
Microtubule Regulation in Plants: From Morphological Development to Stress Adaptation

1
Department of Biochemistry and Metabolism, John Innes Centre, Norwich Research Park, Norwich NR4 7UH, UK
2
Cell Biology Core Lab, Institute of Plant and Microbial Biology, Academia Sinica, Taipei 115, Taiwan
*
Author to whom correspondence should be addressed.
Biomolecules 2023, 13(4), 627; https://doi.org/10.3390/biom13040627
Submission received: 16 January 2023
/
Revised: 9 March 2023
/
Accepted: 25 March 2023
/
Published: 30 March 2023
(This article belongs to the Special Issue Molecular Functions of Microtubules)
{kind=link}
{kind=link}
Abstract
:Microtubules (MTs) are essential elements of the eukaryotic cytoskeleton and are critical for various cell functions. During cell division, plant MTs form highly ordered structures, and cortical MTs guide the cell wall cellulose patterns and thus control cell size and shape. Both are important for morphological development and for adjusting plant growth and plasticity under environmental challenges for stress adaptation. Various MT regulators control the dynamics and organization of MTs in diverse cellular processes and response to developmental and environmental cues. This article summarizes the recent progress in plant MT studies from morphological development to stress responses, discusses the latest techniques applied, and encourages more research into plant MT regulation.
1. Introduction
Microtubules (MTs) are highly conserved cytoskeletal structures in both plant and mammal cells [1,2]. Like mammal MTs, plant MTs consist of α- and β-tubulin subunits [3,4], and some tubulin isoforms are expressed in specialized cells or tissues during development [5,6,7,8]. The formation of α/β-tubulin heterodimers needs a large chaperone complex and guanosine 5′-triphosphate (GTP) [9,10]. Hence, recombinant tubulins cannot be efficiently produced in Escherichia coli because of the lack of a proper protein folding machinery in prokaryotes [11]. The longitudinal head-to-tail interactions between α/β-tubulin heterodimers via GTP hydrolysis to guanosine diphosphate build up the basic units of MTs, protofilaments [12,13]. The GTP cap at the plus end ensures MT growth, while the loss of the GTP cap results in MT shrinkage. The co-existence of growing and shrinking MTs driven by the restoration and hydrolysis of GTP was proposed as a “dynamic instability” model based on the observation of in vitro-reconstituted MTs [14]. The dynamic behaviour of MTs is thought to be an intrinsic property, as demonstrated by a MT polymerization experiment conducted from purified tubulin without external factors [15].
MT polymerization can occur spontaneously in vitro without any pre-formed templates when sufficiently high concentrations of purified tubulins are warmed up in the presence of GTP [16]. However, in cells, tubulin molecules tend to form a nucleation seed for efficiently initiating polymer growth and the construction of dynamic polar MTs under spatial and temporal control [17,18]. The evolutionarily conserved MT nucleating template is known as the γ-tubulin-containing ring complex (γ-TuRC) [19,20,21]. It initiates MT nucleation at a particular subcellular location, primarily regulated by Augmin [22,23,24]. Katanin internally breaks MTs dependent on adenosine 5′-triphosphate (ATP), particularly at MT crossover positions, where the detached daughter MTs can translocate via treadmilling to form new configurations of MT arrays [25,26,27,28,29]. Thus, both γ-TuRC and katanin are thought to be central components in synthesizing new treadmilling MTs at the plant cell cortex [25,30]. The dynamic nature enables MTs to alter their organization in response to internal and external signals for the needs of the cell, and it is regulated by various proteins [31,32].
Eukaryotes have conserved MT-associated proteins (MAPs) that bind along the MT lattice and have stabilizing or destabilizing effects on MT assembly [32,33,34]. However, plants possess a set of MAPs specific to plant morphology and physiology [35,36,37]. Conventional MAPs include motor proteins such as kinesins that utilize MTs as tracks to transport cargo and structural MAPs or severing proteins such as MAP65 and katanin (with the catalytic subunit p60 and a regulatory subunit p80) involved in MT organization via binding, bundling, or cleavage of MTs. MTs plus tip-associated proteins, such as cytoplasmic linker-associated proteins (CLASPs), regulate MT dynamics via their binding and interactions at the plus-end of growing MTs [13,36,38,39,40,41,42,43].
MTs arise from centrosomes in animal cells [44], but MTs in acentrosomal plant cells are thought to self-organize into structured arrays [45]. The plant-specific structures (i.e., cell wall and stomata) and the sessile nature of land plants lead to distinct MT regulation, affecting plant growth, development, and stress adaptation. This article summarizes the latest research progress (mostly after 2018) in plant MT regulation from the cellular to organism level and discusses cell division and intracellular trafficking, morphogenesis and development, as well as stress responses. By introducing the latest technologies in studying plant MTs, we aim to encourage more research to discover the fundamental rules governing the dynamics and molecular mechanism of how cells specify the functional MT array patterns under various circumstances.
2. MTs in Plant Cell Division
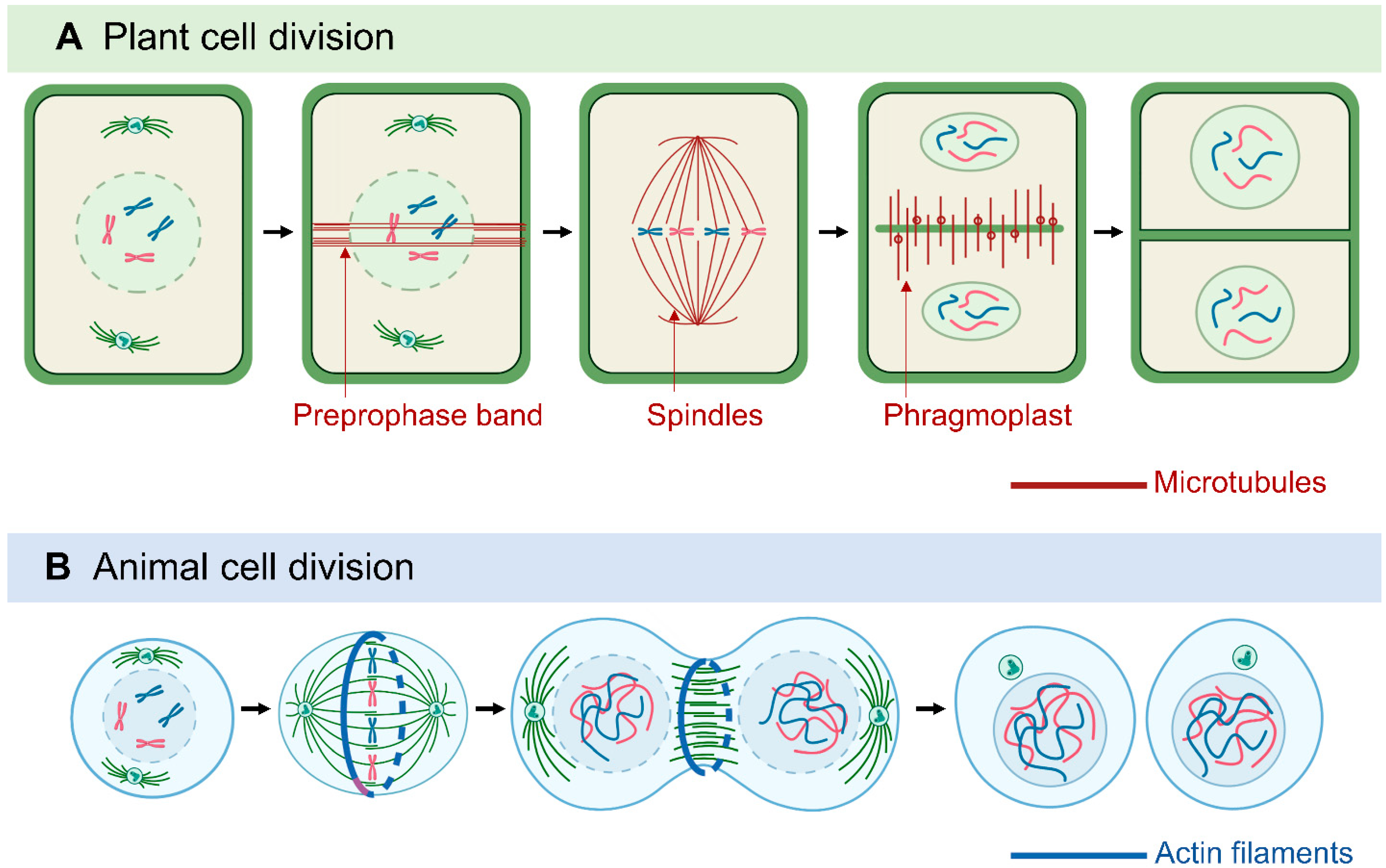
Sessile plants cannot move as quickly as multicellular animals to escape environmental challenges. Thus, besides forming organs and various cell types for morphological development, cell division in plants is also important for adaptation to environmental conditions: adjusting growth under stress by enhancing, reducing, and redirecting cell growth [46]. In dividing plant cells, MTs form distinct structures, including the preprophase band (PPB), the acentrosomal mitotic spindle, and the phragmoplast [47,48,49] (Figure 1). The PPB, which is a plant-specific cortical MT ring, marks the orientation of the cell division plane and determines the spindle positioning in metaphase [50,51,52]. The PPB tunes the orientation of spindles in a mode similar to that of centrioles and astral MTs in animal cells, which implies the importance of spindle orientation [51,53].
MT-based spindles separate chromosomes during mitosis [54]. Most animal spindle-assembly factors are well conserved in plants, but plants lack two major elements: centrosomal components and the cytoplasmic dynein complex [48]. Animal spindle MTs nucleate from centrosomes [55], whereas plant MTs appear to nucleate from the nuclear envelope surface [56]. In animals and fungi, cytoplasmic dynein is a processive minus-end-directed motor that pulls on astral MTs from the cell cortex for efficient and accurate spindle assembly and positioning [57]. In contrast, plants lack cytoplasmic dynein but contain many minus-end–directed kinesin-14 proteins, which were thought to be involved in the sliding of anti-parallel microtubules throughout mitosis [46,58,59,60]. Kinesins with a calponin homology domain (KCH) are a distinguished subclass of kinesin-14 found only in Plantae [61]. Rice OsKCH2 exhibits processive minus-end-directed motility on MTs to potentially compensate for the loss of dynein [62], whereas moss KCH drives MT-based nuclear transport reminiscent of animal dynein [63]. In animals, plus-end–directed kinesin-5 and kinesin-12 facilitate the spindle arrangement [64,65] and it seems to be conserved in plants, as shown by Arabidopsis KINESIN-12E controlling spindle MT organization and size during mitosis [66]. MTs are protein–protein interaction sites for the spindle assembly checkpoint (SAC) protein complex and signaling network. Plants have conserved the SAC network with some variations from animals and yeast [67]. The dissection of proteins associated with SAC led to the discovery of novel aspects of plant SAC regulation [68,69], which may be relevant for plant breeding studies because ploidy alternations likely rely on SAC properties.
Another plant-specific MT machinery, the phragmoplast, is formed between the reconstituting daughter nuclei at the end of telophase, as a hallmark of cytokinesis [70]. The phragmoplast expands and transports Golgi-derived vesicles containing building materials to facilitate the construction of the cell plate. The assembly, crosslinking, and turnover of phragmoplast MTs are regulated by various MAPs, kinesin motors, and regulatory enzymes [49,53,70,71,72]. Among them, plant-specific Cortical MT Disordering 4 tethers the conserved MT-severing protein katanin to facilitate phragmoplast expansion and accelerate cytokinesis [73]. Cytokinesis-specific MAP65-3 plays primary roles in phragmoplast integrity and efficient cell plate formation [74,75,76]. Phragmoplast dynamics during cytokinesis is closely related to the phosphorylation of MAP65-3, regulated by mitogen-activated protein kinase 4 and aurora kinase [77,78,79]. As a positive regulation mechanism, benzimidazole-3 proteins interact with MAP65-3 and promote MT bundling for phragmoplast expansion [80]. The MT motor protein KINESIN12 is critical for maintaining MT plus-ends in the phragmoplast midzone. Indeed, the Arabidopsis double-mutant pok1/pok2 (two kinesin-12 orthologs) revealed chaotic division sites and a slower phragmoplast expansion rate compared to the wild-type [81,82]. Overall, because they lack structurally defined centrosomes but have flexible and distributed PPB and phragmoplasts, plant cells can assemble bipolar spindles and determine the division plane with a great deal of plasticity, thereby compensating for the restraints in cell movement caused by the stiff plant cell wall [46,83].
