The Mode of Action of Cyclic Monoterpenes (−)-Limonene and (+)-α-Pinene on Bacterial Cells


1
State Research Institute of Genetics and Selection of Industrial Microorganisms of the National Research Centre “Kurchatov Institute”, Kurchatov Genomic Center, 117545 Moscow, Russia
2
Institute of Molecular Genetics of the National Research Center “Kurchatov Institute”, 123182 Moscow, Russia
*
Author to whom correspondence should be addressed.
Biomolecules 2021, 11(6), 806; https://doi.org/10.3390/biom11060806
Submission received: 4 April 2021
/
Revised: 24 May 2021
/
Accepted: 26 May 2021
/
Published: 29 May 2021
(This article belongs to the Section Natural and Bio-inspired Molecules)
Abstract
:A broad spectrum of volatile organic compounds’ (VOCs’) biological activities has attracted significant scientific interest, but their mechanisms of action remain little understood. The mechanism of action of two VOCs—the cyclic monoterpenes (−)-limonene and (+)-α-pinene—on bacteria was studied in this work. We used genetically engineered Escherichia coli bioluminescent strains harboring stress-responsive promoters (responsive to oxidative stress, DNA damage, SOS response, protein damage, heatshock, membrane damage) fused to the luxCDABE genes of Photorhabdus luminescens. We showed that (−)-limonene induces the PkatG and PsoxS promoters due to the formation of reactive oxygen species and, as a result, causes damage to DNA (SOSresponse), proteins (heat shock), and membrane (increases its permeability). The experimental data indicate that the action of (−)-limonene at high concentrations and prolonged incubation time makes degrading processes in cells irreversible. The effect of (+)-α-pinene is much weaker: it induces only heat shock in the bacteria. Moreover, we showed for the first time that (−)-limonene completely inhibits the DnaKJE–ClpB bichaperone-dependent refolding of heat-inactivated bacterial luciferase in both E. coli wild type and mutant ΔibpB strains. (+)-α-Pinene partially inhibits refolding only in ΔibpB mutant strain.
1. Introduction
In recent years essential oils have attracted significant scientific interest because they exhibit a broad spectrum of bioactivities, such as antibacterial, antifungal, antiviral, and insecticidal activities [1,2,3]. Moreover, their main active compounds—aldehydes, terpenes, and phenols—are widely used according to the recommendations of the US Food and Drug Administration as food additives, providing a significant reduction in the level of microbial contamination [1,2,4]. Volatile terpenes are derived from the terpene-building units dimethylallyl pyrophosphate and isopentenyl pyrophosphate. However, recent studies have revealed that terpenes are produced not only by plants but also by bacteria, fungi, and amoebae [5,6,7].
Volatile organic compounds (VOCs) limonene and pinene are of great interest in terms of practical application. They are widespread in plants: lemon and other citrus fruits contain limonene, and the main active compound of essential oil obtained from conifers is pinene. These compounds belong to the class of VOCs known as cyclic monoterpenes (Figure 1). Limonene and α-pinene inhibit the growth of Bacillus strains, Staphylococcus aureus, Listeria monocytogenes, Salmonella enterica, Saccharomyces cerevisiae, Zygosaccharomyces rouxii, Sclerotinia sclerotiorum, and Rhizoctonia solani [6,7,8,9,10,11,12]. Some studies on the enantiomers of limonene and pinene have shown that the enantiomeric configuration influences biological activity [10,11,12]. For example, it was revealed that in general, (−)-limonene is more active than (+)-limonene [11], and only the positive enantiomers of pinene have antimicrobial activity against bacterial and fungal cells [10]. The studies mainly aimed to identify these VOCs’ biological activity [12], but their detailed mechanisms of action remain little understood. It was found that limonene, if present in the growth medium during incubation for 4–24 h, destroys the cell membrane of Z. rouxii yeast cells, which is accompanied by leakage of nucleic acids and proteins, as well as the degradation of proteins and their synthesis inhibition [8]. An increase in permeability and degradation of the cell membrane of Gram-negative E. coli and Gram-positive S. aureus bacteria were also shown when they were incubated in a nutrient medium containing finger citron essential oil (FCEO) with limonene (~50%) and also α-pinene as the main components [13]. Biological activities related to limonene and α-pinene demonstrate the importance of these terpenes and their optical isomers as promising therapeutic agents [3,14]. Limonene and α-pinene have been shown to have the potential as enhancers of antituberculosis drugs (ethambutol, rifampicin, and isoniazid) [5].
