Association between Resistin Gene Polymorphisms and Atopic Dermatitis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Sample Collection and Handling
2.3. Genetic Analysis
DNA Extraction
2.4. Molecular Analysis
2.4.1. PCR Procedure
2.4.2. Restriction Fragment Length Polymorphism
2.5. Resistin Serum Level
2.6. Statistical Analysis
3. Results
3.1. Characteristics of the Participants
3.2. Association between Resistin Gene Polymorphisms and Atopic Dermatitis
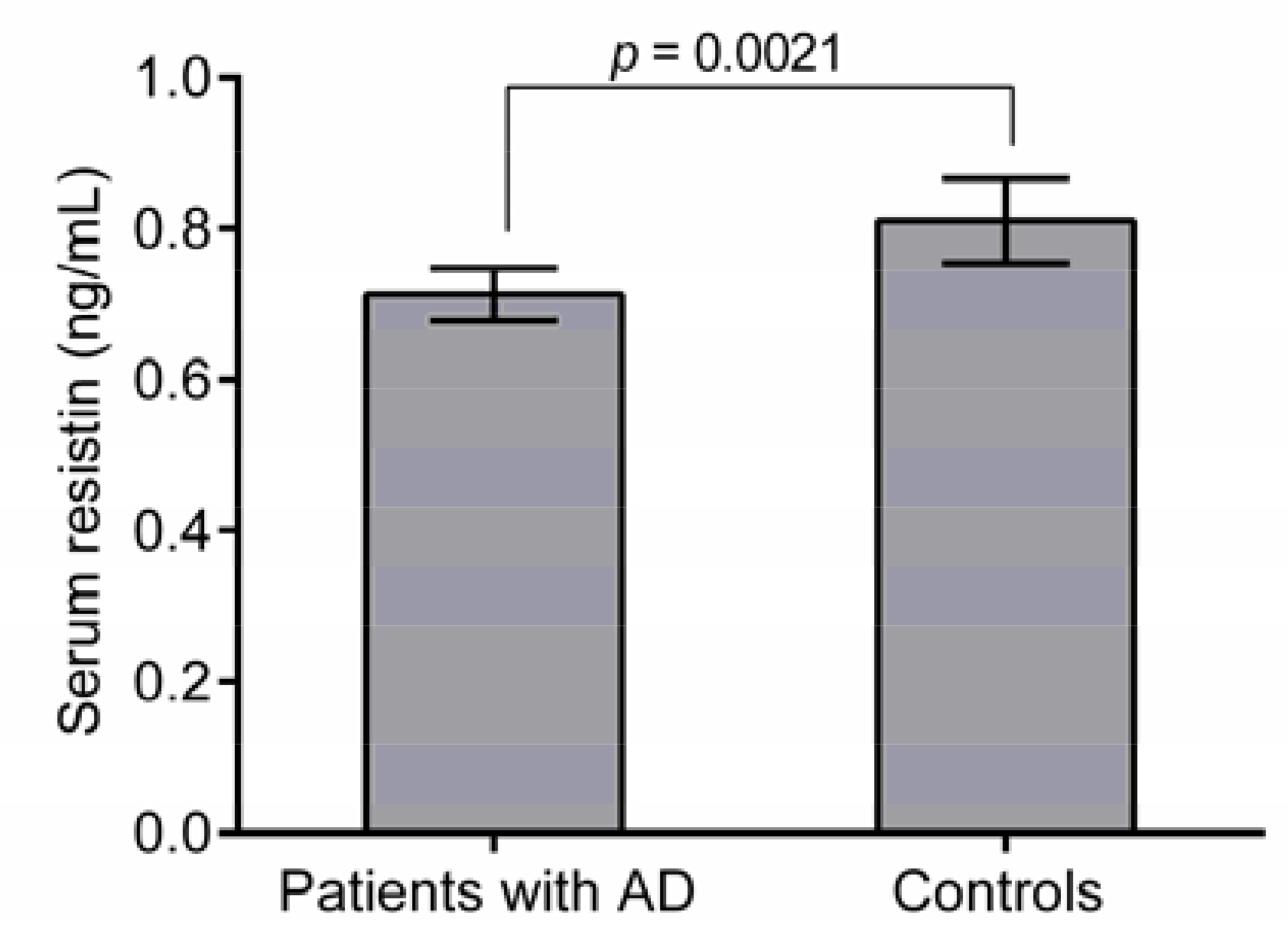
3.3. Association between Resistin Protein Level and AD
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Odhiambo, J.A.; Williams, H.C.; Clayton, T.O.; Robertson, C.F.; Asher, M.I.; Group, I.P.T.S. Global variations in prevalence of eczema symptoms in children from ISAAC phase three. J. Allergy Clin. Immunol. 2009, 124, 1251–1258 e1223. [Google Scholar] [CrossRef] [PubMed]
- Williams, H.; Robertson, C.; Stewart, A.; Ait-Khaled, N.; Anabwani, G.; Anderson, R.; Asher, I.; Beasley, R.; Bjorksten, B.; Burr, M.; et al. Worldwide variations in the prevalence of symptoms of atopic eczema in the international study of asthma and allergies in childhood. J. Allergy Clin. Immunol. 1999, 103, 125–138. [Google Scholar] [CrossRef]
- Williams, H.C. Is the prevalence of atopic dermatitis increasing? Clin. Exp. Dermatol. 1992, 17, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Asher, M.I.; Montefort, S.; Bjorksten, B.; Lai, C.K.; Strachan, D.P.; Weiland, S.K.; Williams, H.; Group, I.P.T.S. Worldwide time trends in the prevalence of symptoms of asthma, allergic rhinoconjunctivitis, and eczema in childhood: ISAAC phases one and three repeat multicountry cross-sectional surveys. Lancet 2006, 368, 733–743. [Google Scholar] [CrossRef]
- Eichenfield, L.F.; Hanifin, J.M.; Beck, L.A.; Lemanske, R.F., Jr.; Sampson, H.A.; Weiss, S.T.; Leung, D.Y. Atopic dermatitis and asthma: Parallels in the evolution of treatment. Pediatrics 2003, 111, 608–616. [Google Scholar] [CrossRef] [PubMed]
- Von Mutius, E. Gene-environment interactions in asthma. J. Allergy Clin. Immunol. 2009, 123, 3–11; quiz 12–13. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Maenhaut, N.; Van de Voorde, J. Regulation of vascular tone by adipocytes. BMC Med. 2011, 9, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Q.; Tan, H.; Irwin, D.M. Evolution of the vertebrate resistin gene family. PLoS One 2015, 10, e0130188. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Lee, K.; Moon, Y.S.; Sul, H.S. A cysteine-rich adipose tissue-specific secretory factor inhibits adipocyte differentiation. J. Biol. Chem. 2001, 276, 11252–11256. [Google Scholar] [CrossRef] [PubMed]
- Steppan, C.M.; Bailey, S.T.; Bhat, S.; Brown, E.J.; Banerjee, R.R.; Wright, C.M.; Patel, H.R.; Ahima, R.S.; Lazar, M.A. The hormone resistin links obesity to diabetes. Nature 2001, 409, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Patel, L.; Buckels, A.C.; Kinghorn, I.J.; Murdock, P.R.; Holbrook, J.D.; Plumpton, C.; Macphee, C.H.; Smith, S.A. Resistin is expressed in human macrophages and directly regulated by PPAR gamma activators. Biochem. Biophys. Res. Commun. 2003, 300, 472–476. [Google Scholar] [CrossRef]
