

Mitochondria Play Essential Roles in Intracellular Protection against Oxidative Stress—Which Molecules among the ROS Generated in the Mitochondria Can Escape the Mitochondria and Contribute to Signal Activation in Cytosol?

 , ,
, ,  , , , , , , , ,
, , , , , , , , 
Abstract
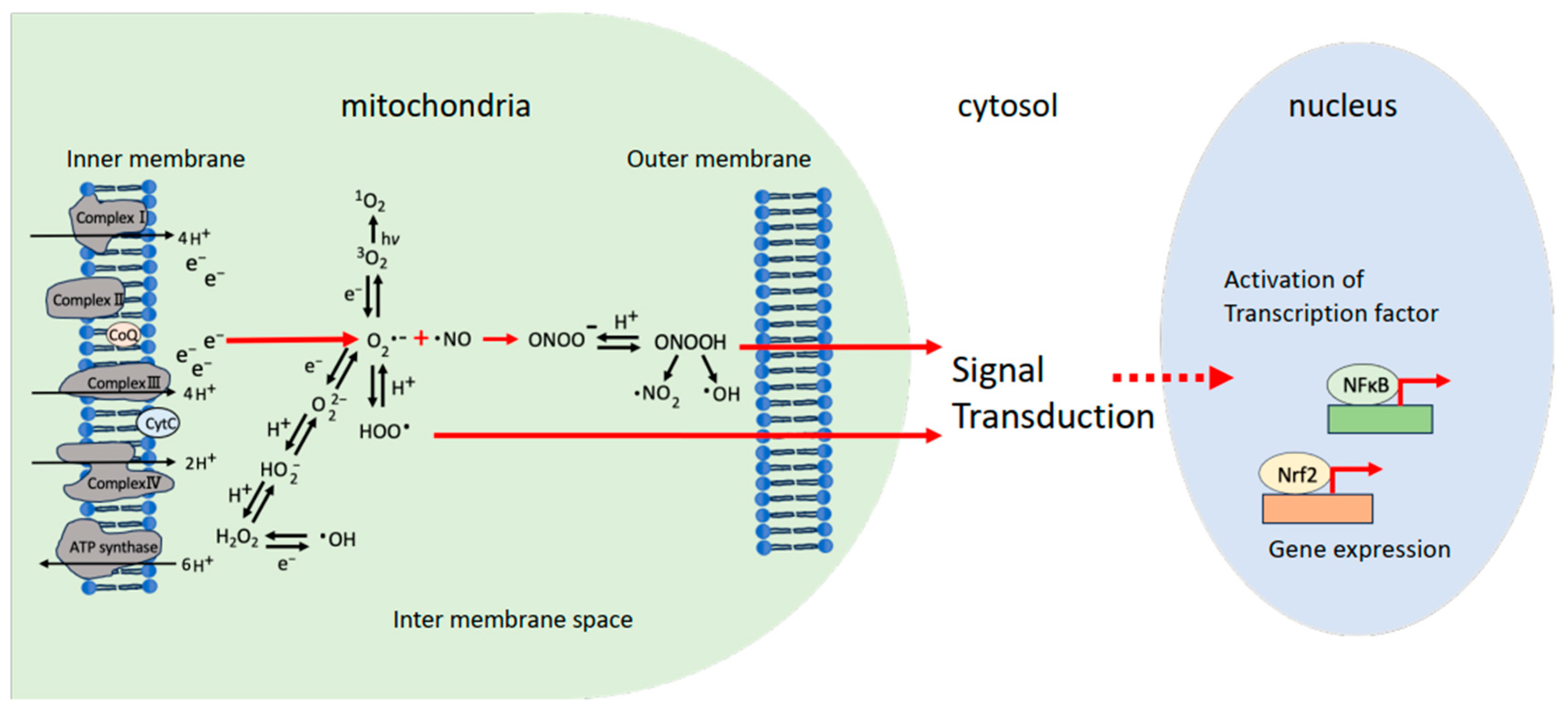
:1. Introduction
2. Materials and Methods
2.1. Theoretical Calculations of Dipole Moments for ROS and RNS
2.2. Predictive Performance of Mitochondria-Originating Reactive Oxygen Species
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Halliwell, B.; Whiteman, M. Measuring reactive species and oxidative damage in vivo and in cell culture: How should you do it and what do the results mean? Br. J. Pharmacol. 2004, 142, 231–255. [Google Scholar] [CrossRef]
- Majima, H.J.; Indo, H.P.; Suenaga, S.; Kaneko, T.; Matsui, H.; Yen, H.-C.; Ozawa, T. Mitochondria as source of free radicals. In Free Radical Biology in Digestive Diseases; Naito, Y., Suematsu, M., Yoshikawa, T., Eds.; Karger Publisher: Basel, Switzerland, 2011; Volume 29, pp. 12–22. [Google Scholar] [CrossRef]
- Indo, H.P.; Davidson, M.; Yen, H.-C.; Suenaga, S.; Tomita, K.; Nishii, T.; Higuchi, M.; Koga, Y.; Ozawa, T.; Majima, H.J. Evidence of ROS generation by mitochondria in cells with impaired electron transport chain and mitochondrial DNA damage. Mitochondrion 2007, 7, 106–118. [Google Scholar] [CrossRef]
- DiMauro, S.; Schon, E.A. Mitochondrial respiratory-chain diseases. N. Engl. J. Med. 2003, 348, 2656–2668. [Google Scholar] [CrossRef]
- DiMauro, S.; Schon, E.A.; Carelli, V.; Hirano, M. The clinical maze of mitochondrial neurology. Nat. Rev. Neurol. 2013, 9, 429–444. [Google Scholar] [CrossRef]
- Salvador, A.; Sousa, J.; Pinto, R.E. Hydroperoxyl, superoxide and pH gradients in the mitochondrial matrix: A theoretical assessment. Free Radic. Biol. Med. 2001, 31, 1208–1215. [Google Scholar] [CrossRef]
- Tengan, C.H.; Moraes, C.T. NO control of mitochondrial function in normal and transformed cells. Biochim. Biophys. Acta Bioenerg. 2017, 1858, 573–581. [Google Scholar] [CrossRef]
- Onyango, A.N. Endogenous Generation of singlet oxygen and ozone in human and animal tissues: Mechanisms, Biological significance, and influence of dietary components. Oxid. Med. Cell Longev. 2016, 2016, 2398573. [Google Scholar] [CrossRef]
- Kerver, E.D.; Vogels, I.M.; Bosch, K.S.; Vreeling-Sindelárová, H.; Van den Munckhof, R.J.; Frederiks, W.M. In situ detection of spontaneous superoxide anion and singlet oxygen production by mitochondria in rat liver and small intestine. Histochem. J. 1997, 29, 229–237. [Google Scholar] [CrossRef]
- McCord, J.M.; Fridovich, I. Superoxide dismutase. An enzymic function for erythrocuprein (hemocuprein). J. Biol. Chem. 1969, 244, 6049–6055. [Google Scholar] [CrossRef]
- Weisiger, R.A.; Fridovich, I. Mitochondrial superoxide dismutase. J. Biol. Chem. 1973, 248, 4793–4796. [Google Scholar] [CrossRef]
- Fukai, T.; Siegfried, M.R.; Ushio-Fukai, M.; Cheng, Y.; Kojda, G.; Harrison, D.G. Regulation of the vascular extracellular superoxide dismutase by nitric oxide and exercise training. J. Clin. Investig. 2000, 105, 1631–1639. [Google Scholar] [CrossRef]
- Okado-Matsumoto, A.; Fridovich, I. Subcellular distribution of superoxide dismutases (SOD) in rat liver Cu,Zn-SOD in mitochondria. J. Biol. Chem. 2001, 276, 38388–38393. [Google Scholar] [CrossRef]
- Indo, H.P.; Yen, H.-C.; Nakanishi, I.; Matsumoto, K.I.; Tamura, M.; Nagano, Y.; Matsui, H.; Gusev, O.; Cornette, R.; Okuda, T.; et al. A mitochondrial superoxide theory for oxidative stress diseases and aging. J. Clin. Biochem. Nutr. 2015, 56, 1–7. [Google Scholar] [CrossRef]
- Liochev, S.I.; Fridovich, I. Superoxide and iron: Partners in crime. IUBMB Life 1999, 48, 157–161. [Google Scholar] [CrossRef]
- Liochev, S.I.; Fridovich, I. The Haber-Weiss cycle—70 years later: An alternative view. Redox Rep. 2002, 7, 55–57; author reply 59–60. [Google Scholar] [CrossRef]
- Liochev, S.I.; Fridovich, I. The effects of superoxide dismutase on H2O2 formation. Free Radic. Biol. Med. 2007, 42, 1465–1469. [Google Scholar] [CrossRef]
