
Role of Pyroptosis in Intervertebral Disc Degeneration and Its Therapeutic Implications


1
The Clinical Medical College, Guizhou Medical University, Guiyang 550004, China
2
The First Clinical Medical College, Lanzhou University, Lanzhou 730000, China
3
Department of Endocrinology, First Hospital of Lanzhou University, Lanzhou 730000, China
4
Gansu Province Clinical Research Center for Endocrine Disease, Lanzhou 730000, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Biomolecules 2022, 12(12), 1804; https://doi.org/10.3390/biom12121804
Submission received: 3 November 2022
/
Revised: 27 November 2022
/
Accepted: 28 November 2022
/
Published: 2 December 2022
Abstract
:Intervertebral disc degeneration (IDD), a progressive and multifactorial pathological process, is predominantly associated with low back pain and permanent disability. Pyroptosis is a type of lytic programmed cell death triggered by the activation of inflammasomes and caspases. Unlike apoptosis, pyroptosis is characterized by the rupture of the plasma membrane and the release of inflammatory mediators, accelerating the destruction of the extracellular matrix (ECM). Recent studies have shown that pyrin domain-containing 3 (NLRP3) inflammasome-mediated pyroptosis in nucleus pulposus (NP) cells is activated in the progression of IDD. Furthermore, targeting pyroptosis in IDD demonstrates the excellent capacity of ECM remodeling and its anti-inflammatory properties, suggesting that pyroptosis is involved in the IDD process. In this review, we briefly summarize the molecular mechanism of pyroptosis and the pathogenesis of IDD. We also focus on the role of pyroptosis in the pathological progress of IDD and its targeted therapeutic application.
1. Introduction
Intervertebral disc (IVD) degeneration (IDD) is a leading cause of low back pain (LBP), affecting more than 80% of the population worldwide [1]. The global burden of disease study 2016 [2] ranked LBP as the leading cause of increased disability, with a lifetime prevalence of approximately 84%. In addition to lowering quality of life, IDD is also associated with a heavy socioeconomic burden. In the United States alone, healthcare expenditures for diseases related to IDD have reached USD 134.5 billion [3]. Notably, epidemiological surveys have reported a higher incidence of IDD in the elderly [4], postmenopausal women [5], and manual workers [6]. The pathogenesis of IDD is complex, involving an interplay of multiple factors which remain incompletely understood. It is generally believed that nucleus pulposus (NP) cell death [7], extracellular matrix (ECM) metabolic disorder [8], and secondary inflammation [9] are the three main causes of IDD. Macroscopically, IDD manifests as structural destruction and progressive dysfunction, including endplate (EP) sclerosis, osteophyte formation, and limited movement [10]. IDD can be improved with medical treatment in its early stages [11]. While the symptoms cannot be entirely relieved by conservative treatment, surgery is usually necessary to relieve the pain caused by the rupture or herniation of the IVD and to restore partial motor function. However, in essence, surgical treatment is still not an etiological treatment and fails to prevent or delay disease progression [12]. Additionally, its unclear curative effect, frequent complications, and high risk of recurrence cannot be ignored [13].
Initially observed in Shigella flexneri-infected macrophages in 1992 [14], pyroptosis was determined to be a form of proinflammatory cell death activated by caspase-1 in 2001 [15]. The term was coined from the Greek word pyro (fire or fever), reflecting its inflammatory nature, and ptosis (falling), meaning cell death. As it disrupts cell integrity features and belongs to a kind of programmed death driven by caspases, pyroptosis morphologically appears to be a combination of necrosis and apoptosis [16]. However, as a new type of cell death, pyroptosis has its specificity. In 2018, pyroptosis was defined as a form of cell death that was pore-formed by the gasdermin protein family and is often—but not always—activated by inflammatory caspase [17]. In recent years, pyroptosis has participated in many disease processes, including viral infection [18], tumors [19], and neurodegenerative diseases [20], etc. A broad set of studies have shown that NLRP3 inflammasome-mediated pyroptosis plays a significant role in IDD, and the inhibition of pyroptosis can alleviate IDD progression [21,22,23].
In this review, we discuss the role of pyroptosis in IDD progression based on the molecular mechanism of pyroptosis and the progression of IDD. Additionally, we summarize IDD therapeutic strategies targeting pyroptosis, providing a theoretical basis for further research.
2. Pyroptosis
2.1. Canonical Inflammasome-Induced Pyroptosis
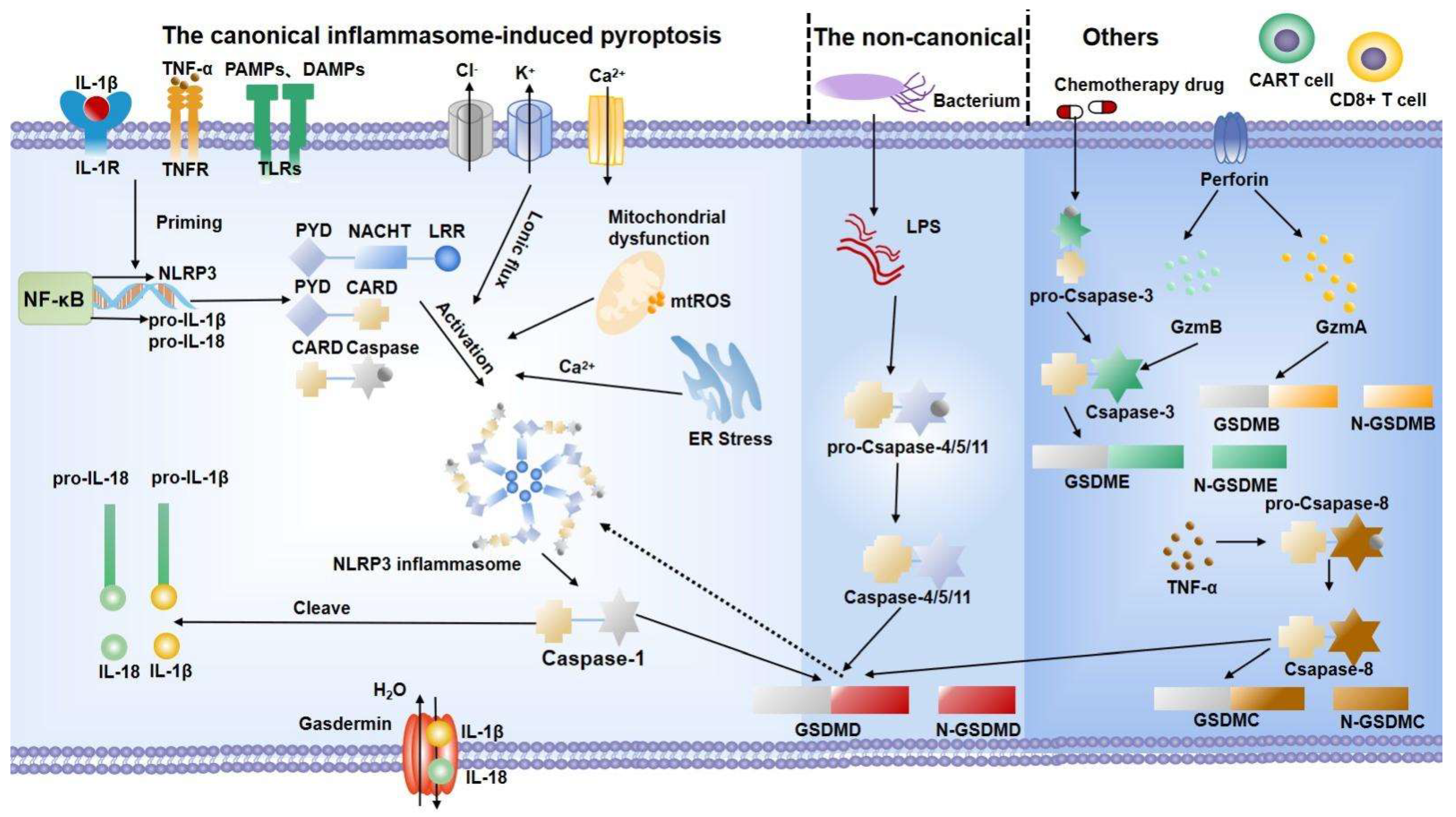
The canonical inflammasome-induced pyroptosis pathway begins with inflammasome assembly [24] (Figure 1). As a multiprotein complex in the cytoplasm, the inflammasome can be divided into three parts according to its domains: pattern recognition receptors (PRRs), apoptosis-associated speck-like protein containing caspase recruitment domain (ASC), and pro-caspase-1. Among them, PRRs comprise AIM2-like receptors, nucleotide-binding oligomerization domain-like receptors (NLRs), and pyrin [25]. PRRs, so-called cytoplasmic sensors, can recognize some molecules and then deliver them inside the host. These molecules contain “pathogen-associated molecular patterns (PAMPs)”, including bacteria, fungi, viruses, etc., as “non-self” signatures, and “damage-associated molecular patterns (DAMPs)”, such as ATP, cholesterol, interleukin (IL)-1β, and IL-18 [26]. Acting as the “adhesive” of the inflammasome, ASC contains a pyrin domain (PYD) and a caspase activation and recruitment domain (CARD). Stimulated by PRRs, pro-caspase-1 is recruited through typical protein–protein interactions, including homotypic ASC-PYD and PRRs-PYD, as well as ASC-CARD and pro-caspase-1-CARD interactions [27]. Finally, performing as an “effector”, pro-caspase-1 is an inactive precursor of active caspase-1. After the inflammasome complex is assembled, it obtains proteolytic activity by self-activating pro-caspase-1 to caspase-1, which in turn cleaves pro-IL-1β and pro-IL-18 into their mature secreted forms [25]. On the other hand, caspase-1 cleaves gasdermin D (GSDMD) by aggregating and self-processing it into caspase-1 p33/p10 [28]. GSDMD is a member of the gasdermin protein family, which consists of two conserved domains: the N-terminal pore-forming domain (PFD) and the C-terminal repressor domain (RD). Under normal circumstances, GSDMD is found in a quiescent state as a result of the autoinhibition produced by the interaction between PFD and RD [29]. After being cleaved by caspase-1, PFD binds to phosphatidylinositol of the eukaryotic cell membrane and cardiolipin of the prokaryotic cell membrane, and then assembles into a hollow oligomer with a diameter of 10–14 nm [30]. Exceeding the limitation of the endosomal sorting complex required for transport (ESCRT)-dependent membrane repair will lead to osmotic cell swelling and membrane perforation, releasing IL-1β and IL-18 [31].
Of note, it is usually postulated that the NLRP3 inflammasome is a key effector molecule of pyroptosis due to its multiple layers of transcriptional and post-translational modifications [32]. The NLRP3 inflammasome is generally expressed at low levels and remains ubiquitinated. Conventionally, the activation of the NLRP3 inflammasome can be divided into two phases: the “priming phase” and the “activating phase” [33]. The essential task of the priming signal is to deal with the increased expression and the post-translational modification of the NLRP3 inflammasome. To be more specific, toll-like receptors (TLRs) or cytokine receptors, including IL-1 receptors and tumor necrosis factor (TNF)-α receptors, can activate the transcription factor NF-κB and upregulate the expression of NLRP3 and pro-IL-1β by identifying ligands (lipopolysaccharide (LPS), TNF-α, IL-1, etc.) individually [34]. Additionally, deubiquitination [35] and phosphorylation [36] are required in the priming step. The activating phase of the NLRP3 inflammasome is triggered by the intracellular disturbance caused by multiple agonists. Among them, ionic flux, reactive oxygen species (ROS), and mitochondrial dysfunction are considered to be central factors [33].