Luxbiosensors are Escherichia coli cells that contain a hybrid plasmid with two main inserted elements—a regulatory system (a promoter–operator region) and a group of transcriptionally fused reporter genes, luxCDABE. Luxbiosensors are widely used to investigate the functional and ecological role (environmental monitoring) of compounds with different chemical structures [15,16,17,18,19,20,21].
In this paper, we present the results on the reactions of microorganisms after the addition of (−)-limonene and (+)-α-pinene to the nutrient medium. These results are obtained using genetically engineered Escherichia coli strains harboring stress-responsive promoters fused to the bioluminescent reporter gene system of lux operon.
2. Materials and Methods
2.1. Bacterial Strains
Escherichia coli strains used in this study are listed in Table 1. The bacterial E. coli K12 strains MG1655, JW3914-1, JW3933-3, QC868, and QC871 were used to measure the activity of VOCs as inductors of oxidative stress, SOSresponse, and heatshock.
The E. coli strains BW25113 and JW3663 ibpB::kan were used to measure the activity of VOCs as inhibitors of the DnaKJE-dependent refolding.
The E. coli strains MG1655, QC871, JW3914-1, and JW3933-3 were used to measure the antibacterial activity of VOCs.
2.2. Plasmids
The plasmids pIbpA’::lux (for detection of heat shock), pColD’::lux (for detection of SOS response), pKatG’::lux, and pSoxS’::lux (for detection of oxidative stress) were constructed before [20]. The plasmid pFabA’::lux [17,19,21] was used for the detection of membrane damage. The plasmids pMerR’::lux, pCopA’::lux, and pZntA’::lux were used as the negative control [26,27]. Vector pDEW201(Apr), containing the Photorhabdus luminescensluxCDABE genes without a promoter [28], was the parent plasmid for the genetic constructs.
The plasmid pXen7 containing the P. luminescensluxCDABE genes under the lac promoter was used [29] to measure the inhibition of bacterial luciferaseenzymatic activity by (−)-limonene and (+)-α-pinene.
Kinetics and the level of the DnaKJE-dependent refolding of heat-inactivated luciferase were measuredin vivoin E. coli cells containing pLeo1 plasmid. The plasmid pLeo1 is the pUC18 derivative bearing the Photobacterium leiognathi luxCDABE genes encoding the α and β subunits of luciferase and reductase under the control of the lac promoter [30].
The plasmid pLR was used to measure the membrane permeability for D-luciferin—substrate of firefly luciferase. The plasmid pLR (Apr) (kindly provided by Dr. N.N. Ugarova, Moscow State University, Russia) contains the Luciola mingrelicaluc geneunder the Pr promoter of Aliivibrio fischeri lux operon [31].
Constructed hybrid plasmids were introduced into the cells of various strains of E. coli. Excretion of plasmid DNA, restriction and ligation of DNA fragments, and transformation of cells were conducted according to [32].
2.3. Nutrient Media and Growth Conditions
Bacterial strains were cultivated in the liquid Luria–Bertani (LB) medium or on Petri dishes with agarized LB medium (LA) supplemented with appropriate antibiotics. The cells were grown under aeration at 30 °C until the early exponential phase. Antibiotics were added at the following concentrations: streptomycin (Sm; 25 µg/mL), kanamycin (Km; 20 µg/mL), ampicillin (Ap; 100 µg/mL), chloramphenicol (Cm; 25 µg/mL).