- Davies, K.J. Oxidative stress, antioxidant defenses, and damage removal, repair, and replacement systems. IUBMB Life 2000, 50, 279–289. [Google Scholar] [CrossRef]
- Friedmann Angeli, J.P.; Schneider, M.; Proneth, B.; Tyurina, Y.Y.; Tyurin, V.A.; Hammond, V.J.; Herbach, N.; Aichler, M.; Walch, A.; Eggenhofer, E.; et al. Inactivation of the ferroptosis regulator Gpx4 triggers acute renal failure in mice. Nat. Cell Biol. 2014, 16, 1180–1191. [Google Scholar] [CrossRef]
- Azuma, K.; Koumura, T.; Iwamoto, R.; Matsuoka, M.; Terauchi, R.; Yasuda, S.; Shiraya, T.; Watanabe, S.; Aihara, M.; Imai, H.; et al. Mitochondrial glutathione peroxidase 4 is indispensable for photoreceptor development and survival in mice. J. Biol. Chem. 2022, 298, 101824. [Google Scholar] [CrossRef]
- Goldstein, S.; Rabai, J. Mechanism of nitrite formation by nitrate photolysis in aqueous solutions: The role of peroxynitrite, nitrogen dioxide, and hydroxyl radical. J. Am. Chem. Soc. 2007, 129, 10597–10601. [Google Scholar] [CrossRef]
- Kissner, R.; Nauser, T.; Kurz, C.; Koppenol, W.H. Peroxynitrous acid--where is the hydroxyl radical? IUBMB Life 2003, 55, 567–572. [Google Scholar] [CrossRef]
- Finkel, T. Signal transduction by mitochondrial oxidants. J. Biol. Chem. 2012, 287, 4434–4440. [Google Scholar] [CrossRef]
- Zhang, D.X.; Gutterman, D.D. Mitochondrial reactive oxygen species-mediated in endothelial cells. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H2023–H2031. [Google Scholar] [CrossRef]
- Cosentino-Goes, D.; Rocco-Machado, N.; Meyer-Fernandes, J.R. Cell signaling through protein kinase C oxidation and activation. Int. J. Mol. Sci. 2012, 13, 10697–10721. [Google Scholar] [CrossRef]
- Holmström, K.M.; Finkel, T. Cellular mechanisms and physiological consequences of redox-dependent signalling. Nat. Rev. Mol. Cell Biol. 2014, 15, 411–421. [Google Scholar] [CrossRef]
- Chandel, N.S. Evolution of Mitochondria as Signaling Organelles. Cell Metab. 2015, 22, 204–206. [Google Scholar] [CrossRef]
- Shadel, G.S.; Horvath, T.L. Mitochondrial ROS signaling in organismal homeostasis. Cell 2015, 163, 560–569. [Google Scholar] [CrossRef]
- Itoh, K.; Ye, P.; Matsumiya, T.; Tanji, K.; Ozaki, T. Emerging functional cross-talk between the Keap1-Nrf2 system and mitochondria. J. Clin. Biochem. Nutr. 2015, 56, 91–97. [Google Scholar] [CrossRef]
- Brand, M.D. Mitochondrial generation of superoxide and hydrogen peroxide as the source of mitochondrial redox signaling. Free Radic. Biol. Med. 2016, 100, 14–31. [Google Scholar] [CrossRef]
- Bouchez, C.; Devin, A. Mitochondrial biogenesis and mitochondrial reactive oxygen species (ROS): A complex relationship regulated by the cAMP/PKA signaling pathway. Cells 2019, 8, 287. [Google Scholar] [CrossRef]
- Brillo, V.; Chieregato, L.; Leanza, L.; Muccioli, S.; Costa, R. Mitochondrial dynamics, ROS, and cell signaling: A blended overview. Life 2021, 11, 332. [Google Scholar] [CrossRef]
- Kasai, S.; Kokubu, D.; Mizukami, H.; Itoh, K. Mitochondrial reactive oxygen species, insulin resistance, and Nrf2-mediated oxidative stress response-toward an actionable strategy for anti-aging. Biomolecules 2023, 13, 1544. [Google Scholar] [CrossRef]
- Indo, H.P.; Hawkins, C.L.; Nakanishi, I.; Matsumoto, K.-I.; Matsui, H.; Suenaga, S.; Davies, M.J.; St Clair, D.K.; Ozawa, T.; Majima, H.J. Role of Mitochondrial reactive oxygen species in the activation of cellular signals, molecules, and function. Handb. Exp. Pharmacol. 2017, 240, 439–456. [Google Scholar] [CrossRef]
- Indo, H.P.; Masuda, D.; Sriburee, S.; Ito, H.; Nakanishi, I.; Matsumoto, K.-I.; Mankhetkorn, S.; Chatatikun, M.; Surinkaew, S.; Udomwech, L.; et al. Evidence of Nrf2/Keap1 Signaling regulation by mitochondria-generated reactive oxygen species in RGK1 cells. Biomolecules 2023, 13, 445. [Google Scholar] [CrossRef]
- Cooper, G. Structure of the plasma membrane. In The Cell: A Molecular Approach, 2nd ed.; ASM Press: Washington, DC, USA, 2000; ISBN 0-87893-106-6. Available online: https://www.ncbi.nlm.nih.gov/books/NBK9898/ (accessed on 14 April 2023).
