The Differential DNA Hypermethylation Patterns of microRNA-137 and microRNA-342 Locus in Early Colorectal Lesions and Tumours

 , , ,
, , ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Genomic DNA Extraction
2.3. Sodium Bisulfite Treatment
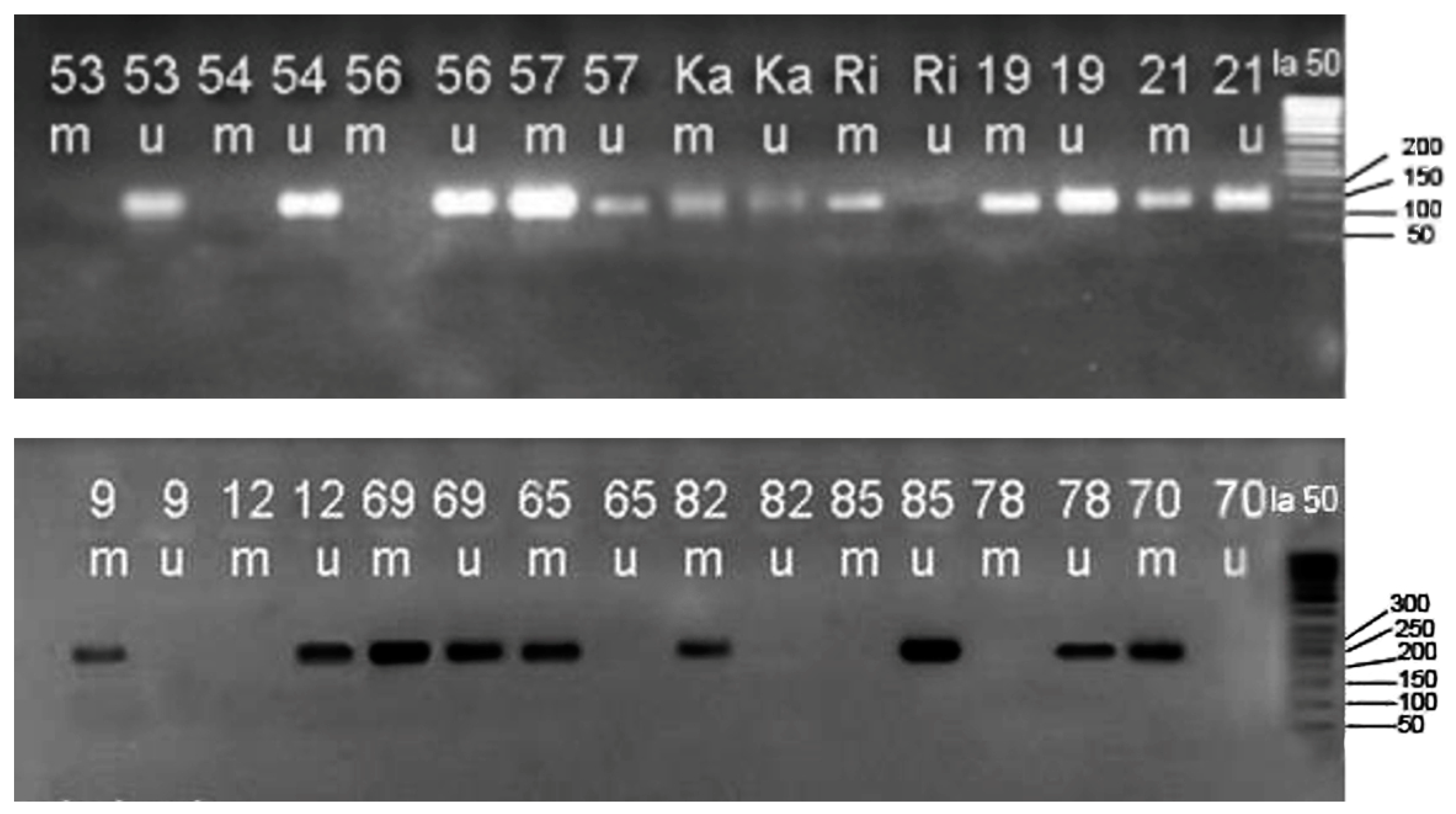
2.4. Methylation-Specific Polymerase Chain Reaction (MSP)
2.5. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Galanopoulos, M.; Tsoukalas, N.; Papanikolaou, I.S.; Tolia, M.; Gazouli, M.; Mantzaris, G.J. Abnormal DNA methylation as a cell-free circulating DNA biomarker for colorectal cancer detection: A review of literature. World J. Gastrointest. Oncol. 2017, 9, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Testa, U.; Pelosi, E.; Castelli, G. Colorectal Cancer: Genetic Abnormalities, Tumor Progression, Tumor Heterogeneity, Clonal Evolution and Tumor-Initiating Cells. Med. Sci. 2018, 6, 31. [Google Scholar] [CrossRef] [PubMed]
- Clark, S.K. Management of genetically determined colorectalcancer. Surgeon 2019, 17, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Mármol, I.; Sánchez-de-Diego, C.; Pradilla Dieste, A.; Cerrada, E.; Rodriguez Yoldi, M.J. Colorectal Carcinoma: A General Overview and Future Perspectives in Colorectal Cancer. Int. J. Mol. Sci. 2017, 18, 197. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.A.; Morris, M.; Idrees, K.; Gimbel, M.I.; Rosenberg, S.; Zeng, Z.; Li, F.; Gan, G.; Shia, J.; laquaglia, M.P.; et al. Colorectal Cancer in the Very Young: A Comparative Study of Tumor Markers, Pathology and Survival in Early Onset and Adult Onset Patients. J. Pediatr. Surg. 2016, 51, 1812–1817. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Li, Z.; Wei, H.; Hu, J.; Jiao, L.; Zhang, J.; Xiong, Y. Identification of gene-specific DNA methylation signature for Colorectal Cancer. Cancer Genet. 2018, 5–11, 228–229. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wu, J.; Meng, X.; Ying, X.; Zuo, Y.; Liu, R.; Pan, Z.; Kang, T.; Huang, W. MicroRNA-342 inhibits colorectal cancer cell proliferation and invasion by directly targeting DNA methyltransferase 1. Carcinogenesis 2011, 32, 1033–1042. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.-J.; Cao, J. MicroRNAs in colorectal cancer as markers and targets: Recent advances. World J. Gastroenterol. 2014, 20, 4288–4299. [Google Scholar] [CrossRef]
- Hur, K. MicroRNAs: Promising biomarkers for diagnosis and therapeutic targets in human colorectal cancer metastasis. BMB Rep. 2015, 48, 217–222. [Google Scholar] [CrossRef]
- Yang, Y.; Gu, X.; Zhou, M.; Xiang, J.; Chen, Z. Serum microRNAs: A new diagnostic method for colorectal cancer. Biomed. Rep. 2013, 1, 495–498. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.N.; Liu, T.T.; Wu, H.; Chen, Y.J.; Tseng, Y.J.; Yao, C.; Weng, S.Q.; Dong, L.; Shen, X.Z. Serum microRNA signatures and metabolomics have high diagnostic value in colorectal cancer using two novel methods. Cancer Sci. 2018, 109, 1185–1194. [Google Scholar] [CrossRef] [PubMed]
- Eslamizadeh, S.; Heidari, M.; Agah, S.; Faghihloo, E.; Ghazi, H.; Mirzaei, A.; Akbari, A. The role of MicroRNA signature as diagnostic biomarkers in different clinical stages of colorectal cancer. Cell J. 2018, 20, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Feinberg, A.P.; Vogelstein, B. Hypomethylation distinguishes genes of some human cancers from their normal counterparts. Nature 1983, 301, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, K.; Dai, T.; Dai, Y.; Yamamoto, F.; Perucho, M. Genetics supersedes epigenetics in colon cancer phenotype. Cancer Cell 2003, 4, 121–131. [Google Scholar] [CrossRef] [Green Version]