2.4. Enzymes and Chemicals
All the chemicals were of analytical grade. Antibiotics (streptomycin, kanamycin, chloramphenicol, and ampicillin) were obtained from Sigma Chemical Co (St. Louis, MO, USA), Merck (Darmstadt, Germany), and Biopharm (Moscow, Russia). Methyl viologen (paraquat), mitomycin C, dimethyl sulfoxide (DMSO), D-luciferin, and VOC compounds (−)-limonene (96% purity, CAS # 5989-54-8) and (+)-α-pinene (98% purity, CAS # 80-56-8) were purchased from Sigma-Aldrich Chimie GmbH (Steinheim, Germany). Triton X-100 was purchased from ServaElectrophoresis GmbH (Heidelberg, Germany), andethanol (96%) was purchased from LLC Donskoy (Tula region, Kimovsky district, rp. Epifan, Russia). Hydrogen peroxide (H2O2), HgCl2, and ZnSO4 were purchased from Merck (Darmstadt, Germany), CuSO4 was purchased from Sigma-Aldrich Chimie GmbH (Steinheim, Germany).
We used an uncoupler of oxidative phosphorylation protonophore carbonyl cyanide—3-chlorophenylhydrazone (CCCP)—to reduce the ATP content in cells. CCCP was obtained from Sigma-Aldrich Chimie GmbH (Steinheim, Germany). ATP concentration was assayed with firefly luciferase [33].
The required concentration of stock solutions of paraquat, H2O2, Triton X-100, and HgCl2was obtained by dissolving the compounds mentioned above in distilled water.
The enzymes used in this work were provided by Fermentas (Vilnius, Lithuania). According to manufacturer protocol, the cell lysate containing the firefly luciferase was obtained using the Dual-Luciferase Reporter Assay System (Promega, Madison, WI, USA).
All substances were stored at a temperature specified by the manufacturer. All test solutions were prepared just before their use.
2.5. Measurement of Production and Activity of Bacterial Luciferase
Bacterial luciferase (E) catalyzes the oxidation of long-chain aldehydes (RCHO) by atmospheric oxygen (O2) in the presence of reduced flavin mononucleotide (FMNH2):
FMNH2 + RCHO + O2→ FMN + RCOOH + H2O + a quantum of light (λmax = 490 nm)
Recombinant luminescent bacteria (luxbiosensors) were pregrown at 30 °C in LB medium supplemented with appropriate antibiotics overnight. Then the culture was diluted up to 107 CFU/mL by fresh LB and grown at 30 °C under aeration until the early exponential phase. The cell aliquots (200 μL)were placed in special cuvettes, one serving as a control to which 4 µL of DMSO was added, while 4 µL of (−)-limonene or (+)-α-pinene in DMSO solution in various concentrations were added in the others. The prepared samples with lux-biosensor cells were placed in front of a photomultiplier in the Luminometer Photometer LMA01 (Beckman Coulter, Praha, Czech Republic). After selected time intervals, the intensity of the bioluminescence of the cellular suspension was measured. Luminescence values were expressed in terms of the instrument’s arbitrary relative light units (RLU). The samples were incubated at room temperature. Three main parameters that characterize the quality of the luxbiosensor were estimated: response amplitude (RA) or induction factor (R) (if the luminescence intensity of the control preparation is practically constant in the time interval 0–t), the minimal response time (tmin), and threshold sensitivity (P)—the concentration of the inductor when RA or R is approximately equal to 2. The RA was determined by the formula RA = It− I0/Ik− I0, where I0 is the intensity of the preparation bioluminescence at the moment of inductor addition (t = 0), Ik is the intensity of the bioluminescence of the control preparation (in the absence of the inductor) at moment t, and It is the intensity of the bioluminescence of the test preparation at moment t. Induction factor R = It/I0.
For induction of luxbiosensors with the PkatG, PsoxS, PcolD, PfabA, and PibpA promoters, various concentrations of H2O2, paraquat (stimulate the formation of superoxide anion radical (O2.−)), mitomycin C (forms both DNA adducts and interstrand cross-links [34]), Triton X-100, and ethanol, respectively, were used as a positive control.
