New Insights into Structural and Functional Roles of Indole-3-acetic acid (IAA): Changes in DNA Topology and Gene Expression in Bacteria




 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Growth Conditions, and Plasmids
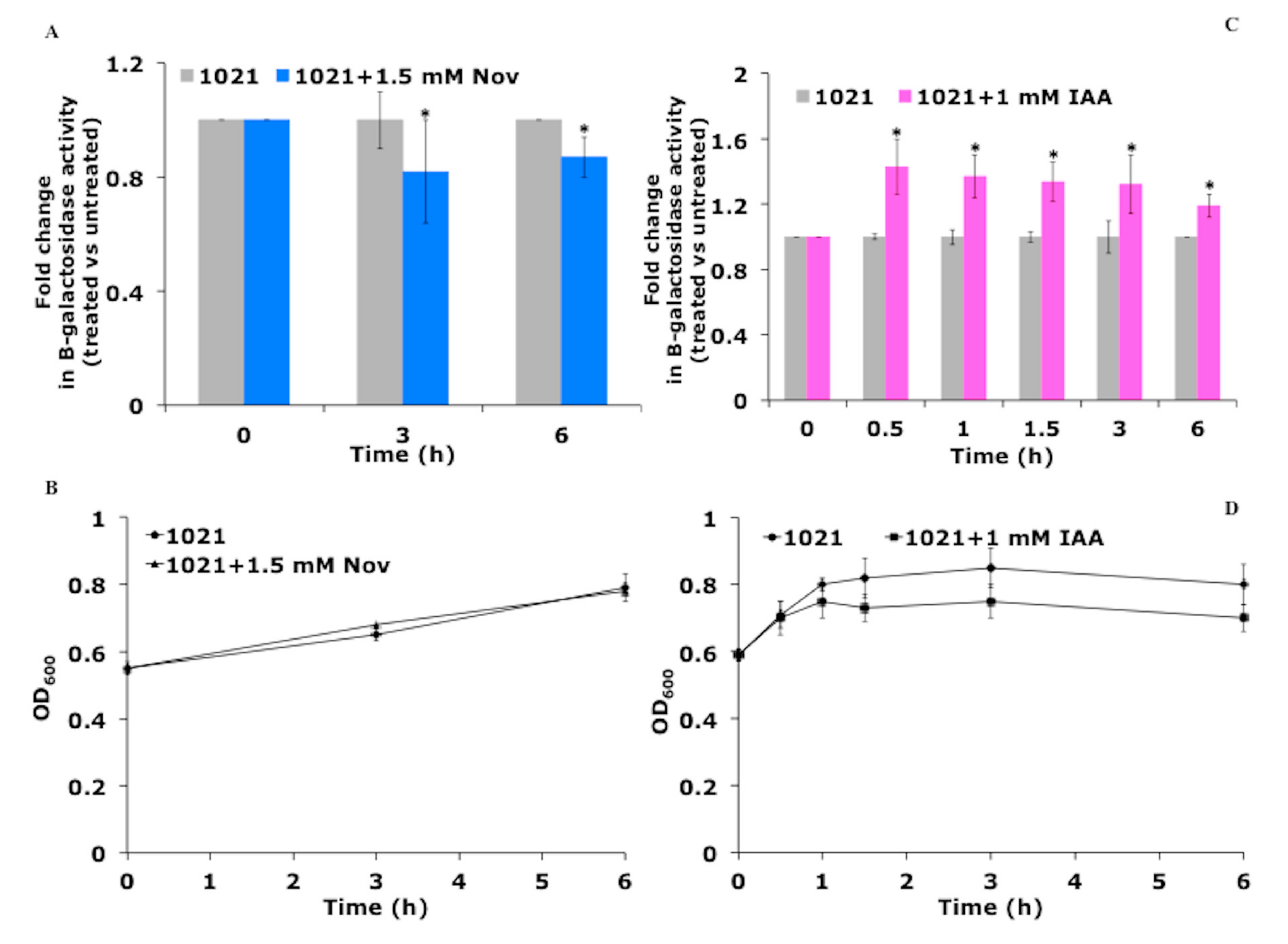
2.2. Evaluation of DNA Supercoiling In Vivo
2.3. DNA Binding Experiments
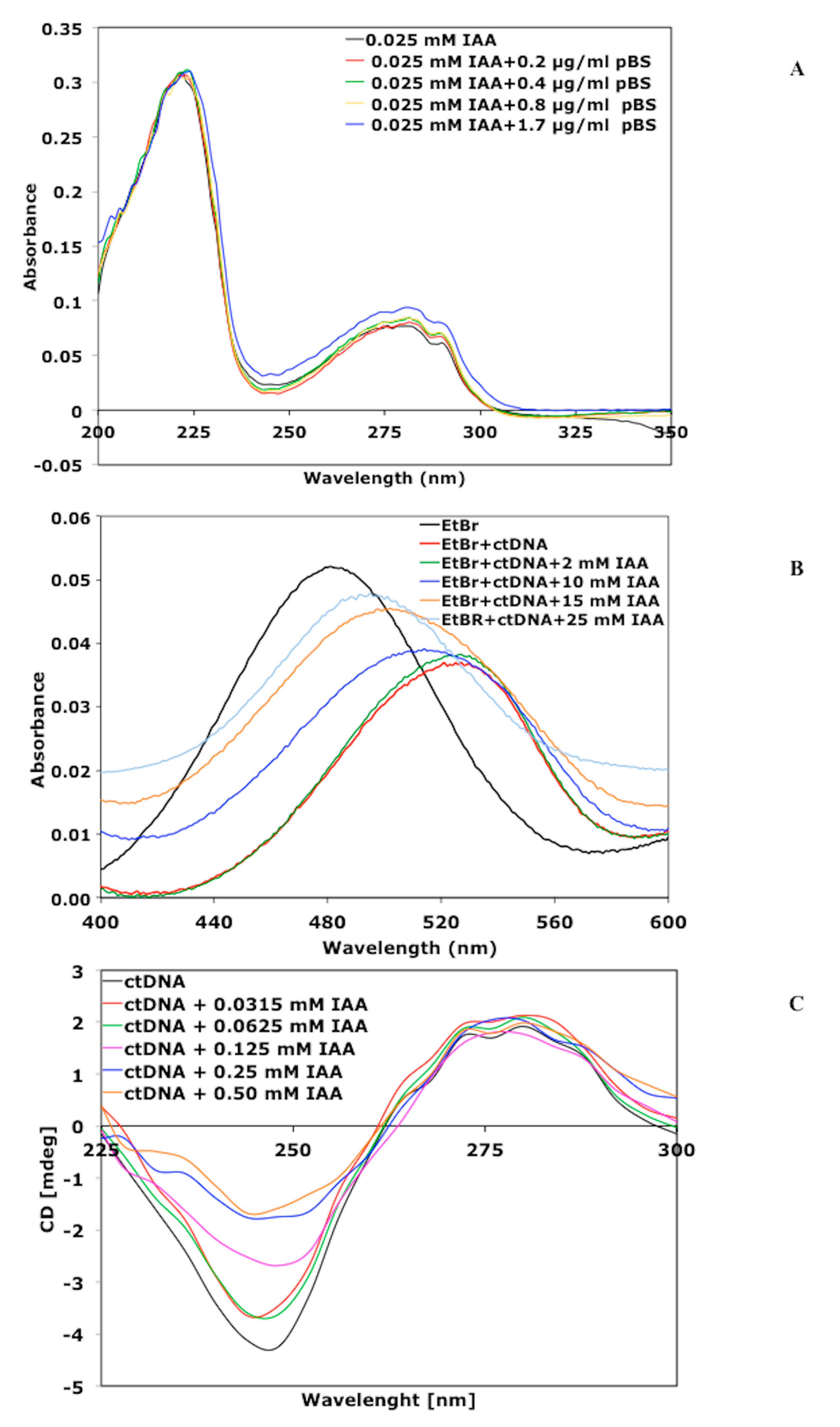
2.3.1. Absorption Spectra Titration
2.3.2. Competitive Study with EtBr
2.3.3. Circular Dichroism (CD) Analysis
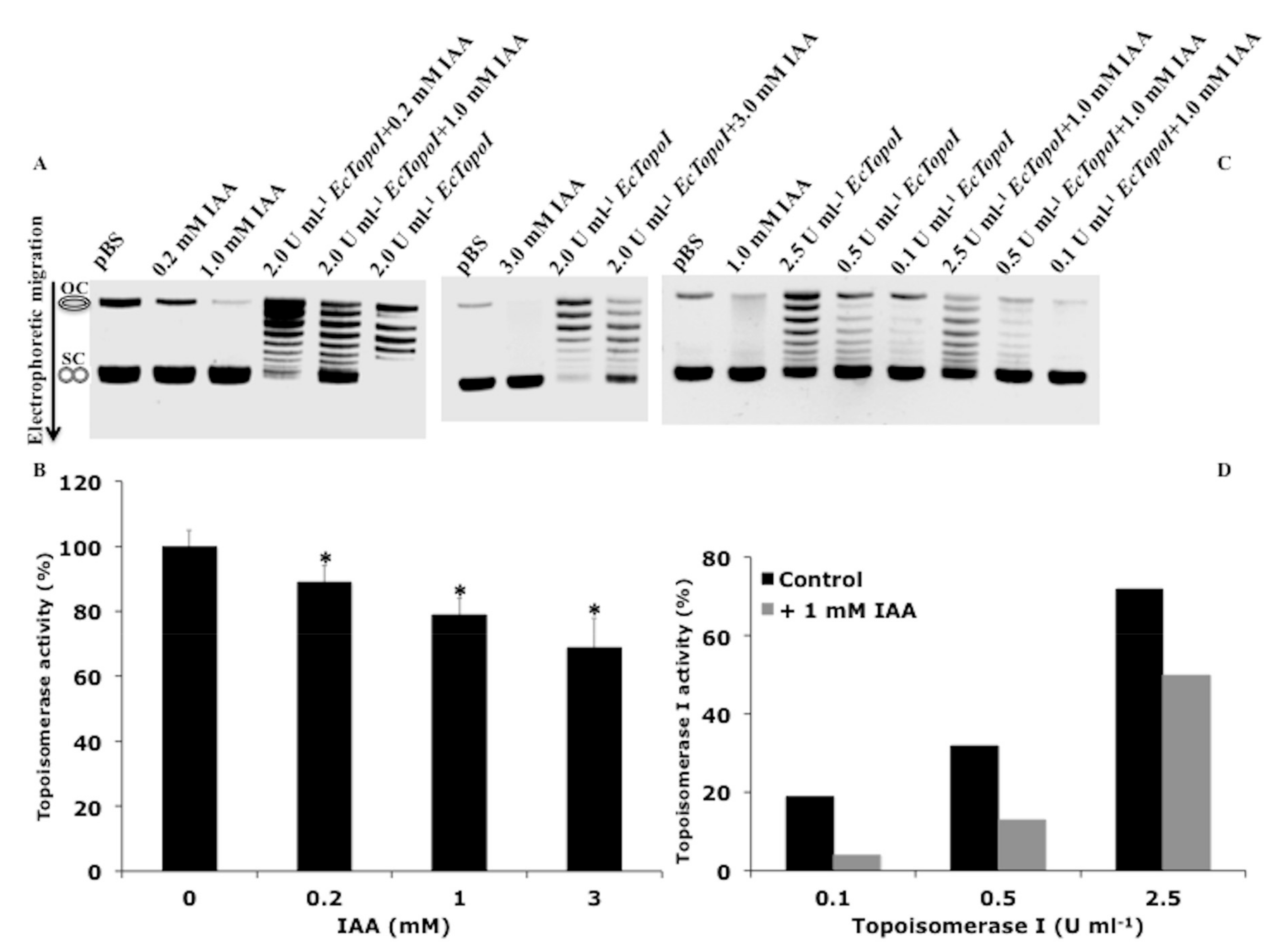
2.4. Topoisomerase-Mediated DNA Relaxation Assay
2.5. DNA Gyrase Assay
2.6. Quantitative Real-Time PCR (qRT-PCR) Analysis
3. Results
3.1. IAA Influences DNA Supercoiling In Vivo
3.2. IAA Interacts with DNA Inducing Alteration in Its Structure
3.2.1. Absorption Spectra of IAA in the Presence of Plasmid DNA
3.2.2. Competitive Studies with Ethidium Bromide (EtBr)
3.2.3. Circular dichroism (CD) Spectroscopic Analysis
3.3. IAA Affects the Relaxation Activity of E. coli DNA Topoisomerases
3.4. IAA Triggers the Expression of Genes Sensitive to a DNA Gyrase Inhibitor
3.5. nifA Promoter Activity is Stimulated by IAA and Inhibited by DNA Gyrase Inhibitor
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Darwin, C. The power of Movement in Plants; John Murray: London, UK, 1880. [Google Scholar]
- Went, F.W.; Thimann, K.V. Phytohormones. New York The Macmillan Company 1937, 251–283. [Google Scholar]
- De Melo, M.P.; Pithon-Curi, T.C.; Curi, R. Indole-3- acetic acid increases glutamine utilization by high peroxidase activity-presenting leucocytes. Life Sci. 2004, 75, 1713–1725. [Google Scholar] [CrossRef] [PubMed]
- Woodward, A.W.; Bartel, B. Auxin: Regulation, action, and interaction. Ann. Bot. 2005, 95, 707–735. [Google Scholar] [CrossRef] [PubMed]
- Prusty, R.; Grisafi, P.; Fink, G.R. The plant hormone indoleacetic acid induces invasive growth in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2004, 101, 4153–4157. [Google Scholar] [CrossRef] [PubMed]