2.2. Noncanonical Inflammasome-Induced Pyroptosis
The noncanonical inflammasome-induced pyroptosis pathway is mediated by human caspase-4/5 and mouse homologous caspase-11. In this way, LPS, a component of Gram-negative bacteria, directly activates caspase-11/4/5, playing a similar function to caspase-1 [37]. These inflammatory caspases cleave GSDMD to incur pyroptosis [38]. However, instead of converting pro-IL-1β and pro-IL-18 directly, caspase-4/5/11 improves the maturation of IL-1β and IL-18 through the NLRP3-caspase-1 pathway [39]. K+ behaves as a bridge, connecting the noncanonical pathway with the NLRP3 inflammasome. While GSDMD activation triggers membrane perforation, the efflux of K+ engages in the NLRP3 inflammasome activation [40]. Another study found that, upon intracellular LPS stimulation, caspase-11 specifically modifies the pannexin-1 channel, resulting in ATP release. ATP, the NLRP3 agonist, acting through P2X7 receptors on the cell membrane, triggers K+ efflux [41].
2.3. Other Relevant Approaches
The discovery that apoptosis-associated caspases-3/8 induce pyroptosis by cleaving GSDME contributes to a deeper understanding of pyroptosis. It has previously been demonstrated that chemotherapy drugs such as mitoxantrone, actinomycin-D, and cisplatin can convert the apoptosis phenotype to caspase-3/GSDME-dependent pyroptosis. Interestingly, the type of cell death is relevant to the expression level of GSDME [42]. New insights into the crosstalk between GSDMD inhibition in caspase-3 and activation in caspase-8 may elucidate interesting mechanisms concerning pyroptosis [43]. Furthermore, caspase-independent pyroptosis has been found in some cells. Chimeric antigen receptor T (CAR-T) cells can release granzyme B to activate caspase-3 [44] and lymphocytes produce granzyme A, acting on GSDMB directly [45]. Elastase, the serine protease in neutrophils, can also cleave GSDMD [46]. Further efforts are required to shed more light on this underlying mechanism.
3. The Pathogenesis of IDD
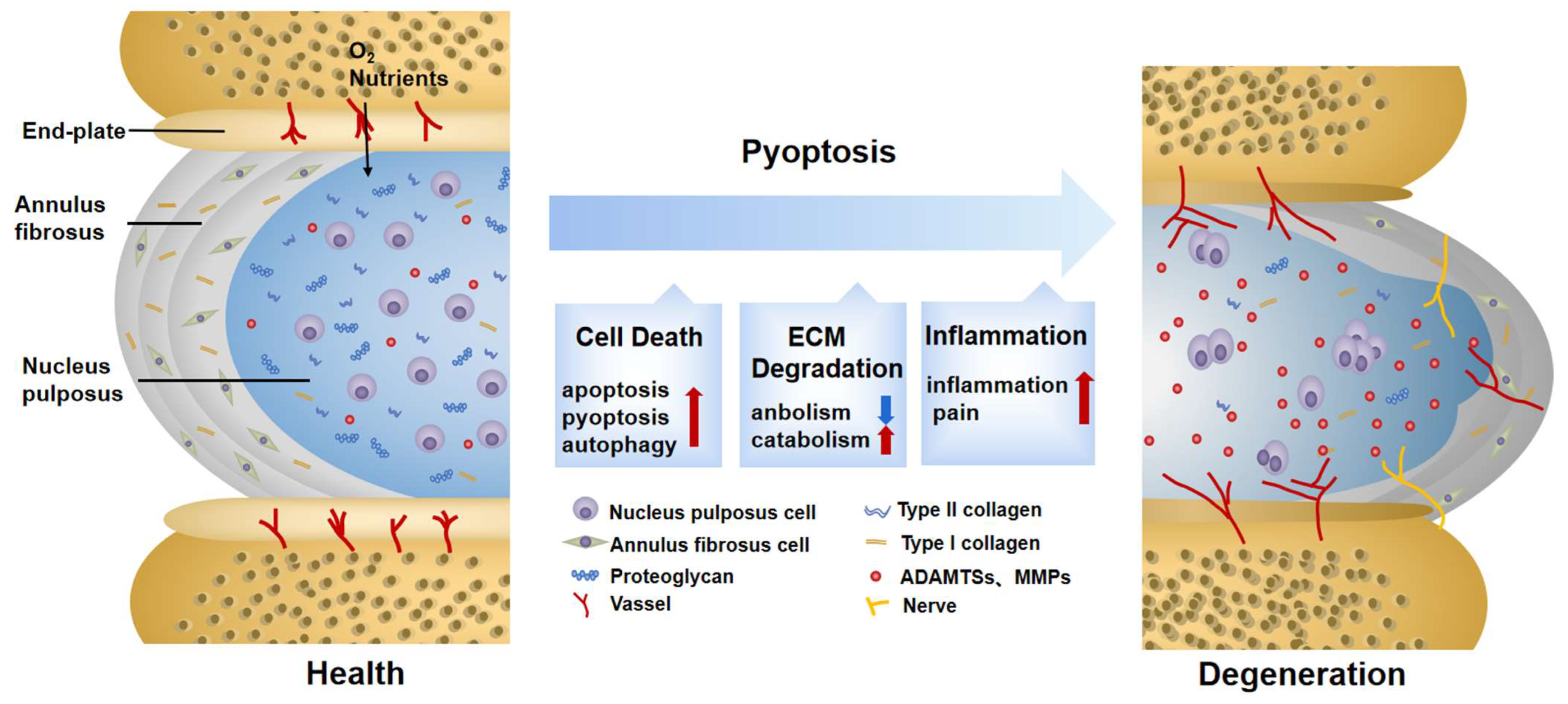
IDD is considered an age-related disease. After going through a degenerative cascade, it ultimately results in the loss of structural integrity, limited movement, pain, and decreased quality of life [10]. Causative factors of IDD can be categorized into passive destruction, including a violent blow or postural abrasion, and progressive changes. The latter is associated with aging and external environmental factors such as smoking, obesity, inflammation, and oxidation, etc. [47]. All of the above play a pathogenic role by changing the structure and metabolism of the IVD. A normal IVD is an avascular and innervated airtight fibrocartilaginous tissue located between the vertebrae. It consists of three parts: the nucleus pulposus (NP), the annulus fibrosus (AF), and the endplate (EP) [48]. Centrally located, the NP is surrounded by the AF, with the outermost EP wrapping the two structures together. Together, they constitute a closed cushioning system against stress, which enables the IVD to absorb and disperse loads from the vertebrae [10].
When IDD occurs, one of the most striking changes is the ECM disorder caused by NP cell damage [49]. Collagens (mainly type I and II collagens), proteoglycans, and elastins comprise the main components of the ECM. Among them, aggrecan is the main proteoglycan found in IVDs and is a material basis of ECM homeostasis [50]. Matrix metalloproteinases (MMPs) [51], mainly including MMP1, 2, 3, 7, 8, 10, and 13, as well as a disintegrin and metalloproteinase with thrombospondin motifs (ADAMTSs) [52], mainly including ADAMTS-1, 4, and 5, largely contribute to the degradation of the ingredients outlined above. The critical step of ECM disorder is proteoglycan degradation, accounting for the decreased hydration capacity of IVDs. This feature reduces the compression capacity and easily causes the expansion of the NP, resulting in cascading changes [53]. Divided into the outer, middle, and inner layers, the AF is a lamellar structure rich in collagens which can disperse mechanical stress to maintain the flexibility and stability of the spine. The nerve fibers and blood vessels distributed in the AF can transport nutrients to maintain the metabolism balance in the NP [54]. Apart from providing nutrients, the AF can disperse stress to prevent the radial disc bulge of the NP. At the same time, the additional stress caused by NP denaturation adds a burden to the outer layer of the AF. Specifically, the interval between the AF and the NP is vague, and the gaps between each layer are increased or even broken, eventually accelerating the loss of proteoglycan and type I collagen fibers [55]. The destruction of this structure leads to a functional change. In the AF, this is manifested by a decrease in antistretching ability and increased angiogenesis, triggering neuropathic pain and further impairing tissue homeostasis [56]. Likewise, the EP has the function of fixing and dispersing mechanical stress. In addition, the EP also performs as a translucent barrier between the avascular IVD and highly vascularized vertebrae, transporting oxygen, glucose, and metabolites through diffusive movement [57]. During the IDD process, EP becomes thinner and more calcified [58], resulting in a vicious circle as a result of hindered solute diffusion. Eventually, inflammation, hypoxia, acidity, and nutrient deprivation can be witnessed in the microenvironment of IDD. Biochemical and pathological changes, as noted above, aggravate the loss of proteoglycans, inhibit collagen cross-linking, and form osteophytes [59]. T1-weighted images show the diagnostic indicator of hyperintensity on the torn AF, whereas T2-weighted images display low signals of atrophy and dehydration in the NP region [60].
4. Roles of Pyroptosis in IDD
4.1. Pyroptosis Triggers Cell Death in IDD
Cell death, especially NP cell death, is one of the most critical factors triggering IDD. It leads to a loss of cell function and a decline in nutrient synthesis and repair capability and, moreover, increases inflammation and oxidative stress [7]. All of this aggravates the degenerative cascade in IVDs. A positive correlation between NLRP3 expression and the degenerative score was found in 45 clinical samples [61]. This result demonstrated that pyroptosis is widely involved in IDD progression, which is mainly induced by NLRP3 inflammasome (Figure 2).
Besides directly causing cell death, pyroptosis and other programmed-cell-death pathways are jointly cross-regulated in the complex and harsh degenerative environment. It was found that, after lumbar spine instability surgery, the height of the IVD was reduced in mice, and fissures and folds were formed between the AF layers, with osteophyte formation in the EP. The TUNEL assay and pyroptosis-related protein detection showed that both apoptosis and pyroptosis in the IDD-model group were potentiated [62]. Studies on anti-inflammatory protein A20 further revealed that when pyroptosis was inhibited, the expression of inflammatory cytokines coupled with apoptosis was also markedly reduced [63]. The above results suggest that there is an interplay between apoptosis and pyroptosis in IDD. The molecular mechanism behind the interplay may be explained by PANoptosis [64], which is characterized by pyroptosis, apoptosis, and necroptosis. Li et al. [65] first discovered PANoptosis, consisting of pyroptosis and apoptosis, in IDD lesions. They used different concentrations of TNF-α to treat NP cells and found that both pyroptosis and apoptosis were upregulated. However, pyroptosis was predominant at low concentrations, whereas apoptosis was predominant at high concentrations, implying that the major type of cell death may vary during different periods of IDD progression [65]. This interconverting relationship may be partially explained by the altered concentrations of inflammatory mediators in the IDD microenvironment and the activation of the CASP3-GSDME pathway in the crosstalk of pyroptosis and apoptosis [66]. Moreover, the relationship between pyroptosis and autophagy is also intriguing. In bromodomain-containing protein 4-inhibited rat NP cells, enhanced autophagy was able to reduce pyroptosis to relieve degenerative development. Whether autophagy enhancement is linked with attenuated pyroptosis requires further investigation; however, it has been demonstrated that the NF-κB signaling pathway is involved [67]. Similarly, one study found a tendency for autophagic vacuoles to increase with higher microtubule-associated protein 1 light chain 3 (LC3)-II/LC3-I ratios in NLRP3 knockout NP cells, demonstrating that pyroptosis and autophagy are antagonistic to each other [68]. Moreover, some researchers have tried to explore the mechanism underlying this phenomenon. One study pointed out that autophagy protects against LPS-induced NP cell pyroptosis. The explicit regulatory relationship between them depends on SQSTM1/p62, one of the autophagy-related proteins, co-localizing with GSDMD-N. It degraded GSDMD-N through the autophagy–lysosome pathway, thereby attenuating pyroptosis [69]. In conclusion, the interplay between pyroptosis and other programmed cell deaths of NP cells in IDD currently remains questionable. It is suggested that subsequent experiments focus on this aspect, which may have a significant role in revealing the progression of IDD.