- Lehrke, M.; Reilly, M.P.; Millington, S.C.; Iqbal, N.; Rader, D.J.; Lazar, M.A. An inflammatory cascade leading to hyperresistinemia in humans. PLoS Med. 2004, 1, e45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donath, M.Y.; Shoelson, S.E. Type 2 diabetes as an inflammatory disease. Nat. Rev. Immunol. 2011, 11, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Abate, N.; Sallam, H.S.; Rizzo, M.; Nikolic, D.; Obradovic, M.; Bjelogrlic, P.; Isenovic, E.R. Resistin: An inflammatory cytokine. Role in cardiovascular diseases, diabetes and the metabolic syndrome. Curr. Pharm. Des. 2014, 20, 4961–4969. [Google Scholar] [CrossRef] [PubMed]
- Nagel, G.; Koenig, W.; Rapp, K.; Wabitsch, M.; Zoellner, I.; Weiland, S.K. Associations of adipokines with asthma, rhinoconjunctivitis, and eczema in German schoolchildren. Pediatr. Allergy Immunol. 2009, 20, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.N.; Irvine, A.D.; Terron-Kwiatkowski, A.; Zhao, Y.; Liao, H.; Lee, S.P.; Goudie, D.R.; Sandilands, A.; Campbell, L.E.; Smith, F.J.; et al. Common loss-of-function variants of the epidermal barrier protein filaggrin are a major predisposing factor for atopic dermatitis. Nat. Genet. 2006, 38, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.C.; Shieh, W.Y.; Chen, C.Y.; Hsu, S.C.; Chen, H.L. Lipopolysaccharide increases resistin gene expression in vivo and in vitro. FEBS Lett. 2002, 530, 158–162. [Google Scholar] [CrossRef]
- Younis, S.; Blumenberg, M.; Javed, Q. Resistin gene polymorphisms are associated with acne and serum lipid levels, providing a potential nexus between lipid metabolism and inflammation. Arch. Dermatol. Res. 2016, 308, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Faraz, A.; Iqbal, T. The RETN gene rs1862513 polymorphism as a novel predisposing marker for familial acne vulgaris in a pakistani population. Iran J. Basic Med. Sci. 2015, 18, 526–528. [Google Scholar] [PubMed]
- Silha, J.V.; Krsek, M.; Skrha, J.V.; Sucharda, P.; Nyomba, B.L.; Murphy, L.J. Plasma resistin, adiponectin and leptin levels in lean and obese subjects: Correlations with insulin resistance. Eur. J. Endocrinol. 2003, 149, 331–335. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Silverberg, J.I. Association of atopic dermatitis with being overweight and obese: A systematic review and metaanalysis. J. Am. Acad. Dermatol. 2015, 72, 606–616 e604. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Han, K.D.; Jung, H.M.; Youn, Y.H.; Lee, J.Y.; Park, Y.G.; Lee, S.H.; Park, Y.M. Association between obesity, abdominal obesity, and adiposity and the prevalence of atopic dermatitis in young Korean adults: The korea national health and nutrition examination survey 2008-2010. Allergy Asthma Immunol. Res. 2016, 8, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Machura, E.; Szczepanska, M.; Ziora, K.; Ziora, D.; Swietochowska, E.; Barc-Czarnecka, M.; Kasperska-Zajac, A. Evaluation of adipokines: Apelin, visfatin, and resistin in children with atopic dermatitis. Mediators Inflamm. 2013, 2013, 760691. [Google Scholar] [CrossRef] [PubMed]