3. Cortical MTs in Intra- and Intercellular Trafficking
Cortical MTs are tightly anchored to the plasma membrane and interact with plasmodesmata and several endomembrane systems. Therefore, they are involved in intra- and intercellular transportation [84,85]. Recent studies showed that MT-dependent homeostasis of the trans-Golgi network (TGN) is regulated by TGNap1 [86], whereas MTs and kinesins participate in vacuole trafficking and endoplasmic reticulum (ER) movement as revealed by modified transport to the vacuole proteins [87,88]. MTs are a scaffold for stromule extension, branching, and kinking [89,90] (see Section 7) and are also involved in nuclear transport to regulate tip cell growth in moss and Arabidopsis root hairs [63,91]. MTs have a role in the intercellular movement as shown by KINESIN G facilitating the cell-to-cell movement of short-root along MTs [92]. Additionally, mobile microRNAs are regulated by MTs and KATANIN1 via ARGONUATE1 of the RNA-induced silencing complex by an unknown mechanism to exit the cell [93].
4. MTs in Morphological Development
MTs are essential in cell wall formation by guiding the movement of cellulose synthase complexes on the plasma membrane. Therefore, they function as templates for the deposition of cell wall cellulose [94,95,96]. The orientation of cellulose microfibrils affects cell wall stiffness and cell growth direction, and in turn, the mechanical stress generated by the growing tissue orients the MTs, thus forming a feedback loop [97]. Perturbation of MTs often leads to changes in cell wall composition and cell stiffness, thus affecting cell expansion and plant architecture [98]. Hence, plant cell morphogenesis requires a dynamic interplay between cell expansion and MT-templated cell wall construction [37].
4.1. Seed Germination
When dormant seeds are imbibed, MTs appear as thick bundles in random orientations [99]. Dormancy release in the embryo triggers MT self-organization and alignment with tensile stress before germination and anisotropic growth [99]. Various MAPs and hormone signaling are involved in seed germination. Wave-Dampened2-Like 4 modulates auxin distribution to promote apical hook opening in Arabidopsis [100]. Additionally, the elongated hypocotyl cells create a variety of MT array patterns with differing degrees of polymer coalignment and orientation to the cell growth axis [101]. These patterns are regulated by light and ethylene signaling via Armadillo Repeat Kinesin 2 and Microtubule-Destabilizing Protein 60 [102,103].
4.2. Root
Recent studies have highlighted the importance of MTs in root morphogenesis and root hair and apical meristem growth by investigating the MAPs TANGLED 1, Auxin-induced-in-roots 9, CLASP, and Rho of Plant interactors and extensive study of KATANIN 1 [104,105,106,107]. Functioning in nutrient and water uptake, the initiation and directional growth of lateral roots are largely regulated by the dynamics of MT networks [108,109]. MAP70-5 is necessary for spatially defined MT organization and endodermis remodeling during lateral root morphogenesis [110], whereas MT reorganization via MT-stabilizing protein TPX2-LIKE5 during lateral root initiation is negatively regulated by Elongated Hypocotyl 5 [111]. Similarly, rhizoid tip growth in the basal land plant Marchantia polymorpha is controlled by the MT organization with different MAPs from flower plants: Wave-Dampened 2-Like [112] and MpNEK1 [113]. The plant hormones auxin and brassinosteroid (BR) regulate root growth via dynamic MT organization as shown by FERONIA mediating root nutation growth by affecting MT polymerization and polar auxin transport [114]. Additionally, Tetratricopeptide-repeat Thioredoxin-Like 3 interacts with MTs and participates in BR signaling during root system morphogenesis [115].
4.3. Leaf
Correct timing is needed to orchestrate MT dynamics and cell wall biogenesis during leaf development [116]. Leaf shape depends on cortical MT-mediated cellulose deposition along the adaxial–abaxial axis in the internal cell wall [117]. MTs may act as sensors or as part of sensing mechanisms for mechanical stress [118]. Mechanical stress promotes the MT response to stress by increasing KATANIN severing activity [119] and initiates and sustains the morphogenesis of wavy leaf epidermal cells [120]. Cortical MTs are thought to align with maximal tension, whereas adjustment of MT stabilization with a tension–adhesion feedback loop with disturbance of this linkage to the wall could provide “noise,” allowing for greater adaptive response to new inputs, both mechanical and developmental [121,122]. MT dynamics during pavement cell morphogenesis are regulated by BR and auxin [123,124] as well as various MAPs such as IPGA1-ANGUSTIFOLIA [125], IQ67 DOMAIN 5 (IQD5) [126], KATANIN and CLASP [127], and basic proline-rich protein [128].
4.4. Xylem
Cortical MT alignment regulates distinct deposition patterns of the secondary cell wall in xylem vessels [129]. Spatiotemporal control of MT nucleation and KATANIN-regulated MT band formation are critical for xylem wall patterning [130,131]. Xylem pit formation requires mutually exclusive plasma membrane tethering by the Microtubule Depletion Domain1–Kinesin13A complex and IQD13 for MT depolymerization and stabilization [132,133,134,135,136]. Recently, the MT regulator MAP20 was reported to be involved in metaxylem pit development [137] and Cortical MT Disordering1 for proper deposition patterns in metaxylem vessels [138].
4.5. Flower
Cortical MTs play important roles in plant cell polar growth/expansion and have been reported to guide the growth and shape of sepals and petals [139]. MAPs were found to modulate cortical MT arrangement in flower organ growth and fertility: Increased Petal Growth Anisotropy 1, QWRF1, and QWRF2 affect MT organization and stability to regulate petal growth anisotropy [140,141], and KATANIN 1 mediates MT organization to regulate petal conical cell shape [142,143] and the pollen tube path [144]. KATANIN 1-mediated MT severing is critical for stamen filament development, as shown by the crosstalk between MT severing and BR signaling [145].
4.6. Fruit, Grain, and Seed
Cortical MT arrays play a critical role in plant cell shape determination [146]. Controlling the shape and architecture of a plant could help improve agricultural yields. The plant-specific IQD family emerged as regulators of fruit shape and grain size in tomatoes [147] and rice [148]. By interacting with MT arrays, TONNEAU 1-recruiting motif (TRM) protein Grain Weight 7 controls grain dimension traits in wheat [149], whereas Reducing Plant Height 1 controls plant and ear height in maize [150]. The plant seed coat protects the embryo and transmits information regarding the external environment [151]. TRM4 and IQD9 maintain cortical MT organization to orchestrate cellulose patterning in Arabidopsis seed mucilage [152,153]. MTs are important for the development and diameter determination of cotton fiber [154], an extension of seed coat [155], as shown by a recent finding that GhMAP20L5 is involved in cotton fiber elongation by interacting with GhTUB13 [156].
5. MTs in Abiotic Stress Responses
5.1. Temperature
Sessile plants have evolved specific mechanisms to perceive environmental stresses and generate appropriate responses. The plant cytoskeleton plays central roles in stress-induced signaling pathways, either as a direct target or a signal transducer [157]. Cell wall remodeling and MT rearrangements are essential for plants to adapt to growth and development in response to environmental stresses [158]. Temperature stresses such as coldness and heat restrict plant growth and development and thus affect crop production. In grapevine, cold-induced disassembly of MTs is regulated by a key transcription factor, Cold Box Factor 4 [159], and MTs were proposed to serve as modulators of cold sensitivity by mediating the transportation of cold signaling vesicles into the plasma membrane [160,161]. Rice MT motor Dual Localisation Kinesin enters the nucleus under cold stress, but whether MTs are involved in cold-induced transcriptional regulation needs further investigation [162]. Recent studies showed that phospholipase D δ regulated reactive oxygen species-mediated MT organization and stomatal movement upon heat shock, and heat shock protein 70-3 bound to MTs and interacted with phospholipase D δ to stabilize cortical MTs upon heat stress [163,164], which suggests the importance of controlling MT dynamics in the heat stress response.
5.2. Salinity
At the cellular level, salinity stress induces rapid depolymerization of cortical MTs and further reassembly into altered MT networks [165,166]. Companion of cellulose synthase (CC) proteins control MT reassembly and support cellulose synthase activity to sustain plant growth under saline conditions [167,168]. A recent proteomic analysis revealed the upregulation of MT proteins (TUB3, TUB4, and TUB9) in salt-adapted Arabidopsis. Loss of function of TUB4 enhanced salinity tolerance, and mutation in TUB9 caused hypersensitivity [169], but the exact mechanism still awaits discovery. Various MAPs play a significant role in mediating dynamic MT changes under salinity; these include Arabidopsis KATANIN 1 [170], MAP65 [171], SPIRAL 2-LIKE [172], and rice microtubule-associated RING finger protein 1 [173]. Moreover, ethylene signaling regulates MT reassembly in response to salt stress in Arabidopsis [174]. Analysis of the rice OsTUB1–Kinesin13A complex revealed the function of tubulin and kinesin in regulating MT organization and ionic homeostasis to increase the survival of rice plants under salt stress, thus providing novel genes for salt-insensitive rice breeding in areas with high soil salinity [175].
5.3. Drought
The precise regulation of stomatal movement is critical for plant adaptation to drought stress [176]. The disassembly of MTs is critical for stomatal closure in response to abscisic acid (ABA) and drought stress [177]. E3 ligases were recently reported to participate in ABA-mediated MT depolymerization, stomatal closure, and the drought stress response; these ligases include MT-related E3 ligase 57 [178] and JAV1-associated ubiquitin ligase 1 [179]. SUN-interacting nuclear envelope protein 1 (SINE1) and SINE2 as well as SPIRAL1 were also found involved in MT reorganization during ABA-induced stomatal closure [180,181], and tubulin perturbation affected guard cell behavior, delaying drought-induced stomatal closure and clustering [182,183].
6. Protein Disorder Regulates MTs
Intrinsically disordered proteins (IDPs) are a group of functional proteins without defined 3D structures. They can adopt various conformations when binding to different partners and serve as interaction specialists in cellular signaling and the regulation of macromolecular machine assembly [184,185,186]. The roles of IDPs in regulating adaptive responses and the dynamics of the cytoskeletal systems have been summarized in animal cells [187]. Both MT-binding MAP2 and Tau proteins are intrinsically disordered [188]. The disordered regions of Tau are important for MT-binding and pathological aggregation, which cause neurodegeneration in Alzheimer’s disease and related dementias [189]. In plants, Arabidopsis CC1 controls MT bundling and dynamics to sustain plant growth under salt stress via its intrinsically disordered N-terminus through a Tau-like mechanism [168]. Recently, a rice disordered repetitive proline-rich protein was found as a novel mechanism of a plant IDP controlling highly ordered actin filaments and MTs to adapt root growth under water deficit [190]. Even though IDPs can control MT dynamics in both mammals and plants, the results are very different: neuropathology in mammals and stress adaptation in plants. In comparison to well-understood mammal protein disorders in MT regulation and function in neurodegenerative diseases, plant research about how protein disorders regulate MTs is lagging. Given that the disordered nature of IDPs plays regulatory roles in highly ordered MTs, this field needs more attention.
7. MTs in Biotic Stress Responses
Upon pathogen attack, the plant cytoskeleton undergoes rapid remodeling to coordinate the movement of intracellular organelles and the formation of immune microdomain complexes as well as the transportation of defense compounds and the turnover of recognizing receptors [191]. Although actin remodeling is thought to play an essential role during plant innate immunity, recent studies have highlighted that immunity-related protein secretion and cell wall-based defense depend on MT-based transport [192]. The role of MTs in intracellular trafficking and secretory pathways may have a positive or negative impact on plant–pathogen interactions. Cortical MT-associated ER sites are essential for the replication and cell-to-cell spread of the tobacco mosaic virus [193]. Chloroplast stromules facilitate the transport of pro-defense signals into the nucleus during innate immunity [194], and stromule extensions through the ER are directed by MTs [195,196], which implies the importance of MTs in chloroplast–nucleus communication. Additionally, MTs but not actin cytoskeleton exclusively control the dynamics of aquaporin AtPIP2;1 during stomatal closure in response to flg22 [197], which suggests the unique function of MTs rather than actin filaments.