4.2. Pyroptosis Provokes ECM Disorder in IDD
The composition, synthesis, and reconstruction of the ECM are extremely important for the stability of the IVD internal environment. When the ECM metabolism is severely disturbed, the hydration capacity of the IVD decreases, becoming stiffer and more fragile, thus affecting biological function [70]. Previous observations have shown that pyroptosis can induce ECM degradation. For example, the downregulation of nicotinamide phosphoribosyl transferase blocks the priming phase of NLRP3 through the NF-κB and MAPK pathways, reducing the degradation of aggrecan and collagen II [71]. Cholesterol accumulates in IDD, showing a repression of aggrecan and collagen II, as well as an elevation of MMP13 and ADTAMTS5. This is related to the activation of endoplasmic reticulum (ER) stress, an activating phase of NLRP3 [72]. Although their signals differ, both of them are DAMPs, which not only activate pyroptosis in NP cells, but also reduce ECM synthesis and increase ECM degradation. In addition to directly disrupting the balance of ECM synthesis and decomposition by causing NP cell death, the inflammatory mediators and cellular contents released by pyroptosis are largely involved in ECM disorder as well. NP cells are instrumental in the ECM metabolism [73]. When healthy NP cells co-culture with pyroptotic ones, the expressions of MMP13, ADAMTS4, and ADAMTS5 are upregulated [22]. The fragment products they hydrolyze damage the stress-resistance capability of the ECM and provoke secondary inflammatory responses [74]. TNF-α and IL-1β are acknowledged factors driving the synthesis and release of MMPs and ADAMTSs [75], which are also in close contact with pyroptosis. The mediators mentioned above may result in ECM degradation caused by pyroptosis. Additionally, this phenomenon is also found in Modic EP changes and degenerated AF tissues and is thus not limited to NP cells [61,62].
In addition, ECM disorder also aggravates pyroptosis. Previous research found that anaerobic glycolysis in degenerated NP cells creates an acidic environment which activates ROS/NLRP3/caspase-1-mediated pyroptosis [23]. However, one study reached the opposite conclusion. With increased acidification, ECM degradation may be aggravated by exogenous factors rather than the decreased NLRP3 in NP cells [76]. On the other hand, the accumulation of metabolites, such as advanced glycation end products, hardens collagen fibers and facilitates the assembly of NLRP3 [77]. This shows promise in regard to restoring ECM disturbances and delaying IDD progression by inhibiting pyroptosis. An injection of VX-76, a caspase-1 inhibitor, can prevent the replacement of proteoglycans by fibrous tissue and slow the aging and fibrosis of the IVD effectively [69].
4.3. Pyroptosis Induces Secondary Inflammation in IDD
Increased secondary inflammatory cytokines are regarded as a significant feature of symptomatic IDD [78]. The inner region of the IVD lacks vascularized structures and is isolated from the host immune system. NP cells usually lack immune tolerance but are capable of phagocyting and secreting inflammatory factors. When exposed to an inflammatory environment, they will produce a solid autoimmune and inflammatory cascade [79,80]. The inflammation in IDD manifests in three ways, which are partly assigned to pyroptosis.
Changes in IVD structure and physiology lead to the production of inflammatory factors, which is the most distinctive feature of the first stage [81]. Characterized by a typical inflammatory cell death mode, pyroptosis unleashes proinflammatory factors in IVD cells either directly or indirectly [17]. As a direct product of pyroptosis, IL-1β is recognized as a key cytokine involved in discogenic back pain [82]. At the same time, when exposed to IL-1β, the expression of inflammatory factors such as IL-6 and IL-8 is significantly elevated [83]. It is not uncommon to see that some substances in IDD can trigger an inflammatory reaction by inducing pyroptosis. Extracellular ATP (eATP) is a representative co-activator of NLRP3 and P2X7R from the ATP-gated ion channel family. When strongly stimulated, P2X7R transfers to the cytoplasm to co-locate with NLRP3 and release transforming growth factor-β (TGF-β) and IL-1. It is also related to IVD inflammation [84]. However, whether eATP causes inflammation directly or through pyroptosis remains to be further elaborated. Although IDD is mainly characterized by aseptic inflammation at this stage, infectious factors have gradually attracted attention in recent years. Among them, propionibacterium acnes account for 13% to 44% of the causes of IDD [85]. It was reported that propionibacterium acnes could induce the pyroptosis of NP cells through the ROS-NLRP3 signal pathway. Pyrolytic products aggravate the aggregation of inflammatory factors in IDD [22]. On the contrary, an injection of the NLRP3 inhibitor MCC950 successfully improves the inflammatory environment of IDD [86], suggesting that targeted pyroptosis may be a therapeutic strategy to prevent infection factors from aggravating the inflammatory progress of IDD. However, to date, in-depth insights into the specific antigens of the infectious factors inducing pyroptosis are still lacking. In addition, although pyroptosis is involved in a heightened inflammatory response in IDD, whether pyroptosis is the initial cause of IDD inflammation deserves further discussion.
The infiltration of leukocytes and the ingrowth of blood vessels and nerves are second-stage events. The results of this incremental process are closely related to the third stage, so we will expound them together. The third stage is identified by the pain symptoms caused by the sensitization of nerve endings and nociceptive neuron infiltration [81]. Sensory nerve fibers and blood vessels are usually distributed in the outer layer of AF tissue for nutrient transport [87]. In the mouse lumbar-spine-instability model, vascular bundles and sensory nerves grew in the inner and outer regions of the AF, with calcitonin gene-related peptide (CGRP)-positive cells increasing [62]. This may be attributed to the fact that cytokines released by pyroptosis aggravate the loss of proteoglycans in the ECM and promotes angiogenesis [88]. CGRP is an important nociceptive neurotransmitter for controlling inflammation and pain [89]. Additionally, a large number of inflammatory factors released by cell rupture constantly stimulate sensory nerves, which is also a significant pain factor. A study utilizing NF-κB inhibitor Bay11-7082 on the lumbar-disc-herniation rat model found that the assembly of NLRP3 was blocked, and CGRP and pain-related behaviors such as allodynia and heat pain thresholds were ameliorated. This suggests that NF-κB, the standard signal of pain and pyroptosis, is involved in relieving neuropathic pain [90]. Moreover, although the relationship between ion channels and inflammation has not been fully explained, Ca2+-dependent Pizol1 channels activated by mechanical stretching [91] and ASIC3 channels marked by acid sensing [23] can promote pyroptosis, which may influence the sensitization of pain in the IVD later [92].
5. Therapeutic Strategies Targeting Pyroptosis and NLRP3 Inflammasome in IDD
5.1. Exogenous Drugs
Exogenous drugs mainly refer to plant-derived natural compounds and chemical compounds that affect the human body through external administration. Thanks to their low toxicity, rich sources, and extensive pharmacological effects, plant-derived natural compounds have gradually become a hot topic in drug discovery [93]. Under the guidance of precise chemical structures, chemical compounds are essential for developing new chemical skeletons or tapping the activity of old drugs [94]. In IDD, exogenous drugs mainly target the NLRP3 inflammasome to inhibit pyroptosis (Table 1). As mentioned above, activating the NLRP3 inflammasome requires priming and activating phase signals. The activation of the transcriptional regulator dominates the priming signal to increase the expression of the NLRP3 inflammasome [33]. TLR4 is the most representative molecule of the Toll family, and can recognize ligand-independent signaling to recruit adapter proteins MyD88, TIRAP, and TRAM, and ultimately participate in the robust activation of NF-κB to increase NLRP3 transcription [95]. In the H2O2-mediated-degeneration model, ganoderic acid significantly inhibited the expression of TLR4 and NLRP3. However, it still remains uncertain whether ganoderic acid directly acts on TLR4 or triggers the secondary reduction in TLR4 by scavenging H2O2 [96]. In addition to H2O2, high-mobility group box-1 protein (HMGB1) and DAMPs also act on TLR4 receptors [97]. Magnoflorine reduced HMGB1 and blocked the recruitment of the Myd88 adapter, and then inhibited the NF-κB/NLRP3/caspase-1 pathway [98]. The released HMGB1 induced by LPS-medicated noncanonical inflammasome-induced pyroptosis [99] propagates the inflammatory response further through phagocytosis, and performs as a DAMP, finally activating caspase-1-dependent pyroptosis [100]. Detecting caspase-11 in this model may further reveal the role of magnoflorine. Oseltamivir [101] and Bay11-7082 [90] can also inhabit the NLRP3 inflammasome through the NF-κB pathway to delay the progression of IDD.
The second signals are related to various intracellular signal disorders concerning ionic flux and mitochondrial dysfunction [33]. Atorvastatin, primarily used to treat cardiovascular and inflammatory diseases, is found to reduce cholesterol content, inhibit inflammatory infiltration, and relieve ER stress to remodel the ECM in IDD [72]. Lipid disturbance caused by cholesterol increases ER pressure and is associated with mitochondria releasing Ca2+ [102]. An acid environment can also promote Ca2+. Under the intervention of paeoniflorin, the dropped PH and Ca2+ release was well-controlled, alleviating NLRP3-mediated pyroptosis and significantly improving the MRI pathological score of IDD [103]. ROS is also a key molecule for NLRP3 activation. Gamma oryzanol is a dietary supplement that can reduce the intracellular ROS and mitochondrial superoxide levels in NP cells. Its mechanism is possibly related to a reduction in thioredoxin-interacting protein (TXNIP), an ROS-sensitive protein that promotes NLRP3 oligomerization [104,105]. Morin [106] and honokiol [107] are potential small molecules for treating IDD through the TXNIP/NLRP3 pathways.
Although great strides have been made to explore exogenous components to delay IDD by regulating pyroptosis, explorations of their metabolic distribution and therapeutic doses are still lacking. Furthermore, almost no in vivo experiments have been carried out on humans thus far. Long-term toxicity and the mode of administration may be challenging issues for subsequent development.

{kind=link}
{kind=link}
Table 1.