2.6. Thermal Inactivation and Refolding
(−)-Limonene and (+)-α-pinene were placed in small plastic tubes (volume 1 mL) containing bacterial cells (2–3 × 108 CFU/mL) in LB medium. The tubes were tightly sealed with two layers of Parafilm M (Pechiney Plastic Packaging Company, Chicago, IL, USA). The controls were performed in the absence of terpenes. For in vivo heatinactivation of Photobacterium leiognathi luciferase, bacterial cells were transferred to 46 °C in a water bath for 5 min. The production of luciferase and heat shock proteins was stopped by adding chloramphenicol to a final concentration of 167 μg/mL. For subsequent recovery (refolding) of P. leiognathi luciferase, bacterial cells were transferred back to lower temperatures (22 °C). The cell aliquots (200 μL) were transferred to a luminometer in which bioluminescence intensity was measured as a function of the incubation time, and the values were plotted as a percentage of the initial activity. For the highest DnaKJE and ClpB levels, E. coli cells were first incubated at 42 °C for 30 min without chloramphenicol (preliminary “heat shock”). All experiments were repeated four to six times with two to three tubes per experiment.
2.7. Antibacterial Activity
Antibacterial activity of (−)-limonene and (+)-α-pinene was tested using the agar diffusion method [35]. First, 100 µL of bacterial suspension (107 CFU/mL) was spread on nutrient agar (NA) medium. Then, a sterile filter paper disc (diameter = 6 mm) containing 4 µL (3.36 mg) of the (sample) (−)-limonene was placed on the surface of the plate. All the plates incubating at 37 °C for 18 h were observed for zones of growth inhibition. The inhibition zones were measured in millimeters from the circumference of the discs to the circumference of the inhibition zone. All the assays were carried out in triplicate.
3. Results
3.1. The Action of (−)-Limonene and (+)-α-Pinene on the LuxBiosensors
3.1.1. Oxidative Stress
The induction of oxidative stress by VOCs, hydrogen peroxide, and paraquat is shown in Figure 2, Figure 3 and Figure 4. The experiment was carried out with the luxbiosensors responsive to oxidative stress—E. coli MG1655 (pKatG’::lux) and MG1655 (pSoxS’::lux) strains. Different concentrations of H2O2 increased the bioluminescence intensity of E. coli MG1655 (pKatG’::lux) (Figure 2A).
Data shown in Figure 2, Figure 3, Figure 4, Figure 5, Figure 6 and Figure 7 are mean values of light emission units (RLU) in at least four experiments. Standard deviations from the mean in these experiments did not exceed 25%.
The induction factor (R) gradually increases up to 6–7 times at 5 μM of H2O2, up to 15–20 times at 25 μM, up to 40 times at 50 μM, and up to 60 times at a concentration of 100 μM. The minimal incubation time required for inducing luminescence (tmin) is ~10 min. It should be noticed that the maximum of induction factor R for all concentrations is reached at ~35–40 min, and then it almost does not decrease. The action of different concentrations of (−)-limonene (1–100 μM) is shown in Figure 2B. (−)-Limonene at concentration 1 μM (P—threshold sensitivity) increases bioluminescence by 2–3 times. Initially, a slight decrease in bioluminescence by 1.5 times at concentration 5 μM and by 5–6 times at 100 μM is observed. Then, with the increase in incubation time, the luminescence rapidly increases at 5 μM up to 80 times compared to the control (which is comparable to the action of ~50–100 μM of H2O2, Figure 2A) and after 50 min reaches a plateau. The induction factor R decreases at higher concentrations of (−)-limonene, e.g., at 100 μM, R = 15. Different concentrations of (+)-α-pinene (up to 100 μM) do not induce the PkatG promoter (Figure 2C).
The curves of induction of the PkatG promoter by (−)-limonene in the wild-type strain (E. coli MG1655 (pKatG’::lux)) and the ΔkatG mutant strain (E. coli JW3914-1 ΔkatG (pKatG’::lux)) are shown in Figure 3.
There is a significant increase in the induction factor R in the ΔkatG mutant strain compared to the wild-type strain. (−)-Limonene at concentration 1 μM causes increased luminescence intensity by ~3 times in the wild-type strain and 10–15 times in the ΔkatG mutant. The higher concentration of (−)-limonene (10 μM) causes the same patterns. A high level of induction of the PkatG promoter under the action of (−)-limonene, comparable to the effect of hydrogen peroxide, is observed. Thus, the absence of catalase enzyme in the bacterial cell in strain JW3914-1 ΔkatG729::kan leads to the significant increase in the induction factor R in the ΔkatG mutant strain compared to wild-type strain and indicates that the mechanism of action of (−)-limonene is associated with the formation of hydrogen peroxide in this process.