7.1. Plant–Pathogen Interaction
Pathogens produce effectors targeting the cytoskeleton to achieve pathogenicity [191]. By interacting with MAPs, pathogens can control the MT network in the host cells [195,198,199]. For example, the Pseudomonas syringae pv. tomato (Pto) effector HopE1 dissociates MAP65-1 from MTs to inhibit protein secretion and cell wall-based extracellular immunity to benefit pathogen Pto growth [199]. MAPs are targets of bacterial pathogen effectors, and other pathogens also co-opt MAPs for facilitating pathogenic penetration into host cells [200]. The effector protein ROP-Interactive Peptide 1 of the powdery mildew fungus Blumeria graminis f.sp. hordei (Bgh) is involved in Bgh virulence and also destabilizes cortical MTs in barley cells [201]. The nematode effector GpSPRY-414-2 targets MTs to facilitate infection [202]. By targeting MAPs, pathogens may weaken the host defense responses regulated by MTs, such as the delivery of antimicrobial molecules to the sites of infection, and thus increase pathogen growth/proliferation [203]. However, the anisotropic patterning of cortical MTs is required for regulating immunity-related genes in distal cells, as revealed by cortical MT-mediated mechanotransduction of pathogen-derived cues facilitating disease resistance caused by the fungus Sclerotinia sclerotiorum [204]. Figure 2 shows examples of MT network changes in response to environmental stresses.
7.2. Symbiosis
Besides non-beneficial plant–pathogen interactions, MTs also function in beneficial symbiosis processes between plants and symbiotic microorganisms [205]. Active cytoskeletal rearrangements are required at different stages of nodule development, from root-hair curling for entrapment of rhizobia to infection thread growth [206,207,208]. Endoplasmic MTs appear to support infection thread growth, infection droplet formation, and bacterial release into the host cytoplasm in nodules of legume species, whereas irregular cortical MT arrangements provide a possibility for isodiametric cell growth that allows a notable increase in cell size for hosting numerous symbiosomes [209]. The reorganization of MT networks during symbiosis is regulated by MAPs. In Medicago, Developmentally regulated plasma membrane polypeptides trigger MT fragmentation in symbiosis-specific membrane nanodomains to establish symbiotic associations [210]. TPXL and MAP65, together with AURORA 1, form a mitotic module to regulate MT functions for supporting the infection-thread formation of rhizobia during legume endosymbiosis [211]. Rhizobia hijack a plant-specific kinesin motor to crosslink MTs with actin filaments for controlling central vacuole formation to achieve symbiosome development and nitrogen fixation [212]. Substantial MT remodeling has been observed in arbuscule-containing cells [213,214]. A recent report revealed that a Solanaceae-specific MAP, tomato similar to SB401, is involved in MT bundling and rearrangements occurring in arbuscule-containing cells, which are required for proper arbuscule development and activity [215].
8. Key Questions in MT Regulation
Several key questions in plant MT regulation remain to be answered. Compared to fungi and animals, factors that regulate spindle MT remodeling in anaphase are largely unknown in plants. What are the novel regulatory proteins and their fine-tuned features associated with MT dynamics during cell division as well as the impact on plant polyploidization? Plants perceive various intrinsic and extrinsic mechanical stress signals during developmental processes. How is the stress in the cell wall transferred into cortical MTs, and what is the role of MTs as a stress sensor in the regulation of plant cell growth? When plants encounter environmental challenges, MTs are involved in various abiotic stress tolerance responses. How do IDPs regulate plant MTs for stress tolerance? What are the roles of MTs in stress sensing and signaling and which MAPs participate in stress adaptation?
9. New Techniques for Studying Plant MTs
To answer the aforementioned key questions, here we discuss several examples of new technology developed recently for plant MT research in silico, in vitro, and in vivo.
9.1. In Silico
Studies describing cytoskeleton dynamics rely on qualitative/quantitative analyses of cytoskeleton images. A new filament segregation algorithm, Implicit Laplacian of Enhanced Edge (ILEE), was created to provide accurate and robust analyses of cytoskeleton 2D and 3D images, thus eliminating the traditional limitation and user bias of the approach involving manual global thresholding. Indeed, ILEE can process plant MT images with satisfying performance [216]. Because cortical MT organization is a determining factor for division plane formation [53], a computer simulation method was developed to capture the effects of cell geometry on MT organization. Additionally, this method describes the stabilization of MTs at cell edges and includes the effects of polar auxin signaling on local MT stability to improve division plane prediction [217].
9.2. In Vitro
In vitro studies are essential for the biochemical characterization of interactions between various MT regulators and MTs. Even though eukaryotic tubulin proteins are highly conserved and the assembly and disassembly mechanisms of MT are thought to be similar, the kinetics of MT dynamic instability differs for animal and plant tubulins [218]. Interactions with MT-interacting proteins may differ for tubulins isolated from different organisms [219]. Thus, plant tubulin but not porcine brain tubulin must be used for in vitro studies of plant MT regulation. A TOG-based column (named after the human MT regulator tumor overexpressed gene [TOG]) was used to purify assembly-competent tubulin from fungal, animal, and algae sources [220]. The breakthrough technology also allowed for purifying wild-type and recombinant functional tubulin from Arabidopsis and tobacco [221,222]. This will facilitate in vitro functional studies of tubulin mutants, tubulin posttranslational modifications, and interactions with various plant MT regulators and motor molecules, for example [221].
9.3. In Vivo
Genetically encoded MT markers in the dicot model plant Arabidopsis have been well developed [223]. However, cytoskeleton fluorescent marker lines were recently generated in the monocot model plant rice [224]. In contrast to Arabidopsis and maize [150,223], the use of tubulin subunits cannot label MTs well in rice [224]. Conversely, the use of the mammalian MAP4 [225] in the mScarlet-MAP4 rice line allowed for live-imaging observation of MT dynamics during pollen development, root cell division, salt response, and cytoskeletal mutant characterization [224]. This research not only provides a valuable resource for studying the rice cytoskeleton but also demonstrates that species-specific MT marker design is necessary, especially for crop plants.
10. Conclusions
Recent progress in plant MT studies has highlighted the importance of MTs in basic plant biology research. The phenotypic diversity of plant organs, regulated by MTs via cell division and wall patterning, is critical for the successful marketing of a wide array of foods such as fruits, vegetables, seeds (grains), leaves, and tubers [226]. How plants use MT regulation to respond to multiple environmental signals to change their growth and development is an area of future research that could help make crop plants more resistant to climate change. With the discovery of more MAPs and the development of new techniques, our understanding of the mechanism of plant MT regulation will facilitate sustainable agriculture.
Author Contributions
A.-S.H. developed the concept and wrote the manuscript. J.-Y.H. prepared figures. All authors have read and agreed to the published version of the manuscript.
Funding
A.-S.H. was supported by a John Innes Centre ISF grant (CX593W13A) led by Michael Webster.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
This study did not generate any new data.
Acknowledgments
Figures were created with www.BioRender.com, accessed on 16 January 2023.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Paluh, J.L.; Killilea, A.N.; Detrich, H.W., 3rd; Downing, K.H. Meiosis-specific failure of cell cycle progression in fission yeast by mutation of a conserved beta-tubulin residue. Mol. Biol. Cell 2004, 15, 1160–1171. [Google Scholar] [CrossRef] [PubMed]
- Rayevsky, A.V.; Sharifi, M.; Samofalova, D.A.; Karpov, P.A.; Blume, Y.B. Structural and functional features of lysine acetylation of plant and animal tubulins. Cell Biol. Int. 2019, 43, 1040–1048. [Google Scholar] [CrossRef] [PubMed]
- Mohri, H. Amino-acid composition of “Tubulin” constituting microtubules of sperm flagella. Nature 1968, 217, 1053–1054. [Google Scholar] [CrossRef] [PubMed]
- Little, M.; Seehaus, T. Comparative analysis of tubulin sequences. Comp. Biochem. Physiol. B 1988, 90, 655–670. [Google Scholar] [CrossRef]
- Kopczak, S.D.; Haas, N.A.; Hussey, P.J.; Silflow, C.D.; Snustad, D.P. The small genome of Arabidopsis contains at least six expressed α-tubulin genes. Plant Cell 1992, 4, 539–547. [Google Scholar]
- Snustad, D.P.; Haas, N.A.; Kopczak, S.D.; Silflow, C.D. The small genome of Arabidopsis contains at least nine expressed β-tubulin genes. Plant Cell 1992, 4, 549–556. [Google Scholar]
- Breviario, D.; Giani, S.; Morello, L. Multiple tubulins: Evolutionary aspects and biological implications. Plant J. 2013, 75, 202–221. [Google Scholar] [CrossRef]
- Hashimoto, T. Dissecting the cellular functions of plant microtubules using mutant tubulins. Cytoskeleton 2013, 70, 191–200. [Google Scholar] [CrossRef]
- Steinborn, K.; Maulbetsch, C.; Priester, B.; Trautmann, S.; Pacher, T.; Geiges, B.; Küttner, F.; Lepiniec, L.; Stierhof, Y.D.; Schwarz, H.; et al. The Arabidopsis PILZ group genes encode tubulin-folding cofactor orthologs required for cell division but not cell growth. Genes Dev. 2002, 16, 959–971. [Google Scholar] [CrossRef] [Green Version]
- Szymanski, D. Tubulin folding cofactors: Half a dozen for a dimer. Curr. Biol. 2002, 12, R767–R769. [Google Scholar] [CrossRef] [Green Version]
- Lewis, S.A.; Tian, G.; Cowan, N.J. The α- and β-tubulin folding pathways. Trends Cell Biol. 1997, 7, 479–485. [Google Scholar] [CrossRef]