Mechanisms of exogenous compounds for IDD treatment by inactivating pyroptosis.
| Type | Compound | Dose | Model Type | Mechanism | References | ||||
|---|---|---|---|---|---|---|---|---|---|
| Pathway | Cell Death | ECM | Inflammation | Others | |||||
| Plant-derived natural compounds | Ganoderic Acid A | 08 μM | Rat NP tissue (in vivo/in vitro) | Inactivated TLR4/NLRP3 | Suppressed apoptosis | Suppressed MMP3, MMP13, ADAMTS4, ADAMTS5. Upregulated Col II, aggrecan | Alleviated IL-6, IL-1β, TNF-α | Reduced oxidative stress by restoring GSH, SOD, GPX | [96] |
| Plant-derived natural compounds | Magnoflorine | 100 μg/mL | Human NP cells (in vitro) | Inactivated HMGB1/ MyD88/NF-κB/NLRP3 | Suppressed apoptosis | Suppressed MMP3, MMP13, ADAMTS4, ADAMTS5. Upregulated Col II, aggrecan | Alleviated HMGB1, IL-1β, IL-6, TNF-α, IL-18 | Alleviated “M1” polarized macrophage | [98] |
| Plant-derived natural compounds | Paeoniflorin | 20 mg/kg | Rat NP tissue (in vivo/in vitro) | Inactivated IL-1β/NLRP3 | Suppressed pyroptosis | Suppressed MMP3, MMP13. Upregulated Col II, aggrecan | Alleviated IL-1β, IL-18 | Reduced calcium concentration | [103] |
| Plant-derived natural compounds | Gamma oryzanol | 40 μM | Rat NP tissue (in vivo/in vitro) | Inactivated IL-1β/NF-κB/NLRP3 | Suppressed pyroptosis | Suppressed MMP13, ADAMTS5. Upregulated Col II, aggrecan | Alleviated IL-1β, IL-18 | —— | [104] |
| Plant-derived natural compounds | Morin | 50 mg/kg | Rat NP tissue (in vivo/in vitro) | Inactivated TXNIP/ NLRP3 | Suppressed pyroptosis | —— | Alleviated IL-1β, IL-18, TNF-α | —— | [106] |
| Plant-derived natural compounds | Honokiol | 40 μM | Rat NP tissue (in vivo/in vitro) | Inactivated NF-κB, JNK, TXNIP/NLRP3 | Suppressed apoptosis | Suppressed MMP3, MMP13, ADAMTS4, ADAMTS5. Upregulated Col II, SOX9 | Alleviated IL-6, COX-2, iNOS | Reduced oxidative stress by restoring MDA, SOD, GPX | [107] |
| Chemical compound | Bay11-7082 | 5 mg/kg | Rat NP tissue (in vivo/in vitro) | Inactivated NF-κB/NLRP3 | —— | —— | Alleviated IL-1β, IL-18. Downregulated CGRP in DRG neurons | —— | [90] |
| Chemical compound | Atorvastatin | 20 μM | Rat NP tissue (in vivo/in vitro) | Inactivated NF-κB/NLRP3 | Promoted autophagy. Suppressed pyroptosis | Suppressed MMP3, MMP13, ADAMTS4, ADAMTS5. Upregulated Col II, aggrecan | Alleviated IL-1β, IL-18, TNF-α | Alleviated SREBP1 mediated cholesterol-induced pyroptosis and ER stress | [72] |
5.2. Endogenous Small Molecules
Endogenous small molecules are substances with physiological functions and biological activities that exist naturally in human or mammalian bodies. They have strong pharmacological effects and few side effects [108]. Endogenous small molecules from a wide range of sources affect IDD through multiple pyroptosis-related signaling pathways. Nicotinamide phosphoribosyl transferase inhibitor [71] and platelet-derived growth factor BB [109] can inhibit NLRP3-mediated pyroptosis and play a protective role in ECM by NAMPT/MAPK/NF-κB and MAPK/PI3K/ERK, respectively. Maresin 1 [110], melatonin [111], cortistatin [112], platelet-rich plasma [113], MFG-E8 [114], A20 [63], lipoxin A4 [115], and SIRT1 [116] can reduce antioxidative stress and inhibit inflammatory factors via the NF-κB, NAMPT/MAPK, Keap1-Nrf2, Nrf2/TXNIP, and JNK1/Declin-1/PI3KC3 pathways. The specific mechanisms are shown in Table 2. Although increasing attention has been paid to the development of endogenous small molecules, the direct administration of endogenous molecules or the induction of the expression of endogenous molecules through activators is still an issue worthy of consideration. Additionally, if administered directly, how to prevent degradation as a result of pharmacokinetics is also an inevitable question.
5.3. Stem Cells and Bioengineering
Due to the special anatomical structure and complex physiological environment of the IVD, exogenous cells struggle to survive, and the efficiency of the drug delivery route is poor [117]. At present, stem cells and bioengineering are used to overcome the issues related to physiological structure and mechanical characteristics, opening new therapeutic avenues for IDD treatment [118]. Stem cells can release exosomes, which are extracellular vesicles 50–100 nm in diameter. They have a low risk of rejection and efficient drug administration [117]. It has been proven that IDD can be alleviated by inhibiting pyroptosis-related signaling pathways through exosomes. Exosomes derived from human umbilical cord mesenchymal stem cells deliver miR-26a-5p to inhibit m6A methylation in NLRP3-medicated pyroptosis by reducing methyltransferase METTL14 in NP cells [119]. Mesenchymal stem-cell-derived exosomal miR-410 [120] and platelet-plasma-derived exosomal miR-141-3p [113] can also target NLRP3 as efficient therapy options to treat IDD. On the basis of the above results, how to improve the targeting efficiency and durability of exosomes is worth discussing. Xing et al. [121] designed an injectable exosome-functionalized extractive-matrix hydrogel by fixing the exosomes extracted from adipose-tissue-derived mesenchymal stem cells on the ECM gel. This device not only directly replenished the ECM, but also continuously increased the loading rate of exosomes to maintain the stability of the ECM by inhibiting pyroptosis. Apart from exosomes, extracellular vesicles (EVs) generated by stem cells are also a good therapeutic choice. Zhang et al. [122] constructed a Cavin-2-modified EV system derived from mesenchymal stem cells, which was loaded with NLRP3 inflammasome antagonist peroxiredoxin-2. The system increased the uptake and permeability rate of EV under TNF-α-agglomerated proinflammatory conditions in IDD. Esterase-responsive ibuprofen nanomicelle premodified embryo-derived nucleus pulposus progenitor cells show outstanding advantages. Esterase-responsive ibuprofen nanomicelles can release ibuprofen into the acidic environment of IDD, which improves the microenvironment of IDD through inhibiting COX2/NF-κB/caspase-1-mediated pyroptosis [123]. Furthermore, ensuring AF cell resistance against NP stress is another feasible research direction. Zhang et al. [124] prepared a fabrication of a polylactide-glycolide/poly-ε-caprolactone/dextran/plastrum testudinis extract (PTE) composite with an anti-inflammation nanofiber membrane. Loaded with PTE, it inhibited NLRP3 and its upstream and downstream signaling pathways and promoted AF cell regeneration.
The therapeutic approaches designed by bioengineering methods targeting pyroptosis and the NLRP3 inflammasome in IDD have broad prospects. In the future, exploring different methods of transportation and bioactive molecules loaded with biomaterials tailored to pyroptosis could be a potential direction. At the same time, it could be promising for the follow-up bioengineering research on IDD to focus on other markers of pyroptosis, such as CASP1, GSDMD, and ASC.
6. Summary and Outlook
Pyroptosis, characterized by the secretion of inflammatory factors and cell destruction, is a new type of cell death that has attracted much attention in the last decade. It has been proven to be associated with IDD. Specifically, pyroptosis is involved in the microscopic progression of IDD by triggering cell death, ECM disorder, and secondary inflammatory responses. These three processes can also further influence pyroptosis, which eventually develops into clinically typical structural destruction and the progressive dysfunction of the IVD, severely reducing patient quality of life.
Although the relationship between pyroptosis and IDD has been recognized, there are still deficiencies. First, a large number of homogenization studies have hindered the search for new mechanisms. The research on IDD primarily focuses on NLRP3 inflammasome-mediated pyroptosis, and little is known about NLRP1/4/9 and AIM2. NLRP1, AIM2 and GSDME are usually reduced or missing in livestock such as pigs, cats, and cattle, making them more resistant to bacterial invasion [125]. The above pyroptotic components may partly explain the reason why humans are more vulnerable to IDD compared to other vertebrates. In other words, the animal-model in vivo experiments should be chosen with caution; otherwise, the results may not reflect the real situations in humans. Secondly, considering the three anatomical structures that coordinate the function of IVDs, the current research on pyroptosis in IDD is chiefly limited to the NP, and the doubts concerning the AF and the EP are yet to be investigated. Moreover, a considerable number of studies have concentrated on molecular biology, but only a few researchers have evaluated changes in the macrostructure of the IVD, which also hinders the foundation of clinical biomarkers. Additionally, no IDD model can replicate the human IDD phenomenon; most experiments at present lack clinical validation and are not sufficiently convincing. The absence of application to the human body also limits our understanding of the required dosage and side effects of pyroptosis therapy. Apart from solving this dilemma, the next step could be to focus on the interaction of pyroptosis with other forms of death in IDD. In conclusion, the inhibition of pyroptosis is a promising strategy for treating IDD.
Author Contributions
S.F. contributed to the conception and design of the manuscript. J.L. and Y.Y. outlined and revised the manuscript. X.W. and X.C. analyzed and finalized the figures and tables. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Construction Plan of the Gansu Province Clinical Research Center for Endocrine Disease (20JR10FA667); the National Key R&D Program of China (2017YFC1310700 and 2017YFC1310702); the Project of the Gansu Natural Science Foundation (22JR5RA914); the National innovation and entrepreneurship training program for College Students (202210730175 and 202110660016); the Guizhou Provincial innovation and entrepreneurship training program for College Students (202110660036); and Lanzhou University 2022 College Students Innovation and Entrepreneurship (20220060133).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Maher, C.; Underwood, M.; Buchbinder, R. Non-specific low back pain. Lancet 2017, 389, 736–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Disease, G.B.D.; Injury, I.; Prevalence, C. Global, regional, and national incidence, prevalence, and years lived with disability for 328 diseases and injuries for 195 countries, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet 2017, 390, 1211–1259. [Google Scholar] [CrossRef] [Green Version]
