
Mutations in Cancer Cause Gain of Cysteine, Histidine, and Tryptophan at the Expense of a Net Loss of Arginine on the Proteome Level


Abstract
:1. Introduction
2. Results
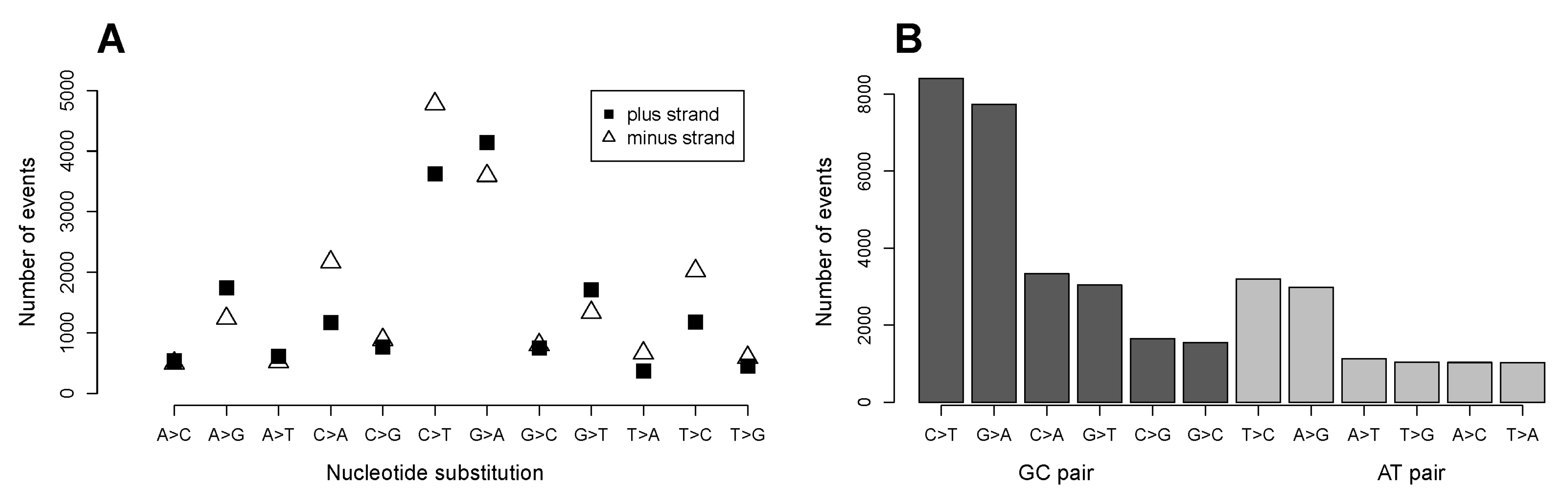
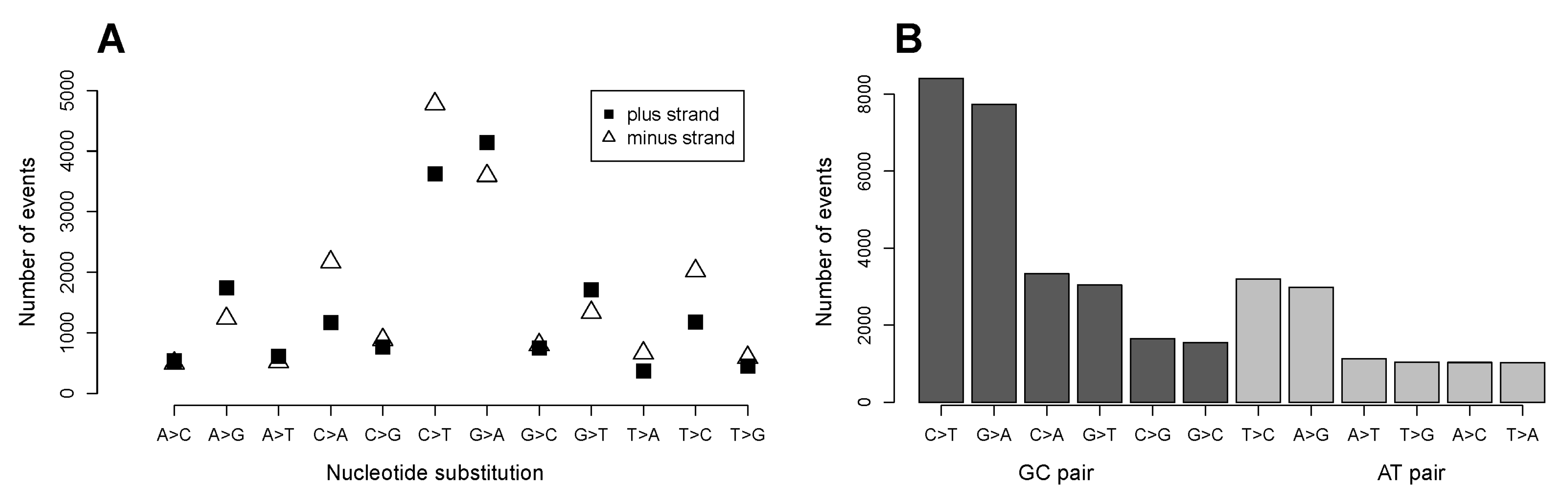
2.1. C>T and G>A Changes Are the Most Numerous Nucleotide Transformations
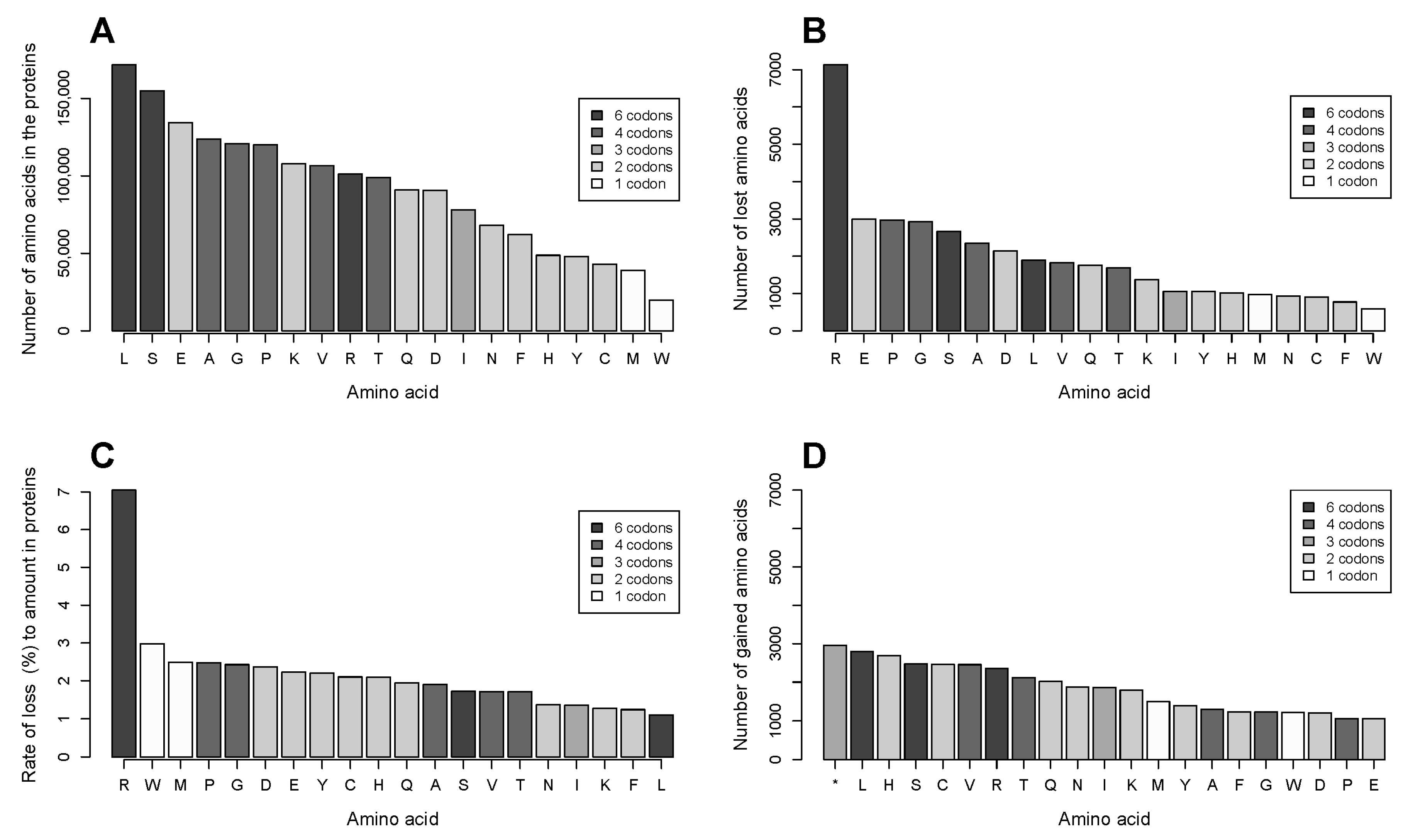
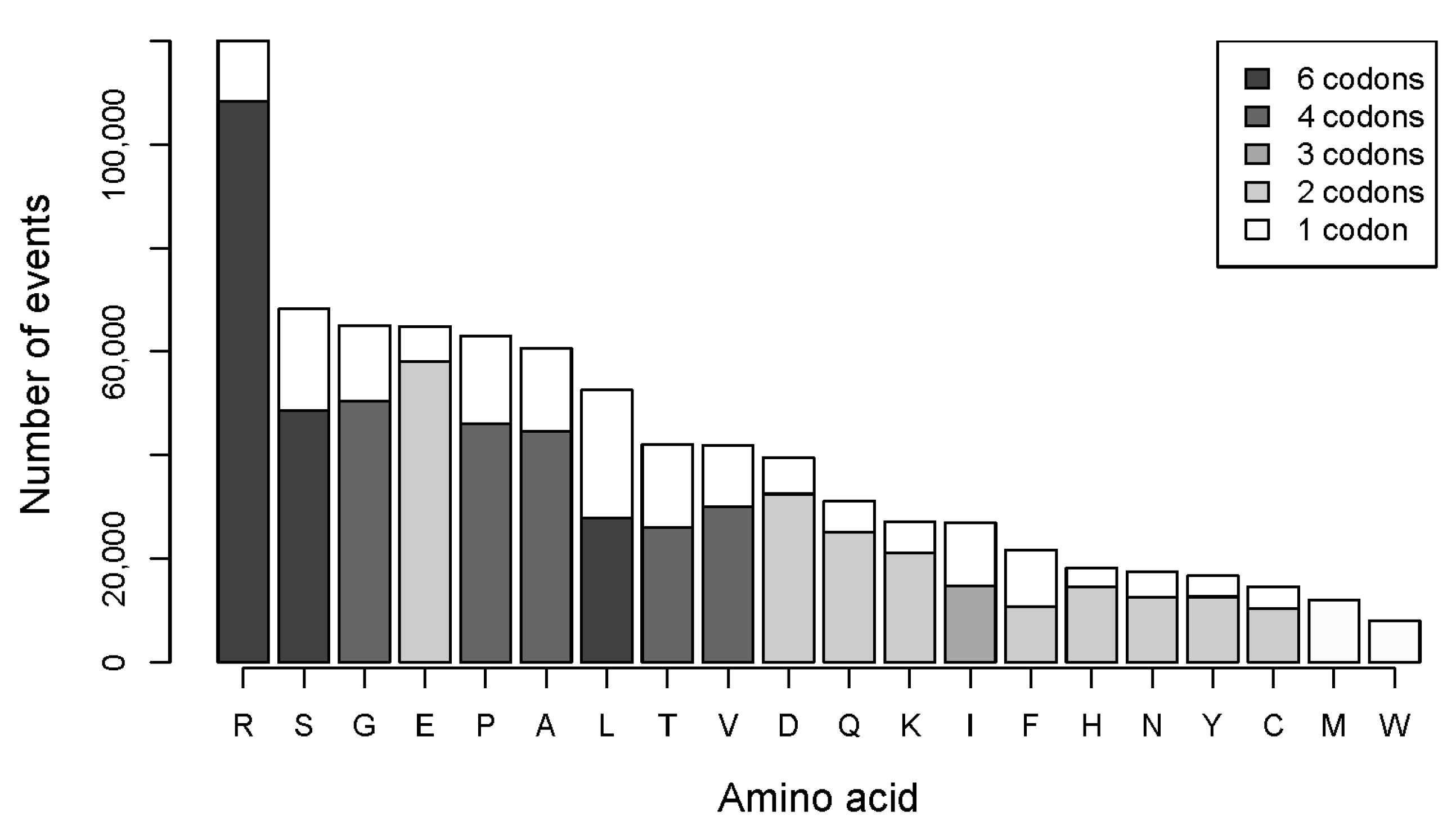
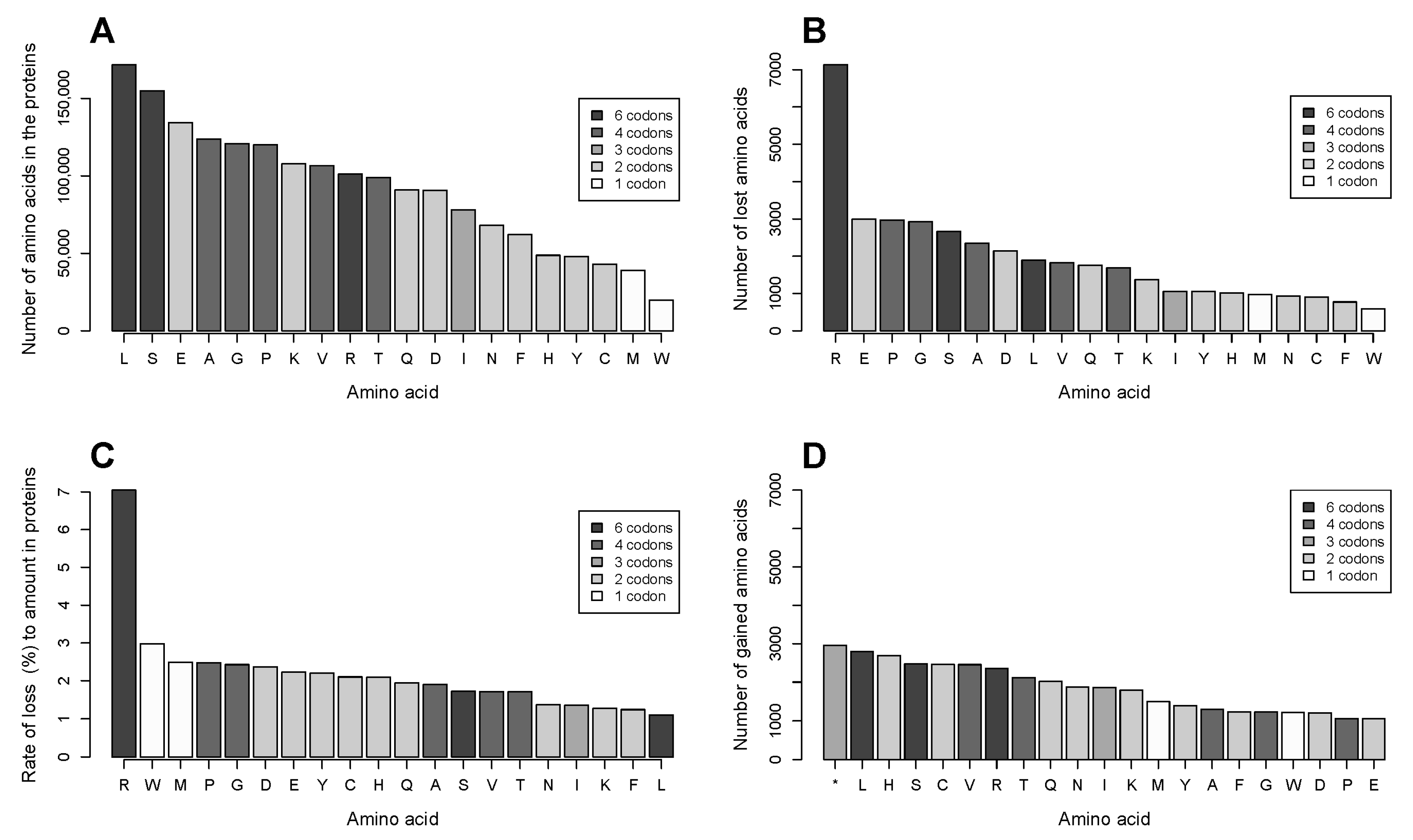
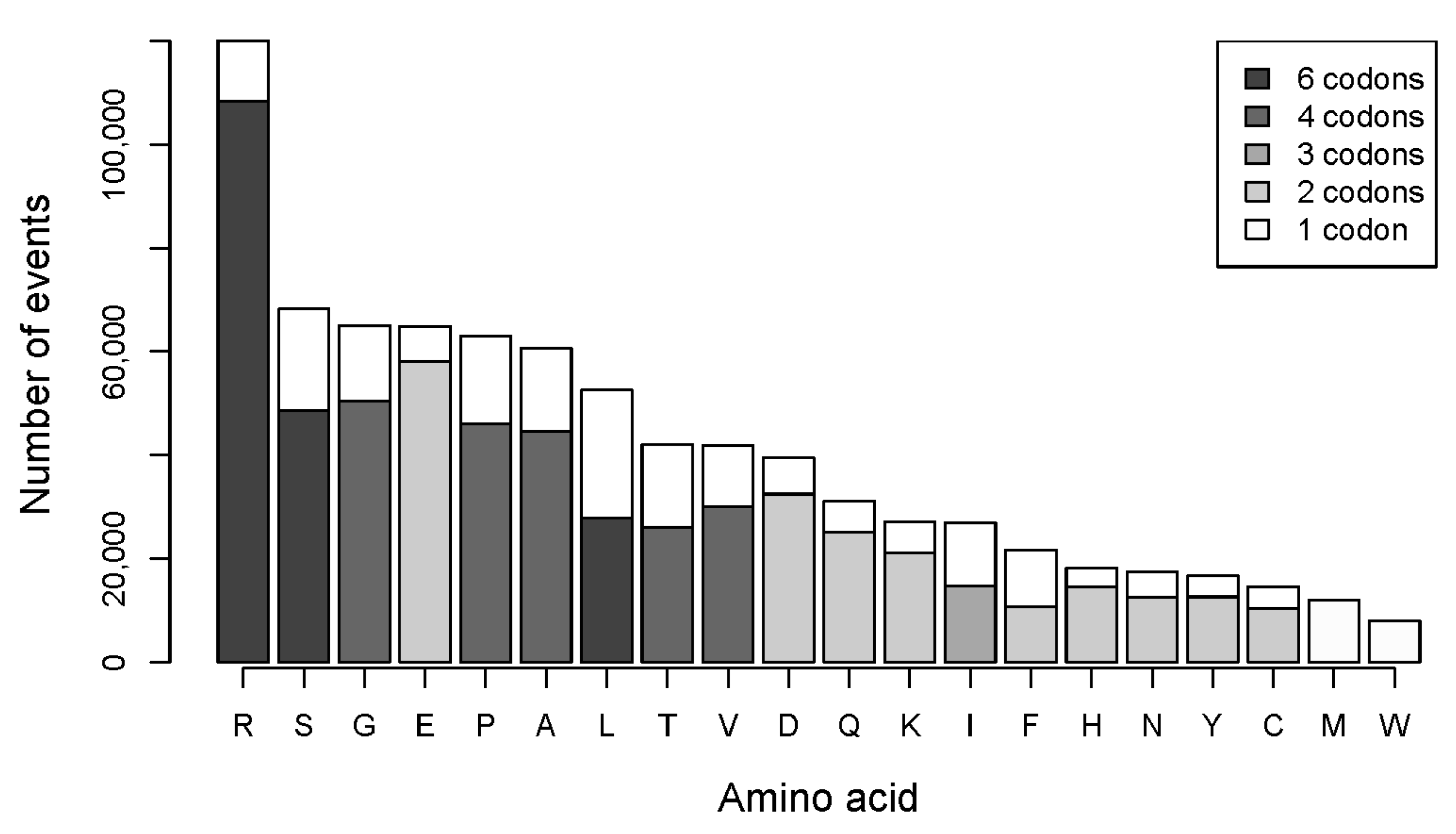
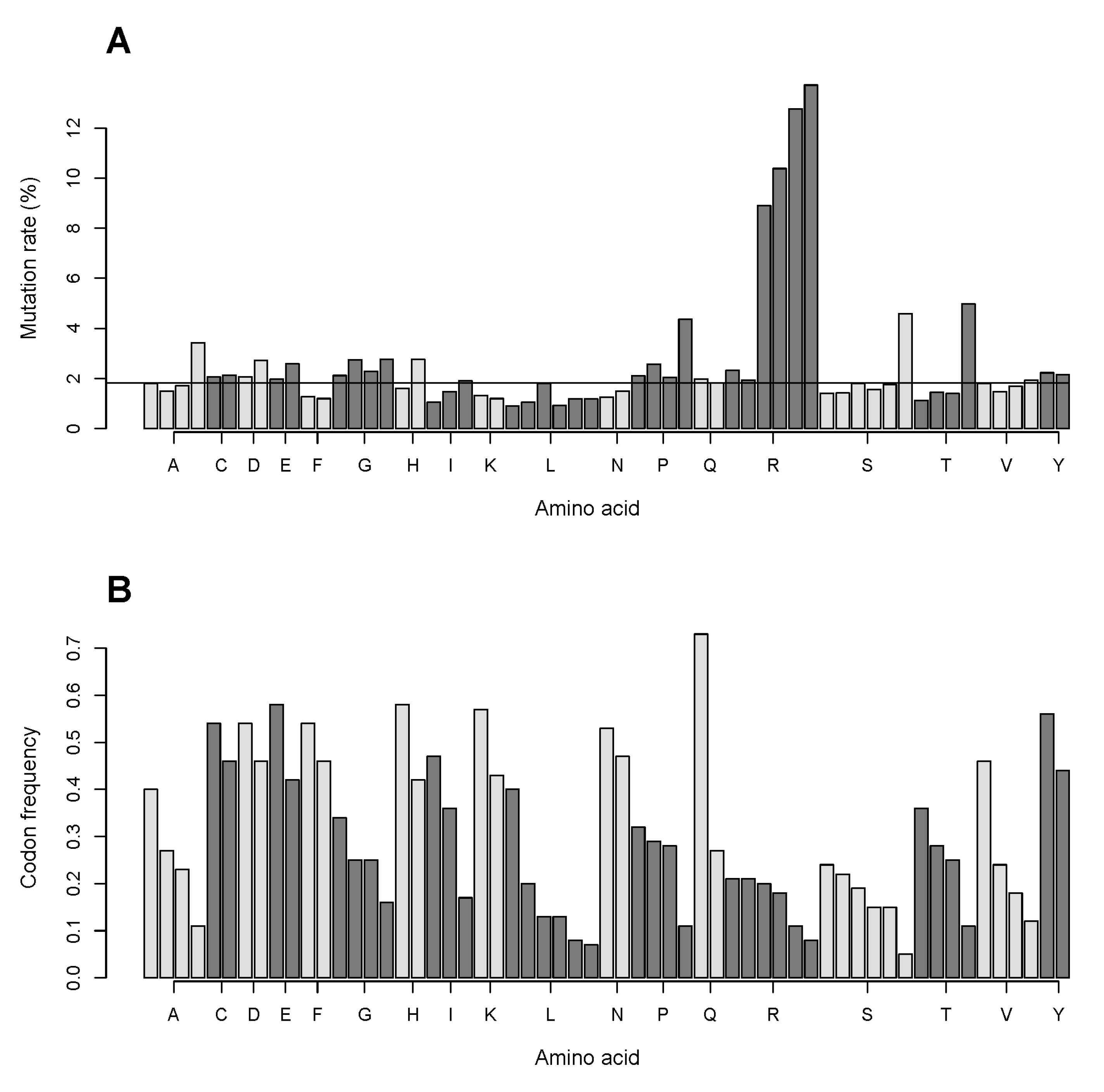
2.2. The Number of Coding Transformations of Amino Acids Is Not Proportional to Their Content in Proteins. Arginine Is Most Affected by Mutations Despite Not Being Most Abundant
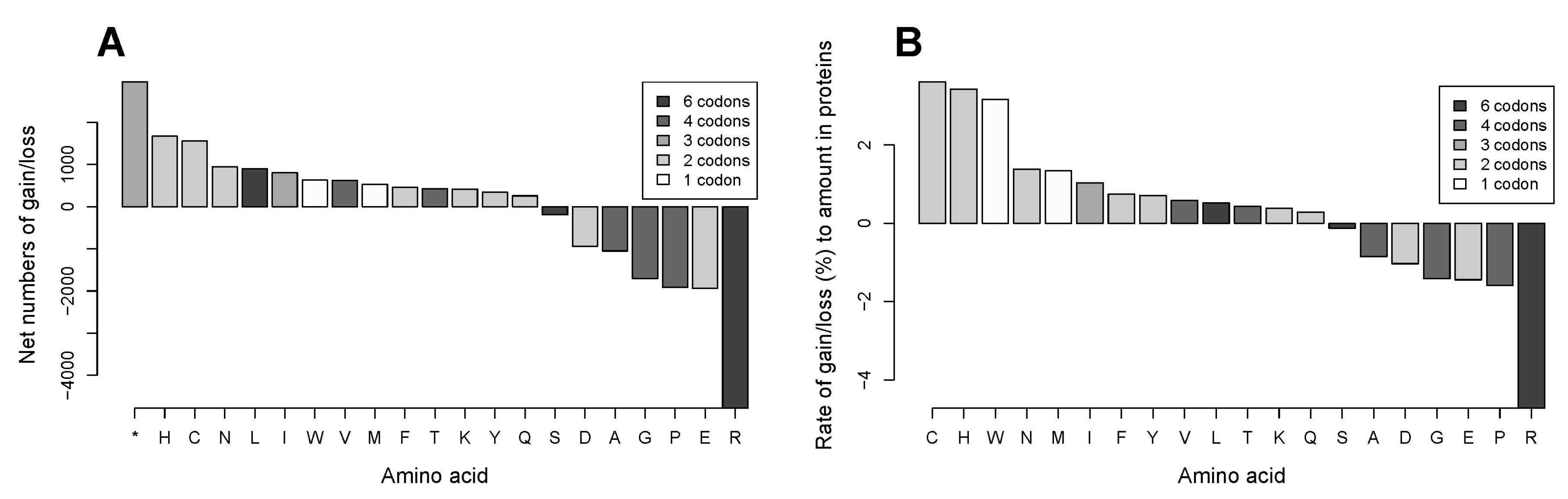
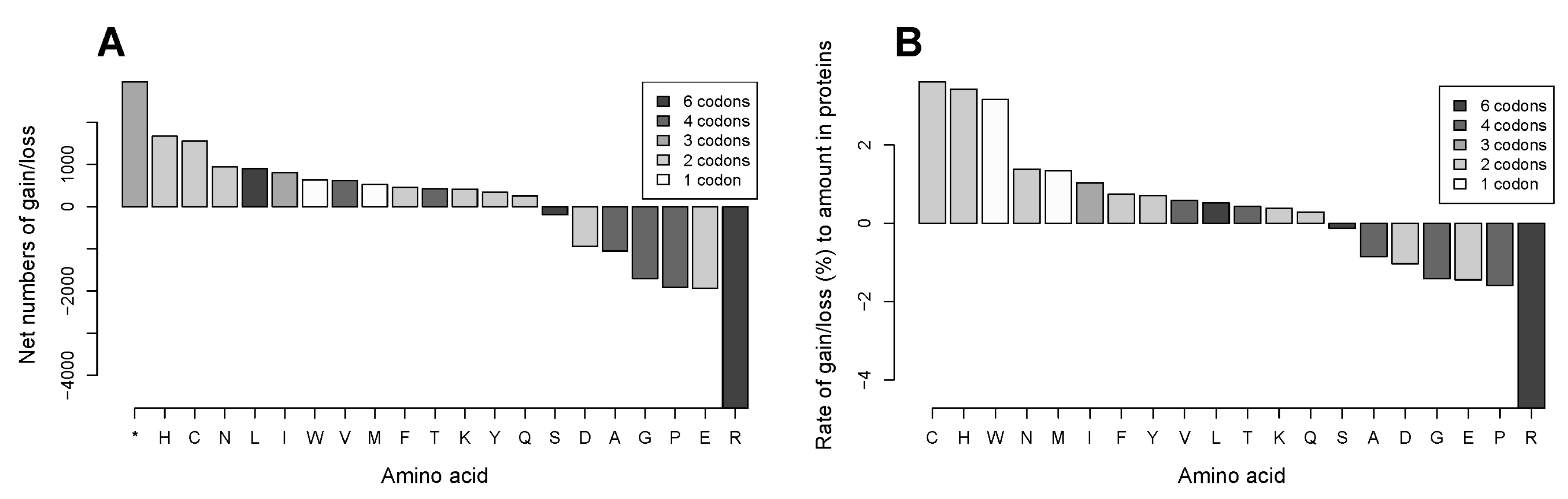
2.3. Gains of Histidine, Cysteine, and Tryptophan and Loss of Arginine Are Characteristic for the Total Outcome of All Coding Mutations in the Proteome
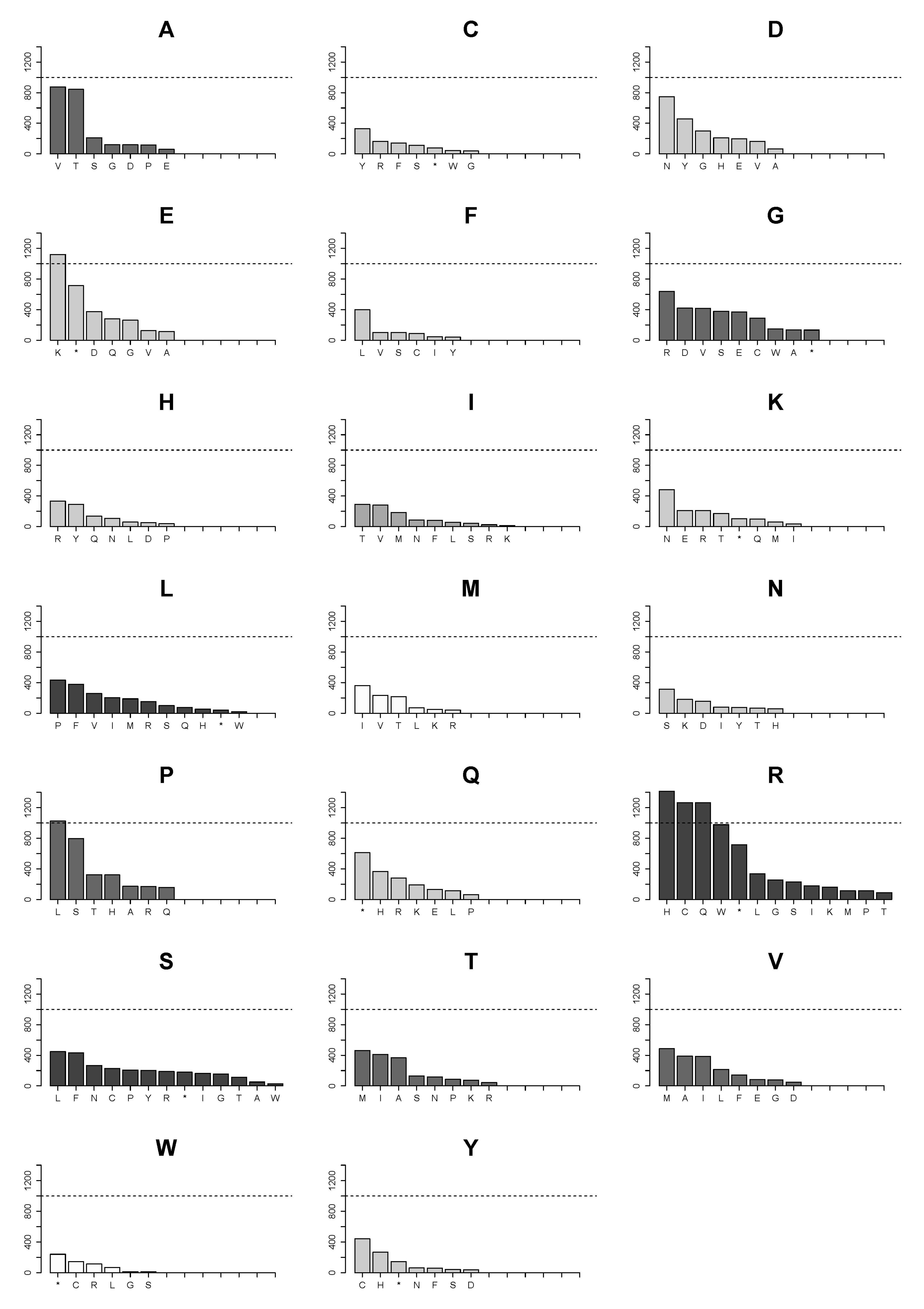
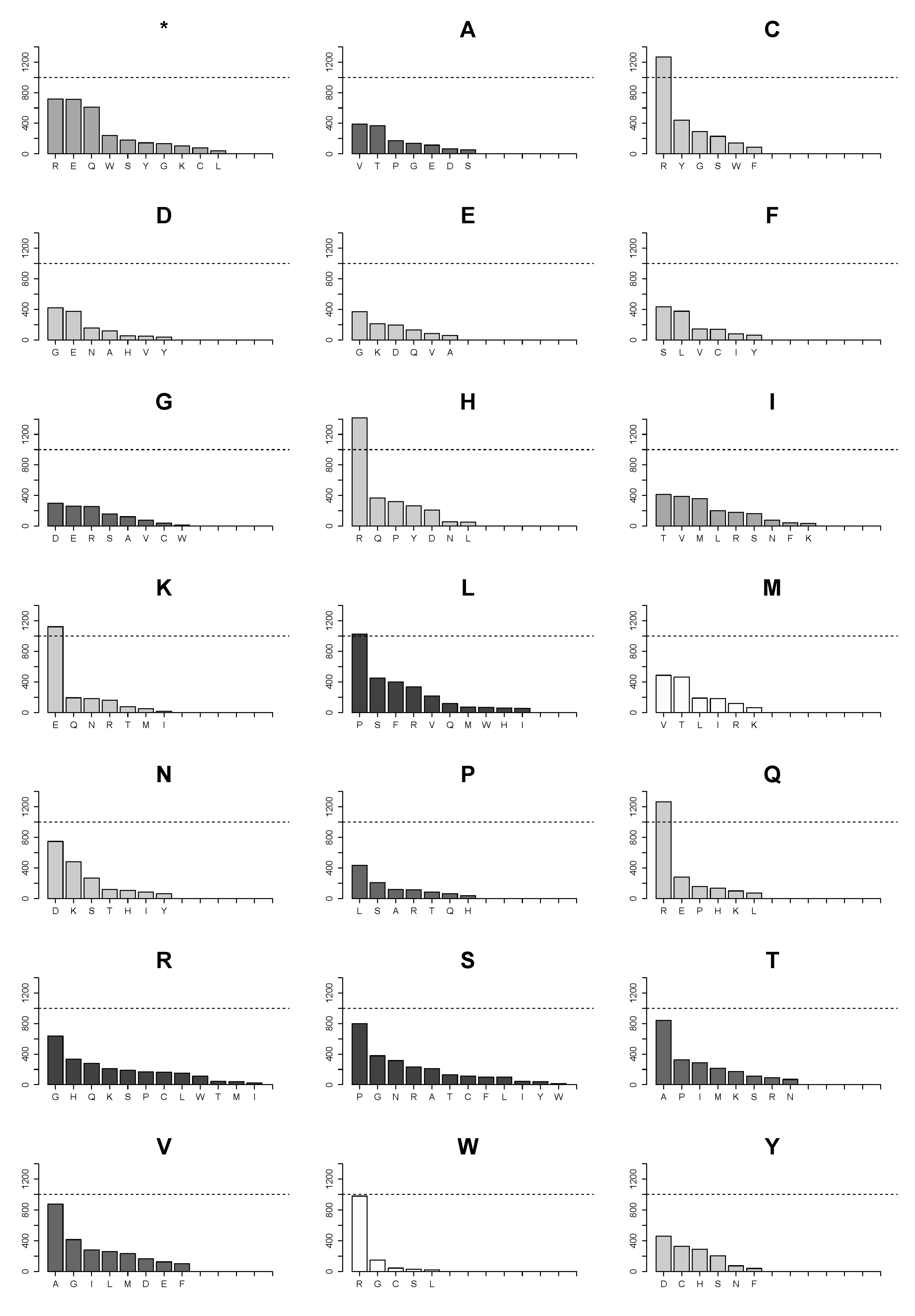
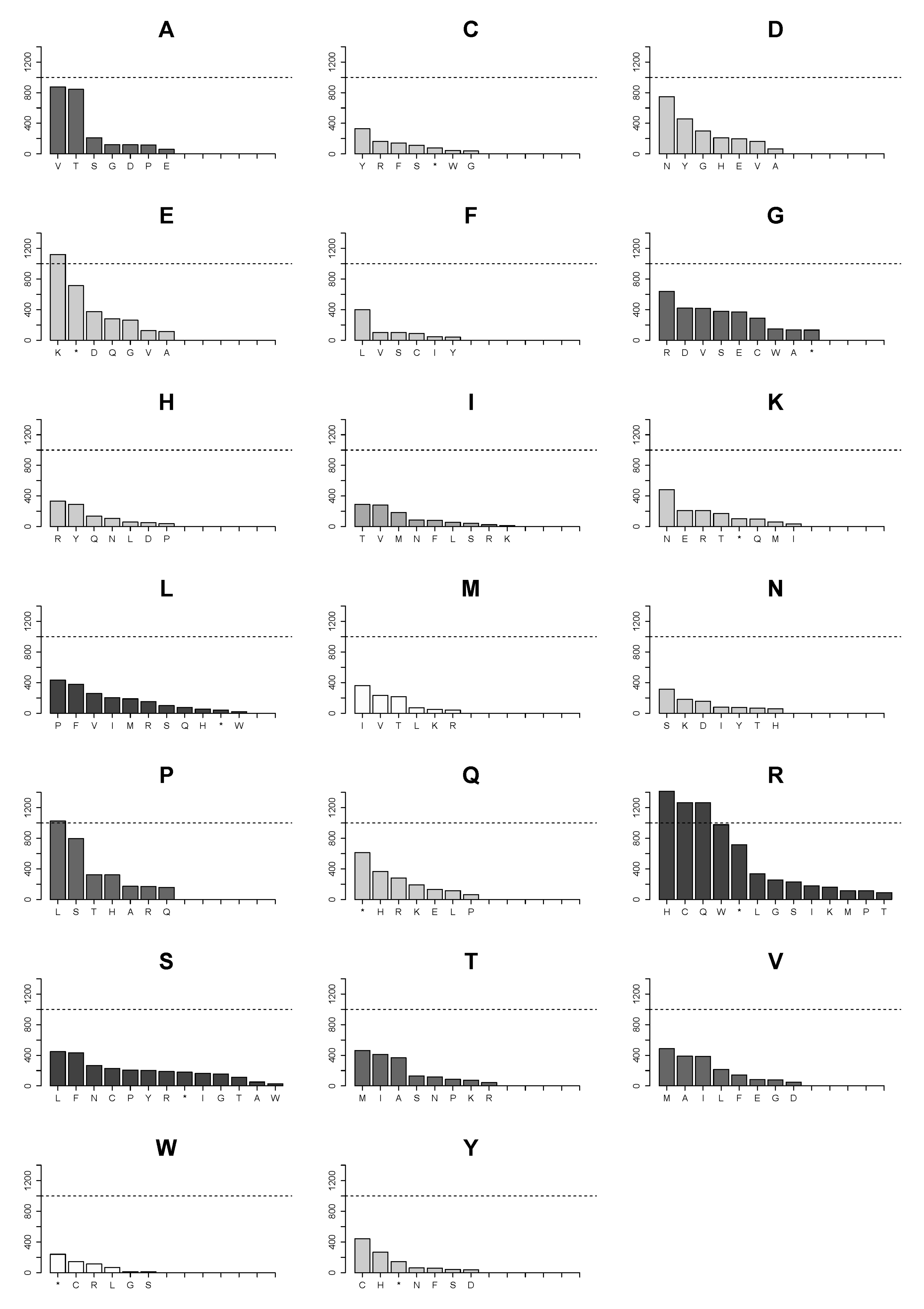
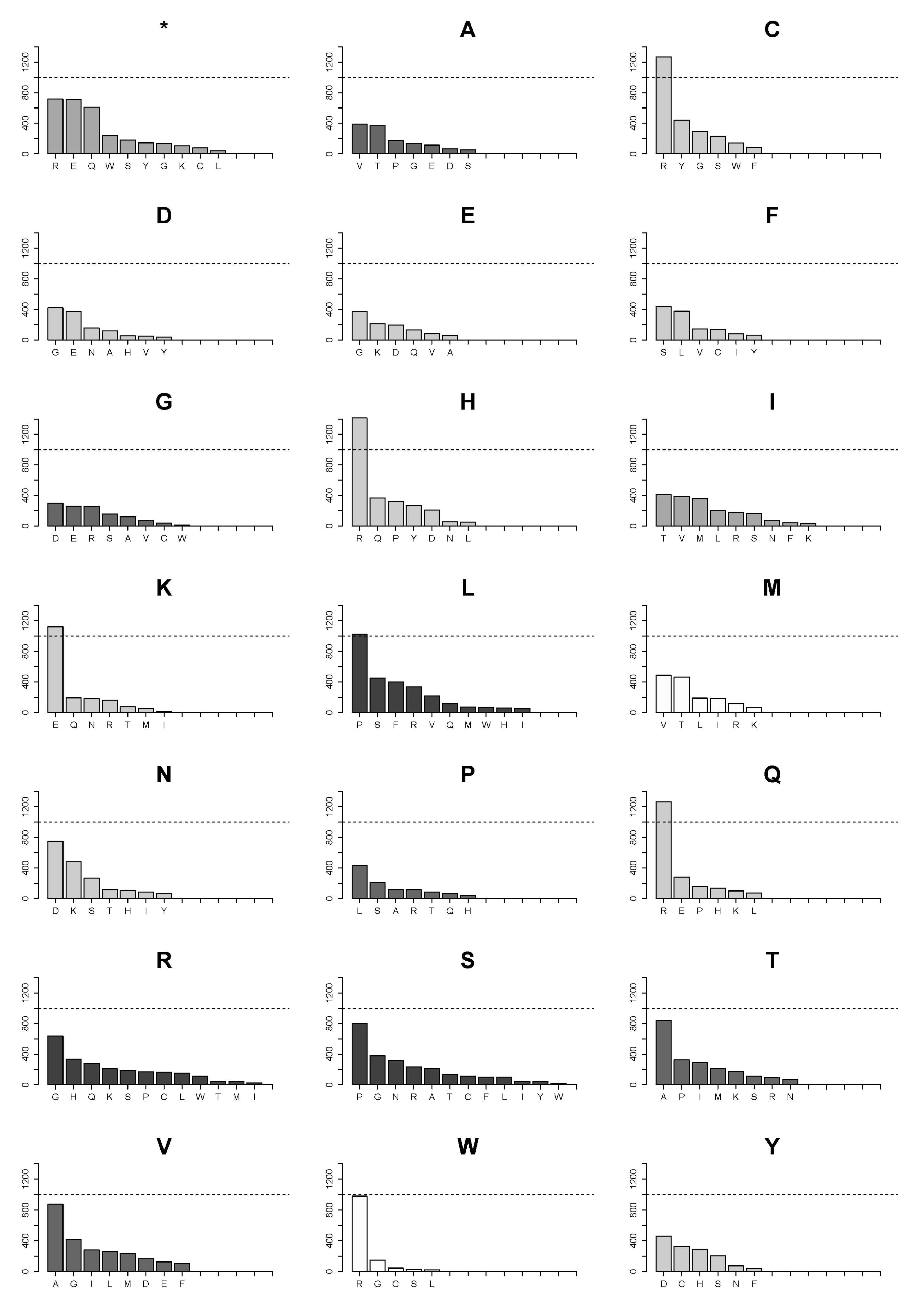