- Goel, A.; Boland, C.R. Epigenetics of colorectal cancer. Gastroenterology 2012, 143, 1442–1460.e1. [Google Scholar] [CrossRef]
- Luo, L.; Chen, W.D.; Pretlow, T.P. CpG island methylation in aberrant crypt foci and cancers from the same patients. Int. J. Cancer 2005, 115, 747–751. [Google Scholar] [CrossRef]
- Subramaniam, M.M.; Chan, J.Y.; Soong, R.; Ito, K.; Yeoh, K.G.; Wong, R.; Guenther, T.; Will, O.; Chen, C.L.; Kumarasinghe, M.P.; et al. RUNX3 inactivation in colorectal polyps arising through different pathways of colonic carcinogenesis. Am. J. Gastroenterol. 2009, 104, 426–436. [Google Scholar] [CrossRef]
- Wendt, M.K.; Johanesen, P.A.; Kang-Decker, N.; Binion, D.G.; Shah, V.; Dwinell, M.B. Silencing of epithelial CXCL12 expression by DNA hypermethylation promotes colonic carcinoma metastasis. Oncogene 2006, 25, 4986–4997. [Google Scholar] [CrossRef] [Green Version]
- Umetani, N.; Takeuchi, H.; Fujimoto, A.; Shinozaki, M.; Bilchik, A.J.; Hoon, D.S. Epigenetic inactivation of ID4 in colorectal carcinomas correlates with poor differentiation and unfavorable prognosis. Clin. Cancer Res. 2004, 10, 7475–7483. [Google Scholar] [CrossRef]
- Goel, A.; Nagasaka, T.; Arnold, C.N.; Inoue, T.; Hamilton, C.; Niedzwiecki, D.; Compton, C.; Mayer, R.J.; Goldberg, R.; Bertagnolli, M.M.; et al. The CpG island methylator phenotype and chromosomal instability are inversely correlated in sporadic colorectal cancer. Gastroenterology 2007, 132, 127–138. [Google Scholar] [CrossRef]
- Notterman, D.A.; Alon, U.; Sierk, A.J.; Levine, A.J. Transcriptional gene expression profiles of colorectal adenoma, adenocarcinoma, and normal tissue examined by oligonucleotide arrays. Cancer Res. 2001, 61, 3124–3130. [Google Scholar] [PubMed]
- Lyberopoulou, A.; Galanopoulos, M.; Aravantinos, G.; Theodoropoulos, G.E.; Marinos, E.; Efstathopoulos, E.P.; Gazouli, M. Identification of Methylation Profiles of Cancer-related Genes in Circulating Tumor Cells Population. Anticancer Res. 2017, 1105–1112. [Google Scholar] [CrossRef]
- Lima, C.R.; Gomes, C.C.; Santos, M.F. Role of microRNAs in endocrine cancer metastasis. Mol. Cell. Endocrinol. 2017, 456, 62–75. [Google Scholar] [CrossRef] [PubMed]
- Orang, A.V.; Barzegari, A. MicroRNAs in Colorectal Cancer: From Diagnosis to Targeted Therapy. Asian Pac. J. Cancer Prev. 2014, 15, 6989–6999. [Google Scholar] [CrossRef] [PubMed]
- To, K.K.; Tong, C.W.; Wu, M.; Cho, W.C. MicroRNAs in the prognosis and therapy of colorectal cancer: From bench to bedside. World J. Gastroenterol. 2018, 24, 2949–2973. [Google Scholar] [CrossRef] [PubMed]
- Slattery, M.L.; Lee, F.Y.; Pellatt, A.J.; Mullany, L.E.; Stevens, J.R.; Samowitz, W.S.; Wolff, R.K.; Herrick, J.S. Infrequently expressed miRNAs in colorectal cancer tissue and tumour molecular phenotype. Mod. Pathol. 2017, 30, 1152–1169. [Google Scholar] [CrossRef] [PubMed]
- Rajewsky, N. microRNA target predictions in animals. Nat. Genet. 2006, 38, S8–S13. [Google Scholar] [CrossRef] [PubMed]
- Ushijima, T.; Suzuki, H. The Origin of CIMP, at Last. Cancer Cell 2019, 35, 165–167. [Google Scholar] [CrossRef] [PubMed]