- Müller-Reichert, T.; Chrétien, D.; Severin, F.; Hyman, A.A. Structural changes at microtubule ends accompanying GTP hydrolysis: Information from a slowly hydrolyzable analogue of GTP, guanylyl (alpha, beta)methylenediphosphonate. Proc. Natl. Acad. Sci. USA 1998, 95, 3661–3666. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, T. Microtubules in plants. Arab. Book 2015, 13, e0179. [Google Scholar] [CrossRef] [Green Version]
- Mitchison, T.; Kirschner, M. Dynamic instability of microtubule growth. Nature 1984, 312, 237–242. [Google Scholar] [CrossRef]
- Cassimeris, L. Regulation of microtubule dynamic instability. Cell Motil. Cytoskelet. 1993, 26, 275–281. [Google Scholar] [CrossRef]
- Carlier, M.F.; Pantaloni, D. Kinetic analysis of cooperativity in tubulin polymerization in the presence of guanosine di- or triphosphate nucleotides. Biochemistry 1978, 17, 1908–1915. [Google Scholar] [CrossRef]
- Job, D.; Valiron, O.; Oakley, B. Microtubule nucleation. Curr. Opin. Cell Biol. 2003, 15, 111–117. [Google Scholar] [CrossRef]
- Hashimoto, T. A ring for all: G-tubulin-containing nucleation complexes in acentrosomal plant microtubule arrays. Curr. Opin. Plant Biol. 2013, 16, 698–703. [Google Scholar] [CrossRef]
- Teixido-Travesa, N.; Roig, J.; Luders, J. The where, when and how of microtubule nucleation—One ring to rule them all. J. Cell Sci. 2012, 125, 4445–4456. [Google Scholar] [CrossRef] [Green Version]
- Lin, T.C.; Neuner, A.; Schiebel, E. Targeting of γ-tubulin complexes to microtubule organizing centers: Conservation and divergence. Trends Cell Biol. 2015, 25, 296–307. [Google Scholar] [CrossRef]
- Roostalu, J.; Surrey, T. Microtubule nucleation: Beyond the template. Nat. Rev. Mol. Cell Biol. 2017, 18, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Kollman, J.M.; Polka, J.K.; Zelter, A.; Davis, T.N.; Agard, D.A. Microtubule nucleating gamma-TuSC assembles structures with 13-fold microtubule-like symmetry. Nature 2010, 466, 879–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, C.-M.K.; Hotta, T.; Kong, Z.; Zeng, C.J.T.; Sun, J.; Lee, Y.-R.J.; Liu, B. Augmin plays a critical role in organizing the spindle and phragmoplast microtubule arrays in Arabidopsis. Plant Cell 2011, 23, 2606–2618. [Google Scholar] [PubMed] [Green Version]
- Nakaoka, Y.; Miki, T.; Fujioka, R.; Uehara, R.; Tomioka, A.; Obuse, C.; Kubo, M.; Hiwatashi, Y.; Goshima, G. An inducible RNA interference system in Physcomitrella patens reveals a dominant role of augmin in phragmoplast microtubule generation. Plant Cell 2012, 24, 1478–1493. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, M.; Ehrhardt, D.W.; Hashimoto, T. Microtubule and katanin-dependent dynamics of microtubule nucleation complexes in the acentrosomal Arabidopsis cortical array. Nat. Cell Biol. 2010, 12, 1064–1070. [Google Scholar] [CrossRef]
- Lindeboom, J.J.; Nakamura, M.; Hibbel, A.; Shundyak, K.; Gutierrez, R.; Ketelaar, T.; Emons, A.M.; Mulder, B.M.; Kirik, V.; Ehrhardt, D.W. A mechanism for reorientation of cortical microtubule arrays driven by microtubule severing. Science 2013, 342, 1245533. [Google Scholar] [CrossRef]
- Wightman, R.; Chomicki, G.; Kumar, M.; Carr, P.; Turner, S.R. SPIRAL2 determines plant microtubule organization by modulating microtubule severing. Curr. Biol. 2013, 23, 1902–1907. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Fishel, E.; Bertroche, T.; Dixit, R. Microtubule severing at crossover sites by katanin generates ordered cortical microtubule arrays in Arabidopsis. Curr. Biol. 2013, 23, 2191–2195. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Liu, W.; Wang, G.; Li, J.; Dong, L.; Han, L.; Wang, Q.; Tian, J.; Yu, Y.; Gao, C.; et al. KTN80 confers precision to microtubule severing by specific targeting of katanin complexes in plant cells. EMBO J. 2017, 36, 3435–3447. [Google Scholar] [CrossRef]
- Ehrhardt, D.W.; Shaw, S.L. Microtubule dynamics and organization of the plant cortical array. Annu. Rev. Plant Biol. 2006, 57, 859–875. [Google Scholar] [CrossRef]
- Horio, T.; Murata, T. The role of dynamic instability in microtubule organization. Front. Plant Sci. 2014, 5, 511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodson, H.V.; Jonasson, E.M. Microtubules and Microtubule-Associated Proteins. Cold Spring Harb. Perspect. Biol. 2018, 10, a022608. [Google Scholar] [CrossRef]
- Bodakuntla, S.; Jijumon, A.S.; Villablanca, C.; Gonzalez-Billault, C.; Janke, C. Microtubule-Associated Proteins: Structuring the Cytoskeleton. Trends Cell Biol. 2019, 29, 804–819. [Google Scholar] [CrossRef] [PubMed]
- Gottschalk, A.C.; Hefti, M.M. The evolution of microtubule associated proteins—A reference proteomic perspective. BMC Genomics. 2022, 23, 266. [Google Scholar] [CrossRef] [PubMed]
- Buschmann, H.; Lloyd, C.W. Arabidopsis mutants and the network of microtubule-associated functions. Mol. Plant 2008, 1, 888–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardiner, J. The evolution and diversification of plant microtubule associated proteins. Plant J. 2013, 75, 219–229. [Google Scholar] [CrossRef]
- Elliott, A.; Shaw, S.L. Update: Plant cortical microtubule arrays. Plant Physiol. 2018, 176, 94–105. [Google Scholar] [CrossRef] [Green Version]
- Lloyd, C.; Hussey, P. Microtubule-associated proteins in plants—Why we need a map. Nat. Rev. Mol. Cell Biol. 2001, 2, 40–47. [Google Scholar] [CrossRef]
- Akhmanova, A.; Steinmetz, M.O. Tracking the ends: A dynamic protein network controls the fate of microtubule tips. Nat. Rev. Mol. Cell Biol. 2008, 9, 309–322. [Google Scholar] [CrossRef]
- Sedbrook, J.C.; Kaloriti, D. Microtubules, MAPs and plant directional cell expansion. Trends Plant Sci. 2008, 13, 303–310. [Google Scholar] [CrossRef]
- Hamada, T. Microtubule organization and microtubule-associated proteins in plant cells. Int. Rev. Cell Mol. Biol. 2014, 312, 1–52. [Google Scholar]
- Li, S.; Sun, T.; Ren, H. The functions of the cytoskeleton and associated proteins during mitosis and cytokinesis in plant cells. Front. Plant Sci. 2015, 6, 282. [Google Scholar] [CrossRef] [Green Version]
- Krtková, J.; Benáková, M.; Schwarzerová, K. Multifunctional microtubule-associated proteins in plants. Front. Plant Sci. 2016, 7, 474. [Google Scholar] [CrossRef] [Green Version]
- Desai, A.; Mitchison, T.J. Microtubule polymerization dynamics. Annu. Rev. Cell Dev. Biol. 1997, 13, 83–117. [Google Scholar] [CrossRef] [Green Version]
- Dixit, R.; Cyr, R. The cortical microtubule array: From dynamics to organization. Plant Cell 2004, 16, 2546–2552. [Google Scholar] [CrossRef] [Green Version]
- Motta, M.R.; Schnittger, A. A microtubule perspective on plant cell division. Curr. Biol. 2021, 31, R547–R552. [Google Scholar] [CrossRef]
- Pickett-Heaps, J.D.; Northcote, D.H. Organization of microtubules and endoplasmic reticulum during mitosis and cytokinesis in wheat meristems. J. Cell Sci. 1966, 1, 109–120. [Google Scholar] [CrossRef]
- Yamada, M.; Goshima, G. Mitotic spindle assembly in land plants: Molecules and mechanisms. Biology 2017, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Smertenko, A. Phragmoplast expansion: The four-stroke engine that powers plant cytokinesis. Curr. Opin. Plant Biol. 2018, 46, 130–137. [Google Scholar] [CrossRef]
- Marcus, A.I.; Dixit, R.; Cyr, R.J. Narrowing of the preprophase microtubule band is not required for cell division plane determination in cultured plant cells. Protoplasma 2005, 226, 169–174. [Google Scholar] [CrossRef]
- Schaefer, E.; Belcram, K.; Uyttewaal, M.; Duroc, Y.; Goussot, M.; Legland, D.; Laruelle, E.; de Tauzia-Moreau, M.L.; Pastuglia, M.; Bouchez, D. The preprophase band of microtubules controls the robustness of division orientation in plants. Science 2017, 356, 186–189. [Google Scholar] [CrossRef] [PubMed]
- Vavrdová, T.; Samaj, J.; Komis, G. Phosphorylation of plant microtubule-associated proteins during cell division. Front. Plant Sci. 2019, 10, 238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livanos, P.; Müller, S. Division plane establishment and cytokinesis. Annu. Rev. Plant Biol. 2019, 70, 239–267. [Google Scholar] [CrossRef] [PubMed]
- Gadde, S.; Heald, R. Mechanisms and molecules of the mitotic spindle. Curr. Biol. 2004, 14, R797–R805. [Google Scholar] [CrossRef] [Green Version]
- Conduit, P.T.; Richens, J.H.; Wainman, A.; Holder, J.; Vicente, C.C.; Pratt, M.B.; Dix, C.I.; Novak, Z.A.; Dobbie, I.M.; Schermelleh, L.; et al. A molecular mechanism of mitotic centrosome assembly in drosophila. eLife 2014, 3, e03399. [Google Scholar] [CrossRef]
- Bannigan, A.; Lizotte-Waniewski, M.; Riley, M.; Baskin, T.I. Emerging molecular mechanisms that power and regulate the anastral mitotic spindle of flowering pants. Cell Motil. Cytoskelet. 2008, 65, 1–11. [Google Scholar] [CrossRef]
- Gassmann, R. Dynein at the kinetochore. J. Cell Sci. 2023, 136, jcs220269. [Google Scholar] [CrossRef]
- Reddy, A.S.; Day, I.S. Kinesins in the Arabidopsis genome: A comparative analysis among eukaryotes. BMC Genomics. 2001, 2, 2. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Ho, C.M.; Kong, Z.; Lee, Y.R.; Qian, Q.; Liu, B. Evaluating the microtubule cytoskeleton and its interacting proteins in monocots by mining the rice genome. Ann. Bot. 2009, 103, 387–402. [Google Scholar] [CrossRef] [Green Version]
- Lucas, J.; Geisler, M. Sequential loss of dynein sequences precedes complete loss in land plants. Plant Physiol. 2022, 189, 1237–1240. [Google Scholar] [CrossRef]
- Lee, Y.R.; Liu, B. Cytoskeletal motors in Arabidopsis. Sixty-one kinesins and seventeen myosins. Plant Physiol. 2004, 136, 3877–3883. [Google Scholar] [CrossRef] [Green Version]
- Tseng, K.F.; Wang, P.; Lee, Y.J.; Bowen, J.; Gicking, A.M.; Guo, L.; Liu, B.; Qiu, W. The preprophase band-associated kinesin-14 OsKCH2 is a processive minus-end-directed microtubule motor. Nat. Commun. 2018, 9, 1067. [Google Scholar] [CrossRef] [Green Version]
- Yamada, M.; Goshima, G. The KCH kinesin drives nuclear transport and cytoskeletal coalescence to promote tip cell growth in Physcomitrella patens. Plant Cell 2018, 30, 1496–1510. [Google Scholar] [CrossRef] [Green Version]
- Ferenz, N.P.; Gable, A.; Wadsworth, P. Mitotic functions of kinesin-5. Semin. Cell Dev. Biol. 2010, 21, 255–259. [Google Scholar] [CrossRef] [Green Version]
- Hancock, W.O. Mitotic kinesins: A reason to delve into kinesin-12. Curr. Biol. 2014, 24, R968–R970. [Google Scholar] [CrossRef] [Green Version]