2.4. Changes of Arg>Cys, Arg>His, Arg>Gln, Arg>Trp, Glu>Lys and Pro>Leu Are the Few High Frequency Substitutions among 160 Possible Transformations for All Amino Acids in Single Nucleotide Substitutions in the Analyzed Proteome Subset
2.5. Landscape of Amino Acids Introduced by Mutations Is Manifestly Dominated by Those Originating from Arginine Substitutions
2.6. Validation of the CCLE Data with the Cosmic Database Confirmed Net Gains of Cysteine, Histidine and Tryptophan and Net Loss of Arginine in the Analyzed Subset of the Proteome; Arginine Was Also Found to Have the Smallest Ratio of Silent to Coding Mutations
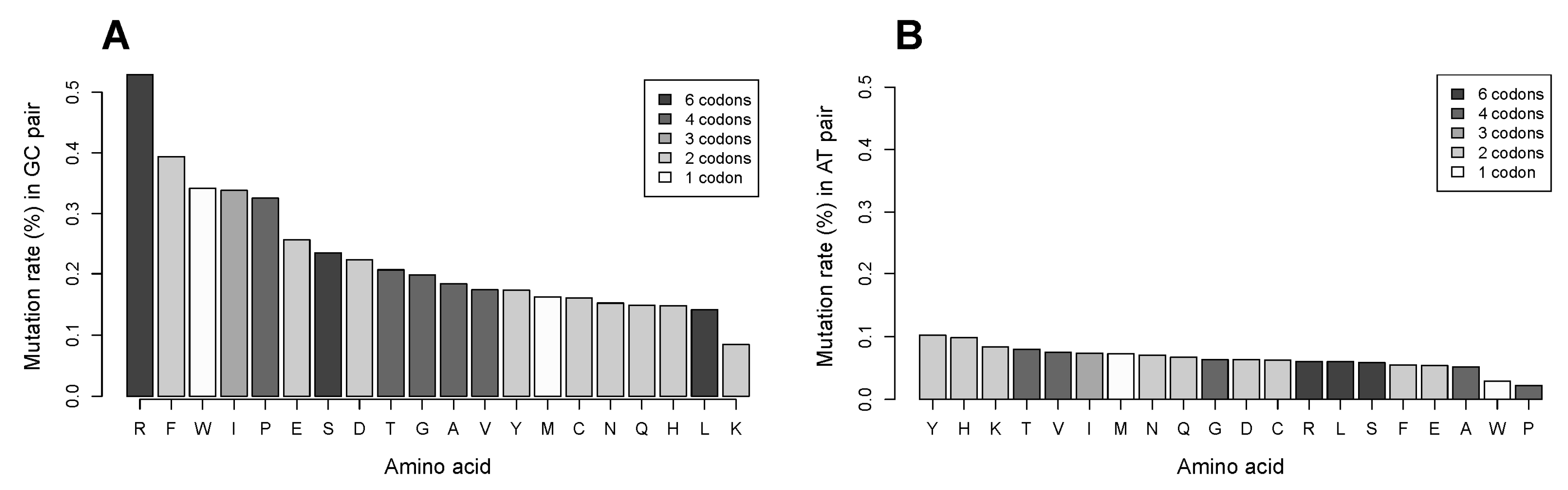
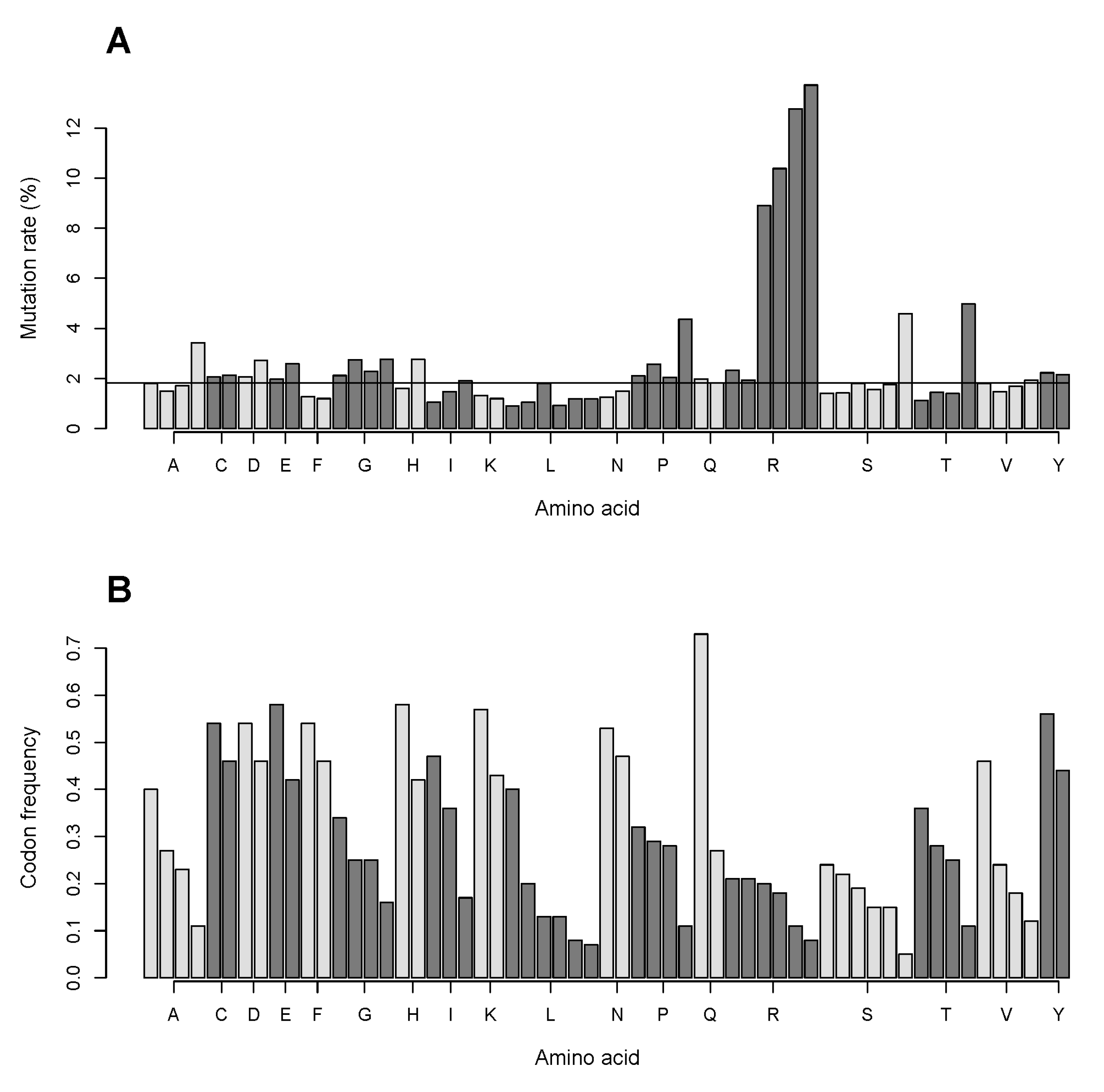
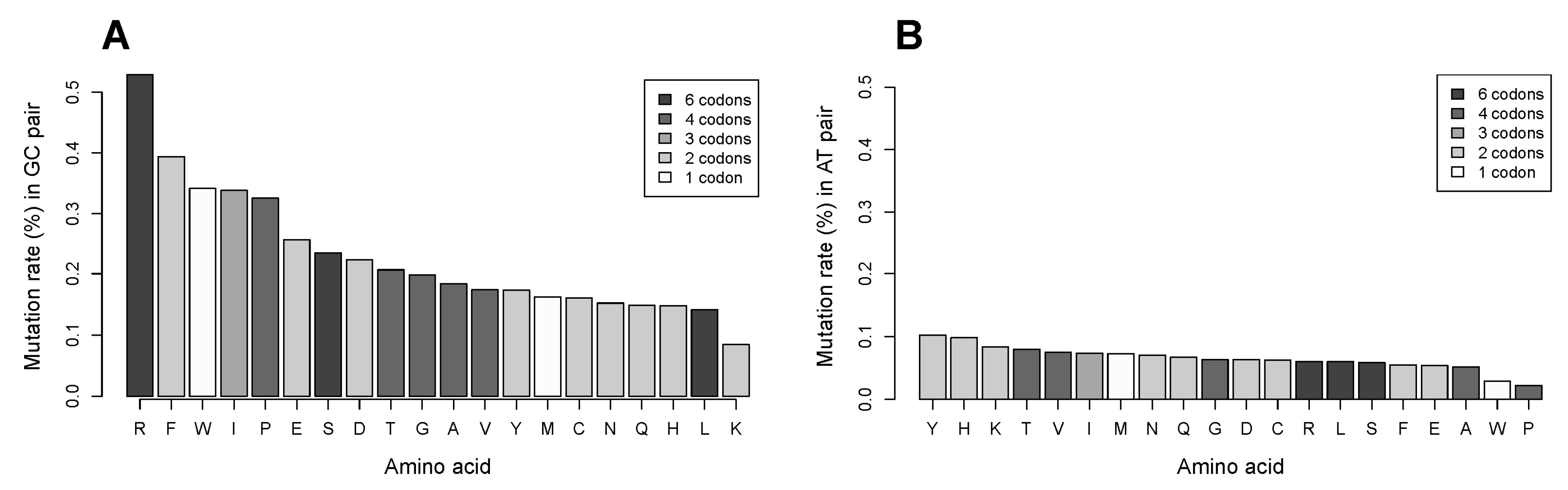
2.7. The GC Pair of Nucleotides in Codons Coding for Arginine Is More Frequently Targeted in Single Nucleotide Substitutions Compared to That for Other Amino Acids. Relationship of Mutation Frequency and Codon Usage Demonstrates an Extraordinarily High Rate for Four of Six Codons of Arginine Compared to Two Other Codons of Arginine and to All Codons of Other Amino Acids
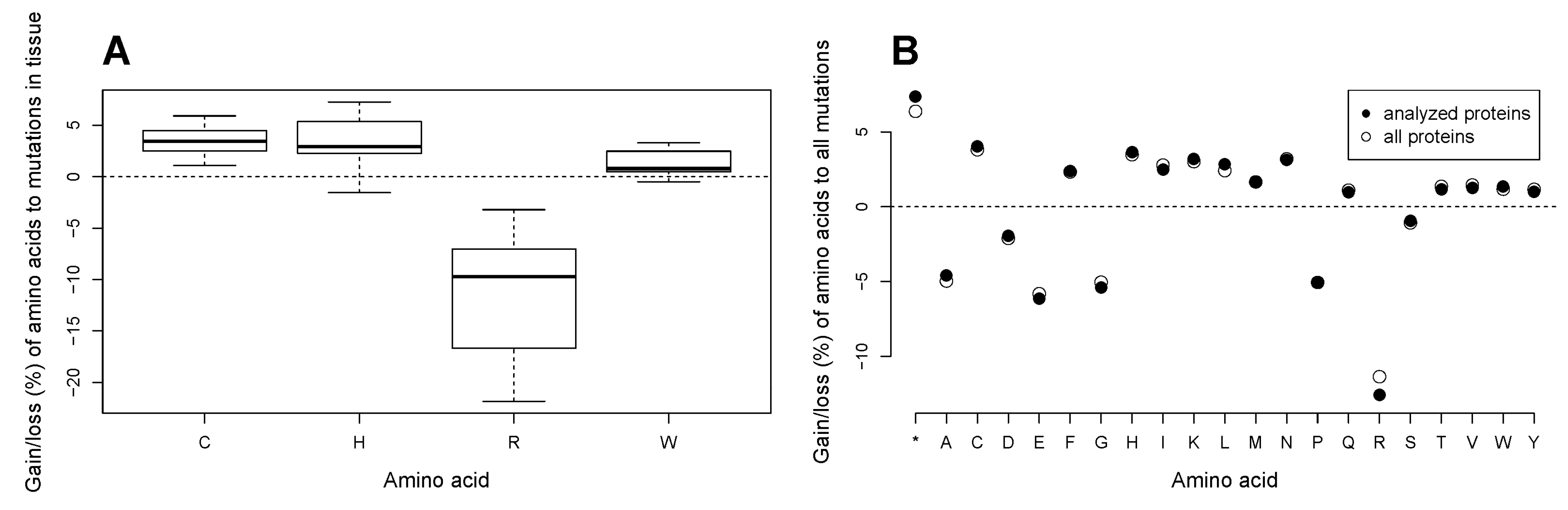