- Dang, J.; Bian, Y.Q.; Sun, J.Y.; Chen, F.; Dong, G.Y.; Liu, Q.; Wang, X.W.; Kjems, J.; Gao, S.; Wang, Q.T. MicroRNA-137 promoter methylation in oral lichen planus and oral squamous cell carcinoma. J. Oral Pathol. Med. 2013, 42, 315–321. [Google Scholar] [CrossRef]
- Chen, Q.; Chen, X.; Zhang, M.; Fan, Q.; Luo, S.; Cao, X. miR-137 is frequently down-regulated in gastric cancer and is a negative regulator of Cdc42. Dig. Dis. Sci. 2011, 56, 2009–2016. [Google Scholar] [CrossRef]
- Lim, B.; Kim, H.J.; Heo, H.; Huh, N.; Baek, S.J.; Kim, J.H.; Bae, D.H.; Seo, E.H.; Lee, S.L.; Song, K.S.; et al. Epigenetic silencing of miR-1271 enhances MEK1 and TEAD4 expression in gastric cancer. Cancer Med. 2018, 7, 3411–3424. [Google Scholar] [CrossRef] [PubMed]
- Yap, T.; Seers, C.; Koo, K.; Cheng, L.; Vella, L.J.; Hill, A.F.; Reynolds, E.; Nastri, A.; Cirillo, N.; McCullough, M. Non-invasive screening of a microRNA-based dysregulation signature in oral cancer and oral potentially malignant disorders. Oral Oncol. 2019, 96, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Bemis, L.T.; Chen, R.; Amato, C.M.; Classen, E.H.; Robinson, S.E.; Coffey, D.G.; Erickson, P.F.; Shellman, Y.G.; Robinson, W.A. MicroRNA-137 targets microphthalmia-associated transcription factor in melanoma cell lines. Cancer Res. 2008, 68, 1362–1368. [Google Scholar] [CrossRef] [PubMed]
- Wood, H. MicroRNA-132 master regulator of neuronal health? Nat. Rev. Neurol. 2018, 14, 508–509. [Google Scholar] [CrossRef] [PubMed]
- Takwi, A.A.; Wang, Y.M.; Wu, J.; Michaelis, M.; Cinatl, J.; Chen, T. miR-137 regulates the constitutive androstane receptor and modulates doxorubicin sensitivity in parental and doxorubicin-resistant neuroblastoma cells. Oncogene 2014, 33, 3717–3729. [Google Scholar] [CrossRef] [PubMed]
- Langevin, S.M.; Stone, R.A.; Bunker, C.H.; Lyons-Weiler, M.A.; LaFramboise, W.A.; Kelly, L.; Seethala, R.R.; Grandis, J.R.; Sobol, R.W.; Taioli, E. MicroRNA-137 promoter methylation is associated with poorer overall survival in patients with squamous cell carcinoma of the head and neck. Cancer 2011, 117, 1454–1462. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Kyung, H.M. Identification of eight meta-signature miRNAs as potential biomarkers for oropharyngeal cancers. Cancer Genet. 2019, 233–234, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Langevin, S.M.; Stone, R.A.; Bunker, C.H.; Grandis, J.R.; Sobol, R.W.; Taioli, E. MicroRNA-137 promoter methylation in oral rinses from patients with squamous cell carcinoma of the head and neck is associated with gender and body mass index. Carcinogenesis 2010, 31, 864–870. [Google Scholar] [CrossRef] [Green Version]
- Lim, Y.; Sun, C.X.; Tran, P.; Punyadeera, C. Salivary epigenetic biomarkers in head and neck squamous cell carcinomas. Biomark. Med. 2016, 10, 301–313. [Google Scholar] [CrossRef]
- Balaguer, F.; Link, A.; Lozano, J.J.; Cuatrecasas, M.; Nagasaka, T.; Boland, C.R.; Goel, A. Epigenetic silencing of miR-137 is an early event in colorectal carcinogenesis. Cancer Res. 2010, 70, 6609–6618. [Google Scholar] [CrossRef]
- Kaur, S.; Lotsari-Salomaa, J.E.; Seppänen-Kaijansinkko, R.; Peltomäki, P. MicroRNA Methylation in Colorectal Cancer. Adv. Exp. Med. Biol. 2016, 937, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Menigatti, M.; Staiano, T.; Manser, C.N.; Bauerfeind, P.; Komljenovic, A.; Robinson, M.; Jiricny, J.; Buffoli, F.; Marra, G. Epigenetic silencing of monoallelically methylated miRNA loci in precancerous colorectal lesions. Oncogenesis 2013, 2, e56. [Google Scholar] [CrossRef]