- Herrmann, A.; Livanos, P.; Zimmermann, S.; Berendzen, K.; Rohr, L.; Lipka, E.; Müller, S. KINESIN-12E regulates metaphase spindle flux and helps control spindle size in Arabidopsis. Plant Cell 2021, 33, 27–43. [Google Scholar] [CrossRef]
- Komaki, S.; Schnittger, A. The spindle checkpoint in plants-a green variation over a conserved theme? Curr. Opin. Plant Biol. 2016, 34, 84–91. [Google Scholar] [CrossRef]
- Komaki, S.; Schnittger, A. The spindle assembly checkpoint in Arabidopsis is rapidly shut off during severe stress. Dev. Cell 2017, 43, 172–185. [Google Scholar] [CrossRef] [Green Version]
- Su, H.; Liu, Y.; Wang, C.; Liu, Y.; Feng, C.; Sun, Y.; Yuan, J.; Birchler, J.A.; Han, F. Knl1 participates in spindle assembly checkpoint signaling in maize. Proc. Natl. Acad. Sci. USA 2021, 118, e2022357118. [Google Scholar] [CrossRef]
- Smertenko, A.; Hewitt, S.L.; Jacques, C.N.; Kacprzyk, R.; Liu, Y.; Marcec, M.J.; Moyo, L.; Ogden, A.; Oung, H.M.; Schmidt, S.; et al. Phragmoplast microtubule dynamics—A game of zones. J. Cell Sci. 2018, 131, jcs203331. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Liu, X.; Zhang, W.; Li, J.; Liu, H.; Yang, L.; Lei, P.; Zhang, H.; Yu, F. MOR1/MAP215 acts synergistically with katanin to control cell division and anisotropic cell elongation in Arabidopsis. Plant Cell 2022, 34, 3006–3027. [Google Scholar] [CrossRef] [PubMed]
- Mills, A.M.; Morris, V.H.; Rasmussen, C.G. The localization of PHRAGMOPLAST ORIENTING KINESIN1 at the division site depends on the microtubule-binding proteins TANGLED1 and AUXIN-INDUCED IN ROOT CULTURES9 in Arabidopsis. Plant Cell 2022, 34, 4583–4599. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, T.; Tsutsumi, M.; Otomo, K.; Murata, T.; Yagi, N.; Nakamura, M.; Nemoto, T.; Hasebe, M.; Oda, Y. A novel katanin-tethering machinery accelerates cytokinesis. Curr. Biol. 2019, 29, 4060–4070. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.M.; Hotta, T.; Guo, F.; Roberson, R.W.; Lee, Y.R.; Liu, B. Interaction of antiparallel microtubules in the phragmoplast is mediated by the microtubule-associated protein MAP65-3 in Arabidopsis. Plant Cell 2011, 23, 2909–2923. [Google Scholar] [CrossRef] [Green Version]
- Ho, C.M.; Lee, Y.R.; Kiyama, L.D.; Dinesh-Kumar, S.P.; Liu, B. Arabidopsis microtubule-associated protein MAP65-3 cross-links antiparallel microtubules toward their plus ends in the phragmoplast via its distinct c-terminal microtubule binding domain. Plant Cell 2012, 24, 2071–2085. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Xiao, Y.; Song, Y.; Gan, C.; Deng, X.; Wang, P.; Liu, J.; Jiang, Z.; Peng, L.; Zhou, D.; et al. Rice microtubule-associated protein OsMAP65-3.1, but not OsMAP65-3.2, plays a critical role in phragmoplast microtubule organization in cytokinesis. Front. Plant Sci. 2022, 13, 1030247. [Google Scholar] [CrossRef]
- Beck, M.; Komis, G.; Müller, J.; Menzel, D.; Samaj, J. Arabidopsis homologs of nucleus- and phragmoplast-localized kinase 2 and 3 and mitogen-activated protein kinase 4 are essential for microtubule organization. Plant Cell 2010, 22, 755–771. [Google Scholar] [CrossRef] [Green Version]
- Beck, M.; Komis, G.; Ziemann, A.; Menzel, D.; Šamaj, J. Mitogen-activated protein kinase 4 is involved in the regulation of mitotic and cytokinetic microtubule transitions in Arabidopsis thaliana. New Phytol. 2011, 189, 1069–1083. [Google Scholar] [CrossRef]
- Boruc, J.; Weimer, A.K.; Stoppin-Mellet, V.; Mylle, E.; Kosetsu, K.; Cedeño, C.; Jaquinod, M.; Njo, M.; De Milde, L.; Tompa, P.; et al. Phosphorylation of MAP65-1 by Arabidopsis aurora kinases is required for efficient cell cycle progression. Plant Physiol. 2017, 173, 582–599. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Deng, X.; Sun, B.; Lee Van, S.; Kang, Z.; Lin, H.; Lee, Y.J.; Liu, B. Role of the BUB3 protein in phragmoplast microtubule reorganization during cytokinesis. Nat. Plants 2018, 4, 485–494. [Google Scholar] [CrossRef]
- Lee, Y.R.; Li, Y.; Liu, B. Two Arabidopsis phragmoplast-associated kinesins play a critical role in cytokinesis during male gametogenesis. Plant Cell 2007, 19, 2595–2605. [Google Scholar] [CrossRef] [Green Version]
- Herrmann, A.; Livanos, P.; Lipka, E.; Gadeyne, A.; Hauser, M.T.; Van Damme, D.; Müller, S. Dual localized kinesin-12 POK2 plays multiple roles during cell division and interacts with MAP65-3. EMBO Rep. 2018, 19, e46085. [Google Scholar] [CrossRef]
- Liu, B.; Lee, Y.J. Spindle assembly and mitosis in plants. Annu. Rev. Plant Biol. 2022, 73, 227–254. [Google Scholar] [CrossRef]
- Oda, Y. Emerging roles of cortical microtubule-membrane interactions. J. Plant Res. 2018, 131, 5–14. [Google Scholar] [CrossRef]
- Christensen, J.R.; Reck-Peterson, S.L. Hitchhiking across kingdoms: Cotransport of cargos in fungal, animal, and plant cells. Annu. Rev. Cell Dev. Biol. 2022, 38, 155–178. [Google Scholar] [CrossRef]
- Renna, L.; Stefano, G.; Slabaugh, E.; Wormsbaecher, C.; Sulpizio, A.; Zienkiewicz, K.; Brandizzi, F. TGNap1 is required for microtubule-dependent homeostasis of a subpopulation of the plant trans-Golgi network. Nat. Commun. 2018, 9, 5313. [Google Scholar] [CrossRef] [Green Version]
- Delgadillo, M.O.; Ruano, G.; Zouhar, J.; Sauer, M.; Shen, J.; Lazarova, A.; Sanmartín, M.; Lai, L.T.F.; Deng, C.; Wang, P.; et al. MTV proteins unveil ER- and microtubule-associated compartments in the plant vacuolar trafficking pathway. Proc. Natl. Acad. Sci. USA 2020, 117, 9884–9895. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, M.; Qi, X.; Doyle, C.; Zheng, H. Armadillo-repeat kinesin1 interacts with Arabidopsis atlastin RHD3 to move ER with plus-end of microtubules. Nat. Commun. 2020, 11, 5510. [Google Scholar] [CrossRef]
- Erickson, J.L.; Schattat, M.H. Shaping plastid stromules-principles of in vitro membrane tubulation applied in planta. Curr. Opin. Plant Biol. 2018, 46, 48–54. [Google Scholar] [CrossRef]
- Hanson, M.R.; Conklin, P.L. Stromules, functional extensions of plastids within the plant cell. Curr. Opin. Plant Biol. 2020, 58, 25–32. [Google Scholar] [CrossRef]
- Brueggeman, J.M.; Windham, I.A.; Nebenführ, A. Nuclear movement in growing Arabidopsis root hairs involves both actin filaments and microtubules. J. Exp. Bot. 2022, 73, 5388–5399. [Google Scholar] [CrossRef] [PubMed]
- Spiegelman, Z.; Lee, C.M.; Gallagher, K.L. KinG is a plant-specific kinesin that regulates both intra- and intercellular movement of SHORT-ROOT. Plant Physiol. 2018, 176, 392–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, L.; Zhang, C.; Gao, B.; Zhang, Y.; Stewart, E.; Jez, J.; Nakajima, K.; Chen, X. Microtubules promote the non-cell autonomous action of microRNAs by inhibiting their cytoplasmic loading onto ARGONAUTE1 in Arabidopsis. Dev. Cell 2022, 57, 995–1008.e5. [Google Scholar] [CrossRef] [PubMed]
- Paredez, A.R.; Somerville, C.R.; Ehrhardt, D.W. Visualization of cellulose synthase demonstrates functional association with microtubules. Science 2006, 312, 1491–1495. [Google Scholar] [CrossRef] [Green Version]
- Crowell, E.F.; Bischoff, V.; Desprez, T.; Rolland, A.; Stierhof, Y.D.; Schumacher, K.; Gonneau, M.; Höfte, H.; Vernhettes, S. Pausing of Golgi bodies on microtubules regulates secretion of cellulose synthase complexes in Arabidopsis. Plant Cell 2009, 21, 1141–1154. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, R.; Lindeboom, J.J.; Paredez, A.R.; Emons, A.M.; Ehrhardt, D.W. Arabidopsis cortical microtubules position cellulose synthase delivery to the plasma membrane and interact with cellulose synthase trafficking compartments. Nat. Cell Biol. 2009, 11, 797–806. [Google Scholar] [CrossRef]
- Hong, L.; Dumond, M.; Zhu, M.; Tsugawa, S.; Li, C.B.; Boudaoud, A.; Hamant, O.; Roeder, A.H.K. Heterogeneity and robustness in plant morphogenesis: From cells to organs. Annu. Rev. Plant Biol. 2018, 69, 469–495. [Google Scholar] [CrossRef]
- Bao, Z.; Xu, Z.; Zang, J.; Bürstenbinder, K.; Wang, P. The morphological diversity of plant organs: Manipulating the organization of microtubules may do the trick. Front. Cell Dev. Biol. 2021, 9, 649626. [Google Scholar] [CrossRef]
- Yan, H.; Chaumont, N.; Gilles, J.F.; Bolte, S.; Hamant, O.; Bailly, C. Microtubule self-organisation during seed germination in Arabidopsis. BMC. Biol. 2020, 18, 44. [Google Scholar] [CrossRef]
- Deng, J.; Wang, X.; Liu, Z.; Mao, T. The microtubule-associated protein WDL4 modulates auxin distribution to promote apical hook opening in Arabidopsis. Plant Cell 2021, 33, 1927–1944. [Google Scholar] [CrossRef]
- Elliott, A.; Shaw, S.L. Microtubule array patterns have a common underlying architecture in hypocotyl cells. Plant Physiol. 2018, 176, 307–325. [Google Scholar] [CrossRef] [Green Version]
- Ma, Q.; Wang, X.; Sun, J.; Mao, T. Coordinated regulation of hypocotyl cell elongation by light and ethylene through a microtubule destabilizing protein. Plant Physiol. 2018, 176, 678–690. [Google Scholar] [CrossRef] [Green Version]
- Lan, M.; Kang, E.; Liu, X.; Fu, Y.; Zhu, L. Stable ARMADILLO REPEAT KINESIN 2 in light inhibits hypocotyl elongation and facilitates light-induced cortical microtubule reorientation in Arabidopsis. J. Exp. Bot. 2022, 74, erac473. [Google Scholar] [CrossRef]
- Mir, R.; Morris, V.H.; Buschmann, H.; Rasmussen, C.G. Division plane orientation defects revealed by a synthetic double mutant phenotype. Plant Physiol. 2018, 176, 418–431. [Google Scholar] [CrossRef] [Green Version]
- Halat, L.; Gyte, K.; Wasteneys, G. The microtubule-associated protein CLASP Is translationally regulated in light-dependent root apical meristem growth. Plant Physiol. 2020, 184, 2154–2167. [Google Scholar] [CrossRef]
- Ovečka, M.; Luptovčiak, I.; Komis, G.; Šamajová, O.; Samakovli, D.; Šamaj, J. Spatiotemporal pattern of ectopic cell divisions contribute to mis-shaped phenotype of primary and lateral roots of katanin1 mutant. Front. Plant Sci. 2020, 11, 734. [Google Scholar] [CrossRef]
- Feiguelman, G.; Cui, X.; Sternberg, H.; Hur, E.B.; Higa, T.; Oda, Y.; Fu, Y.; Yalovsky, S. Microtubule-associated ROP interactors affect microtubule dynamics and modulate cell wall patterning and root hair growth. Development 2022, 149, dev200811. [Google Scholar] [CrossRef]