- Stein, W.D.; Lieb, W.R. Transport and Diffusion across Cell Membranes; Academic Press: New York, NY, USA, 1986. [Google Scholar] [CrossRef]
- Serricchio, M.; Vissa, A.; Kim, P.K.; Yip, C.M.; McQuibban, G.A. Cardiolipin synthesizing enzymes form a complex that interacts with cardiolipin-dependent membrane organizing proteins. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2018, 1863, 447–457. [Google Scholar] [CrossRef]
- Gasanoff, E.S.; Yaguzhinsky, L.S.; Garab, G. Cardiolipin, non-bilayer structures and mitochondrial bioenergetics: Relevance to cardiovascular disease. Cells 2021, 10, 1721. [Google Scholar] [CrossRef]
- Indo, H.P.; Inanami, O.; Koumura, T.; Suenaga, S.; Yen, H.C.; Kakinuma, S.; Matsumoto, K.; Nakanishi, I.; St Clair, W.; St Clair, D.K.; et al. Roles of mitochondria-generated reactive oxygen species on X-ray-induced apoptosis in a human hepatocellular carcinoma cell line, HLE. Free Radic. Res. 2012, 46, 1029–1043. [Google Scholar] [CrossRef]
- Clough, S.A.; Beers, Y.; Klein, G.P.; Rothman, L.S. Dipole moment of water from Stark measurements of H2O, HDO, and D2O. Chem. Phys. 1973, 59, 2254–2259. [Google Scholar] [CrossRef]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Nakatsuji, H.; Li, X.; et al. Gaussian 09, Revision A.02; Gaussian, Inc.: Wallingford, CT, USA, 2009; Available online: https://gaussian.com/g09citation/ (accessed on 30 April 2021).
- Becke, A.D. Density-functional thermochemistry. III. The role of exact exchange. J. Chem. Phys. 1993, 98, 5648–5652. [Google Scholar] [CrossRef]
- Lee, C.; Yang, W.; Parr, R.G. Development of the Colle-Salavati correlation-energy formula into a functional of the electron density. Phys. Rev. B Condens. Matter 1988, 37, 785–789. [Google Scholar] [CrossRef]
- Hay, J.J.; Wadt, R.W. Ab initio effective core potentials for molecular calculations. Potentials for the transition metal atoms Sc to Hg. J. Chem. Phys. 1985, 82, 270–283. [Google Scholar] [CrossRef]
- Olsen, J.; Jørgensen, P.P. Linear and nonlinear response functions for an exact state and for an MCSCF state. J. Chem. Phys. 1985, 82, 3235–3264. [Google Scholar] [CrossRef]
- Dennington, R.; Keith, T.; Millam, J. Gauss View, Version 6.1.1; Semichem, Inc.: Wallingford, CT, USA, 2003. [Google Scholar]
- Finkelstein, A.; Cass, A. Effect of cholesterol on the water permeability of thin lipid membranes. Nature 1967, 216, 717–718. [Google Scholar] [CrossRef]
- Chance, B.; Sies, H.; Boveris, A. Hydroperoxide metabolism in mammalian organs. Physiol. Rev. 1979, 59, 527–605. [Google Scholar] [CrossRef]
- Takahashi, M.; Asada, K. Superoxide anion permeability of phospholipid membranes and chloroplast thylakoids. Arch. Biochem. Biophys. 1983, 226, 558–566. [Google Scholar] [CrossRef]
- Gus’kova, R.A.; Ivanov, I.I.; Kol’tover, V.K.; Akhobadze, V.V.; Rubin, A.B. Permeability of bilayer lipid membranes for superoxide (O2−) radicals. Biochim. Biophys. Acta 1984, 778, 579–585. [Google Scholar] [CrossRef]
- Subczynski, W.K.; Lomnicka, M.; Hyde, J.S. Permeability of nitric oxide through lipid bilayer membranes. Free Radical. Res. 1996, 24, 343–349. [Google Scholar] [CrossRef]
- Mendiara, S.N.; Perissinotti, L.J. Dissociation equilibrium of dinitrogen tetroxide in organic solvents: An electron paramagnetic resonance measurement. Appl. Magn. Reson. 2003, 25, 323–346. [Google Scholar] [CrossRef]
- Denicola, A.; Souza, J.M.; Radi, R. Diffusion of peroxynitrite across erythrocyte membranes. Proc. Natl. Acad. Sci. USA 1998, 95, 3566–3571. [Google Scholar] [CrossRef]
- Marla, S.S.; Lee, J.; Groves, J.T. Peroxynitrite rapidly permeates phospholipid membranes. Proc. Natl. Acad. Sci. USA 1997, 94, 14243–14248. [Google Scholar] [CrossRef]
- Khairutdinov, R.F.; Coddington, J.W.; Hurst, J.K. Permeation of phospholipid membranes by peroxynitrite. Biochemistry 2000, 39, 14238–14249. [Google Scholar] [CrossRef] [PubMed]
- Trujillo, M.; Piacenza, L.; Radi, R. Reactivity of mitochondrial peroxiredoxins with biological hydroperoxides. Redox Biochem. Chem. 2023, 5–6, 100017. [Google Scholar] [CrossRef]
- Möller, M.N.; Lancaster, J.R., Jr.; Denicola, A. Chapter 2 The interaction of reactive oxygen and nitrogen species with membranes. In Current Topics in Membranes; Matalon, S., Ed.; Academic Press Inc.: New York, NY, USA, 2008; Volume 61, pp. 23–42. [Google Scholar] [CrossRef]
- Möller, M.N.; Cuevasanta, E.; Orrico, F.; Lopez, A.C.; Thomson, L.; Denicola, A. Diffusion and Transport of Reactive Species Across Cell Membranes. Adv. Exp. Med. Biol. 2019, 1127, 3–19. [Google Scholar] [CrossRef]