- Alonso, S.; González, B.; Alibés, A.; Perucho, M. Analysis of Somatic DNA Methylation Alterations of Genes Encoding Cell Surface Metallopeptidases in Colorectal Cancer. In Proteases and Cancer. Methods in Molecular Biology; Cal, S., Obaya, A., Eds.; Humana Press: New York, NY, USA, 2018; Volume 1731. [Google Scholar] [CrossRef]
- Huang, Z.; Huang, D.; Ni, S.; Peng, Z.; Sheng, W.; Du, X. Plasma microRNAs are promising novel biomarkers for early detection of colorectal cancer. Int. J. Cancer 2010, 127, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Kokuda, R.; Watanabe, R.; Okuzaki, D.; Akamatsu, H.; Oneyama, C. MicroRNA-137-mediated Src oncogenic signaling promotes cancer progression. Genes Cells 2018, 23, 8. [Google Scholar] [CrossRef] [PubMed]
- Oneyama, C.; Okada, M. MicroRNAs as the fine-tuners of Src oncogenic signalling. J. Biochem. 2015, 157, 431–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slezak-Prochazka, I.; Kluiver, J.; Jong, D.D.; Kortman, G.; Halsema, N.; Poppema, S.; Kroesen, B.-J.; van den Berg, A. Cellular localization and processing of primary transcripts of exonic microRNAs. PLoS ONE 2013, 8, e76647. [Google Scholar] [CrossRef] [PubMed]
- Louloupi, A.; Ntini, E.; Liz, J.; Ørom, U.A. Microprocessor dynamics shows co- and post-transcriptional processing of pri-miRNAs. RNA 2017, 23, 892–898. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.R.; Marquez, R.T.; Tsao, W.; Pathak, S.; Roy, A.; Ping, J.; Wilkerson, B.; Lan, L.; Meng, W.; Neufeld, K.L.; et al. Tumor suppressive microRNA-137 negatively regulates Musashi-1 and colorectal cancer progression. Oncotarget 2015, 6, 12558–12573. [Google Scholar] [CrossRef]
- Bellizzi, A.M.; Frankel, W.L. Colorectal cancer due to deficiency in DNA mismatch repair function: A review. Adv. Anat. Pathol. 2009, 16, 405–417. [Google Scholar] [CrossRef]
- Egawa, J.; Nunokawa, A.; Shibuya, M.; Watanabe, Y.; Kaneko, N.; Igeta, H.; Someya, T. Resequencing and association analysis of MIR137 with schizophrenia in a Japanese population. Psychiatry Clin. Neurosci. 2013, 67, 277–279. [Google Scholar] [CrossRef]
- Kuswanto, C.N.; Sum, M.Y.; Qiu, A.; Sitoh, Y.Y.; Liu, J.; Sim, K. The impact of genome wide supported microRNA-137 (MIR137) risk variants on frontal and striatal white matter integrity, neurocognitive functioning, and negative symptoms in schizophrenia. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2015, 168B, 317–326. [Google Scholar] [CrossRef]
- Fang, Z.; Tang, J.; Bai, Y.; Lin, H.; You, H.; Jin, H.; Lin, L.; You, P.; Li, L.; Dai, Z.; et al. Plasma levels of microRNA-24, microRNA-320a, and microRNA-423-5p are potential biomarkers for colorectal carcinoma. J. Exp. Clin. Cancer Res. 2015, 34, 86. [Google Scholar] [CrossRef]
- Chang, P.Y.; Chen, C.C.; Chang, Y.S.; Tsai, W.S.; You, J.F.; Lin, G.P.; Chen, T.W.; Chen, J.S.; Chan, E.C. MicroRNA-223 and microRNA-92a in stool and plasma samples act as complementary biomarkers to increase colorectal cancer detection. Oncotarget 2016, 7, 10663–10675. [Google Scholar] [CrossRef]
- Carter, J.V.; Galbraith, N.J.; Yang, D.; Burton, J.F.; Walker, S.P.; Galandiuk, S. Blood-based microRNAs as biomarkers for the diagnosis of colorectal cancer: A systematic review and meta-analysis. Br. J. Cancer 2017, 116, 762–774. [Google Scholar] [CrossRef]