- Vilches Barro, A.; Stöckle, D.; Thellmann, M.; Ruiz-Duarte, P.; Bald, L.; Louveaux, M.; von Born, P.; Denninger, P.; Goh, T.; Fukaki, H.; et al. Cytoskeleton dynamics are necessary for early events of lateral root initiation in Arabidopsis. Curr. Biol. 2019, 29, 2443–2454.e5. [Google Scholar] [CrossRef] [Green Version]
- Kirchhelle, C.; Garcia-Gonzalez, D.; Irani, N.G.; Jérusalem, A.; Moore, I. Two mechanisms regulate directional cell growth in Arabidopsis lateral roots. eLife 2019, 8, e47988. [Google Scholar] [CrossRef]
- Stöckle, D.; Reyes-Hernández, B.J.; Barro, A.V.; Nenadić, M.; Winter, Z.; Marc-Martin, S.; Bald, L.; Ursache, R.; Fujita, S.; Maizel, A.; et al. Microtubule-based perception of mechanical conflicts controls plant organ morphogenesis. Sci. Adv. 2022, 8, eabm4974. [Google Scholar] [CrossRef]
- Qian, Y.; Wang, X.; Liu, Y.; Wang, X.; Mao, T. HY5 inhibits lateral root initiation in Arabidopsis through negative regulation of the microtubule-stabilizing protein TPXL5. Plant Cell 2022, 13, koac358. [Google Scholar] [CrossRef] [PubMed]
- Champion, C.; Lamers, J.; Jones, V.A.S.; Morieri, G.; Honkanen, S.; Dolan, L. Microtubule associated protein WAVE DAMPENED2-LIKE (WDL) controls microtubule bundling and the stability of the site of tip-growth in Marchantia polymorpha rhizoids. PLoS Genet. 2021, 17, e1009533. [Google Scholar] [CrossRef] [PubMed]
- Otani, K.; Ishizaki, K.; Nishihama, R.; Takatani, S.; Kohchi, T.; Takahashi, T.; Motose, H. An evolutionarily conserved NIMA-related kinase directs rhizoid tip growth in the basal land plant Marchantia polymorpha. Development 2018, 145, dev154617. [Google Scholar] [CrossRef] [Green Version]
- Li, E.; Wang, G.; Zhang, Y.L.; Kong, Z.; Li, S. FERONIA mediates root nutating growth. Plant J. 2020, 104, 1105–1116. [Google Scholar] [CrossRef] [PubMed]
- Xin, P.; Schier, J.; Šefrnová, Y.; Kulich, I.; Dubrovsky, J.G.; Vielle-Calzada, J.P.; Soukup, A. The Arabidopsis TETRATRICOPEPTIDE-REPEAT THIOREDOXIN-LIKE (TTL) family members are involved in root system formation via their interaction with cytoskeleton and cell wall remodeling. Plant J. 2022, 112, 946–965. [Google Scholar] [CrossRef]
- Harding, S.A.; Hu, H.; Nyamdari, B.; Xue, L.J.; Naran, R.; Tsai, C.J. Tubulins, rhythms and cell walls in poplar leaves: It’s all in the timing. Tree Physiol. 2018, 38, 397–408. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.; Du, F.; Oliveri, H.; Zhou, L.; Ali, O.; Chen, W.; Feng, S.; Wang, Q.; Lü, S.; Long, M.; et al. Microtubule-mediated wall anisotropy contributes to leaf blade flattening. Curr. Biol. 2020, 30, 3972–3985.e6. [Google Scholar] [CrossRef]
- Hamant, O.; Inoue, D.; Bouchez, D.; Dumais, J.; Mjolsness, E. Are microtubules tension sensors? Nat. Commun. 2019, 10, 2360. [Google Scholar] [CrossRef] [Green Version]
- Sampathkumar, A.; Krupinski, P.; Wightman, R.; Milani, P.; Berquand, A.; Boudaoud, A.; Hamant, O.; Jönsson, H.; Meyerowitz, E.M. Subcellular and supracellular mechanical stress prescribes cytoskeleton behavior in Arabidopsis cotyledon pavement cells. eLife 2014, 3, e01967. [Google Scholar] [CrossRef]
- Bidhendi, A.J.; Altartouri, B.; Gosselin, F.P.; Geitmann, A. Mechanical stress initiates and sustains the morphogenesis of wavy leaf epidermal cells. Cell Rep. 2019, 28, 1237–1250.e6. [Google Scholar] [CrossRef]
- Verger, S.; Long, Y.; Boudaoud, A.; Sampathkumar, O. A tension-adhesion feedback loop in plant epidermis. eLife 2018, 7, e34460. [Google Scholar] [CrossRef]
- Schneider, R.; Ehrhardt, D.W.; Meyerowitz, E.M.; Sampathkumar, A. Tethering of cellulose synthase to microtubules dampens mechano-induced cytoskeletal organization in Arabidopsis pavement cells. Nat. Plants 2022, 8, 1064–1073. [Google Scholar] [CrossRef]
- Belteton, S.A.; Sawchuk, M.G.; Donohoe, B.S.; Scarpella, E.; Szymanski, D.B. Reassessing the roles of PIN proteins and anticlinal microtubules during pavement cell morphogenesis. Plant Physiol. 2018, 176, 432–449. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Yang, Q.; Wang, Y.; Wang, L.; Fu, Y.; Wang, X. Brassinosteroids regulate pavement cell growth by mediating BIN2-induced microtubule stabilization. J. Exp. Bot. 2018, 69, 1037–1049. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; Dang, X.; Bai, W.; Liu, M.; Li, Y.; Zhu, L.; Yang, Y.; Yu, P.; Ren, H.; Huang, D.; et al. The IPGA1-ANGUSTIFOLIA module regulates microtubule organisation and pavement cell shape in Arabidopsis. New Phytol. 2022, 236, 1310–1325. [Google Scholar] [CrossRef]
- Mitra, D.; Klemm, S.; Kumari, P.; Quegwer, J.; Möller, B.; Poeschl, Y.; Pflug, P.; Stamm, G.; Abel, S.; Bürstenbinder, K. Microtubule-associated protein IQ67 DOMAIN5 regulates morphogenesis of leaf pavement cells in Arabidopsis thaliana. J. Exp. Bot. 2019, 70, 529–543. [Google Scholar] [CrossRef] [Green Version]
- Eng, R.C.; Schneider, R.; Matz, T.W.; Carter, R.; Ehrhardt, D.W.; Jönsson, H.; Nikoloskim, Z.; Sampathkumar, A. KATANIN and CLASP function at different spatial scales to mediate microtubule response to mechanical stress in Arabidopsis cotyledons. Curr. Biol. 2021, 31, 3262–3274.e6. [Google Scholar] [CrossRef]
- Wong, J.H.; Kato, T.; Belteton, S.A.; Shimizu, R.; Kinoshita, N.; Higaki, T.; Sakumura, Y.; Szymanski, D.B.; Hashimoto, T. Basic proline-rich protein-mediated microtubules are essential for lobe growth and flattened cell geometry. Plant Physiol. 2019, 181, 1535–1551. [Google Scholar] [CrossRef]
- Oda, Y.; Mimura, T.; Hasezawa, S. Regulation of secondary cell wall development by cortical microtubules during tracheary element differentiation in Arabidopsis cell suspensions. Plant Physiol. 2005, 137, 1027–1036. [Google Scholar] [CrossRef] [Green Version]
- Schneider, R.; Klooster, K.V.; Picard, K.L.; van der Gucht, J.; Demura, T.; Janson, M.; Sampathkumar, A.; Deinum, E.E.; Ketelaar, T.; Persson, S. Long-term single-cell imaging and simulations of microtubules reveal principles behind wall patterning during proto-xylem development. Nat. Commun. 2021, 12, 669. [Google Scholar] [CrossRef]
- Jacobs, B.; Schneider, R.; Molenaar, J.; Filion, L.; Deinum, E.E. Microtubule nucleation complex behavior is critical for cortical array homogeneity and xylem wall patterning. Proc. Natl. Acad. Sci. USA 2022, 119, e2203900119. [Google Scholar] [CrossRef] [PubMed]
- Oda, Y.; Iida, Y.; Kondo, Y.; Fukuda, H. Wood cell-wall structure requires local 2D-microtubule disassembly by a novel plasma membrane-anchored protein. Curr. Biol. 2010, 20, 1197–1202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oda, Y.; Fukuda, H. Initiation of cell wall pattern by a Rho- and microtubule-driven symmetry breaking. Science 2012, 337, 1333–1336. [Google Scholar] [CrossRef] [PubMed]
- Oda, Y.; Fukuda, H. Rho of plant GTPase signaling regulates the behavior of Arabidopsis kinesin-13A to establish secondary cell wall patterns. Plant Cell 2013, 25, 4439–4450. [Google Scholar] [CrossRef] [Green Version]
- Bourdon, M.; Kalmbach, L.; Helariutta, Y. Plant vasculature: Selective membrane-to-microtubule tethering patterns the xylem cell wall. Curr Biol. 2017, 27, R842–R844. [Google Scholar] [CrossRef] [Green Version]
- Sugiyama, Y.; Wakazaki, M.; Toyooka, K.; Fukuda, H.; Oda, Y. A novel plasma membrane-anchored protein regulates xylem cell-wall deposition through microtubuledependent lateral inhibition of Rho GTPase domains. Curr. Biol. 2017, 27, 2522–2528. [Google Scholar] [CrossRef]
- Smertenko, T.; Turner, G.; Fahy, D.; Brew-Appiah, R.A.T.; Alfaro-Aco, R.; de Almeida Engler, J.; Sanguinet, K.A.; Smertenko, A. Brachypodium distachyon MAP20 functions in metaxylem pit development and contributes to drought recovery. New Phytol. 2020, 227, 1681–1695. [Google Scholar] [CrossRef]
- Sasaki, T.; Fukuda, H.; Oda, Y. CORTICAL MICROTUBULE DISORDERING1 is required for secondary cell wall patterning in xylem vessels. Plant Cell 2017, 29, 3123–3139. [Google Scholar] [CrossRef] [Green Version]
- Hervieux, N.; Tsugawa, S.; Fruleux, A.; Dumond, M.; Routier-Kierzkowska, A.L.; Komatsuzaki, T.; Boudaoud, A.; Larkin, J.C.; Smith, R.S.; Li, C.B.; et al. Mechanical shielding of rapidly growing cells buffers growth heterogeneity and contributes to organ shape reproducibility. Curr. Biol. 2017, 27, 3468–3479.e4. [Google Scholar] [CrossRef]
- Ma, H.; Xu, L.; Fu, Y.; Zhu, L. Arabidopsis QWRF1 and QWRF2 redundantly modulate cortical microtubule arrangement in floral organ growth and fertility. Front. Cell Dev. Biol. 2021, 9, 634218. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, B.; Dang, X.; Zhu, L.; Rao, J.; Ren, H.; Lin, C.; Qin, Y.; Lin, D. Arabidopsis IPGA1 is a microtubule-associated protein essential for cell expansion during petal morphogenesis. J. Exp Bot. 2019, 70, 5231–5243. [Google Scholar] [CrossRef] [Green Version]
- Ren, H.; Dang, X.; Cai, X.; Yu, P.; Li, Y.; Zhang, S.; Liu, M.; Chen, B.; Lin, D. Spatiotemporal orientation of microtubules controls conical cell shape in Arabidopsis thaliana petals. PLoS Genet. 2017, 13, e1006851. [Google Scholar] [CrossRef] [Green Version]
- Dang, X.; Yu, P.; Li, Y.; Yang, Y.; Zhang, Y.; Ren, H.; Chen, B.; Lin, D. Reactive oxygen species mediate conical cell shaping in Arabidopsis thaliana petals. PLoS Genet. 2018, 14, e1007705. [Google Scholar] [CrossRef]
- Riglet, L.; Rozier, F.; Kodera, C.; Bovio, S.; Sechet, J.; Fobis-Loisy, I.; Gaude, T. KATANIN-dependent mechanical properties of the stigmatic cell wall mediate the pollen tube path in Arabidopsis. eLife 2020, 9, e57282. [Google Scholar] [CrossRef]
- Wang, J.; Wang, G.; Liu, W.; Yang, H.; Wang, C.; Chen, W.; Zhang, X.; Tian, J.; Yu, Y.; Li, J.; et al. Brassinosteroid signals cooperate with katanin-mediated microtubule severing to control stamen filament elongation. EMBO J. 2022, 42, e111883. [Google Scholar] [CrossRef]
- Hashimoto, T. Microtubule and cell shape determination. In The Plant Cytoskeleton; Chapter 11; Liu, B., Ed.; Springer: New York, NY, USA, 2011; pp. 245–257. [Google Scholar]
- Xiao, H.; Jiang, N.; Schaffner, E.; Stockinger, E.J.; van der Knaap, E. A retrotransposon-mediated gene duplication underlies morphological variation of tomato fruit. Science 2008, 319, 1527–1530. [Google Scholar] [CrossRef]
- Yang, B.; Wendrich, J.R.; De Rybel, B.; Weijers, D.; Xue, H.W. Rice microtubule-associated protein IQ67-DOMAIN14 regulates grain shape by modulating microtubule cytoskeleton dynamics. Plant Biotechnol. J. 2020, 18, 1141–1152. [Google Scholar] [CrossRef]