- Dieleman, J.L.; Cao, J.; Chapin, A.; Chen, C.; Li, Z.; Liu, A.; Horst, C.; Kaldjian, A.; Matyasz, T.; Scott, K.W.; et al. US Health Care Spending by Payer and Health Condition, 1996–2016. JAMA 2020, 323, 863–884. [Google Scholar] [CrossRef] [PubMed]
- Cheung, K.M.; Karppinen, J.; Chan, D.; Ho, D.W.; Song, Y.Q.; Sham, P.; Cheah, K.S.; Leong, J.C.; Luk, K.D. Prevalence and pattern of lumbar magnetic resonance imaging changes in a population study of one thousand forty-three individuals. Spine 2009, 34, 934–940. [Google Scholar] [CrossRef] [PubMed]
- de Schepper, E.I.; Damen, J.; van Meurs, J.B.; Ginai, A.Z.; Popham, M.; Hofman, A.; Koes, B.W.; Bierma-Zeinstra, S.M. The association between lumbar disc degeneration and low back pain: The influence of age, gender, and individual radiographic features. Spine 2010, 35, 531–536. [Google Scholar] [CrossRef]
- Jo, M.; Chae, S.W. Stress analysis of intervertebral disc during occupational activities. Comput. Methods Programs Biomed. 2021, 208, 106298. [Google Scholar] [CrossRef]
- Ding, F.; Shao, Z.W.; Xiong, L.M. Cell death in intervertebral disc degeneration. Apoptosis 2013, 18, 777–785. [Google Scholar] [CrossRef]
- Liang, H.; Luo, R.; Li, G.; Zhang, W.; Song, Y.; Yang, C. The Proteolysis of ECM in Intervertebral Disc Degeneration. Int. J. Mol. Sci. 2022, 23, 1715. [Google Scholar] [CrossRef]
- Navone, S.E.; Marfia, G.; Giannoni, A.; Beretta, M.; Guarnaccia, L.; Gualtierotti, R.; Nicoli, D.; Rampini, P.; Campanella, R. Inflammatory mediators and signalling pathways controlling intervertebral disc degeneration. Histol. Histopathol. 2017, 32, 523–542. [Google Scholar] [CrossRef]
- Raj, P.P. Intervertebral disc: Anatomy-physiology-pathophysiology-treatment. Pain Pract. 2008, 8, 18–44. [Google Scholar] [CrossRef]
- Zhao, L.; Manchikanti, L.; Kaye, A.D.; Abd-Elsayed, A. Treatment of Discogenic Low Back Pain: Current Treatment Strategies and Future Options—A Literature Review. Curr. Pain Headache Rep. 2019, 23, 86. [Google Scholar] [CrossRef] [PubMed]
- Eisenstein, S.M.; Balain, B.; Roberts, S. Current Treatment Options for Intervertebral Disc Pathologies. Cartilage 2020, 11, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.H.; Kim, H.S.; Jang, I.T. Intervertebral Disc Diseases PART 2: A Review of the Current Diagnostic and Treatment Strategies for Intervertebral Disc Disease. Int. J. Mol. Sci. 2020, 21, 2135. [Google Scholar] [CrossRef] [Green Version]
- Zychlinsky, A.; Prevost, M.C.; Sansonetti, P.J. Shigella flexneri induces apoptosis in infected macrophages. Nature 1992, 358, 167–169. [Google Scholar] [CrossRef]
- Cookson, B.T.; Brennan, M.A. Pro-inflammatory programmed cell death. Trends Microbiol. 2001, 9, 113–114. [Google Scholar] [CrossRef]
- Bergsbaken, T.; Fink, S.L.; Cookson, B.T. Pyroptosis: Host cell death and inflammation. Nat. Rev. Microbiol. 2009, 7, 99–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galluzzi, L.; Vitale, I.; Aaronson, S.A.; Abrams, J.M.; Adam, D.; Agostinis, P.; Alnemri, E.S.; Altucci, L.; Amelio, I.; Andrews, D.W.; et al. Molecular mechanisms of cell death: Recommendations of the Nomenclature Committee on Cell Death 2018. Cell Death Differ. 2018, 25, 486–541. [Google Scholar] [CrossRef]
- Wang, Z.; Yu, H.; Zhuang, W.; Chen, J.; Jiang, Y.; Guo, Z.; Huang, X.; Liu, Q. Cell pyroptosis in picornavirus and its potential for treating viral infection. J. Med. Virol. 2022, 94, 3570–3580. [Google Scholar] [CrossRef]
- Yu, P.; Zhang, X.; Liu, N.; Tang, L.; Peng, C.; Chen, X. Pyroptosis: Mechanisms and diseases. Signal Transduct. Target Ther. 2021, 6, 128. [Google Scholar] [CrossRef]
- Voet, S.; Srinivasan, S.; Lamkanfi, M.; van Loo, G. Inflammasomes in neuroinflammatory and neurodegenerative diseases. EMBO Mol. Med. 2019, 11, e10248. [Google Scholar] [CrossRef]
- Tang, P.; Zhu, R.; Ji, W.P.; Wang, J.Y.; Chen, S.; Fan, S.W.; Hu, Z.J. The NLRP3/Caspase-1/Interleukin-1beta Axis is Active in Human Lumbar Cartilaginous Endplate Degeneration. Clin. Orthop. Relat. Res. 2016, 474, 1818–1826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, G.; Han, X.; Lin, Z.; Qian, H.; Chen, B.; Zhou, C.; Chen, Y.; Jiang, W. Propionibacterium acnes Accelerates Intervertebral Disc Degeneration by Inducing Pyroptosis of Nucleus Pulposus Cells via the ROS-NLRP3 Pathway. Oxid. Med. Cell. Longev. 2021, 2021, 4657014. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; An, R.; Xiang, Q.; Li, G.; Wang, K.; Song, Y.; Liao, Z.; Li, S.; Hua, W.; Feng, X.; et al. Acid-sensing ion channels regulate nucleus pulposus cell inflammation and pyroptosis via the NLRP3 inflammasome in intervertebral disc degeneration. Cell Prolif. 2021, 54, e12941. [Google Scholar] [CrossRef] [PubMed]
- Martinon, F.; Burns, K.; Tschopp, J. The inflammasome: A molecular platform triggering activation of inflammatory caspases and processing of proIL-beta. Mol. Cell 2002, 10, 417–426. [Google Scholar] [CrossRef]
- Broz, P.; Dixit, V.M. Inflammasomes: Mechanism of assembly, regulation and signalling. Nat. Rev. Immunol. 2016, 16, 407–420. [Google Scholar] [CrossRef]
- Burdette, B.E.; Esparza, A.N.; Zhu, H.; Wang, S. Gasdermin D in pyroptosis. Acta Pharm. Sin. B 2021, 11, 2768–2782. [Google Scholar] [CrossRef]
- Hornung, V.; Ablasser, A.; Charrel-Dennis, M.; Bauernfeind, F.; Horvath, G.; Caffrey, D.R.; Latz, E.; Fitzgerald, K.A. AIM2 recognizes cytosolic dsDNA and forms a caspase-1-activating inflammasome with ASC. Nature 2009, 458, 514–518. [Google Scholar] [CrossRef] [Green Version]
- Boucher, D.; Monteleone, M.; Coll, R.C.; Chen, K.W.; Ross, C.M.; Teo, J.L.; Gomez, G.A.; Holley, C.L.; Bierschenk, D.; Stacey, K.J.; et al. Caspase-1 self-cleavage is an intrinsic mechanism to terminate inflammasome activity. J. Exp. Med. 2018, 215, 827–840. [Google Scholar] [CrossRef] [Green Version]
- Kuang, S.; Zheng, J.; Yang, H.; Li, S.; Duan, S.; Shen, Y.; Ji, C.; Gan, J.; Xu, X.W.; Li, J. Structure insight of GSDMD reveals the basis of GSDMD autoinhibition in cell pyroptosis. Proc. Natl. Acad. Sci. USA 2017, 114, 10642–10647. [Google Scholar] [CrossRef] [Green Version]
- Ding, J.; Wang, K.; Liu, W.; She, Y.; Sun, Q.; Shi, J.; Sun, H.; Wang, D.C.; Shao, F. Pore-forming activity and structural autoinhibition of the gasdermin family. Nature 2016, 535, 111–116. [Google Scholar] [CrossRef]
- Ruhl, S.; Shkarina, K.; Demarco, B.; Heilig, R.; Santos, J.C.; Broz, P. ESCRT-dependent membrane repair negatively regulates pyroptosis downstream of GSDMD activation. Science 2018, 362, 956–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newton, K.; Dixit, V.M.; Kayagaki, N. Dying cells fan the flames of inflammation. Science 2021, 374, 1076–1080. [Google Scholar] [CrossRef] [PubMed]
- Kelley, N.; Jeltema, D.; Duan, Y.; He, Y. The NLRP3 Inflammasome: An Overview of Mechanisms of Activation and Regulation. Int. J. Mol. Sci. 2019, 20, 3328. [Google Scholar] [CrossRef] [Green Version]
- Bauernfeind, F.G.; Horvath, G.; Stutz, A.; Alnemri, E.S.; MacDonald, K.; Speert, D.; Fernandes-Alnemri, T.; Wu, J.; Monks, B.G.; Fitzgerald, K.A.; et al. Cutting edge: NF-kappaB activating pattern recognition and cytokine receptors license NLRP3 inflammasome activation by regulating NLRP3 expression. J. Immunol. 2009, 183, 787–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Py, B.F.; Kim, M.S.; Vakifahmetoglu-Norberg, H.; Yuan, J. Deubiquitination of NLRP3 by BRCC3 critically regulates inflammasome activity. Mol. Cell 2013, 49, 331–338. [Google Scholar] [CrossRef] [Green Version]
- Song, N.; Liu, Z.S.; Xue, W.; Bai, Z.F.; Wang, Q.Y.; Dai, J.; Liu, X.; Huang, Y.J.; Cai, H.; Zhan, X.Y.; et al. NLRP3 Phosphorylation is an Essential Priming Event for Inflammasome Activation. Mol. Cell 2017, 68, 185–197.e186. [Google Scholar] [CrossRef] [Green Version]
- Downs, K.P.; Nguyen, H.; Dorfleutner, A.; Stehlik, C. An overview of the non-canonical inflammasome. Mol. Asp. Med. 2020, 76, 100924. [Google Scholar] [CrossRef]
- Aglietti, R.A.; Estevez, A.; Gupta, A.; Ramirez, M.G.; Liu, P.S.; Kayagaki, N.; Ciferri, C.; Dixit, V.M.; Dueber, E.C. GsdmD p30 elicited by caspase-11 during pyroptosis forms pores in membranes. Proc. Natl. Acad. Sci. USA 2016, 113, 7858–7863. [Google Scholar] [CrossRef] [Green Version]
- Man, S.M.; Karki, R.; Kanneganti, T.D. Molecular mechanisms and functions of pyroptosis, inflammatory caspases and inflammasomes in infectious diseases. Immunol. Rev. 2017, 277, 61–75. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Chen, Z.M.; Wu, X.; Zhang, L.; Cao, Y.; Zhou, P. Distinct Molecular Mechanisms Underlying Potassium Efflux for NLRP3 Inflammasome Activation. Front. Immunol. 2020, 11, 609441. [Google Scholar] [CrossRef]
- Yang, D.; He, Y.; Munoz-Planillo, R.; Liu, Q.; Nunez, G. Caspase-11 Requires the Pannexin-1 Channel and the Purinergic P2X7 Pore to Mediate Pyroptosis and Endotoxic Shock. Immunity 2015, 43, 923–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Gao, W.; Shi, X.; Ding, J.; Liu, W.; He, H.; Wang, K.; Shao, F. Chemotherapy drugs induce pyroptosis through caspase-3 cleavage of a gasdermin. Nature 2017, 547, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Rogers, C.; Fernandes-Alnemri, T.; Mayes, L.; Alnemri, D.; Cingolani, G.; Alnemri, E.S. Cleavage of DFNA5 by caspase-3 during apoptosis mediates progression to secondary necrotic/pyroptotic cell death. Nat. Commun. 2017, 8, 14128. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhang, Y.; Xia, S.; Kong, Q.; Li, S.; Liu, X.; Junqueira, C.; Meza-Sosa, K.F.; Mok, T.M.Y.; Ansara, J.; et al. Gasdermin E suppresses tumour growth by activating anti-tumour immunity. Nature 2020, 579, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; He, H.; Wang, K.; Shi, X.; Wang, Y.; Su, Y.; Wang, Y.; Li, D.; Liu, W.; Zhang, Y.; et al. Granzyme A from cytotoxic lymphocytes cleaves GSDMB to trigger pyroptosis in target cells. Science 2020, 368, eaaz7548. [Google Scholar] [CrossRef] [PubMed]