- Bianco, C.; Imperlini, E.; Calogero, R.; Senatore, B.; Amoresano, A.; Carpentieri, A.; Pucci, P.; Defez, R. Indole-3-acetic acid improves Escherichia coli defence to stress. Arch. Microbiol. 2006, 185, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Bianco, C.; Imperlini, E.; Calogero, R.; Senatore, B.; Pucci, P.; Defez, R. Indole-3-acetic acid regulates the central metabolic pathways in Escherichia coli. Microbiology 2006, 152, 2421–2431. [Google Scholar] [CrossRef] [PubMed]
- Defez, R.; Spena, A. Method to control gene expression in bacteria, namely Rhizobiaceae, to improve root nodule development, nitrogen fixation and plant biomass production. US patent EP98/830674.2, 9 November 1998. [Google Scholar]
- Pandolfini, T.; Storlazzi, A.; Calabria, E.; Defez, R.; Spena, A. The spliceosomal intron of the rolA gene of Agrobacterium rhizogenes is a prokaryotic promoter. Mol. Microbiol. 2000, 35, 1326–1334. [Google Scholar] [CrossRef]
- Defez, R.; Esposito, R.; Angelini, A.; Bianco, C. Overproduction of indole-3-acetic acid in free-living rhizobia induces transcriptional changes resembling those occurring in nodule bacteroids. Mol. Plant-Microbe Interact. 2016, 29, 484–495. [Google Scholar] [CrossRef]
- Imperlini, E.; Bianco, C.; Lonardo, E.; Camerini, S.; Cermola, M.; Moschetti, G.; Defez, R. Effect of indole-3-acetic acid on Sinorhizobium meliloti survival and on symbiotic nitrogen fixation and stem dry weight production. Appl. Microbiol. Biotechnol. 2009, 83, 727–738. [Google Scholar] [CrossRef]
- Bianco, C.; Defez, R. Auxins upregulate nif and fix genes. Plant Signal. Behav. 2010, 5, 1290–1294. [Google Scholar] [CrossRef] [PubMed]
- Defez, R.; Andreozzi, A.; Bianco, C. The overproduction of indole-3-acetic acid (IAA) in endophytes upregulates nitrogen fixation in both bacterial cultures and inoculated rice plants. Microb. Ecol. 2017, 74, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Andreozzi, A.; Prietro, P.; Mercado-Blanco, J.; Monaco, S.; Zampieri, E.; Romano, S.; Valé, G.; Defez, R.; Bianco, C. The co-inoculation with nitrogen-fixing and indole-3-acetic (IAA)-producing bacterial endophytes positively affects rice (Oryza sativa L. cv. Baldo) rice. Env. Microbiol. 2019. [Google Scholar] [CrossRef]