- Armstrong, D.A.; Huie, R.E.; Lymar, S.; Koppenol, W.H.; Merényi, G.; Neta, P.; Stanbury, D.M.; Steenken, S.; Wardman, P. Standard electrode potentials involving radicals in aqueous solution: Inorganic radicals. BioInorg. React. Mech. 2013, 9, 59–61. [Google Scholar] [CrossRef]
- Buettner, G.R.; Jurkiewicz, B.A. Catalytic metals, ascorbate and free radicals: Combinations to avoid. Radiat. Res. 1996, 145, 532–541. [Google Scholar] [CrossRef] [PubMed]
- Warren, J.J.; Tronic, T.A.; Mayer, J.M. Thermochemistry of proton-coupled electron transfer reagents and its implications. Chem. Rev. 2010, 110, 6961–7001. [Google Scholar] [CrossRef] [PubMed]
- Bartberger, M.D.; Liu, W.; Ford, E.; Miranda, K.M.; Switzer, C.; Fukuto, J.M.; Farmer, P.J.; Wink, D.A.; Houk, K.N. The reduction potential of nitric oxide (NO) and its importance to NO biochemistry. Proc. Natl. Acad. Sci. USA 2002, 99, 10958–10963. [Google Scholar] [CrossRef]
- Koppenol, W.H.; Stanbury, D.M.; Bounds, P.L. Electrode potentials of partially reduced oxygen species, from dioxygen to water. Free Radic. Biol. Med. 2010, 49, 317–322. [Google Scholar] [CrossRef]
- Finosh, G.Y.; Joyabalan, M. Reactive oxygen species—Control and management using amphiphilic biosynthetic hydrogels for cardiac applications. Adv. Biosci. Biotech. 2013, 4, 1134–1146. [Google Scholar] [CrossRef]
- Karogodina, T.Y.; Sergeeva, S.V.; Stass, D.V. Stability and reactivity of free radicals: A physicochemical perspective with biological implications. Hemoglobin 2011, 35, 262–275. [Google Scholar] [CrossRef]
- Phaniendra, A.; Jestadi, D.B.; Periyasamy, L. Free radicals: Properties, sources, targets, and their implication in various diseases. Indian J. Clin. Biochem. 2015, 30, 11–26. [Google Scholar] [CrossRef]
- Pryor, W.A. Oxy-radicals and related species: Their formation, lifetimes, and reactions. Annu. Rev. Physiol. 1986, 48, 657–667. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. Strategies of antioxidant defense. Eur. J. Biochem. 1993, 215, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Beckman, J.S. Peroxynitrite versus hydroxyl radical: The role of nitric oxide in superoxide-dependent cerebral injury. Ann. N. Y. Acad. Sci. 1994, 738, 69–75. [Google Scholar] [CrossRef]
- Bonini, M.G.; Augusto, O. Carbon dioxide stimulates the production of thiyl, sulfinyl, and disulfide radical anion from thiol oxidation by peroxynitrite. J. Biol. Chem. 2001, 276, 9749–9754. [Google Scholar] [CrossRef]
- Augusto, O.; Miyamoto, S. Oxygen radicals and related species. In Principles of Free Radical Biomedicine; Pantopoulos, K., Schipper, H.M., Eds.; Nova Science Publishers: Hauppauge, NY, USA, 2011; Volume 1, pp. 1–23. [Google Scholar]
- Winterbourn, C.C. Reconciling the chemistry and biology of reactive oxygen species. Nat. Chem. Biol. 2008, 4, 278–286. [Google Scholar] [CrossRef]
- Sueishi, M.; Hori, M.; Ishikawa, M.; Matsu-Ura, K.; Kamogawa, E.; Honda, Y.; Kita, M.; Ohara, K. Scavenging rate constants of hydrophilic antioxidants against multiple reactive oxygen species. J. Clin. Biochem. Nutr. 2014, 54, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Keszler, A.; Zhang, Y.; Hogg, N. Reaction between nitric oxide, glutathione, and oxygen in the presence and absence of protein: How are S-nitrosothiols formed? Free Radic. Biol. Med. 2010, 48, 55–64. [Google Scholar] [CrossRef]
- Adams, G.E.; Boag, J.W.; Currant, J.; Michael, B.D. Absolute rate constants for the reaction of the hydroxyl radical with organic compounds. In Pulse Radiolysis; Ebert, M., Keene, J.P., Swallow, A.J., Baxendale, J.H., Eds.; Academic Press: New York, NY, USA, 1965; pp. 131–143. [Google Scholar]
- Liphard, M.; Bothe, E.; Schulte-Frohlinde, D. The influence of glutathione on single-strand breakage in single-stranded DNA irradiated in aqueous solution in the absence and presence of oxygen. Int. J. Radiat. Biol. 1990, 58, 589–602. [Google Scholar] [CrossRef]
- Gupta, V.; Carroll, K.S. Sulfenic acid chemistry, detection and cellular lifetime. Biochim. Biophys. Acta 2014, 1840, 847–875. [Google Scholar] [CrossRef]
- Ross, F.; Alberta, B.; Ross, A.B. Selected Specific Rates of Reactions of Transients from Water in Aqueous Solution. III. Hydroxyl Radical and Perhydroxyl Radical and Their Radical Ions; Natl. Stand. Ref. Data Syst. No. 59; National Bureau of Standards: Washington, DC, USA, 1977.