- Wang, W.; Pan, Q.; Tian, B.; He, F.; Chen, Y.; Bai, G.; Akhunova, A.; Trick, H.N.; Akhunov, E. Gene editing of the wheat homologs of TONNEAU1-recruiting motif encoding gene affects grain shape and weight in wheat. Plant J. 2019, 100, 251–264. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Ge, F.; Qiang, Z.; Zhu, L.; Zhang, S.; Chen, L.; Wang, X.; Li, J.; Fu, Y. Maize ZmRPH1 encodes a microtubule-associated protein that controls plant and ear height. Plant Biotechnol. J. 2020, 18, 1345–1347. [Google Scholar] [CrossRef] [Green Version]
- Radchuk, V.; Borisjuk, L. Physical, metabolic and developmental functions of the seed coat. Front. Plant Sci. 2014, 5, 510. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Voiniciuc, C.; Fu, L.; Dieluweit, S.; Klose, H.; Usadel, B. TRM4 is essential for cellulose deposition in Arabidopsis seed mucilage by maintaining cortical microtubule organization and interacting with CESA3. New Phytol. 2019, 221, 881–895. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Stamm, G.; Bürstenbinder, K.; Voiniciuc, C. Microtubule-associated IQD9 orchestrates cellulose patterning in seed mucilage. New Phytol. 2022, 235, 1096–1110. [Google Scholar] [CrossRef] [PubMed]
- Graham, B.P.; Haigler, C.H. Microtubules exert early, partial, and variable control of cotton fiber diameter. Planta 2021, 253, 47. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Llewellyn, D.J.; Singh, S.P.; Green, A.G. Cotton seed development: Opportunities to add value to a byproduct of fiber production. In Flowering and Fruiting in Cotton; Oosterhuis, D.M., Cothren, J.T., Eds.; The Cotton Foundation Press: Cordova, TN, USA, 2012; pp. 131–162. [Google Scholar]
- Song, Q.; Gao, W.; Du, C.; Wang, J.; Zuo, K. Cotton microtubule-associated protein GhMAP20L5 mediates fiber elongation through the interaction with the tubulin GhTUB13. Plant Sci. 2022, 327, 111545. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Jeevaraj, T.; Yunus, M.H.; Chakraborty, S.; Chakraborty, N. The plant cytoskeleton takes center stage in abiotic stress responses and resilience. Plant Cell Environ. 2023, 46, 5–22. [Google Scholar] [CrossRef]
- Kesten, C.; Menna, A.; Sánchez-Rodríguez, C. Regulation of cellulose synthesis in response to stress. Curr. Opin. Plant Biol. 2017, 40, 106–113. [Google Scholar] [CrossRef]
- Wang, L.; Sadeghnezhad, E.; Riemann, M.; Nick, P. Microtubule dynamics modulate sensing during cold acclimation in grapevine suspension cells. Plant Sci. 2019, 280, 18–30. [Google Scholar] [CrossRef]
- Wang, L.; Nick, P. Cold sensing in grapevine-Which signals are upstream of the microtubular “thermometer”. Plant Cell Environ. 2017, 40, 2844–2857. [Google Scholar] [CrossRef]
- Wang, L.; Sadeghnezhad, E.; Nick, P. Upstream of gene expression: What is the role of microtubules in cold signalling? J. Exp. Bot. 2020, 71, 36–48. [Google Scholar] [CrossRef]
- Xu, X.; Walter, W.J.; Liu, Q.; Machens, I.; Nick, P. A rice class-XIV kinesin enters the nucleus in response to cold. Sci. Rep. 2018, 8, 3588. [Google Scholar] [CrossRef]
- Song, P.; Jia, Q.; Chen, L.; Jin, X.; Xiao, X.; Li, L.; Chen, H.; Qu, Y.; Su, Y.; Zhang, W.; et al. Involvement of Arabidopsis phospholipase D δ in regulation of ROS-mediated microtubule organization and stomatal movement upon heat shock. J. Exp. Bot. 2020, 71, 6555–6570. [Google Scholar] [CrossRef]
- Song, P.; Jia, Q.; Xiao, X.; Tang, Y.; Liu, C.; Li, W.; Li, T.; Li, L.; Chen, H.; Zhang, W.; et al. HSP70-3 interacts with phospholipase Dδ and participates in heat stress defense. Plant Physiol. 2021, 185, 1148–1165. [Google Scholar] [CrossRef]
- Wang, C.; Li, J.; Yuan, M. Salt tolerance requires cortical microtubule reorganization in Arabidopsis. Plant Cell Physiol. 2007, 48, 1534–1547. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Kurepa, J.; Hashimoto, T.; Smalle, J.A. Salt stress–induced disassembly of Arabidopsis cortical microtubule arrays involves 26S proteasome–dependent degradation of SPIRAL1. Plant Cell 2011, 23, 3412–3427. [Google Scholar] [CrossRef] [Green Version]
- Endler, A.; Kesten, C.; Schneider, R.; Zhang, Y.; Ivakov, A.; Froehlich, A.; Funke, N.; Persson, S. A mechanism for sustained cellulose synthesis during salt stress. Cell 2015, 162, 1353–1364. [Google Scholar] [CrossRef] [Green Version]
- Kesten, C.; Wallmann, A.; Schneider, R.; McFarlane, H.E.; Diehl, A.; Khan, G.A.; van Rossum, B.J.; Lampugnani, E.R.; Szymanski, W.G.; Cremer, N. The companion of cellulose synthase 1 confers salt tolerance through a Tau-like mechanism in plants. Nat. Commun. 2019, 10, 857. [Google Scholar] [CrossRef] [Green Version]
- Chun, H.J.; Baek, D.; Jin, B.J.; Cho, H.M.; Park, M.S.; Lee, S.H.; Lim, L.H.; Cha, Y.J.; Bae, D.W.; Kim, S.T. Microtubule dynamics plays a vital role in plant adaptation and tolerance to salt stress. Int. J. Mol. Sci. 2021, 22, 5957. [Google Scholar] [CrossRef]
- Yang, J.; An, B.; Luo, H.; He, C.; Wang, Q. AtKATANIN1 modulates microtubule depolymerization and reorganization in response to salt stress in Arabidopsis. Int. J. Mol. Sci. 2019, 21, 138. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Chen, Q.; Li, X.; Li, Y. MAP65-1 is required for the depolymerization and reorganization of cortical microtubules in the response to salt stress in Arabidopsis. Plant Sci. 2017, 264, 112–121. [Google Scholar] [CrossRef]
- Yu, B.; Zheng, W.; Xing, L.; Zhu, J.K.; Persson, S.; Zhao, Y. Root twisting drives halotropism via stress-induced microtubule reorientation. Dev. Cell 2022, 57, 2412–2425.e6. [Google Scholar] [CrossRef]
- Park, Y.C.; Chapagain, S.; Jang, C.S. The microtubule-associated RING finger protein 1 (OsMAR1) acts as a negative regulator for salt-stress response through the regulation of OCPI2 (O. sativa chymotrypsin protease inhibitor 2). Planta 2018, 247, 875–886. [Google Scholar] [CrossRef] [PubMed]
- Dou, L.; He, K.; Higaki, T.; Wang, X.; Mao, T. Ethylene signaling modulates cortical microtubule reassembly in response to salt stress. Plant Physiol. 2018, 176, 2071–2081. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Xuan, W.; Zhao, P.; Yao, X.; Peng, C.; Tian, Y.; Ye, J.; Wang, B.; He, J.; Chi, W. OsTUB1 confers salt insensitivity by interacting with Kinesin13A to stabilize microtubules and ion transporters in rice. New Phytol. 2022, 235, 1836–1852. [Google Scholar] [CrossRef] [PubMed]
- Siqueira, J.A.; de Oliveira, H.O.; Nunes-Nesi, A.; Araújo, W.L. Guard cell regulation: Pulling the strings behind the scenes. Trends Plant Sci. 2021, 26, 1093–1095. [Google Scholar] [CrossRef]
- Jiang, Y.; Wu, K.; Lin, F.; Qu, Y.; Liu, X.; Zhang, Q. Phosphatidic acid integrates calcium signaling and microtubule dynamics into regulating ABA-induced stomatal closure in Arabidopsis. Planta 2014, 239, 565–575. [Google Scholar] [CrossRef]
- Dou, L.; He, K.; Peng, J.; Wang, X.; Mao, T. The E3 ligase MREL57 modulates microtubule stability and stomatal closure in response to ABA. Nat. Commun. 2021, 12, 2181. [Google Scholar] [CrossRef]
- Yu, S.G.; Kim, J.H.; Cho, N.H.; Oh, T.R.; Kim, W.T. Arabidopsis RING E3 ubiquitin ligase JUL1 participates in ABA-mediated microtubule depolymerization, stomatal closure, and tolerance response to drought stress. Plant J. 2020, 103, 824–842. [Google Scholar] [CrossRef]
- Biel, A.; Moser, M.; Meier, I. Arabidopsis KASH proteins SINE1 and SINE2 are involved in microtubule reorganization during ABA-Induced stomatal closure. Front. Plant Sci. 2020, 11, 575573. [Google Scholar] [CrossRef]
- Wang, P.; Qi, S.; Wang, X.; Dou, L.; Jia, M.A.; Mao, T.; Guo, Y.; Wang, X. The OPEN STOMATA1-SPIRAL1 module regulates microtubule stability during abscisic acid-induced stomatal closure in Arabidopsis. Plant Cell 2023, 35, 260–278. [Google Scholar] [CrossRef]
- Swamy, P.S.; Hu, H.; Pattathil, S.; Maloney, V.J.; Xiao, H.; Xue, L.J.; Chung, J.D.; Johnson, V.E.; Zhu, Y.; Peter, G.F. Tubulin perturbation leads to unexpected cell wall modifications and affects stomatal behaviour in Populus. J. Exp. Bot. 2015, 66, 6507–6518. [Google Scholar] [CrossRef] [Green Version]
- Kállai, B.M.; Kourová, H.; Chumová, J.; Papdi, C.; Trögelová, L.; Kofroňová, O.; Hozák, P.; Filimonenko, V.; Mészáros, T.; Magyar, Z.; et al. γ-Tubulin interacts with E2F transcription factors to regulate proliferation and endocycling in Arabidopsis. J. Exp. Bot. 2020, 71, 1265–1277. [Google Scholar]
- Tompa, P.; Schad, E.; Tantos, A.; Kalmar, L. Intrinsically disordered proteins: Emerging interaction specialists. Curr. Opin. Struct. Biol. 2015, 35, 49–59. [Google Scholar] [CrossRef]
- Wright, P.E.; Dyson, H.J. Intrinsically disordered proteins in cellular signalling and regulation. Nat. Rev. Mol. Cell Biol. 2015, 16, 18–29. [Google Scholar] [CrossRef]
- Hsiao, A.S. Plant protein disorder: Spatial regulation, broad specificity, switch of signaling and physiological status. Front. Plant Sci. 2022, 13, 904446. [Google Scholar] [CrossRef]
- Guharoy, M.; Szabo, B.; Contreras Martos, S.; Kosol, S.; Tompa, P. Intrinsic structural disorder in cytoskeletal proteins. Cytoskeleton 2013, 70, 550–571. [Google Scholar] [CrossRef] [Green Version]
- Mukrasch, M.D.; Bibow, S.; Korukottu, J.; Jeganathan, S.; Biernat, J.; Griesinger, C.; Mandelkow, E.; Zweckstetter, M. Structural polymorphism of 441-residue tau at single residue resolution. PLoS Biol. 2009, 7, e34. [Google Scholar] [CrossRef]
- Mukrasch, M.D.; Mukrasch, M.D.; von Bergen, M.; Biernat, J.; Fischer, D.; Griesinger, C.; Mandelkow, E.; Zweckstetter, M. The “jaws” of the tau-microtubule interaction. J. Biol Chem. 2007, 282, 12230–12239. [Google Scholar] [CrossRef] [Green Version]
- Hsiao, A.S.; Wang, K.; Ho, T.D. An intrinsically disordered protein interacts with the cytoskeleton for adaptive root growth under stress. Plant Physiol. 2020, 183, 570–587. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Lian, N.; Zhang, Y.; Man, Y.; Chen, L.; Yang, H.; Lin, J.; Jing, Y. The cytoskeleton in plant immunity: Dynamics, regulation, and function. Int. J. Mol. Sci. 2022, 23, 15553. [Google Scholar] [CrossRef]
- Li, J.; Staiger, C.J. Understanding cytoskeletal dynamics during the plant immune response. Annu. Rev. Phytopathol. 2018, 56, 513–533. [Google Scholar] [CrossRef]
- Pitzalis, N.; Heinlein, M. The roles of membranes and associated cytoskeleton in plant virus replication and cell-to-cell movement. J. Exp. Bot. 2017, 69, 117–132. [Google Scholar] [CrossRef] [PubMed]