- Fisher, H.M. Genetic regulation of nitrogen fixation in rhizobia. Microbiol. Rev. 1994, 58, 352–386. [Google Scholar]
- Liu, Y.-J.; Hu, B.; Zhu, J.-B.; Shen, S.-J.; Yu, G.-Q. nifH promoter is regulated by DNA supercoiling in Sinorhizobium meliloti. Acta Bioch. Bioph. Sin. 2005, 37, 221–226. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Witham, F.H.; Hendry, L.B.; Chapman, O.L. Chirality and stereochemical recognition in DNA-phytohormone interactions: A model approach. Origin of Life 1978, 9, 7–15. [Google Scholar] [CrossRef]
- Jacobsen, J.V. Regulation of ribonucleic acid metabolism by plant hormones. Ann. Rev. Plant Physiol. 1977, 28, 537–564. [Google Scholar] [CrossRef]
- Dorman, C. DNA supercoiling and bacterial gene expression. Sci. Prog. 2006, 89, 151–166. [Google Scholar] [CrossRef]
- Ma, J.; Wang, M.D. DNA supercoiling during transcription. Biophys. Rev. 2016, 8, S75–S87. [Google Scholar] [CrossRef]
- Zhi, X.; Dages, S.; Dages, K.; Liu, Y.; Hua, Z.-C.; Makemson, J.; Leng, F. Transient and dynamic DNA supercoiling potently stimulates the leu-500 promoter in Escherichia coli. J. Biol. Chem. 2017, 292, 14566–14575. [Google Scholar] [CrossRef]
- Champoux, J.J. DNA topoisomerases: Structure, function, and mechanism. Annu. Rev. Biochem. 2001, 70, 369–413. [Google Scholar] [CrossRef]
- Galibert, F.; Finan, T.M.; Long, S.R.; Puhler, A.; Abola, P.; Ampe, F.; Barloy-Hubler, F.; Barnett, M.J.; Becker, A.; Boistard, P.; et al. The composite genome of the legume symbiont Sinorhizobium meliloti. Science 2001, 293, 668–672. [Google Scholar] [CrossRef]
- Ditta, G.; Virts, E.; Palomares, A.; Kim, C.H. The nifA gene of Rhizobium meliloti is oxygen regulated. J. Bacteriol. 1987, 169, 3217–3223. [Google Scholar] [CrossRef]