- Andersen, Y.M.F.; Egeberg, A.; Gislason, G.H.; Skov, L.; Knop, F.K.; Thyssen, J.P. Adult atopic dermatitis and the risk of type 2 diabetes. J. Allergy Clin. Immunol. 2017, 139, 1057–1059. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, D.R.; Lazar, M.A. Human resistin: Found in translation from mouse to man. Trends Endocrinol. Metab. 2011, 22, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.; White, S.P.; Ling, C.; Zhou, W. Resistin and cardiovascular disease. Trends Cardiovasc. Med. 2011, 21, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.E.; Kim, H.S. Human resistin in cardiovascular disease. J. Smooth Muscle Res. 2012, 48, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Nieva-Vazquez, A.; Perez-Fuentes, R.; Torres-Rasgado, E.; Lopez-Lopez, J.G.; Romero, J.R. Serum resistin levels are associated with adiposity and insulin sensitivity in obese hispanic subjects. Metab. Syndr. Relat. Disord. 2014, 12, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Menzaghi, C.; Bacci, S.; Salvemini, L.; Mendonca, C.; Palladino, G.; Fontana, A.; De Bonis, C.; Marucci, A.; Goheen, E.; Prudente, S.; et al. Serum resistin, cardiovascular disease and all-cause mortality in patients with type 2 diabetes. PLoS ONE 2014, 8, e64729. [Google Scholar]
- Mathew, H.; Castracane, V.D.; Mantzoros, C. Adipose tissue and reproductive health. Metabolism 2017. [Google Scholar] [CrossRef] [PubMed]
- Nagaev, I.; Smith, U. Insulin resistance and type 2 diabetes are not related to resistin expression in human fat cells or skeletal muscle. Biochem. Biophys. Res. Commun. 2001, 285, 561–564. [Google Scholar] [CrossRef] [PubMed]
- Kaser, S.; Kaser, A.; Sandhofer, A.; Ebenbichler, C.F.; Tilg, H.; Patsch, J.R. Resistin messenger-RNA expression is increased by proinflammatory cytokines in vitro. Biochem. Biophys. Res. Commun. 2003, 309, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Harrison, W.J.; Bull, J.J.; Seltmann, H.; Zouboulis, C.C.; Philpott, M.P. Expression of lipogenic factors galectin-12, resistin, SREBP-1, and SCD in human sebaceous glands and cultured sebocytes. J. Invest. Dermatol. 2007, 127, 1309–1317. [Google Scholar] [CrossRef] [PubMed]
- Khabour, O.F.; Mesmar, F.S.; Alatoum, M.A.; Gharaibeh, M.Y.; Alzoubi, K.H. Associations of polymorphisms in adiponectin and leptin genes with men's longevity. Aging Male 2010, 13, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Cherneva, R.V.; Georgiev, O.B.; Petrova, D.S.; Mondeshki, T.L.; Ruseva, S.R.; Cakova, A.D.; Mitev, V.I. Resistin—The link between adipose tissue dysfunction and insulin resistance in patients with obstructive sleep apnea. J. Diabetes Metab. Disord. 2013, 12, 5. [Google Scholar] [CrossRef] [PubMed]