- Kozaki, K.; Imoto, I.; Mogi, S.; Omura, K.; Inazawa, J. Exploration of tumor-suppressive microRNAs silenced by DNA hypermethylation in oral cancer. Cancer Res. 2008, 68, 2094–2105. [Google Scholar] [CrossRef]
- Endo, H.; Muramatsu, T.; Furuta, M.; Uzawa, N.; Pimkhaokham, A.; Amagasa, T.; Inazawa, J.; Kozaki, K. Potential of tumor-suppressive miR-596 targeting LGALS3BP as a therapeutic agent in oral cancer. Carcinogenesis 2013, 34, 560–569. [Google Scholar] [CrossRef]
- Tonouchi, E.; Muramatsu, T.; Furuta, M.; Uzawa, N.; Pimkhaokham, A.; Amagasa, T.; Inazawa, J.; Kozaki, K. miR-3140 suppresses tumor cell growth by targeting BRD4 via its coding sequence and downregulates the BRD4-NUT fusion oncoprotein. Sci. Rep. 2018, 8, 4482. [Google Scholar] [CrossRef] [Green Version]
- Bier, A.; Giladi, N.; Kronfeld, N.; Lee, H.K.; Cazacu, S.; Finniss, S.; Xiang, C.; Poisson, L.; de-Carvalho, A.C.; Slavin, S. MicroRNA-137 is downregulated in glioblastoma and inhibits the stemness of glioma stem cells by targeting RTVP-1. Oncotarget 2013, 4, 665–676. [Google Scholar] [CrossRef]
- Su, R.; Cao, S.; Ma, J.; Liu, Y.; Liu, X.; Zheng, J.; Chen, J.; Liu, L.; Cai, H.; Li, Z.; et al. Knockdown of SOX2OT inhibits the malignant biological behaviors of glioblastoma stem cells via up-regulating the expression of miR-194-5p and miR-122. Mol. Cancer 2017, 16, 171. [Google Scholar] [CrossRef]
- Liu, M.; Lang, N.; Qiu, M.; Xu, F.; Li, Q.; Tang, Q.; Chen, J.; Chen, X.; Zhang, S.; Liu, Z.; et al. miR-137 targets Cdc42 expression, induces cell cycle G1 arrest and inhibits invasion in colorectal cancer cells. Int. J. Cancer 2011, 128, 1269–1279. [Google Scholar] [CrossRef]
- Qadir, M.; Parveen, A.; Ali, M. Cdc42: Role in Cancer Management. Chem. Biol. Drug Des. 2015, 86, 432–439. [Google Scholar] [CrossRef]
- Garzon, R.; Liu, S.; Fabbri, M.; Liu, Z.; Heaphy, C.E.; Callegari, E.; Schwind, S.; Pang, J.; Yu, J.; Muthusamy, N.; et al. MicroRNA-29b induces global DNA hypomethylation and tumour suppressor gene re-expression in acute myeloid leukaemia by targeting directly DNMT3A and 3B and indirectly DNMT1. Blood 2009, 113, 6411–6418. [Google Scholar] [CrossRef]
- Chen, Y.L.; Zhang, Z.X.; Shou, L.H.; Di, J.Y. Regulation of DNA methylation and tumour suppression gene expression by miR-29b in leukaemia patients and related mechanisms. Eur. Rev. Med. Pharm. Sci. 2018, 22, 158–165. [Google Scholar] [CrossRef]
- Chen, D.L.; Wang, D.S.; Wu, W.J.; Zeng, Z.L.; Luo, H.Y.; Qiu, M.Z.; Ren, C.; Zhang, D.S.; Wang, Z.Q.; Wang, F.H.; et al. Overexpression of paxillin induced by miR-137 suppression promotes tumour progression and metastasis in colorectal cancer. Carcinogenesis 2013, 34, 803–811. [Google Scholar] [CrossRef]
- Luo, C.; Tetteh, P.W.; Merz, P.R.; Dickes, E.; Abukiwan, A.; Hotz-Wagenblatt, A.; Holland-Cunz, S.; Sinnberg, T.; Schittek, B.; Schadendorf, D.; et al. miR-137 inhibits the invasion of melanoma cells through downregulation of multiple oncogenic target genes. J. Investig. Dermatol. 2013, 133, 768–775. [Google Scholar] [CrossRef]
- Chen, X.; Liu, S.; Zhao, X.; Ma, X.; Gao, G.; Yu, L.; Yan, D.; Dong, H.; Sun, W. Long noncoding RNA ILF3-AS1 promotes cell proliferation, migration, and invasion via negatively regulating miR-200b/a/429 in melanoma. Biosci. Rep. 2017, 37. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, M.G.; Roldán, G.; Magliocco, A.; McIntyre, J.B.; Parney, I.; Easaw, J.C. Determination of the methylation status of MGMT in different regions within glioblastoma multiforme. J. Neuro-Oncol. 2011, 102, 255–260. [Google Scholar] [CrossRef]