- Eriksen, T.E.; Fransson, G. Formation of reducing radicals on radiolysis of glutathione and some related compounds in aqueous solution. J. Chem. Soc. Perkin Trans. 1988, 2, 1117–1122. [Google Scholar] [CrossRef]
- Carballal, S.; Bartesaghi, S.; Radi, R. Kinetic and mechanistic considerations to assess the biological fate of peroxynitrite. Biochim. Biophys. Acta 2014, 1840, 768–780. [Google Scholar] [CrossRef] [PubMed]
- Engstrom, P.C.; Easterling, L.; Baker, R.R.; Matalon, S. Mechanisms of extracellular hydrogen peroxide clearance by alveolar type II pneumocytes. J. Appl. Physiol. 1985, 69, 2078–2084. [Google Scholar] [CrossRef] [PubMed]
- Kramarenko, G.G.; Hummel, S.G.; Martin, S.M.; Buettner, G.R. Ascorbate reacts with singlet oxygen to produce hydrogen peroxide. Photochem. Photobiol. 2006, 82, 1634–1637. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, R.; Yagi, M.; Kikuchi, A. Suppression of riboflavin-sensitized singlet oxygen generation by l-ascorbic acid, 3-O-ethyl-l-ascorbic acid and Trolox. J. Photochem. Photobiol. B 2019, 191, 116–122. [Google Scholar] [CrossRef]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef] [PubMed]
- Ferrer-Sueta, G.; Campolo, N.; Trujillo, M.; Bartesaghi, S.; Carballal, S.; Romero, N.; Alvarez, B.; Radi, R. Biochemistry of peroxynitrite and protein tyrosine nitration. Chem. Rev. 2018, 118, 1338–1408. [Google Scholar] [CrossRef]
- Sheng, Y.; Abreu, I.A.; Cabelli, D.E.; Maroney, M.J.; Miller, A.F.; Teixeira, M.; Valentine, J.S. Superoxide dismutases and superoxide reductases. Chem. Rev. 2014, 114, 854–918. [Google Scholar] [CrossRef]
- Quijano, C.; Castro, L.; Peluffo, G.; Valez, V.; Radi, R. Enhanced mitochondrial superoxide in hyperglycemic endothelial cells: Direct measurements and formation of hydrogen peroxide and peroxynitrite. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H3404–H3414. [Google Scholar] [CrossRef]
- Asada, K.; Kanematsu, S. Reactivity of Thiols with Superoxide Radicals. Agric. Biol. Chem. 1976, 40, 1891–1892. [Google Scholar] [CrossRef]
- Kwon, B.G.; Kim, J.-O.; Kwon, J.-K. An Advanced Kinetic Method for HO2∙/O2-∙ Determination by Using Terephthalate in the Aqueous Solution. Environm. Engin. Res. 2012, 17, 205–210. [Google Scholar] [CrossRef]
- Bartlett, D.; Church, D.F.; Bounds, P.L.; Koppenol, W.H. The kinetics of the oxidation of L-ascorbic acid by peroxynitrite. Free Radic. Biol. Med. 1995, 18, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Hakim, T.S.; Sugimori, K.; Camporesi, E.M.; Anderson, G. Half-life of nitric oxide in aqueous solutions with and without haemoglobin. Physiol. Meas. 1996, 17, 267–277. [Google Scholar] [CrossRef]
- Zhong, Z.J.; Yao, Z.P.; Shi, Z.Q.; Liu, Y.D.; Liu, L.F.; Xin, G.Z. Measurement of intracellular Nnitric oxide with a quantitative mass spectrometry probe approach. Anal. Chem. 2021, 93, 8536–8543. [Google Scholar] [CrossRef]
- Hetrick, E.M.; Schoenfisch, M.H. Analytical chemistry of nitric oxide. Annu. Rev. Anal. Chem. 2009, 2, 409–433. [Google Scholar] [CrossRef]
- Thomas, D.D.; Ridnour, L.A.; Isenberg, J.S.; Flores-Santana, W.; Switzer, C.H.; Donzelli, S.; Hussain, P.; Vecoli, C.; Paolocci, N.; Ambs, S.; et al. The chemical biology of nitric oxide: Implications in cellular signaling. Free Radic. Biol. Med. 2008, 45, 18–31. [Google Scholar] [CrossRef]
- Ford, E.; Hughes, M.N.; Wardman, P. Kinetics of the reactions of nitrogen dioxide with glutathione, cysteine, and uric acid at physiological pH. Free Radic. Biol. Med. 2002, 32, 1314–1323. [Google Scholar] [CrossRef]
- Augusto, O.; Bonini, M.G.; Amanso, A.M.; Linares, E.; Santos, C.C.; De Menezes, S.L. Nitrogen dioxide and carbonate radical anion: Two emerging radicals in biology. Free Radic. Biol. Med. 2002, 32, 841–859. [Google Scholar] [CrossRef]
- Koppenol, W.H.; Moreno, J.J.; Pryor, W.A.; Ischiropoulos, H.; Beckman, J.S. Peroxynitrite, a cloaked oxidant formed by nitric oxide and superoxide. Chem. Res. Toxicol. 1992, 5, 834–842. [Google Scholar] [CrossRef]
- Quijano, C.; Alvarez, B.; Gatti, R.M.; Augusto, O.; Radi, R. Pathways of peroxynitrite oxidation of thiol groups. Biochem. J. 1997, 322 Pt 1, 167–173. [Google Scholar] [CrossRef]