- May, R.C. Gender, immunity and the regulation of longevity. Bioessays 2007, 29, 795–802. [Google Scholar] [CrossRef] [PubMed]
- Ukkola, O.; Kunnari, A.; Kesaniemi, Y.A. Genetic variants at the resistin locus are associated with the plasma resistin concentration and cardiovascular risk factors. Regul. Pept. 2008, 149, 56–59. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.W.; Shin, Y.H.; Lee, K.E.; Kim, E.S.; Sohn, M.H.; Kim, K.E. Relationship between adipokines and manifestations of childhood asthma. Pediatr. Allergy Immunol. 2008, 19, 535–540. [Google Scholar] [CrossRef] [PubMed]
- Pehlivan, Y.; Onat, A.M.; Ceylan, N.; Turkbeyler, I.H.; Buyukhatipoglu, H.; Comez, G.; Babacan, T.; Tarakcioglu, M. Serum leptin, resistin and TNF-alpha levels in patients with systemic sclerosis: The role of adipokines in scleroderma. Int. J. Rheum. Dis. 2012, 15, 374–379. [Google Scholar] [CrossRef] [PubMed]
- Robati, R.M.; Partovi-Kia, M.; Haghighatkhah, H.R.; Younespour, S.; Abdollahimajd, F. Increased serum leptin and resistin levels and increased carotid intima-media wall thickness in patients with psoriasis: Is psoriasis associated with atherosclerosis? J. Am. Acad. Dermatol. 2014, 71, 642–648. [Google Scholar] [CrossRef] [PubMed]
- Machura, E.; Szczepanska, M.; Swietochowska, E.; Halkiewicz, F.; Barc-Czarnecka, M.; Ziora, K.; Ziora, D. Evaluation of adipokines in children with cystic fibrosis. Endokrynol. Pol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Nosalski, R.; Guzik, T.J. Perivascular adipose tissue inflammation in vascular disease. Br. J. Pharmacol. 2017, 174, 3496–3513. [Google Scholar] [CrossRef] [PubMed]
- Grygiel-Gorniak, B.; Kaczmarek, E.; Mosor, M.; Przyslawski, J.; Bogacz, A. Genetic background, adipocytokines, and metabolic disorders in postmenopausal overweight and obese women. Biochem. Genet. 2016, 54, 636–652. [Google Scholar] [CrossRef] [PubMed]
- Khabour, O.F.; Abu-Rumeh, L.; Al-Jarrah, M.; Jamous, M.; Alhashimi, F. Association of adiponectin protein and adipoq gene variants with lumbar disc degeneration. Exp. Ther. Med. 2014, 8, 1340–1344. [Google Scholar] [CrossRef] [PubMed]
- Tay, Y.K.; Kong, K.H.; Khoo, L.; Goh, C.L.; Giam, Y.C. The prevalence and descriptive epidemiology of atopic dermatitis in Singapore school children. Br. J. Dermatol. 2002, 146, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.R.; Burr, M.L.; Williams, H.C. Factors influencing atopic dermatitis-a questionnaire survey of schoolchildren's perceptions. Br. J. Dermatol. 2004, 150, 1154–1161. [Google Scholar] [CrossRef] [PubMed]
- Amorim, M.G.S.; de Oliveira, M.D.; Soares, D.S.; da Silva Borges, L.; Dermargos, A.; Hatanaka, E. Effects of exergaming on cardiovascular risk factors and adipokine levels in women. J. Physiol. Sci. 2017. [Google Scholar] [CrossRef] [PubMed]