2.8. All Tissue Types in Cancer Undergo Loss of Arginine and Gain of Cysteine, and Most Tissue Types Gain Histidine and Tryptophan in Amino Acid Substitutions in the Analyzed Subset of Proteins. The Patterns of a Marked Loss of Arginine and High Gains of Cysteine and Histidine as Well as Uniform Gains of Some Amino Acids and Losses of the Others Are Universal for the Total Cancer Proteome
2.9. Loss of Arginine Is Prominent in Several Tumor Suppressor Proteins
3. Discussion
4. Materials and Methods
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lawrence, M.S.; Stojanov, P.; Polak, P.; Kryukov, G.V.; Cibulskis, K.; Sivachenko, A.; Carter, S.L.; Stewart, C.; Mermel, C.H.; Roberts, S.A.; et al. Mutational heterogeneity in cancer and the search for new cancer genes. Nature 2014, 499, 214–218. [Google Scholar] [CrossRef] [PubMed]
- Alexandrov, L.B.; Nik-Zainal, S.; Wedge, D.C.; Aparicio, S.A.; Behjati, S.; Biankin, A.V.; Bignell, G.R.; Bolli, N.; Borg, A.; Børresen-Dale, A.L.; et al. Signatures of mutational processes in human cancer. Nature 2013, 500, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.; Bao, J.; Zhou, X. Genome-wide mutational spectra analysis reveals significant cancer-specific heterogeneity. Sci. Rep. 2015, 5, 12566. [Google Scholar] [CrossRef] [PubMed]