- Miller, J.H. Esperiments in molecular genetics. In Cold Spring Harbor Laboratory Press; Cold Spring Harbor: New York, NY, USA, 1972. [Google Scholar]
- Smith, D.H.; Davis, B.D. Mode of action of Novobiocin in Escherichia coli. J. Bacteriol. 1967, 93, 71–79. [Google Scholar]
- Crozat, E.; Philippe, N.; Lenski, E.R.; Geiselmann, J.; Schneider, D. Long-term experimental evolution in Escherichia coli. XII. DNA topology as a key target selection. Genetics 2005, 169, 523–532. [Google Scholar] [CrossRef]
- Bettotti, P.; Visone, V.; Lunelli, L.; Perugino, G.; Ciaramella, M.; Valenti, A. Structure and properties of DNA molecules over the full range of biologically relevant supercoiling states. Sci. Rep. 2018, 8, 6163. [Google Scholar] [CrossRef]
- Bianco, C.; Defez, R. A Sinorhizobium meliloti IAA-overproducing strain improves phosphate solubilization and Medicago plant yield. Appl. Environ. Microbiol. 2010, 76, 4626–4632. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2–ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Gilbert, N.; Allan, J. Supercoiling in DNA and chromatin. Curr. Op. Genet. Dev. 2014, 25, 15–21. [Google Scholar] [CrossRef]
- González-Ruiz, V.; Olives, A.I.; Martin, M.A.; Ribelles, P.; Ramos, M.T.; Menéndez, C. An overview of analytical techniques employed to evidence drug-DNA interactions. Applications to the design of genosensors. In Biochemical engineering, Trends, Research and Techologies; NTECH Open Acces Publisher: Rijeka, Croatia, 2011; pp. 65–90. [Google Scholar]
- Biver, T. Use of UV-Vis spectrometry to gain information on the mode of binding of small molecules to DNAs and RNAs. Appl. Spectr. Rev. 2012, 47, 271–325. [Google Scholar] [CrossRef]
- Chang, Y.-M.; Chen, C.K.-M.; Hou, M.-H. Conformational changes in DNA upon ligand binding monitored by circular dichroism. Int. J. Sci. 2012, 13, 3394–3413. [Google Scholar] [CrossRef]
- Caric, D.; Tomisic, V.; Kveder, M.; Galic, N.; Pifat, G.; Magnus, V.; Soskic, M. Absorption and fluorescence spectra of ring-substituted indole-3-acetic acids. Biophys. Chem. 2004, 111, 247–257. [Google Scholar] [CrossRef]