The data on the action of different concentrations of (−)-limonene and (+)-α-pinene on the PsoxS promoter are represented in Figure 4.
Different concentrations of paraquat (methyl viologen) increase bioluminescence intensity of E. coli MG1655 (pSoxS’::lux) (Figure 4A). The induction factor R gradually increases over2 h: up to 10 times at a concentration of 5 μM of paraquat, up to 80–90 timesat 50 μM, and up to 100 times at 100 μM. The minimal incubation time required for inducing luminescence (tmin) is ~20–25 min. The action of different concentrations of (−)-limonene is shown in Figure 4B. The minimum used concentration of (−)-limonene (1 μM) is the same as in the case of induction of the PkatG promoter. However, the induction factor R of E. coli MG1655 (pSoxS’::lux) is lower compared to the R of E. coli MG1655 (pKatG’::lux): R does not exceed 10 times at 5 and 10 μM of (−)-limonene, and at 100 μM, at first, the luminescence decreases by about an order of magnitude and then starts to increase, crosses the control line (luminescence values of strain without VOCs) after 70 min, and after 2 h of incubation exceeds the control by ~5 times. The action of different concentrations of (+)-α-pinene does not induce the PsoxS promoter. However, at a high concentration (100 μM), it reduces the intensity of bacterial luminescence by ~2–3 times (Figure 4C).
Summarizing the obtained data on the induction of oxidative stress by VOCs, we can conclude that in the bacterial cells, (−)-limonene promotes the synthesis of a significant amount of reactive oxygen species (ROS)—hydrogen peroxide and superoxide anion radical. (+)-α-Pinene is not active in this respect.
3.1.2. SOSResponse
The data on the action of different concentrations of (−)-limonene and (+)-α-pinene on the SOSpromoter (PcolD—DNA damage) of the plasmid pColD are presented in Figure 5.
Mitomycin C at a concentration of 10 μM increases bioluminescence intensity of E. coli MG1655 (pColD’::lux): the induction factor R gradually increases up to 10 times, with a tminof ~40 min (Figure 5). (−)-Limonene induces the PcolD promoter, starting from a concentration of 5 μM, and, like mitomycin C, after ~40 min of incubation. At higher concentrations (25–100 μM), at first, the luminescence intensity decreases within 20–30 min of incubation up to 5–6 times and then increases and reaches a plateau after 2 h. The induction factor Rrises no more than 20 times at concentrations of (−)-limonene ≤ 100 μM. (+)-α-Pinene does not induce the PcolD promoter at concentrations ≤ 100 μM.
3.1.3. Cell Membrane Damage
The data on the action of different concentrations of (−)-limonene (2, 4, and 10 µM) on the PfabA promoter (cell membrane damage) are represented in Figure 6A.
Different concentrations of Triton X-100—0.5, 2, and 4.0 µg/mL(used as a positive control [17])—increase bioluminescence intensity of E. coli MG1655 (pFabA’::lux). The response amplitude RA gradually increases up to 3–4 times at a concentration of 2 µg/mLafter 2.5 h of incubation (Figure 6A). (−)-Limonene significantly decreases the luminescence intensity of E. coli MG1655 (pFabA’::lux) in the first minutes of incubation, but then after ~60–70 min of incubation at low concentrations (less than 5 µM), the luminescence intensity begins to gradually recover up to the control level (luminescence of strain without VOCs). At higher concentrations of (−)-limonene, inhibition of luciferase enzymatic activity does not allow evaluating the ability of this VOC to activate the PfabA promoter (Figure 6A).
(−)-Limonene does not induce the activity of the PmerR (Figure 6B), PcopA, and PzntA promoters (data not shown), which specifically react exclusively to mercury, copper, and zinc ions, respectively (E. coli MG1655 (pMerR’::lux), MG1655 (pCopA’::lux) and MG1655 (pZntA’::lux) are used as a negative control [26,27]).
3.1.4. HeatShock
The data on the action of different concentrations of (−)-limonene and (+)-α-pinene on the PibpA promoter (protein damage, heat shock) are represented in Figure 7.