- Shirvani, H.; Arabzadeh, E. Metabolic cross-talk between skeletal muscle and adipose tissue in high-intensity interval training vs. Moderate-intensity continuous training by regulation of PGC-1alpha. Eat Weight Disord. 2018. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Qi, Y.; Huang, F.; Chen, X.; Zhou, Y.; Ye, L.; Wang, W.; Ning, G.; Wang, S. Down-regulated resistin level in consequence of decreased neutrophil counts in untreated grave's disease. Oncotarget. 2016, 7, 78680–78687. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Han, K.; Lee, J.H.; Kim, N.I.; Roh, J.Y.; Seo, S.J.; Song, H.J.; Lee, M.G.; Choi, J.H.; Park, Y.M. Increased risk of chronic spontaneous urticaria in patients with autoimmune thyroid diseases: A nationwide, population-based study. Allergy Asthma Immunol. Res. 2017, 9, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Fabene, P.F.; Bramanti, P.; Constantin, G. The emerging role for chemokines in epilepsy. J. Neuroimmunol. 2010, 224, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Gautam, S.C.; Chopp, M.; Zaloga, C.; Jones, M.L.; Ward, P.A.; Welch, K.M. Expression of monocyte chemoattractant protein-1 and macrophage inflammatory protein-1 after focal cerebral ischemia in the rat. J. Neuroimmunol. 1995, 56, 127–134. [Google Scholar] [CrossRef]
- Hickman, S.E.; El Khoury, J. Mechanisms of mononuclear phagocyte recruitment in Alzheimer's disease. CNS Neurol. Disord. Drug Targets 2010, 9, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Ransohoff, R.M.; Hamilton, T.A.; Tani, M.; Stoler, M.H.; Shick, H.E.; Major, J.A.; Estes, M.L.; Thomas, D.M.; Tuohy, V.K. Astrocyte expression of mRNA encoding cytokines IP-10 and JE/MCP-1 in experimental autoimmune encephalomyelitis. FASEB J. 1993, 7, 592–600. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| SNP ID | Primer Sequence (5’–3’) | PCR Annealing T (°C) | Restriction Enzyme, Incubation Conditions | Fragment Length (bp) |
|---|---|---|---|---|
| rs3745367 | F: GGAAGAAGCCATCAATGAGAGG R: CCTGTTGGTTTGGAGCTAGGTC | 58 | Alu I, 37 °C overnight | GG➔ 14, 71, 243 GA➔ 14, 71, 85, 158, 243 AA➔ 14, 71, 85, 158 |
| rs3219177 | F: AGTGACAGCTGCTCCTGCG R: ATGAGATTTGGTGAGCGCT | 58 | BspCN I, 37 °C overnight | CC➔ 48, 376, 351 CT➔ 48, 54, 322, 376, 424 TT➔ 48, 54, 322, 376 |
| Variable | Control n (%) | Patient n (%) | p-Value |
|---|---|---|---|
| Gender | |||
| Male | 82 (50.9%) | 82 (50.6%) | 0.955 |
| Female | 79 (49.1%) | 80 (49.4%) | |
| Total | 161 | 162 | |
| Age groups | |||
| New Born–10 years | 94 (58.4%) | 92 (56.8%) | 0.885 |
| 11–20 years | 36 (22.4%) | 40 (24.7%) | |
| >20 | 31 (19.2%) | 30 (18.5%) | |
| Genotypes and Alleles | Control n (%) | Patient n (%) | p-Value |
|---|---|---|---|
| rs3219177 | |||
| CC | 113 (70.2%) | 107 (66.0%) | 0.219 NS |
| TT | 4 (2.5%) | 1 (0.6%) | |
| CT | 44 (27.3%) | 54 (33.4%) | |
| Allele C | 157 (76.6%) | 161 (74.5%) | 0.625 NS |
| Allele T | 48 (23.4%) | 55 (25.5%) | |
| rs3745367 | |||
| GG | 52 (32.3%) | 76 (46.9%) | 0.023 |
| AA | 32 (19.9%) | 22 (13.6%) | |
| GA | 77 (47.8%) | 64 (39.5%) | |