- Barretina, J.; Caponigro, G.; Stransky, N.; Venkatesan, K.; Margolin, A.A.; Kim, S.; Wilson, C.J.; Lehár, J.; Kryukov, G.V.; Sonkin, D.; et al. The Cancer Cell Line Encyclopedia enables predictive modeling of anticancer drug sensitivity. Nature 2012, 483, 603–607. [Google Scholar] [CrossRef] [PubMed]
- Forbes, S.A.; Beare, D.; Gunasekaran, P.; Leung, K.; Bindal, N.; Boutselakis, H.; Ding, M.; Bamford, S.; Cole, C.; Ward, S.; et al. COSMIC: Exploring the world’s knowledge of somatic mutations in human cancer. Nucleic Acids Res. 2015, 43, D805–D811. [Google Scholar] [CrossRef] [PubMed]
- Nikolaev, S.I.; Sotiriou, S.K.; Pateras, I.S.; Santoni, F.; Sougioultzis, S.; Edgren, H.; Almusa, H.; Robyr, D.; Guipponi, M.; Saarela, J.; et al. A single-nucleotide substitution mutator phenotype revealed by exome sequencing of human colon adenomas. Cancer Res. 2012, 72, 6279–6289. [Google Scholar] [CrossRef] [PubMed]
- Helleday, T.; Eshtad, S.; Nik-Zainal, S. Mechanisms underlying mutational signatures in human cancers. Nat. Rev. Genet. 2014, 15, 585–598. [Google Scholar] [CrossRef] [PubMed]