- Kissner, R.; Nauser, T.; Bugnon, P.; Lye, P.G.; Koppenol, W.H. Formation and properties of peroxynitrite as studied by laser flash photolysis, high-pressure stopped-flow technique, and pulse radiolysis. Chem. Res. Toxicol. 1997, 10, 1285–1292. [Google Scholar] [CrossRef] [PubMed]
- Radi, R. Peroxynitrite, a stealthy biological oxidant. J. Biol. Chem. 2013, 288, 26464–26472. [Google Scholar] [CrossRef] [PubMed]
- Radi, R. Oxygen radicals, nitric oxide, and peroxynitrite: Redox pathways in molecular medicine. Proc. Natl. Acad. Sci. USA 2018, 115, 5839–5848. [Google Scholar] [CrossRef] [PubMed]
- Bartesaghi, S.; Radi, R. Fundamentals on the biochemistry of peroxynitrite and protein tyrosine nitration. Redox Biol. 2018, 14, 618–625. [Google Scholar] [CrossRef]
- Halliwell, B. Free radicals and antioxidants: A personal view. Nutr. Rev. 1994, 52, 253–265. [Google Scholar] [CrossRef]
- Bianconi, E.; Piovesan, A.; Facchin, F.; Beraudi, A.; Casadei, R.; Frabetti, F.; Vitale, L.; Pelleri, M.C.; Tassani, S.; Piva, F.; et al. An estimation of the number of cells in the human body. Ann. Hum. Biol. 2013, 40, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Majima, H.J.; Oberley, T.D.; Furukawa, K.; Mattson, M.P.; Yen, H.-C.; Szweda, L.I.; St Clair, D.K. Prevention of mitochondrial injury by manganese superoxide dismutase reveals a primary mechanism for alkaline-induced cell death. J. Biol. Chem. 1998, 273, 8217–8224. [Google Scholar] [CrossRef]
- Itoh, K.; Chiba, T.; Takahashi, S.; Ishii, T.; Igarashi, K.; Katoh, Y.; Oyake, T.; Hayashi, N.; Satoh, K.; Hatayama, I.; et al. An Nrf2/small Maf heterodimer mediates the induction of phase II detoxifying enzyme genes through antioxidant response elements. Biochem. Biophys. Res. Commun. 1997, 236, 313–322. [Google Scholar] [CrossRef]
- Itoh, K.; Wakabayashi, N.; Katoh, Y.; Ishii, T.; Igarashi, K.; Engel, J.D.; Yamamoto, M. Keap1 represses nuclear activation of antioxidant responsive elements by Nrf2 through binding to the amino-terminal Neh2 domain. Genes Dev. 1999, 13, 76–86. [Google Scholar] [CrossRef]
- Gross, E.; Bedlack, R.S., Jr.; Loew, L.M. Dual-wavelength ratiometric fluorescence measurement of the membrane dipole potential. Biophys. J. 1994, 67, 208–216. [Google Scholar] [CrossRef]
- Sarkar, S.; Chattopadhyay, A. Membrane dipole potential: An emerging approach to explore membrane organization and function. J. Phys. Chem. B 2022, 126, 4415–4430. [Google Scholar] [CrossRef] [PubMed]
- Salemi, G.; Gueli, M.; D’Amelio, M.; Saia, V.; Mangiapane, P.; Aridon, P.; Ragonese, P.; Lupo, I. Blood levels of homocysteine, cysteine, glutathione, folic acid, and vitamin B12 in the acute phase of atherothrombotic stroke. Neurol. Sci. 2009, 30, 361–364. [Google Scholar] [CrossRef] [PubMed]
- Richie, J.P., Jr.; Skowronski, L.; Abraham, P.; Leutzinger, Y. Blood glutathione concentrations in a large-scale human study. Clin. Chem. 1996, 42, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.D.; Flores-Santana, W.; Switer, C.H.; Wink, D.A.; Ridnour, L.A. Impact of cell signaling processes. In Nitric Oxide Biology and Pathology, 2nd ed.; Section I. Chemical Biology, Chapter 1, Determinants of nitric oxide chemistry; Ignapro, L.J., Ed.; Academic Press: Amsterdam, The Netherlands, 2010; pp. 3–26. [Google Scholar]
- Lynch, R.E.; Fridovich, I. Permeation of the erythrocyte stroma by superoxide radical. J. Biol. Chem. 1978, 253, 4697–4699. [Google Scholar] [PubMed]
- Porcelli, A.; Ghelli, A.; Zanna, C.; Pinton, P.; Rizzuto, R.; Rugolo, M. pH difference across the outer mitochondrial membrane measured with a green fluorescent protein mutant. Biochem. Biophys. Res. Commun. 2005, 326, 799–804. [Google Scholar] [CrossRef]
- Mondal, P.; Ishigami, I.; Gérard, E.F.; Lim, C.; Yeh, S.R.; de Visser, S.P.; Wijeratne, G.B. Proton-coupled electron transfer reactivities of electronically divergent heme superoxide intermediates: A kinetic, thermodynamic, and theoretical study. Chem. Sci. 2021, 12, 8872–8883. [Google Scholar] [CrossRef]
- De Grey, A.D. HO2•: The forgotten radical. DNA Cell Biol. 2002, 21, 251–257. [Google Scholar] [CrossRef]
- Cordeiro, R.M. Reactive oxygen species at phospholipid bilayers: Distribution, mobility and permeation. Biochim. Biophys. Acta 2014, 1838 Pt B, 438–444. [Google Scholar] [CrossRef]