- Dobrev, P.I.; Havlicek, L.; Vagner, M.; Malbeck, J.; Kaminek, M. Purification and determination of plant hormones auxin and abscisic acid using solid phase extraction and two-dimensional high performance liquid chromatography. J. Chromat. 2005, 1075, 159–166. [Google Scholar] [CrossRef]
- Meliani, A.; Bensoltane, A.; Benidire, L.; Oufdou, K. Plant growth-promotion and IAA secretion with Pseudomonas fluorescens and Pseudomonas putida. J. Bot. Sci. 2017, 6, 16–24. [Google Scholar]
- Baneriee, A.; Singh, J.; Dasgupta, D. Fluorescence spectroscopic and calorimetry based approaches to characterize the mode of interaction of small molecules with DNA. J. Fluoresc. 2013, 23, 745–752. [Google Scholar] [CrossRef]
- Sohrabi, N. Binding and UV/Vis spectral investigation of interaction of Ni(II) piroxicam complex with Calf thymus deoxyribonucleic acid (Ct-DNA): A thermodynamic approach. J. Pharm. Sci. Res. 2015, 7, 533–537. [Google Scholar]
- Salem, O.M.; Vilková, M.; Janocková, J.; Jendzelovsky, R.; Fedorocko, P.; Zilecká, E.; Kaspárková, J.; Brabec, V.; Imrich, J.; Kozurková, M. New spiro tria(thia)zolidine–acridines as topoisomerase inhibitors, DNA binders and cytostatic compounds. Int. J. Biol. Macrom. 2016, 86, 690–700. [Google Scholar] [CrossRef]
- Forterre, P.; Gribaldo, S.; Gadelle, D.; Serre, M.C. Origin and evolution of DNA topoisomerases. Biochimie 2007, 89, 427–446. [Google Scholar] [CrossRef]
- Forterre, P.; Gadelle, D. Phylogenomics of DNA topoisomerases: Their origin and putative roles in the emergence of modern organisms. Nucleic Acids Res. 2009, 37, 679–692. [Google Scholar] [CrossRef]
- Vvedenskaya, I.O.; Zhang, Y.; Goldman, S.R.; Valenti, A.; Visone, V.; Taylor, D.M.; Ebright, R.H.; Nickels, B.E. Massively systematic transcript end readout, “MASTER”: Transcription start site selection, transcriptional slippage, and transcript yields. Moll. Cell. 2015, 60, 953–965. [Google Scholar] [CrossRef][Green Version]