- Codon Usage Database. Available online: http://www.kazusa.or.jp/codon/cgi-bin/showcodon.cgi?species=9606&aa=1&style=N (accessed on 14 January 2017).
- Shihab, H.A.; Gough, J.; Cooper, D.N.; Day, I.N.M.; Gaunt, T.R. Predicting the Functional Consequences of Cancer-Associated Amino Acid Substitutions. Bioinformatics 2013, 29, 1504–1510. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Texada, D.E. Low-usage codons and rare codons of Escherichia coli. Gene Ther. Mol. Biol. 2006, 10, 1–12. [Google Scholar]
- Angov, E. Codon usage: Nature’s roadmap to expression and folding of proteins. Biotechnol. J. 2011, 6, 650–659. [Google Scholar] [CrossRef] [PubMed]
- Jordan, I.K.; Kondrashov, F.A.; Adzhubei, I.A.; Wolf, Y.I.; Koonin, E.V.; Kondrashov, A.S.; Sunyaev, S. A universal trend of amino acid gain and loss in protein evolution. Nature 2005, 433, 633–638. [Google Scholar] [CrossRef] [PubMed]
- Klutstein, M.; Nejman, D.; Greenfield, R.; Cedar, H. DNA methylation in cancer and aging. Cancer Res. 2016, 76, 3446–3450. [Google Scholar] [CrossRef] [PubMed]
