in Cell Wall Biogenesis but Is Not Essential for Antagonism Against Plant Pathogens. Biomolecules. 2019; 9(12):781. https://doi.org/10.3390/biom9120781
Chicago/Turabian StyleSuriani Ribeiro, Marcela, Renato Graciano de Paula, Aline Raquel Voltan, Raphaela Georg de Castro, Cláudia Batista Carraro, Leandro José de Assis, Andrei Stecca Steindorff, Gustavo Henrique Goldman, Roberto Nascimento Silva, Cirano José Ulhoa, and et al. 2019. "Endo-β-1,3-glucanase (GH16 Family) from Trichoderma harzianum Participates in Cell Wall Biogenesis but Is Not Essential for Antagonism Against Plant Pathogens" Biomolecules 9, no. 12: 781. https://doi.org/10.3390/biom9120781