- Kambara, H.; Liu, F.; Zhang, X.; Liu, P.; Bajrami, B.; Teng, Y.; Zhao, L.; Zhou, S.; Yu, H.; Zhou, W.; et al. Gasdermin D Exerts Anti-inflammatory Effects by Promoting Neutrophil Death. Cell Rep. 2018, 22, 2924–2936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, M.A.; Roughley, P.J. What is intervertebral disc degeneration, and what causes it? Spine 2006, 31, 2151–2161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, W.C.; Sze, K.L.; Samartzis, D.; Leung, V.Y.; Chan, D. Structure and biology of the intervertebral disk in health and disease. Orthop. Clin. N. Am. 2011, 42, 447–464. [Google Scholar] [CrossRef]
- Guerrero, J.; Hackel, S.; Croft, A.S.; Hoppe, S.; Albers, C.E.; Gantenbein, B. The nucleus pulposus microenvironment in the intervertebral disc: The fountain of youth? Eur. Cell Mater. 2021, 41, 707–738. [Google Scholar] [CrossRef]
- Antoniou, J.; Steffen, T.; Nelson, F.; Winterbottom, N.; Hollander, A.P.; Poole, R.A.; Aebi, M.; Alini, M. The human lumbar intervertebral disc: Evidence for changes in the biosynthesis and denaturation of the extracellular matrix with growth, maturation, ageing, and degeneration. J. Clin. Investig. 1996, 98, 996–1003. [Google Scholar] [CrossRef] [Green Version]
- Visse, R.; Nagase, H. Matrix metalloproteinases and tissue inhibitors of metalloproteinases: Structure, function, and biochemistry. Circ. Res. 2003, 92, 827–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apte, S.S. A disintegrin-like and metalloprotease (reprolysin-type) with thrombospondin type 1 motif (ADAMTS) superfamily: Functions and mechanisms. J. Biol. Chem. 2009, 284, 31493–31497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sivan, S.S.; Wachtel, E.; Roughley, P. Structure, function, aging and turnover of aggrecan in the intervertebral disc. Biochim. Biophys. Acta 2014, 1840, 3181–3189. [Google Scholar] [CrossRef] [PubMed]
- Nerurkar, N.L.; Elliott, D.M.; Mauck, R.L. Mechanical design criteria for intervertebral disc tissue engineering. J. Biomech. 2010, 43, 1017–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, G.; Shi, C.; Lin, J.; Wang, S.; Wang, H.; Liu, T.; Yang, H.; Li, B. Biomechanics in Annulus Fibrosus Degeneration and Regeneration. Adv. Exp. Med. Biol. 2018, 1078, 409–420. [Google Scholar] [CrossRef]
- Anitua, E.; Padilla, S. Biologic therapies to enhance intervertebral disc repair. Regen. Med. 2018, 13, 55–72. [Google Scholar] [CrossRef] [PubMed]
- Ashinsky, B.; Smith, H.E.; Mauck, R.L.; Gullbrand, S.E. Intervertebral disc degeneration and regeneration: A motion segment perspective. Eur. Cell Mater. 2021, 41, 370–380. [Google Scholar] [CrossRef]
- Roberts, S.; Urban, J.P.; Evans, H.; Eisenstein, S.M. Transport properties of the human cartilage endplate in relation to its composition and calcification. Spine 1996, 21, 415–420. [Google Scholar] [CrossRef]
- Wong, J.; Sampson, S.L.; Bell-Briones, H.; Ouyang, A.; Lazar, A.A.; Lotz, J.C.; Fields, A.J. Nutrient supply and nucleus pulposus cell function: Effects of the transport properties of the cartilage endplate and potential implications for intradiscal biologic therapy. Osteoarthr. Cartil. 2019, 27, 956–964. [Google Scholar] [CrossRef]
- Gullbrand, S.E.; Ashinsky, B.G.; Martin, J.T.; Pickup, S.; Smith, L.J.; Mauck, R.L.; Smith, H.E. Correlations between quantitative T2 and T1rho MRI, mechanical properties and biochemical composition in a rabbit lumbar intervertebral disc degeneration model. J. Orthop. Res. 2016, 34, 1382–1388. [Google Scholar] [CrossRef]
- Chen, Z.H.; Jin, S.H.; Wang, M.Y.; Jin, X.L.; Lv, C.; Deng, Y.F.; Wang, J.L. Enhanced NLRP3, caspase-1, and IL- 1beta levels in degenerate human intervertebral disc and their association with the grades of disc degeneration. Anat. Rec. 2015, 298, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Fu, F.; Bao, R.; Yao, S.; Zhou, C.; Luo, H.; Zhang, Z.; Zhang, H.; Li, Y.; Yan, S.; Yu, H.; et al. Aberrant spinal mechanical loading stress triggers intervertebral disc degeneration by inducing pyroptosis and nerve ingrowth. Sci. Rep. 2021, 11, 772. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Zhang, C.; Zhou, Z.M.; Wang, K.; Gao, J.W.; Qian, Z.Y.; Bao, J.P.; Ji, H.Y.; Cabral, V.L.F.; Wu, X.T. A20 attenuates pyroptosis and apoptosis in nucleus pulposus cells via promoting mitophagy and stabilizing mitochondrial dynamics. Inflamm. Res. 2022, 71, 695–710. [Google Scholar] [CrossRef]
- Wang, Y.; Kanneganti, T.D. From pyroptosis, apoptosis and necroptosis to PANoptosis: A mechanistic compendium of programmed cell death pathways. Comput. Struct. Biotechnol. J. 2021, 19, 4641–4657. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wu, X.; Li, J.; Du, L.; Wang, X.; Cao, J.; Li, H.; Huo, Z.; Li, G.; Pan, D.; et al. Circ_0004354 might compete with circ_0040039 to induce NPCs death and inflammatory response by targeting miR-345-3p-FAF1/TP73 axis in intervertebral disc degeneration. Oxid. Med. Cell Longev. 2022, 2022, 2776440. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yuan, Y.; Huang, Z.X.; Chen, H.; Lan, R.; Wang, Z.; Lai, K.; Chen, H.; Chen, Z.; Zou, Z.; et al. GSDME-mediated pyroptosis promotes inflammation and fibrosis in obstructive nephropathy. Cell Death Differ. 2021, 28, 2333–2350. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Li, S.; Markova, D.Z.; Liang, A.; Kepler, C.K.; Huang, Y.; Zhou, J.; Yan, J.; Chen, W.; Huang, D.; et al. Bromodomain-containing protein 4 inhibition alleviates matrix degradation by enhancing autophagy and suppressing NLRP3 inflammasome activity in NP cells. J. Cell Physiol. 2020, 235, 5736–5749. [Google Scholar] [CrossRef]
- Chen, J.; Yan, J.; Li, S.; Zhu, J.; Zhou, J.; Li, J.; Zhang, Y.; Huang, Z.; Yuan, L.; Xu, K.; et al. Atorvastatin inhibited TNF-alpha induced matrix degradation in rat nucleus pulposus cells by suppressing NLRP3 inflammasome activity and inducing autophagy through NF-kappaB signaling. Cell Cycle 2021, 20, 2160–2173. [Google Scholar] [CrossRef]
- Liao, Z.; Li, S.; Liu, R.; Feng, X.; Shi, Y.; Wang, K.; Li, S.; Zhang, Y.; Wu, X.; Yang, C. Autophagic Degradation of Gasdermin D Protects against Nucleus Pulposus Cell Pyroptosis and Retards Intervertebral Disc Degeneration In Vivo. Oxid. Med. Cell Longev. 2021, 2021, 5584447. [Google Scholar] [CrossRef]
- Le Maitre, C.L.; Pockert, A.; Buttle, D.J.; Freemont, A.J.; Hoyland, J.A. Matrix synthesis and degradation in human intervertebral disc degeneration. Biochem. Soc. Trans. 2007, 35, 652–655. [Google Scholar] [CrossRef]
- Huang, Y.; Peng, Y.; Sun, J.; Li, S.; Hong, J.; Zhou, J.; Chen, J.; Yan, J.; Huang, Z.; Wang, X.; et al. Nicotinamide Phosphoribosyl Transferase Controls NLRP3 Inflammasome Activity through MAPK and NF-kappaB Signaling in Nucleus Pulposus Cells, as Suppressed by Melatonin. Inflammation 2020, 43, 796–809. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Li, S.; Zhang, Y.; Deng, Z.; Wu, J.; Huang, Z.; Qin, T.; Xiao, Y.; Zhou, J.; Xu, K.; et al. Cholesterol Induces Pyroptosis and Matrix Degradation via mSREBP1-Driven Endoplasmic Reticulum Stress in Intervertebral Disc Degeneration. Front. Cell Dev. Biol. 2021, 9, 803132. [Google Scholar] [CrossRef] [PubMed]
- Lawson, L.Y.; Harfe, B.D. Developmental mechanisms of intervertebral disc and vertebral column formation. Wiley Interdiscip. Rev. Dev. Biol. 2017, 6, e283. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.J.; Yu, X.H.; Wang, C.; Yang, W.; He, W.S.; Zhang, S.J.; Yan, Y.G.; Zhang, J. MMPs and ADAMTSs in intervertebral disc degeneration. Clin. Chim. Acta 2015, 448, 238–246. [Google Scholar] [CrossRef]
- Wang, Y.; Che, M.; Xin, J.; Zheng, Z.; Li, J.; Zhang, S. The role of IL-1beta and TNF-alpha in intervertebral disc degeneration. Biomed. Pharmacother. 2020, 131, 110660. [Google Scholar] [CrossRef]
- Brand, F.J., 3rd; Forouzandeh, M.; Kaur, H.; Travascio, F.; de Rivero Vaccari, J.P. Acidification changes affect the inflammasome in human nucleus pulposus cells. J. Inflamm. 2016, 13, 29. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Wang, Y.; Zhang, Y.; Geng, W.; Liu, W.; Gao, Y.; Li, S.; Wang, K.; Wu, X.; Kang, L.; et al. Advanced glycation end products regulate anabolic and catabolic activities via NLRP3-inflammasome activation in human nucleus pulposus cells. J. Cell Mol. Med. 2017, 21, 1373–1387. [Google Scholar] [CrossRef]
- Johnson, Z.I.; Schoepflin, Z.R.; Choi, H.; Shapiro, I.M.; Risbud, M.V. Disc in flames: Roles of TNF-alpha and IL-1beta in intervertebral disc degeneration. Eur. Cell Mater. 2015, 30, 104–116; discussion 107–116. [Google Scholar] [CrossRef]
- Capossela, S.; Schlafli, P.; Bertolo, A.; Janner, T.; Stadler, B.M.; Potzel, T.; Baur, M.; Stoyanov, J.V. Degenerated human intervertebral discs contain autoantibodies against extracellular matrix proteins. Eur. Cell Mater. 2014, 27, 251–263; discussion 263. [Google Scholar] [CrossRef]
- Sun, Z.; Liu, B.; Luo, Z.J. The Immune Privilege of the Intervertebral Disc: Implications for Intervertebral Disc Degeneration Treatment. Int. J. Med. Sci. 2020, 17, 685–692. [Google Scholar] [CrossRef]
- Risbud, M.V.; Shapiro, I.M. Role of cytokines in intervertebral disc degeneration: Pain and disc content. Nat. Rev. Rheumatol. 2014, 10, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Le Maitre, C.L.; Freemont, A.J.; Hoyland, J.A. The role of interleukin-1 in the pathogenesis of human intervertebral disc degeneration. Arthritis Res. Ther. 2005, 7, R732–R745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimbo, K.; Park, J.S.; Yokosuka, K.; Sato, K.; Nagata, K. Positive feedback loop of interleukin-1beta upregulating production of inflammatory mediators in human intervertebral disc cells in vitro. J. Neurosurg. Spine 2005, 2, 589–595. [Google Scholar] [CrossRef]
- Penolazzi, L.; Bergamin, L.S.; Lambertini, E.; Poma, V.V.; Sarti, A.C.; De Bonis, P.; Di Virgilio, F.; Piva, R. The P2X7 purinergic receptor in intervertebral disc degeneration. J. Cell Physiol. 2022, 237, 1418–1428. [Google Scholar] [CrossRef]
- Chen, Z.; Cao, P.; Zhou, Z.; Yuan, Y.; Jiao, Y.; Zheng, Y. Overview: The role of Propionibacterium acnes in nonpyogenic intervertebral discs. Int. Orthop. 2016, 40, 1291–1298. [Google Scholar] [CrossRef] [PubMed]
- He, D.; Zhou, M.; Bai, Z.; Wen, Y.; Shen, J.; Hu, Z. Propionibacterium acnes induces intervertebral disc degeneration by promoting nucleus pulposus cell pyroptosis via NLRP3-dependent pathway. Biochem. Biophys. Res. Commun. 2020, 526, 772–779. [Google Scholar] [CrossRef]