- Sproul, D.; Meehan, R.R. Genomic insights into cancer-associated aberrant CpG island hypermethylation. Brief. Funct. Genom. 2013, 12, 174–190. [Google Scholar] [CrossRef] [PubMed]
- Bartkova, J.; Horejsí, Z.; Koed, K.; Krämer, A.; Tort, F.; Zieger, K.; Guldberg, P.; Sehested, M.; Nesland, J.M.; Lukas, C.; et al. DNA damage response as a candidate anti-cancer barrier in early human tumorigenesis. Nature 2005, 434, 864–870. [Google Scholar] [CrossRef] [PubMed]
- Marino, S.M.; Gladyshev, V.N. Cysteine function governs its conservation and degeneration and restricts its utilization on protein surface. J. Mol. Biol. 2010, 404, 902–916. [Google Scholar] [CrossRef] [PubMed]
- Trifonov, E.N. The triplet code from first principles. J. Biomol. Struct. Dyn. 2004, 22, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Miseta, A.; Csutora, P. Relationship between the occurrence of cysteine in proteins and the complexity of organisms. Mol. Biol. Evol. 2000, 17, 1232–1239. [Google Scholar] [CrossRef] [PubMed]
- Maddocks, O.D.K.; Athineos, D.; Cheung, E.C.; Lee, P.; Zhang, T.; van den Broek, N.J.F.; Mackay, G.M.; Labuschagne, C.F.; Gay, D.; Kruiswijk, F.; et al. Modulating the therapeutic response of tumors to dietary serine and glycine starvation. Nature 2017, 544, 372–376. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- RStudio Team. RStudio: Integrated Development for R; RStudio, Inc.: Boston, MA, USA, 2015. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Coding for the Protein | Number of Arginine Substitution Events in the Protein | Total Number of Substitutions in the Protein in the CCLE Database | Number of Arginine Residues in the Protein | Number of Amino Acids in the Protein | Ratio of Substituted Arginines to Total Number of Mutations | Ratio of Substituted Arginines to Number of Arginine Residues | Ratio of all Non-Arginine Substitutions to Total number of Non-Arginine Amino acids |
|---|---|---|---|---|---|---|---|
| TP53 | 188 | 458 | 26 | 393 | 0.41 | 7.23 | 0.74 |
| PTEN | 22 | 74 | 20 | 403 | 0.30 | 1.10 | 0.14 |
| FBXW7 | 26 | 44 | 43 | 707 | 0.59 | 0.60 | 0.03 |
| CASP8 | 11 | 27 | 26 | 479 | 0.41 | 0.42 | 0.04 |
| STK4 | 10 | 14 | 24 | 487 | 0.71 | 0.42 | 0.01 |
| TNNI3K | 13 | 66 | 36 | 835 | 0.20 | 0.36 | 0.07 |
| ADAMTSL3 | 37 | 101 | 106 | 1691 | 0.37 | 0.35 | 0.04 |