- Tikunov, A.; Johnson, C.B.; Pediaditakis, P.; Markevich, N.; Macdonald, J.M.; Lemasters, J.J.; Holmuhamedov, E. Closure of VDAC causes oxidative stress and accelerates the Ca2+-induced mitochondrial permeability transition in rat liver mitochondria. Arch. Biochem. Biophys. 2010, 495, 174–181. [Google Scholar] [CrossRef]

{kind=link}
| ROS or RNS | Calculated Dipole Moment/D | Permeability Coefficient/cm s−1 | |
|---|---|---|---|
| H2O | Water | 1.89 | 2.3 × 10−3 [48] |
| H2O2 | Hydrogen peroxide | 0.00 (permeable) | 6.1 × 10−3, 6.6 × 10−4 [49] |
| •OH | Hydroxyl radical | 1.67 | |
| O2•− | Superoxide | 0 | 1 × 10−6 (pH 7.3, 25 °C) [50] (7.6 + 0.3) × 10−8 [51] |
| HO2• | Hydroperoxyl radical | 2.23 | 4.9 × 10−4 [51] |
| •NO | Nitric oxide | 0.14 (permeable) | 93 (20 °C) [52] |
| •NO2 | Nitrogen dioxide | 0.35 | [53] and discussion in the text |
| ONOO− | Peroxynitrite | 2.14 | Through anion exchanger [54] 8.0 × 10−4 [55] |
| ONOOH | Peroxynitrous acid | 1.77 | 4–13 × 10−4 [56,57,58] |
| ROS or RNS | Half-Life Time | Amount/Cell | Diffusion Distance (µm) | Permeability Coefficients (Pm) (cm s−1) | Eo′; One-Electron Reduction Potential (V) at pH 7 | pKa | k (AscH–)/M−1 s−1 | k (GSH)/M−1 s−1 | |
|---|---|---|---|---|---|---|---|---|---|
| H2O | Water | –– | –– | –– | 3.3 × 10−3 (EYPC) [59] | −2.87 [60] −2.87 [61] | 15.7 [62] | –– | –– |
| O2 | Oxygen | –– | –– | –– | 12 (DMPC) [59] 125 (DMPC) [59] 114 (DOPC) [59] 157 (POPC) [59] 50 (EYPC: 30% Chol) [59] 38 (RBC human) [59] 21 (CHO cells) [59] 42 (CHO cells) [59] | −0.18 (pH 7) [60] −0.33 [61] −0.16 [63] −0.18 (pH 7, 25 °C) [64] | –– | –– | –– |
| •OH | Peroxynitrous acid | 10−9 s [65] 10−9~10−6 s (diffusion-controlled reactivity) [66] 10−10 s [67] 10−9 s (1 M, 37 °C) [68] 10−9 s [69] | –– | 3 Å [70] A large flux of hydroxyl radicals would be required to inactivate a substantial fraction of any biological target [70] 0.02 (GSH+) [71] | –– | +2.32 (pH 7) [60] +2.31 [61] +2.31 (pH 7, 25 °C) [64] +2.31 [72] +2.31 (pH 7) [73] | 11.9 [62] 11.6 [74] | 1.1 × 1010 (pH 7.4) [61] | 1.0 × 1010 [72] 1.64 ± 0.01 × 1010 [74] 1 × 109 [75] 8.8 × 109 (pH 1.0) [76] 9.0 × 109 (pH 7.6) [77] 1 × 1010 [78] 1.1 × 1010 (oxidized GSH) [79] 1.4 × 1010 (reduced GSH) [79] 1.4 ± 0.1 × 1010 (pH 7.8) [80] 4.4 ± 0.5 × 1010 (pH 10.6) [80] 2.3 × 1010 [81] 4.4 ± 0.5 × 1010 (pH 10.6) [82] |
| 1O2 | Singlet oxygen | 10−6 s [65] 10−6 s [67] 10−6 s (solvent, 37 °C) [68] 10−5 s [69] 10−9~10−6 s [83] | –– | –– | –– | +0.81 (pH 7, 25 °C) [64] | –– | 3.2 × 108 [83] 1.8 × 108 [84] | 9.39 ± 0.07 × 108 [74] |
| H2O2 | Hydrogen peroxide | Stable [65] Stable) Stable, decomposed by catalase and GSH peroxidase and by EDTA and ADP [68] Enzymatic [69] 18.1 ± 2.7 min [82] | Physiological condition (proliferation/differentiation/migration/angiogenesis): 0.001~0.1 µM) Stress responses/adaptation (e.g., NRF2): 0.05~5.0 µM [85] Inflammation/fibrogenesis/tumor growth/metastasis: 0.01~10.0 µM [85] Growth arrest/cell death: 1.0~10.0 µM [85] | 1600 (GSH+) [78] | 6 × 10−4 (RBC horse) [59] 3 × 10−3 (peroxisome rat liver) [59] 1.2 × 10−2 (RBC rat) [59] 2 × 10−4 (Jurkat T cells) [59] 3.6 × 10−4 (Chara coralina) [59] 1.6 × 10−3 (Escherichia coli) [59] 4 × 10−4 (PC12 cells) [59] 1.6 × 10−3 (HUVEC cells) [59] 1.1 × 10−3 (IMR-90 cells) [59] 4.4 × 10−4 (HeLa cells) [59] | +0.39 (pH 7) [60] +0.32 [61] +1.77 [72] +1.8 [78] +0.39 (pH 7, 25 °C) [86] | 11.6 [62] 11.75 (pH 7.2) [78] | –– | 9 × 10−1 [72] 9 × 10−1 [73] 9 × 10−1 (pH 7.4, 37 °C) [78] 8.7 × 10−1 [81] |
| O2•− | Superoxide | 10−6 s [65] 1 s (pH 10) [66] 10−6 s (diffusion-controlled reactivity) [66] 10−6 s [67] The lifetime of superoxide in a cellular environment in water would be expected to be very short, too short to permit diffusion for great distances [68] Enzymatic [69] 3000 ms (10−6 M) [87] 175 ms (10−6 M + SOD 10−9 M) [87] hours (10−9 M) [87] 175 ms (10−9 M + SOD 10−9 M) [87] 0.175 ms (10−9 M + SOD 10−6 M) [87] | 28.4 pM (normal condition)/mitochondria [88] Formation rate (to 6 µM/s) [88] MnSOD-catalyzed dismutation (k = 2 × 109 M−1 s−1) [88] 9.15 × 10−8 pmol production/s/mitochondria * 690 nM production/s/mitochondria * 5.5 × 104 superoxide molecules /s/mitochondria * | –– | 2.1 × 10−6 (SBPC) [59] 7.6 × 10−8 (EYPC) [59] | +0.94 [72] +0.94 [73] | –– | 1 × 105 (pH 7.4) [61] 2.7 × 105 (pH 7.4) [61] | ~10 to 103 [72] 2 × 102 [81] 1.1 ± 0.04 × 103 [74] 6.7 × 105 (reduced GSH) (pH 7.8) [89] |