| Allele G | 129 (54.2%) | 140 (61.9%) | 0.091 |
| Allele A | 109 (45.8%) | 86 (38.1%) | |
| Genotypes and Alleles | Control n (%) | Patient n (%) | p-Value |
|---|---|---|---|
| rs3219177 | |||
| CC | 57 (69.5%) | 54 (65.1%) | 0.255 |
| TT | 4 (4.9%) | 1 (1.2%) | |
| CT | 21 (25.6%) | 28 (33.7%) | |
| Allele C | 78 (75.7%) | 82 (73.9%) | 0.755 |
| Allele T | 25 (24.3%) | 29 (26.1%) | |
| rs3745367 | |||
| GG | 26 (31.7%) | 44 (53%) | 0.007 |
| AA | 14 (17.1%) | 5 (6%) | |
| GA | 42 (51.2%) | 34 (41%) | |
| Allele G | 40 (54.8%) | 78 (66.7%) | 0.0003 |
| Allele A | 56 (45.2%) | 39 (33.3%) | |
| Genotypes and Alleles | Control n (%) | Patient n (%) | p-Value |
|---|---|---|---|
| rs3219177 | |||
| CC | 56 (70.9%) | 53 (67.1%) | 0.731 |
| TT | 0 (0%) | 0 (0%) | |
| CT | 23 (29.1%) | 26 (32.9%) | |
| Allele C | 79 (77.5%) | 79 (75.2%) | 0.708 |
| Allele T | 23 (22.5%) | 26 (24.8%) | |
| rs3745367 | |||
| GG | 27 (34.2%) | 30 (38%) | 0.82 |
| AA | 17 (21.5%) | 17 (21.5%) | |
| GA | 35 (44.3%) | 32 (40.5%) | |
| Allele G | 62 (54.4%) | 62 (55.9%) | 0.824 |
| Allele A | 52 (45.6%) | 49 (44.1%) | |
| Age Groups | ||||||
|---|---|---|---|---|---|---|
| Genotypes and Alleles | New Born–10 years | 11–20 years | >20 years | |||
| rs3219177 | Control | Patient | Control | Patient | Control | Patient |
| CC | 64 (68.1%) | 58 (63%) | 27 (75%) | 27 (67.5%) | 22 (71%) | 22(73.3%) |
| TT | 2 (2.1%) | 0 (0%) | 1 (2.8%) | 1 (2.5%) | 1 (3.2%) | 0 (0%) |
| CT | 28 (29.8%) | 34 (37%) | 8 (22.2%) | 12 (30%) | 8 (25.8%) | 8 (26.7%) |
| p-value | 0.24 | 0.798 | 1.00 | |||
| Allele C | 92 (75.4%) | 92 (73%) | 35 (79.5%) | 39 (75%) | 30 (77%) | 30(78.9%) |
| Allele T | 30 (24.6%) | 34 (27%) | 9 (20.5%) | 13 (25%) | 9 (23%) | 8 (21.1%) |
| p-value | 0.667 | 0.598 | 0.83 | |||
| rs3745367 | ||||||
| GG | 30 (31.9%) | 47 (51.1%) | 11 (30.6%) | 16 (40%) | 11 (35.5%) | 13(43.3%) |
| AA | 19 (20.2%) | 12 (13%) | 9 (25%) | 4 (10%) | 4 (12.9%) | 6 (20%) |
| GA | 45 (47.9%) | 33 (35.9%) | 16 (44.4%) | 20 (50%) | 16 (51.6) | 11(36.7%) |
| p-value | 0.028 | 0.243 | 0.516 | |||
| Allele G | 75 (54%) | 80 (64%) | 27 (51.9%) | 36 (60%) | 27 (57.4%) | 24(58.5%) |
| Allele A | 64 (46%) | 45 (36%) | 25 (48.1%) | 24 (40%) | 20 (42.6%) | 17(41.5%) |
| p-value | 0.098 | 0.39 | 0.918 | |||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Banihani, S.A.; Abu-Alia, K.F.; Khabour, O.F.; Alzoubi, K.H. Association between Resistin Gene Polymorphisms and Atopic Dermatitis. Biomolecules 2018, 8, 17. https://doi.org/10.3390/biom8020017
Banihani SA, Abu-Alia KF, Khabour OF, Alzoubi KH. Association between Resistin Gene Polymorphisms and Atopic Dermatitis. Biomolecules. 2018; 8(2):17. https://doi.org/10.3390/biom8020017
Chicago/Turabian StyleBanihani, Saleem A., Khawla F. Abu-Alia, Omar F. Khabour, and Karem H. Alzoubi. 2018. "Association between Resistin Gene Polymorphisms and Atopic Dermatitis" Biomolecules 8, no. 2: 17. https://doi.org/10.3390/biom8020017