- Stefanakis, M.; Al-Abbasi, M.; Harding, I.; Pollintine, P.; Dolan, P.; Tarlton, J.; Adams, M.A. Annulus fissures are mechanically and chemically conducive to the ingrowth of nerves and blood vessels. Spine 2012, 37, 1883–1891. [Google Scholar] [CrossRef]
- Johnson, W.E.; Sivan, S.; Wright, K.T.; Eisenstein, S.M.; Maroudas, A.; Roberts, S. Human intervertebral disc cells promote nerve growth over substrata of human intervertebral disc aggrecan. Spine 2006, 31, 1187–1193. [Google Scholar] [CrossRef]
- Arkless, K.; Argunhan, F.; Brain, S.D. CGRP Discovery and Timeline. Handb. Exp. Pharmacol. 2019, 255, 1–12. [Google Scholar] [CrossRef]
- Zhang, A.; Wang, K.; Ding, L.; Bao, X.; Wang, X.; Qiu, X.; Liu, J. Bay11-7082 attenuates neuropathic pain via inhibition of nuclear factor-kappa B and nucleotide-binding domain-like receptor protein 3 inflammasome activation in dorsal root ganglions in a rat model of lumbar disc herniation. J. Pain Res. 2017, 10, 375–382. [Google Scholar] [CrossRef]
- Sun, Y.; Leng, P.; Song, M.; Li, D.; Guo, P.; Xu, X.; Gao, H.; Li, Z.; Li, C.; Zhang, H. Piezo1 activates the NLRP3 inflammasome in nucleus pulposus cell-mediated by Ca2+/NF-kappaB pathway. Int. Immunopharmacol. 2020, 85, 106681. [Google Scholar] [CrossRef] [PubMed]
- Mamet, J.; Lazdunski, M.; Voilley, N. How nerve growth factor drives physiological and inflammatory expressions of acid-sensing ion channel 3 in sensory neurons. J. Biol. Chem. 2003, 278, 48907–48913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sen, T.; Samanta, S.K. Medicinal plants, human health and biodiversity: A broad review. Adv. Biochem. Eng. Biotechnol. 2015, 147, 59–110. [Google Scholar] [CrossRef] [PubMed]
- Pantziarka, P.; Pirmohamed, M.; Mirza, N. New uses for old drugs. BMJ 2018, 361, k2701. [Google Scholar] [CrossRef]
- Lim, K.H.; Staudt, L.M. Toll-like receptor signaling. Cold Spring Harb. Perspect. Biol. 2013, 5, a011247. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Cai, X.; Xu, F.; Kang, H.; Li, Y.; Feng, R. Ganoderic Acid A alleviates the degeneration of intervertebral disc via suppressing the activation of TLR4/NLRP3 signaling pathway. Bioengineered 2022, 13, 11684–11693. [Google Scholar] [CrossRef]
- Yang, H.; Wang, H.; Andersson, U. Targeting Inflammation Driven by HMGB1. Front. Immunol. 2020, 11, 484. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.; Guo, Z.; Hou, F.; Fan, W.; Wu, B.; Qian, Z. Magnoflorine Alleviates “M1” Polarized Macrophage-Induced Intervertebral Disc Degeneration through Repressing the HMGB1/Myd88/NF-kappaB Pathway and NLRP3 Inflammasome. Front. Pharmacol. 2021, 12, 701087. [Google Scholar] [CrossRef]
- Li, W.; Deng, M.; Loughran, P.A.; Yang, M.; Lin, M.; Yang, C.; Gao, W.; Jin, S.; Li, S.; Cai, J.; et al. LPS Induces Active HMGB1 Release from Hepatocytes into Exosomes through the Coordinated Activities of TLR4 and Caspase-11/GSDMD Signaling. Front. Immunol. 2020, 11, 229. [Google Scholar] [CrossRef]
- Xu, J.; Jiang, Y.; Wang, J.; Shi, X.; Liu, Q.; Liu, Z.; Li, Y.; Scott, M.J.; Xiao, G.; Li, S.; et al. Macrophage endocytosis of high-mobility group box 1 triggers pyroptosis. Cell Death Differ. 2014, 21, 1229–1239. [Google Scholar] [CrossRef]
- Yilmaz, I.; Akalan, H.; Oznam, K.; Karaarslan, N.; Yasar Sirin, D.; Ozbek, H. Does oseltamivir protect human chondrocyte and nucleus pulposus cells from degeneration by inhibiting senescence and proinflammation mediated by the NLRP3 inflammasome and NF-kappaB? Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 4816–4827. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Tong, Z.; Jiang, S.; Zheng, W.; Zhao, J.; Zhou, X. The Roles of Endoplasmic Reticulum in NLRP3 Inflammasome Activation. Cells 2020, 9, 1219. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.Y.; Luo, Z.P. Paeoniflorin inhibits pyroptosis of nucleus pulposus cells in an acidic environment and alleviates the degeneration of the intervertebral disc in rats. Cell. Signal. 2022, 91, 110243. [Google Scholar] [CrossRef]
- Xu, H.; Dai, Z.H.; He, G.L.; Cai, H.C.; Chen, X.Y.; Chen, Y.L.; Xu, C.; Sheng, S.R. Gamma-oryzanol alleviates intervertebral disc degeneration development by intercepting the IL-1beta/NLRP3 inflammasome positive cycle. Phytomedicine 2022, 102, 154176. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Tardivel, A.; Thorens, B.; Choi, I.; Tschopp, J. Thioredoxin-interacting protein links oxidative stress to inflammasome activation. Nat. Immunol. 2010, 11, 136–140. [Google Scholar] [CrossRef]
- Zhou, Y.; Chen, Z.; Yang, X.; Cao, X.; Liang, Z.; Ma, H.; Zhao, J. Morin attenuates pyroptosis of nucleus pulposus cells and ameliorates intervertebral disc degeneration via inhibition of the TXNIP/NLRP3/Caspase-1/IL-1beta signaling pathway. Biochem. Biophys. Res. Commun. 2021, 559, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Tang, P.; Gu, J.M.; Xie, Z.A.; Gu, Y.; Jie, Z.W.; Huang, K.M.; Wang, J.Y.; Fan, S.W.; Jiang, X.S.; Hu, Z.J. Honokiol alleviates the degeneration of intervertebral disc via suppressing the activation of TXNIP-NLRP3 inflammasome signal pathway. Free Radic. Biol. Med. 2018, 120, 368–379. [Google Scholar] [CrossRef]
- Bofill, A.; Jalencas, X.; Oprea, T.I.; Mestres, J. The human endogenous metabolome as a pharmacology baseline for drug discovery. Drug Discov. Today 2019, 24, 1806–1820. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Gong, Y.; Zheng, X.; Qiu, J.; Jiang, T.; Chen, L.; Lu, F.; Wu, X.; Cheng, F.; Hong, Z. Platelet-Derived Growth Factor-BB Inhibits Intervertebral Disc Degeneration via Suppressing Pyroptosis and Activating the MAPK Signaling Pathway. Front. Pharmacol. 2021, 12, 799130. [Google Scholar] [CrossRef]
- Wang, Y.H.; Li, Y.; Wang, J.N.; Zhao, Q.X.; Jin, J.; Wen, S.; Wang, S.C.; Sun, T. Maresin 1 Attenuates Radicular Pain through the Inhibition of NLRP3 Inflammasome-Induced Pyroptosis via NF-kappaB Signaling. Front. Neurosci. 2020, 14, 831. [Google Scholar] [CrossRef]
- Chen, F.; Jiang, G.; Liu, H.; Li, Z.; Pei, Y.; Wang, H.; Pan, H.; Cui, H.; Long, J.; Wang, J.; et al. Melatonin alleviates intervertebral disc degeneration by disrupting the IL-1beta/NF-kappaB-NLRP3 inflammasome positive feedback loop. Bone Res. 2020, 8, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Qiu, C.; Wang, W.; Peng, J.; Cheng, X.; Shangguan, Y.; Xu, M.; Li, J.; Qu, R.; Chen, X.; et al. Cortistatin protects against intervertebral disc degeneration through targeting mitochondrial ROS-dependent NLRP3 inflammasome activation. Theranostics 2020, 10, 7015–7033. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Xie, G.; Yang, W.; Wang, W.; Zuo, Z.; Wang, W. Platelet-rich plasma attenuates intervertebral disc degeneration via delivering miR-141-3p-containing exosomes. Cell Cycle 2021, 20, 1487–1499. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Xie, C.; Chen, Z.; He, G.; Dai, Z.; Cai, H.; Zhang, H.; Lu, H.; Wu, H.; Hu, X.; et al. MFG-E8 alleviates intervertebral disc degeneration by suppressing pyroptosis and extracellular matrix degradation in nucleus pulposus cells via Nrf2/TXNIP/NLRP3 axis. Cell Death Discov. 2022, 8, 209. [Google Scholar] [CrossRef]
- Jin, J.; Xie, Y.; Shi, C.; Ma, J.; Wang, Y.; Qiao, L.; Li, K.; Sun, T. Lipoxin A4 Inhibits NLRP3 Inflammasome Activation in Rats with Non-compressive Disc Herniation through the JNK1/Beclin-1/PI3KC3 Pathway. Front. Neurosci. 2020, 14, 799. [Google Scholar] [CrossRef]
- Ma, Z.; Tang, P.; Dong, W.; Lu, Y.; Tan, B.; Zhou, N.; Hao, J.; Shen, J.; Hu, Z. SIRT1 alleviates IL-1beta induced nucleus pulposus cells pyroptosis via mitophagy in intervertebral disc degeneration. Int. Immunopharmacol. 2022, 107, 108671. [Google Scholar] [CrossRef]
- Hu, Z.L.; Li, H.Y.; Chang, X.; Li, Y.Y.; Liu, C.H.; Gao, X.X.; Zhai, Y.; Chen, Y.X.; Li, C.Q. Exosomes derived from stem cells as an emerging therapeutic strategy for intervertebral disc degeneration. World J. Stem Cells 2020, 12, 803–813. [Google Scholar] [CrossRef]
- Bowles, R.D.; Setton, L.A. Biomaterials for intervertebral disc regeneration and repair. Biomaterials 2017, 129, 54–67. [Google Scholar] [CrossRef]
- Yuan, X.; Li, T.; Shi, L.; Miao, J.; Guo, Y.; Chen, Y. Human umbilical cord mesenchymal stem cells deliver exogenous miR-26a-5p via exosomes to inhibit nucleus pulposus cell pyroptosis through METTL14/NLRP3. Mol. Med. 2021, 27, 91. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, J.; Zhang, Y.; Liu, W.; Ni, W.; Huang, X.; Yuan, J.; Zhao, B.; Xiao, H.; Xue, F. Mesenchymal stem cells-derived exosomes ameliorate intervertebral disc degeneration through inhibiting pyroptosis. J. Cell Mol. Med. 2020, 24, 11742–11754. [Google Scholar] [CrossRef]
- Xing, H.; Zhang, Z.; Mao, Q.; Wang, C.; Zhou, Y.; Zhou, X.; Ying, L.; Xu, H.; Hu, S.; Zhang, N. Injectable exosome-functionalized extracellular matrix hydrogel for metabolism balance and pyroptosis regulation in intervertebral disc degeneration. J. Nanobiotechnology 2021, 19, 264. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.; Liu, H.; Ma, L.; Lei, J.; Tong, B.; Li, G.; Ke, W.; Wang, K.; Feng, X.; Hua, W.; et al. Engineering Extracellular Vesicles Restore the Impaired Cellular Uptake and Attenuate Intervertebral Disc Degeneration. ACS Nano 2021, 15, 14709–14724. [Google Scholar] [CrossRef] [PubMed]
- Xia, K.S.; Li, D.D.; Wang, C.G.; Ying, L.W.; Wang, J.K.; Yang, B.; Shu, J.W.; Huang, X.P.; Zhang, Y.A.; Yu, C.; et al. An esterase-responsive ibuprofen nano-micelle pre-modified embryo derived nucleus pulposus progenitor cells promote the regeneration of intervertebral disc degeneration. Bioact. Mater. 2023, 21, 69–85. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.S.; Yang, J.M.; Ding, R.J.; Liu, X.Z.; Jiang, X.B.; Yang, Z.J.; Ling, Z.M.; Hu, T.X.; Wei, F.X. Fabrication of a Polylactide-Glycolide/Poly-epsilon-Caprolactone/Dextran/Plastrum Testudinis Extract Composite Anti-Inflammation Nanofiber Membrane via Electrospinning for Annulus Fibrosus Regeneration. J. Biomed. Nanotechnol. 2021, 17, 873–888. [Google Scholar] [CrossRef]
- Vrentas, C.E.; Schaut, R.G.; Boggiatto, P.M.; Olsen, S.C.; Sutterwala, F.S.; Moayeri, M. Inflammasomes in livestock and wildlife: Insights into the intersection of pathogens and natural host species. Vet. Immunol. Immunopathol. 2018, 201, 49–56. [Google Scholar] [CrossRef]
Figure 1.