- Dormann, C.J.; Dormann, M.J. DNA supercoiling is a fundamental regulatory principle in the control of bacterial gene expression. Biophys. Rev. 2016, 8, S89–S100. [Google Scholar] [CrossRef]
- Snyder, R.D.; Ewing, D.; Hendry, L.B. DNA intercalative potential of marketed drugs testing positive in in vitro cytogenetics assays. Mut. Res. 2006, 609, 47–59. [Google Scholar] [CrossRef]
- Hendry, L.B.; Mahesh, V.B.; Bransome, E.D.; Ewing, D.E. Small molecule intercalation with double stranded DNA: Implications for normal gene regulation and for predicting the biological efficacy and genotoxicity of drugs and other chemicals. Mut. Res. 2007, 623, 53–71. [Google Scholar] [CrossRef]
- Szafran, M.J.; Gongerowska, M.; Gutkowski, P.; Zakrzewska-Czerwinska, J.; Jakimowicz, D. The coordinate positive regulation of topoisomerase genes maintains topological homeostasis in Streptomyces coelicolor. J. Bacteriol. 2016, 198, 3016–3028. [Google Scholar] [CrossRef]
- Ueshima, R.; Fujita, N.; Hishihama, A. DNA supercoiling and temperature shift the promoter activity of the Escherichia coli rpoH gene encoding the heat-shock sigma subunit of RNA polymerase. Mol. Gen. Genet. 1989, 215, 185–189. [Google Scholar] [CrossRef]
- Ogata, Y.; Mizushima, T.; Kataoka, K.; Kita, K.; Miki, T.; Sekimizu, K. DnaK heat shock protein of Escherichia coli maintains the negative supercoiling of DNA against thermal stress. J. Biol. Chem. 1996, 271, 29407–29414. [Google Scholar] [CrossRef]
- Lee, Y.-R.; Cheng, M.; Lee, J.D.; Zhang, J.; Lin, S.-Y.; Fu, T.-M.; Chen, H.; Ishikawa, T.; Chiang, S.-Y.; Katon, J.; et al. Reactivation of PTEN tumor suppressor for cancer treatment through inhibition of a MYC-WWP1 inhibitory pathway. Science 2019, 364, aau0159. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene | Forward (5′→3′) | Reverse (5′→3′) |
|---|---|---|---|
| SMc04040 | ibpA | GACCTATCCGCCCTACAACA | GAAGCGGACTTGATCTCGAC |
| SMc00646 | rpoH1 | GTGAGGAAGAGGTCGTCTCG | TCAGAACCTTCATGGCATTG |
| SMc02163 | pgi | GGCAAGAAGATCACCGATGT | GTCTCGATCGTGGTGAAGGT |