| HO2• | Hydroperoxyl radical | 51~422 s (pH 2~10) [90] HO2• radicals in organic or lipophilic media could have a longer half-life. The half-life of superoxide cannot be calculated unless the concentrations of SOD and all reactive substrates are known [67] | 9.15 × 10−8 pmol production/s/mitochondria * 690 nM production/s/mitochondria * 5.5 × 104 superoxide molecules/s/mitochondria * | –– | 4.9 × 10−4 (EYPC) [59] | +1.05 (pH 7) [60] +1.06 [72] +1.05 (pH 7, 25 °C) [86] | 4 [62] 4.8 [89] 4.8 [90] 4.8 [91] | 1 × 105 (pH 7.4) [61] 2.7 × 105 (pH 7.4) [61] | –– |
| •NO | Nitric oxide | ms to s depending on the available concentration of O2, otherwise stable [66] Second [67] 1~10 s [69] 445 s [92] •NO:1200 nM in saline: binding with Hb: 2 × 105 M−1 s−1 [92] Seconds [93] | pM~μM [93] pM~µM in physiological milieu [94] cGMP-mediated processes; <1~30 nM [95] Akt phosphorylation; = 30~100 nM stabilization of HIF-1α; = 100~300 nM [95] phosphorylation of p53; > 400 nM [95] nitrosative stress; 1 μM [95] | –– | 73 (EYPC) [59] 66 (EYPC: 30% Chol) [59] 18 (RBC human) [59] | –0.52 (pH 7) [60] –0.35 [63] –0.80 [72] –0.80 [73] | –– | –– | Nondetectable [72] 1.0 × 101 [75] |
| •NO2 | Nitrogen dioxide | Second [67] <10 µs [96] | Typically 0.2~0.3 µM [96] | 0.4 (GSH+) [78] 0.2 in the cytoplasm [96] <0.8 in blood plasma [96] | ~5 (EYPC) [59] | +1.04 (pH 7) [60] +1.04 [63] +1.04 [72] +1.04 [73] | –– | 1.8 × 107 [96] 3.5 × 108 [96] 3.54 × 106 (pH 5.4~6.5, 55 °C) [97] | 3.0 × 107 [72] 2.2 × 107 [75] 3 × 107 [78] 2 × 107 [81] ~2 × 107 [96] |
| ONOO− | Peroxynitrite | 0.8 s (pH 7.4) [64] 10−3 s [67] 0.05~1 s [69] 0.8 s (pH 7.4) [98] 0.9 s [98] Stable [98] Relatively stable [99] Less than 1 s (pH 7.4, 37 °C) [99] 0.8 s (pH 7.4) [100] | A total peroxynitrite and peroxynitrous acid concentration that exceeds 0.1 mM [101] | 60 (GSH+) [78] 0.42 [101] | ––– | –– | –– | 7 × 102 [78] 2.35 ± 0.04 × 102, 25 °C [91] | 6.6 × 102 (pH 7.4, 25 °C) [71] 7.0 × 102 [73] 6.6 × 102 [75] 1.36 × 103 (pH 7.4, 37 °C) [78] 2.81 × 102 (pH 5.75, 37 °C) [100] |
| ONOOH | Peroxynitrous acid | Fairly stable [67] 0.90 s, 25 °C [98] Less than 1 s at physiological pH and 37 °C [99] 0.6 s; 1.13 s−1 in phosphate buffer (pH 7.4, 37 °C) [102] | A total peroxynitrite and peroxynitrous acid concentration that exceeds 0.1 mM [101] | –– | 8 × 10−4 (DMPC) [59] 1.3 × 10−3 (EYPC) [59] 6.3 × 10−4 (DMPC) [59] 4 × 10−4 (DPPC) [59] | +1.40 [72] | 6.8 [86] 6.8 [90] 6.8 [98] 6.8 [103] | –– | 6.6 × 102 [72] 1.35 × 103 [81] |
| RO• | Alkoxyl radicals | 10−6 s [67] 10−6 s (100 mM) [68] 10−6 s [69] | –– | –– | +1.60 [61] +1.60 [72] ~+1.60 [73] | –– | 1.6 × 109(pH 7.4) [61] | 2.76 ± 0.15 × 106 [74] | |
| ROO• | Peroxyl radicals | Seconds to hours depending on conditions [66] 17 s [67] 7 s (100 mM, 37 °C) [68] 7 s [69] | –– | –– | +1.00 [61] +0.77~1.44 [73] +1.00 [72] | –– | 1-2 × 106 (pH 7.4) [61] | –– | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masuda, D.; Nakanishi, I.; Ohkubo, K.; Ito, H.; Matsumoto, K.-i.; Ichikawa, H.; Chatatikun, M.; Klangbud, W.K.; Kotepui, M.; Imai, M.; et al. Mitochondria Play Essential Roles in Intracellular Protection against Oxidative Stress—Which Molecules among the ROS Generated in the Mitochondria Can Escape the Mitochondria and Contribute to Signal Activation in Cytosol? Biomolecules 2024, 14, 128. https://doi.org/10.3390/biom14010128
Masuda D, Nakanishi I, Ohkubo K, Ito H, Matsumoto K-i, Ichikawa H, Chatatikun M, Klangbud WK, Kotepui M, Imai M, et al. Mitochondria Play Essential Roles in Intracellular Protection against Oxidative Stress—Which Molecules among the ROS Generated in the Mitochondria Can Escape the Mitochondria and Contribute to Signal Activation in Cytosol? Biomolecules. 2024; 14(1):128. https://doi.org/10.3390/biom14010128
Chicago/Turabian StyleMasuda, Daisuke, Ikuo Nakanishi, Kei Ohkubo, Hiromu Ito, Ken-ichiro Matsumoto, Hiroshi Ichikawa, Moragot Chatatikun, Wiyada Kwanhian Klangbud, Manas Kotepui, Motoki Imai, and et al. 2024. "Mitochondria Play Essential Roles in Intracellular Protection against Oxidative Stress—Which Molecules among the ROS Generated in the Mitochondria Can Escape the Mitochondria and Contribute to Signal Activation in Cytosol?" Biomolecules 14, no. 1: 128. https://doi.org/10.3390/biom14010128