- Binabaj, M.M.; Bahrami, A.; ShahidSales, S.; Joodi, M.; Joudi Mashhad, M.; Hassanian, S.M.; Anvari, K.; Avan, A. The prognostic value of MGMT promoter methylation in glioblastoma: A meta-analysis of clinical trials. J. Cell. Physiol. 2018, 233, 378–386. [Google Scholar] [CrossRef]
- Kang, N.; Choi, S.Y.; Kim, Y.K.; Yoo, I.F.; Han, D.H.; Lee, D.S.; Kim, Y.S.; Hong, S.H.; Kang, J.H.; Lee, K.Y.; et al. Silencing of miR-137 by aberrant promoter hypermethylation in surgically resected lung cancer. Lung Cancer 2015, 89, 99–103. [Google Scholar] [CrossRef]
- Singh, D.K.; Bose, S.; Kumar, S. Regulation of expression of microRNAs by DNA methylation in lung cancer. Biomarkers 2016, 21, 589–599. [Google Scholar] [CrossRef]
- Bandres, E.; Agirre, X.; Bitarte, N.; Ramirez, N.; Zarate, R.; Roman-Gomez, J.; Prosper, F.; Garcia-Foncillas, J. Epigenetic regulation of microRNA expression in colorectal cancer. Int. J. Cancer 2009, 125, 2737–2743. [Google Scholar] [CrossRef]
- Stevens, J.R.; Herrick, J.S.; Wolff, R.K.; Slattery, M.L. Identifying factors associated with the direction and significance of microRNA tumour-normal expression differences in colorectal cancer. BMC Cancer 2017, 17, 707. [Google Scholar] [CrossRef]
- Hamfjord, J.; Stangeland, A.M.; Hughes, T.; Skrede, M.L.; Tveit, K.M.; Ikdahl, T.; Kure, E.H. Differential expression of miRNAs in colorectal cancer: Comparison of paired tumor tissue and adjacent normal mucosa using high-throughput sequencing. PLoS ONE 2012, 7, e34150. [Google Scholar] [CrossRef]
- Grady, W.M.; Parkin, R.K.; Mitchell, P.S.; Lee, J.H.; Kim, Y.H.; Tsuchiya, K.D.; Washington, M.K.; Paraskeva, C.; Willson, J.K.; Kaz, A.M. Epigenetic silencing of the intronic microRNA hsa-miR-342 and its host gene EVL in colorectal cancer. Oncogene 2008, 27, 3880–3888. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Chuang, C.C.; Zuo, L. Potential roles of microRNAs and ROS in colorectal cancer: Diagnostic biomarkers and therapeutic targets. Oncotarget 2017, 8, 17328–17346. [Google Scholar] [CrossRef]
- Issa, J.; Vertino, P.M.; Boehm, C.D.; Newsham, I.F.; Baylin, S.B. Switch from monoallelic to biallelic human IGF2 promoter methylation during aging and carcinogenesis. Proc. Natl. Acad. Sci. USA 1996, 93, 11757–11762. [Google Scholar] [CrossRef]
- Le Stunff, C.; Castell, A.L.; Todd, N.; Mille, C.; Belot, M.P.; Frament, N.; Brailly-Tabard, S.; Benachi, A.; Fradin, D.; Bougnères, P. Fetal growth is associated with CpG methylation in the P2 promoter of the IGF1 gene. Clin. Epigenet. 2018, 10, 74. [Google Scholar] [CrossRef]
- Zheng, Q.H.; Ma, L.W.; Zhu, W.G.; Zhang, Z.Y.; Tong, T.J. p21Waf1/Cip1 plays a critical role in modulating senescence through changes of DNA methylation. J. Cell. Biochem. 2006, 98, 1230–1248. [Google Scholar] [CrossRef]
- Hassler, M.R.; Klisaroska, A.; Kollmann, K.; Steiner, I.; Bilban, M.; Schiefer, A.I.; Sexl, V.; Egger, G. Antineoplastic activity of the DNA methyltransferase inhibitor 5-aza-2’-deoxycytidine in anaplastic large cell lymphoma. Biochimie 2012, 94, 2297–2307. [Google Scholar] [CrossRef]
- Lanceta, J.; Rough, R.A.; Liang, R.; Wang, E. MicroRNA group disorganization in aging. Exp. Gerontol. 2010, 45, 269–278. [Google Scholar] [CrossRef]