| SMa1227 | fixJ | CTCGTGACGGACCTGAGAAT | GCAGCAACCAGATGTTCAGA |
| SMa0815 | nifA | CCTTGCAAGAGCATTCCTTC | TCTTTGACCTGGCGAGAGTT |
| SMa1225 | fixK1 | CATTCTTTCTTTGCCGAAGC | CGCAAAGATCGACGAGAAAT |
| SMc02857 | dnaK | CCGAGTTCAAGAAGGAGCAG | AGCTTCATCGTCAGGTGCTT |
| SMc01364 SMc02482 SMc02481 SMc02480 SMc00869 SMc00768 | topA sucA sucD icd atpF2 aceA | CATCGACCGTGACTATGTGG AAGACCGTCGTCCAGCTCTA TGTTCCAGACGACCAATGAA AACCTGGACGAATCGATCAC GCTGCTTACGAGCAGGAGTT GGACGCTATTCCATCTGGTC | GCACGTCCTTCCAATTGAGT CGACCTCCTTCAACTGCTTC CCTCGTCTTTCAGGAACTGC TTCCTCGTCGAACACCTTCT GCAAGAGCCTTCGACTTGAT CGAGAACGTTGCGATTGTAG |
| SMc01317 | rpoB | CGTCAACAAGTACGGCTTCA | CGTCCATCAGGTTGATGTTG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Defez, R.; Valenti, A.; Andreozzi, A.; Romano, S.; Ciaramella, M.; Pesaresi, P.; Forlani, S.; Bianco, C. New Insights into Structural and Functional Roles of Indole-3-acetic acid (IAA): Changes in DNA Topology and Gene Expression in Bacteria. Biomolecules 2019, 9, 522. https://doi.org/10.3390/biom9100522
Defez R, Valenti A, Andreozzi A, Romano S, Ciaramella M, Pesaresi P, Forlani S, Bianco C. New Insights into Structural and Functional Roles of Indole-3-acetic acid (IAA): Changes in DNA Topology and Gene Expression in Bacteria. Biomolecules. 2019; 9(10):522. https://doi.org/10.3390/biom9100522
Chicago/Turabian StyleDefez, Roberto, Anna Valenti, Anna Andreozzi, Silvia Romano, Maria Ciaramella, Paolo Pesaresi, Sara Forlani, and Carmen Bianco. 2019. "New Insights into Structural and Functional Roles of Indole-3-acetic acid (IAA): Changes in DNA Topology and Gene Expression in Bacteria" Biomolecules 9, no. 10: 522. https://doi.org/10.3390/biom9100522
APA StyleDefez, R., Valenti, A., Andreozzi, A., Romano, S., Ciaramella, M., Pesaresi, P., Forlani, S., & Bianco, C. (2019). New Insights into Structural and Functional Roles of Indole-3-acetic acid (IAA): Changes in DNA Topology and Gene Expression in Bacteria. Biomolecules, 9(10), 522. https://doi.org/10.3390/biom9100522