- Caplan, J.L.; Kumar, A.S.; Park, E.; Padmanabhan, M.S.; Hoban, K.; Modla, S.; Czymmek, K.; Dinesh-Kumar, S.P. Chloroplast stromules function during innate immunity. Dev. Cell 2015, 34, 45–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erickson, J.L.; Adlung, N.; Lampe, C.; Bonas, U.; Schattat, M.H. The Xanthomonas effector XopL uncovers the role of microtubules in stromule extension and dynamics in Nicotiana benthamiana. Plant J. 2018, 93, 856–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.S.; Park, E.; Nedo, A.; Alqarni, A.; Ren, L.; Hoban, K.; Modla, S.; McDonald, J.H.; Kambhamettu, C.; Dinesh-Kumar, S.P.; et al. Stromule extension along microtubules coordinated with actin-mediated anchoring guides perinuclear chloroplast movement during innate immunity. eLife 2018, 7, e23625. [Google Scholar] [CrossRef]
- Cui, Y.; Zhao, Y.; Lu, Y.; Su, X.; Chen, Y.; Shen, Y.; Lin, J.; Li, X. In vivo single-particle tracking of the aquaporin AtPIP2;1 in stomata reveals cell type-specific dynamics. Plant Physiol. 2021, 185, 1666–1681. [Google Scholar] [CrossRef]
- Lee, A.H.-Y.; Hurley, B.; Felsensteiner, C.; Yea, C.; Ckurshumova, W.; Bartetzko, V.; Wang, P.W.; Quach, V.; Lewis, J.D.; Liu, Y.C.; et al. A bacterial acetyltransferase destroys plant microtubule networks and blocks secretion. PLoS Pathog. 2012, 8, e1002523. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Kim, P.; Li, G.; Elowsky, C.; Alfano, J.R. A bacterial effector co-opts calmodulin to target the plant microtubule network. Cell Host Microbe 2016, 19, 67–78. [Google Scholar] [CrossRef] [Green Version]
- Robin, G.P.; Kleemann, J.; Neumann, U.; Cabre, L.; Dallery, J.F.; Lapalu, N.; O’Connell, R.J. Subcellular localization screening of Colletotrichum higginsianum effector candidates identifies fungal proteins targeted to plant peroxisomes, golgi bodies, and microtubules. Front. Plant Sci. 2018, 9, 562. [Google Scholar] [CrossRef] [Green Version]
- Nottensteiner, M.; Zechmann, B.; McCollum, C.; Hückelhoven, R. A barley powdery mildew fungus non-autonomous retrotransposon encodes a peptide that supports penetration success on barley. J. Exp. Bot. 2018, 69, 3745–3758. [Google Scholar] [CrossRef] [Green Version]
- Mei, Y.; Wright, K.M.; Haegeman, A.; Bauters, L.; Diaz-Granados, A.; Goverse, A.; Gheysen, G.; Jones, J.T.; Mantelin, S. The Globodera pallida SPRYSEC effector GpSPRY-414-2 that suppresses plant defenses targets a regulatory component of the dynamic microtubule network. Front. Plant Sci. 2018, 9, 1019. [Google Scholar] [CrossRef]
- Bhandari, D.D.; Brandizzi, F. Plant endomembranes and cytoskeleton: Moving targets in immunity. Curr. Opin. Plant Biol. 2020, 58, 8–16. [Google Scholar] [CrossRef]
- Léger, O.; Garcia, F.; Khafif, M.; Carrere, S.; Leblanc-Fournier, N.; Duclos, A.; Tournat, V.; Badel, E.; Didelon, M.; Le Ru, A.; et al. Pathogen-derived mechanical cues potentiate the spatio-temporal implementation of plant defense. BMC Biol. 2022, 20, 292. [Google Scholar] [CrossRef]
- Kitaeva, A.B.; Gorshkov, A.P.; Kusakin, P.G.; Sadovskaya, A.R.; Tsyganova, A.V.; Tsyganov, V.E. Tubulin cytoskeleton organization in cells of determinate nodules. Front. Plant Sci. 2022, 13, 823183. [Google Scholar] [CrossRef]
- Timmers, A.C.J. The role of the plant cytoskeleton in the interaction between legumes and rhizobia. J. Microsc. 2008, 231, 247–256. [Google Scholar] [CrossRef]
- Genre, A.; Timmers, T. The symbiotic role of the actin filament cytoskeleton. New Phytol. 2019, 221, 611–613. [Google Scholar] [CrossRef]
- Tsyganov, V.E.; Kitaeva, A.B.; Demchenko, K.N. Comparative analysis of tubulin cytoskeleton rearrangements in nodules of Medicago truncatula and Pisum sativum. In The Model Legume Medicago truncatula; de Bruijn, F.J., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2019; pp. 543–547. [Google Scholar]
- Kitaeva, A.B.; Demchenko, K.N.; Tikhonovich, I.A.; Timmers, A.C.; Tsyganov, V.E. Comparative analysis of the tubulin cytoskeleton organization in nodules of Medicago truncatula and Pisum sativum: Bacterial release and bacteroid positioning correlate with characteristic microtubule rearrangements. New Phytol. 2016, 210, 168–183. [Google Scholar] [CrossRef]
- Su, C.; Klein, M.L.; Hernández-Reyes, C.; Batzenschlager, M.; Ditengou, F.A.; Lace, B.; Keller, J.; Delaux, P.M.; Ott, T. The Medicago truncatula DREPP protein triggers microtubule fragmentation in membrane nanodomains during symbiotic infections. Plant Cell 2020, 32, 1689–1702. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.P.; Jiang, S.; Su, Y.; Xu, P.; Wang, J.; Liang, W.; Liu, C.W.; Murray, J.D. Intracellular infection by symbiotic bacteria requires the mitotic kinase AURORA1. Proc. Natl. Acad. Sci. USA 2022, 119, e2202606119. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, Q.; Wu, J.; Qi, M.; Zhang, C.; Huang, Y.; Wang, G.; Wang, H.; Tian, J.; Yu, Y.; et al. A legume kinesin controls vacuole morphogenesis for rhizobia endosymbiosis. Nat. Plants. 2022, 8, 1275–1288. [Google Scholar] [CrossRef]
- Genre, A.; Bonfante, P. Cytoskeleton-related proteins in tobacco mycorrhizal cells: Gamma-tubulin and clathrin localisation. Eur. J. Histochem. 1999, 43, 105–111. [Google Scholar]
- Blancaflor, E.B.; Zhao, L.; Harrison, M.J. Microtubule organization in root cells of Medicago truncatula during development of an arbuscular mycorrhizal symbiosis with Glomus versiforme. Protoplasma 2001, 217, 154–165. [Google Scholar] [CrossRef] [PubMed]
- Ho-Plágaro, T.; Huertas, R.L.; Tamayo-Navarrete, M.A.I.; Blancaflor, E.; Gavara, N.; García-Garrido, J.M. A novel putative microtubule-associated protein is involved in arbuscule development during arbuscular mycorrhiza formation. Plant Cell Physiol. 2021, 62, 306–320. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Zhang, Z.; Tong, Y.; Foda, B.M.; Day, B. ILEE: Algorithms and toolbox for unguided and accurate quantitative analysis of cytoskeletal images. J. Cell Biol. 2023, 222, e202203024. [Google Scholar] [CrossRef] [PubMed]
- Chakrabortty, B.; Willemsen, V.; de Zeeuw, T.; Liao, C.-Y.; Weijers, D.; Mulder, B.; Scheres, B. A plausible microtubule-based mechanism for cell division orientation in plant embryogenesis. Curr. Biol. 2018, 28, 3031–3043. [Google Scholar] [CrossRef] [Green Version]
- Moore, R.C.; Zhang, M.; Cassimeris, L.; Cyr, R.J. In vitro assembled plant microtubules exhibit a high state of dynamic instability. Cell Motil. Cytoskelet. 1997, 38, 278–286. [Google Scholar] [CrossRef]
- Alonso, M.C.; Drummond, D.R.; Kain, S.; Hoeng, J.; Amos, L.; Cross, R.A. An ATP gate controls tubulin binding by the tethered head of kinesin-1. Science 2007, 316, 120–123. [Google Scholar] [CrossRef] [Green Version]
- Widlund, P.O.; Podolski, M.; Reber, S.; Alper, J.; Storch, M.; Hyman, A.A.; Howard, J.; Drechsel, D.N. One-step purification of assembly-competent tubulin from diverse eukaryotic sources. Mol. Biol. Cell 2012, 23, 4393–4401. [Google Scholar] [CrossRef]
- Hotta, T.; Fujita, S.; Uchimura, S.; Noguchi, M.; Demura, T.; Muto, E.; Hashimoto, T. Affinity purification and characterization of functional tubulin from cell suspension cultures of Arabidopsis and tobacco. Plant Physiol. 2016, 170, 1189–1205. [Google Scholar] [CrossRef] [Green Version]
- Hotta, T.; Hashimoto, T. Affinity purification of tubulin from plant materials. Methods Cell Biol. 2020, 160, 263–280. [Google Scholar]
- Wong, J.H.; Hashimoto, T. Novel Arabidopsis microtubule-associated proteins track growing microtubule plus ends. BMC Plant Biol. 2017, 17, 33. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Østerlund, I.; Ruhnow, F.; Cao, Y.; Huang, G.; Cai, W.; Zhang, J.; Liang, W.; Nikoloski, Z.; Persson, S.; et al. Fluorescent cytoskeletal markers reveal associations between the actin and microtubule cytoskeleton in rice cells. Development 2022, 149, dev200415. [Google Scholar] [CrossRef] [PubMed]
- Marc, J.; Granger, C.L.; Brincat, J.; Fisher, D.D.; Kao, T.; McCubbin, A.G.; Cyr, R.J. A GFP-MAP4 reporter gene for visualizing cortical microtubule rearrangements in living epidermal cells. Plant Cell 1998, 10, 1927–1940. [Google Scholar] [PubMed] [Green Version]
- Lazzaro, M.D.; Wu, S.; Snouffer, A.; Wang, Y.; van der Knaap, E. Plant organ shapes are regulated by protein interactions and associations with microtubules. Front. Plant Sci. 2018, 9, 1766. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Comparison of plant and animal cell division. Plant cell division (A) is characterized by microtubule (MT)-based structures: the preprophase band, the acentrosomal mitotic spindle, and the phragmoplast. In animal cell cytokinesis (B), the contractile ring pinches the cell into two daughter cells, whereas plant phragmoplasts extend and guide vesicle fusion to generate the cell plate.
Figure 1.
Comparison of plant and animal cell division. Plant cell division (A) is characterized by microtubule (MT)-based structures: the preprophase band, the acentrosomal mitotic spindle, and the phragmoplast. In animal cell cytokinesis (B), the contractile ring pinches the cell into two daughter cells, whereas plant phragmoplasts extend and guide vesicle fusion to generate the cell plate.

Figure 2.
MT networks in response to environmental stresses. Environmental stresses cause the change in MT networks in plant cells. Insets illustrate MT network changes after pathogen infection and treatments for heat, salinity, and drought stress.
Figure 2.
MT networks in response to environmental stresses. Environmental stresses cause the change in MT networks in plant cells. Insets illustrate MT network changes after pathogen infection and treatments for heat, salinity, and drought stress.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hsiao, A.-S.; Huang, J.-Y. Microtubule Regulation in Plants: From Morphological Development to Stress Adaptation. Biomolecules 2023, 13, 627. https://doi.org/10.3390/biom13040627
AMA Style
Hsiao A-S, Huang J-Y. Microtubule Regulation in Plants: From Morphological Development to Stress Adaptation. Biomolecules. 2023; 13(4):627. https://doi.org/10.3390/biom13040627
Chicago/Turabian StyleHsiao, An-Shan, and Ji-Ying Huang. 2023. "Microtubule Regulation in Plants: From Morphological Development to Stress Adaptation" Biomolecules 13, no. 4: 627. https://doi.org/10.3390/biom13040627
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.