| GNAS | 10 | 54 | 29 | 394 | 0.19 | 0.34 | 0.12 |
| ZNF423 | 14 | 51 | 47 | 1284 | 0.27 | 0.30 | 0.03 |
| HIP1 | 17 | 33 | 59 | 1037 | 0.52 | 0.29 | 0.02 |
| RB1 | 13 | 58 | 46 | 928 | 0.22 | 0.28 | 0.05 |
| CDK18 | 10 | 18 | 36 | 472 | 0.56 | 0.28 | 0.02 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsuber, V.; Kadamov, Y.; Brautigam, L.; Berglund, U.W.; Helleday, T. Mutations in Cancer Cause Gain of Cysteine, Histidine, and Tryptophan at the Expense of a Net Loss of Arginine on the Proteome Level. Biomolecules 2017, 7, 49. https://doi.org/10.3390/biom7030049
Tsuber V, Kadamov Y, Brautigam L, Berglund UW, Helleday T. Mutations in Cancer Cause Gain of Cysteine, Histidine, and Tryptophan at the Expense of a Net Loss of Arginine on the Proteome Level. Biomolecules. 2017; 7(3):49. https://doi.org/10.3390/biom7030049
Chicago/Turabian StyleTsuber, Viktoriia, Yunus Kadamov, Lars Brautigam, Ulrika Warpman Berglund, and Thomas Helleday. 2017. "Mutations in Cancer Cause Gain of Cysteine, Histidine, and Tryptophan at the Expense of a Net Loss of Arginine on the Proteome Level" Biomolecules 7, no. 3: 49. https://doi.org/10.3390/biom7030049