- Kranjc, T.; Ostanek, B.; Marc, J. Bone microRNAs and Ageing. Curr. Pharm. Biotechnol. 2017, 18, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Ma, Y.; Wang, H.; Qin, H. Reciprocal regulation between microRNAs and epigenetic machinery in colorectal cancer. Oncol. Lett. 2017, 13, 1048–1057. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Liu, J.; Huo, T.; Tian, Y.; Zhao, L. The role of microRNAs in colorectal liver metastasis: Important participants and potential clinical significances. Tumour Biol. 2017, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Li, Q.; Niu, J.; Li, B.; Jiang, D.; Wan, Z.; Yang, Q.; Jiang, F.; Wei, P.; Bai, S. microRNA-342-5p and miR-608 inhibit colon cancer tumorigenesis by targeting NAA10. Oncotarget 2016, 19, 2709–2720. [Google Scholar] [CrossRef]
- Ramassone, A.; Pagotto, S.; Veronese, A.; Visone, R. Epigenetics and MicroRNAs in Cancer. Int. J. Mol. Sci. 2019, 19, 459. [Google Scholar] [CrossRef]
- Lawler, M.; Alsina, D.; Adams, R.A.; Anderson, S.A.; Brown, G.; Fearnhead, N.S.; Fenwick, S.W.; Halloran, S.P.; Hochhauser, D.; Hull, M.A.; et al. Critical research gaps and recommendations to inform research prioritisation for more effective prevention and improved outcomes in colorectal cancer. Gut 2018, 67, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Draht, M.X.G.; Goudkade, D.; Koch, A.; Grabsch, H.I.; Weijenberg, M.P.; van Engeland, M.; Melotte, V.; Smits, K.M. Prognostic DNA methylation markers for sporadic colorectal cancer: A systematic review. Clin. Epigenet. 2018, 10, 35. [Google Scholar] [CrossRef] [PubMed]
- Nikolouzakis, T.K.; Vassilopoulou, L.; Fragkiadaki, P.; Mariolis Sapsakos, T.; Papadakis, G.Z.; Spandidos, D.A.; Tsatsakis, A.M.; Tsiaoussis, J. Improving diagnosis, prognosis and prediction by using biomarkers in CRC patients. Oncol. Rep. 2018, 39, 2455–2472. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Final Concentration | Volume/Reaction (µL) | Reagent |
|---|---|---|
| Distilled Water | 5.45 | - |
| 10x Hot Start Buffer | 1.25 | 1x |
| 5X Q buffer | 2.5 | 1x |
| 25 mM MgCl2 | 0.25 | 1.5–5 mM |
| dNTP * (10 mM of each) | 0.25 | 200 μM of each dNTP |
| Primer Forward (M and U) | 0.75 | 0.1–0.5 μM |
| Primer Reverse (M and U) | 0.75 | 0.1–0.5 μM |
| Template | 1.2 | 1 μg/100 μl reaction |
| Hot Start Taq Polymerase | 0.1 | 2.5 units/reaction |
| Final Volume | 12.5 | - |
| Steps | Duration (sec.) for miR-137 | Temperature (°C) for miR-342 | Duration (sec.) for miR-342 | Temperature (°C) for miR-342 | |
|---|---|---|---|---|---|
| Primary denaturation | 15 | 95 | 15′ | 95 | |
| Cycles | Denaturation | 1 | 95 * | 30 | 92 ** |
| Annealing | 35 | 56 * | 45 | 64 ** | |
| Extension | 1 | 72 * | 30 | 72 ** | |
| Final Extension | 10 | 72 | 10 | 72 | |
| Annealing Temperature (°C) | %CG | Length | Sequence | Primer |
|---|---|---|---|---|
| 53.2 | 57.9 | 19 | 5′-TAGCGGTAGTAGCGGTAGC-3′ | miRNA-137-M* |
| (Forward) | ||||
| 52.4 | 47.6 | 21 | 5′-CTAATACTCTCCTCGACTACG-3 | miRNA-137-M* (Reverse) |
| 52.4 | 47.6 | 21 | 5′-TAGTGGTAGTAGTGGTAGTGG-3′ | miRNA-137-U* |
| (Forward) | ||||
| 51.1 | 40.9 | 22 | 5′-CTAATACTCTCCTCAACTACAC-3′ | miRNA-137-U* (Reverse) |
| 71.15–80.47 | 44 | 43 | 5′-GCGGTCCCAAAAGGGTCAGTTATTTTCGTTCGTTTCGTTTTTC-3′ | miRNA-342-M* (Forward) |
| 72.50–81.79 | 51 | 41 | 5′-GCGGTCCCAAAAGGGTCAGTAAATACGCGCGTTACTATTCG-3′ | MiRNA-342-M* (Reverse) |
| 59.51 58.99- | 35 | 45 | 5′-GCGGTCCCAAAAGGGTCAGTTATTTTTGTTTGTTTTGTTTTTTGT-3′ | miRNA-342-U* |
| (Forward) | ||||
| 69.92–79.44 | 42 | 45 | 5′-GCGGTCCCAAAAGGGTCAGTTAAAATACACACATTACTATTCACC-3′ | miRNA-342-U* |