Pyroptosis pathway. The canonical inflammasome-induced pyroptosis pathway (taken NLRP3 inflammasome pathway) begins with the interaction between receptors and ligands. Then, the NF-κB signal is activated and increases the expression of the NLRP3 inflammasome. Meanwhile, activation signals, including various disturbances, improve the assembly of the NLRP3 inflammasome. Then, the assembled NLPR3 activates caspase-1 to mature IL-18 and IL-1β. The noncanonical inflammasome-induced pyroptosis is mediated by LPS to activate caspase-4/5/11 to cleave GSDMD. In other ways, different cells activate the gasdermin family through caspase-dependent or caspase-independent pathways, which eventually lead to cell expansion, membrane perforation, and the release of cytoplasmic content. IL: interleukin; TNF-α: tumor necrosis factor-α; PAMPs: pathogen-associated molecular patterns; DAMPs: damage-associated molecular patterns; TLRs: toll-like receptors; ROS: reactive oxygen species; ER: endoplasmic reticulum; LPS: lipopolysaccharide; GSDMD: gasdermin D; GzmA: granzyme A; GzmB: granzyme B; CAR-T: chimeric antigen receptor T.
Figure 1.
Pyroptosis pathway. The canonical inflammasome-induced pyroptosis pathway (taken NLRP3 inflammasome pathway) begins with the interaction between receptors and ligands. Then, the NF-κB signal is activated and increases the expression of the NLRP3 inflammasome. Meanwhile, activation signals, including various disturbances, improve the assembly of the NLRP3 inflammasome. Then, the assembled NLPR3 activates caspase-1 to mature IL-18 and IL-1β. The noncanonical inflammasome-induced pyroptosis is mediated by LPS to activate caspase-4/5/11 to cleave GSDMD. In other ways, different cells activate the gasdermin family through caspase-dependent or caspase-independent pathways, which eventually lead to cell expansion, membrane perforation, and the release of cytoplasmic content. IL: interleukin; TNF-α: tumor necrosis factor-α; PAMPs: pathogen-associated molecular patterns; DAMPs: damage-associated molecular patterns; TLRs: toll-like receptors; ROS: reactive oxygen species; ER: endoplasmic reticulum; LPS: lipopolysaccharide; GSDMD: gasdermin D; GzmA: granzyme A; GzmB: granzyme B; CAR-T: chimeric antigen receptor T.

Figure 2.
Changes in normal and degenerative intervertebral discs. The intervertebral disc is composed of nucleus pulposus (NP), annulus fibrosus (AF), and endplate (EP), together constituting a closed buffer system against stress. While in IDD, cell death, ECM degradation, and secondary verification aggravate the vicious cycle, which is closely associated with pyroptosis. ECM: extracellular matrix; ADAMTS: a disintegrin and metalloproteinase with thrombospondin motifs; MMPs: matrix metalloproteinases.
Figure 2.
Changes in normal and degenerative intervertebral discs. The intervertebral disc is composed of nucleus pulposus (NP), annulus fibrosus (AF), and endplate (EP), together constituting a closed buffer system against stress. While in IDD, cell death, ECM degradation, and secondary verification aggravate the vicious cycle, which is closely associated with pyroptosis. ECM: extracellular matrix; ADAMTS: a disintegrin and metalloproteinase with thrombospondin motifs; MMPs: matrix metalloproteinases.
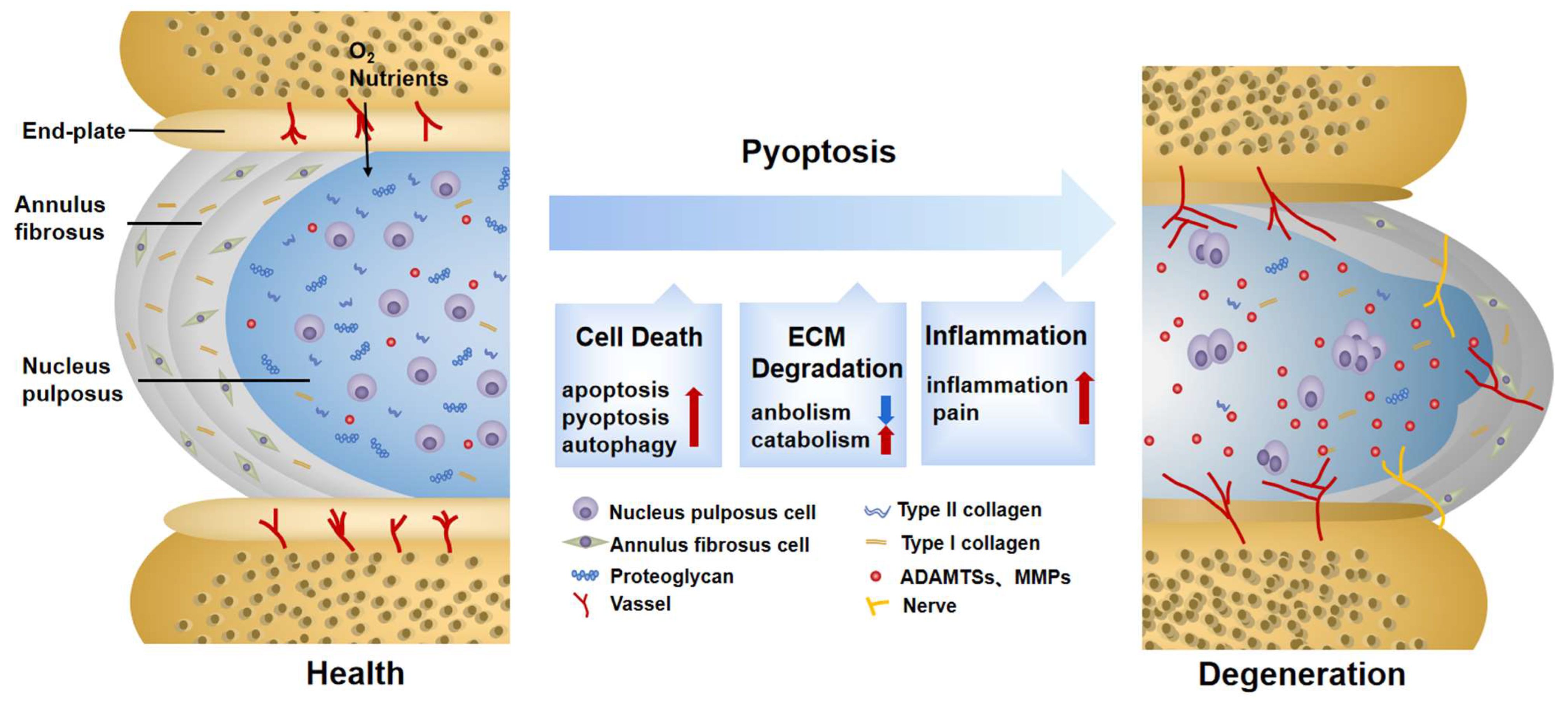
Table 2.
Mechanisms of endogenous molecules for IDD treatment by inactivating pyroptosis.
| Compound | Dose | Model Type | Mechanism | References | ||||
|---|---|---|---|---|---|---|---|---|
| Pathway | Cell Death | ECM | Inflammation | Others | ||||
| Melatonin | 1000 μM | Human (in vitro)/Rat NP tissue (in vivo/in vitro) | Inactivated NAMPT/MAPK/NF-κB/NLRP3 | Suppressed pyroptosis | Upregulated Col II, aggrecan | Suppressed IL-1β, IL-18, TNF-α | —— | [71,111] |
| Platelet-Derived Growth Factor-BB | 50 ng/mL | Rat NP tissue (in vivo/in vitro) | Inactivated MAPK/PI3K/AKT/NLRP3 | Suppressed pyroptosis and apoptosis | Suppressed MMP3, MMP9, ADAMTS4, ADAMTS5. Upregulated Col II, aggrecan | Alleviated IL-1β, IL-18 | —— | [109] |
| Cortistatin | 50 μg/mL | Human (in vitro)/Mice NP tissue (in vitro) | Inactivated NF-κB/NLRP3 | Suppressed apoptosis | Suppressed MMP13, ADAMTS5. Upregulated Col II, aggrecan | Alleviated IL-1β, TNF-α | Alleviated respiratory chain. Suppressed mitochondrial ROS generation | [112] |
| Maresin 1 | 100 ng | Rat NP tissue (in vivo/in vitro) | Inactivated NF-κB/NLRP3 | Suppressed pyroptosis | —— | Alleviated IL-1β, IL-18, TNF-α | Alleviated mechanical allodynia. Improved radicular pain | [110] |
| Platelet-rich plasma | —— | Mice NP tissue (in vivo/in vitro) | Activated the Keap1-Nrf2 | Suppressed pyroptosis | —— | Alleviated IL-1β, IL-18, TGF-β, IL-6 | Alleviated ROS production | [113] |
| A20 | —— | Rat NP tissue (in vivo/in vitro) | Inactivated NF-κB/NLRP3 | Promoted mitophagy. Suppressed pyroptosis and apoptosis | —— | Alleviated iNOS, COX2, TNF-α, IL-1β, IL-6, IL-18 | Stabilized mitochondrial dynamics. Inhibited collapse of mitochondrial membrane potential and ROS | [63] |
| MFG-E8 | 100 ng/ml | Rat NP tissue (in vivo/in vitro) | Inactivated Nrf2/TXNIP/NLRP3 | Suppressed pyroptosis | Suppressed MMP3, ADAMTS5. Upregulated Col II, aggrecan | Alleviated IL-1β, IL-18 | Suppressed ROS and mitochondrial dysfunction | [114] |
| Lipoxin A4 | 10 µL, 100 ng | Rat NP tissue (in vivo/in vitro) | Activated JNK1/Beclin-1/PI3KC3/NLRP3 | Promoted autophagy | —— | Decreased TNF-α, IL-1β, IL-18. Increased IL-4, IL-10, TGF-β | Ameliorated the pain threshold | [115] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Luo, J.; Yang, Y.; Wang, X.; Chang, X.; Fu, S. Role of Pyroptosis in Intervertebral Disc Degeneration and Its Therapeutic Implications. Biomolecules 2022, 12, 1804. https://doi.org/10.3390/biom12121804
AMA Style
Luo J, Yang Y, Wang X, Chang X, Fu S. Role of Pyroptosis in Intervertebral Disc Degeneration and Its Therapeutic Implications. Biomolecules. 2022; 12(12):1804. https://doi.org/10.3390/biom12121804
Chicago/Turabian StyleLuo, Jieya, Yuxuan Yang, Xuan Wang, Xingyu Chang, and Songbo Fu. 2022. "Role of Pyroptosis in Intervertebral Disc Degeneration and Its Therapeutic Implications" Biomolecules 12, no. 12: 1804. https://doi.org/10.3390/biom12121804
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.