| (Reverse) |
| Frequency (% and Number Out of 59) | Conditions | Clinical-Pathological Feature |
|---|---|---|
| Pathology report | Tubular adenomas | 54.2%—(32) |
| Hyperplastic polyps | 25.%—(15) | |
| Invasive adenocarcinoma | 11.9%—(7) | |
| Villous, juvenile and ulcerative lesions | 5.1%—(3) | |
| Other serrated types | 3.4%—(3) | |
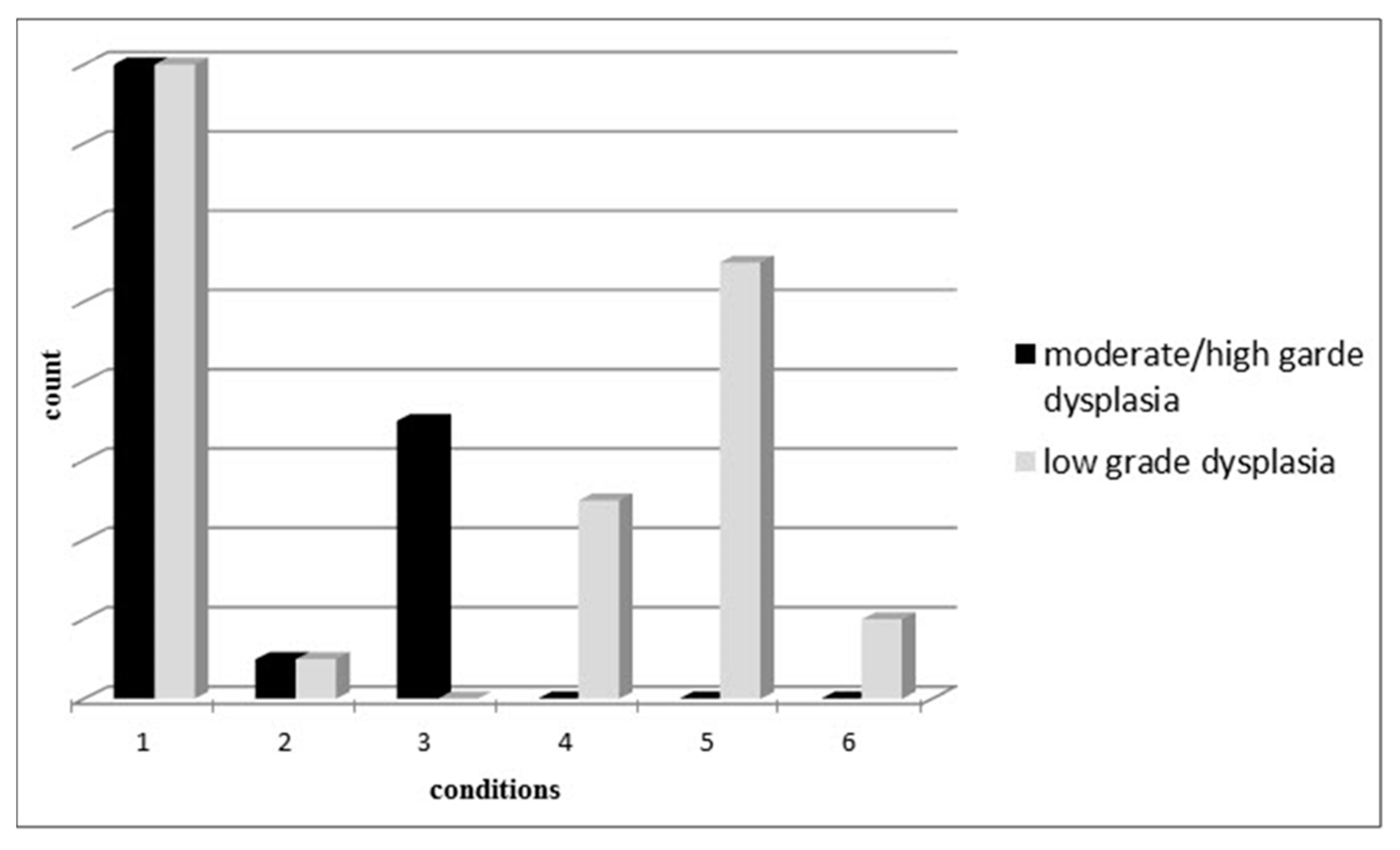
| Dysplasia degree | Low-grade dysplasia | 59.3%—(3–5) |
| High and moderate dysplasia | 40.7%—(24) | |
| Anatomic site and location in colon | Rectum and sigmoid | 40.7%—(24) |
| Descending colon | 22%—(13) | |
| Transverse colon and hepatic flexure | 16.9%—(10) | |
| Ascending colon | 11.9%—(7) | |
| Secom | 8.5%—(5) | |
| Sex | Male | 49.2% (29) |
| Female | 50.8% (30) | |
| Age | 17–45 yrs. | 25.4% (15) |
| 45–65 yrs. | 45.8% (27) | |
| More than 65 yrs. | 28.8% (17) | |
| Body mass index (BMI) | Normal (18.5–25) | 27.1% (16) |
| Overweight (>25) | 71.2% (42) | |
| Underweight (<25) | 1.7% (1) | |
| Family history | Positive | 28.8% (17) |
| Negative | 71.2% (42) | |
| Diabetes | Positive | 10.2% (6) |
| Negative | 89.8% (53) | |
| Hypertension | Positive | 16.9% (10) |
| Negative | 83.1% (49) | |
| Nonsteroidal anti-inflammatory drugs | Positive | 20.3% (12) |
| (NSAIDs) intake | Negative | 79.7% (47) |
| Smoking | Positive | 23.7% (14) |
| Negative | 76.3% (45) | |
| Alcohol consumption | Positive | 15.3% (9) |
| Negative | 84.7% (50) | |
| Addiction | Positive | 3.4% (2) |
| Negative | 96.6% (57) | |
| Regular exercise | Positive | 54.2% (32) |
| Negative | 45.8% (27) | |
| Fruit-vegetable intake | Considerate | 50.8% (30) |
| Inappropriate | 49.2% (29) | |
| Red meat intake | Considerate | 37.3% (22) |
| Inappropriate | 62.7% (37) |
| Pathology Report | miR-137 Methylation Status | miR-342 Methylation Status | ||
|---|---|---|---|---|
| Methylation Frequencies | Unmethylation Frequencies | Methylation Frequencies | Unmethylation Frequencies | |
| Tubular adenoma polyps | 15.6% | 84.4% | 81.3% | 18.8% |
| Villous adenoma polyps | 100% | 0% | 50% | 50% |
| Invasive adenocarcinomas polyp/masses | 28.6% | 71.4% | 75.7% | 14.3% |
| Ulcerated, juvenile and retention polyps | 20% | 80% | 20% | 80% |
| Hyperplastic polyps | 9.1% | 90.9% | 81.8% | 18.2% |
| Other serrated polyps | 0% | 100% | 100% | 0% |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kashani, E.; Hadizadeh, M.; Chaleshi, V.; Mirfakhraie, R.; Young, C.; Savabkar, S.; Irani, S.; Asadzadeh Aghdaei, H.; Ashrafian Bonab, M. The Differential DNA Hypermethylation Patterns of microRNA-137 and microRNA-342 Locus in Early Colorectal Lesions and Tumours. Biomolecules 2019, 9, 519. https://doi.org/10.3390/biom9100519
Kashani E, Hadizadeh M, Chaleshi V, Mirfakhraie R, Young C, Savabkar S, Irani S, Asadzadeh Aghdaei H, Ashrafian Bonab M. The Differential DNA Hypermethylation Patterns of microRNA-137 and microRNA-342 Locus in Early Colorectal Lesions and Tumours. Biomolecules. 2019; 9(10):519. https://doi.org/10.3390/biom9100519
Chicago/Turabian StyleKashani, Elham, Mahrooyeh Hadizadeh, Vahid Chaleshi, Reza Mirfakhraie, Chris Young, Sanaz Savabkar, Shiva Irani, Hamid Asadzadeh Aghdaei, and Maziar Ashrafian Bonab. 2019. "The Differential DNA Hypermethylation Patterns of microRNA-137 and microRNA-342 Locus in Early Colorectal Lesions and Tumours" Biomolecules 9, no. 10: 519. https://doi.org/10.3390/biom9100519