Ethanol (4%) (used as a positive control [15])increases the bioluminescence intensity of E. coli MG1655 (pIbpA’::lux). The response amplitude RA gradually increases up to 10 times and reaches a plateau after ~40–45 min of incubation, with a tminof ~25 min (Figure 7A). The action of (−)-limonene leads to the induction of heat shock (Figure 7A). At the minimum concentration (P = 5 μM) of (−)-limonene, the luminescence begins to increase after ~60 min of incubation and then gradually increases by only 1.5–2 times. At 10 μM, the luminescence gradually increases by 8–10 times; at 15–20 μM, the luminescence begins to increase after 45 min of incubation and then increases up to 20 times after 90–100 min of incubation (Figure 7A).
(+)-α-Pinene induces the PibpA promoter. The induction begins to increase after ~60 min of incubation; Cmin = 5 μM. At 10 μM, the luminescence increases by 3 times; at 20 μM, the luminescence increases by up to 10 times (Figure 7B). However, the action of (+)-α-pinene on the induction of bioluminescence intensity of E. coli MG1655 (pIbpA’::lux) is weaker compared to the action of (−)-limonene.
3.1.5. Effect of VOCs on the Enzymatic Activity of Native Bacterial Luciferase P. luminescens
The decrease in the intensity of cell luminescence is observed in the initial moment after adding (−)-limonene, and to a lesser extent (+)-α-pinene, to luxbiosensors (Figure 3, Figure 4B, Figure 5A, Figure 6 and Figure 7). In this connection, the action of (−)-limonene and (+)-α-pinene on luciferase enzymatic activity, contained in the bacterial cell, was studied. For this purpose, the E. coli MG1655 (pXen7) strain was used. The plasmid pXen7 contains P. luminescens luxCDABE genes under lac promoter that provide constitutive expression of lux genes [29]. The initial luminescence level of the cell suspension in the exponential phase is about 10,000 RLU (Figure 8).
After (−)-limonene is added into the cell suspension at a concentration of less than 5 μM, the luminescence intensity of E. coli MG1655 (pXen7) strain almost does not decrease during subsequent incubation at room temperature. However, a significant decrease in luminescence intensity is observed at concentrations of 10 μM of (−)-limonene and especially at 100 μM, by 10 and 100 times, respectively (Figure 8). Furthermore, simple dilution or centrifugation of the bacterial suspension resulted in the recovery of 80–100% of the initial luminescence intensity (within experimental error). These data indicate that the inhibition effect seen at high concentrations of (−)-limonene was fully reversible. Consequently, the inhibition of luciferase enzymatic activity by (−)-limonene can be explained by the competition of these VOCs with the luciferase substrate, long-chain aldehyde, or FMNH2.
(+)-α-Pinene at concentrations up to 100 μM does not decrease the intensity of cell luminescence (Figure 8). It can be assumed that the insignificant inhibitory effect of (+)-α-pinene is determined by low solubility (2.49 mg/L) and weak penetration into the cell cytoplasm due to the connection of (+)-α-pinene with the membrane. This assumption is supported by the data obtained using cell lysate containing the firefly L. mingrelica luciferase (see Section 3.3).
3.2. Effect of (−)-Limonene and (+)-α-Pinene on the DnaKJE-Dependent Refolding of Heat-Inactivated Bacterial Luciferase
It was shown that such VOCs as ketones 2-heptanone, 2-nonanone, and 2-undecanone effectively inhibit the DnaKJE-dependent refolding of heat-inactivated bacterial luciferases only in the E. coli ΔibpB mutant strain lacking small chaperone IbpB [36]. Chaperone IbpB, which forms a complex with hydrophobic sites in proteins [37,38,39], appears to inhibit the complexation of these sites with hydrophobic ketones. Therefore, it was of interest to determine the ability of terpenes (−)-limonene and (+)-α-pinene to inhibit the DnaKJE-dependent refolding of proteins depending on the presence of small chaperone IbpB in the cell. Kinetics and the level of the DnaKJE-dependent refolding of heat-inactivated luciferase were measuredin vivoin the wild-type strain(E. coli BW25113) and the ΔibpB mutant strain (E. coli JW3663 ibpB::kan), containing plasmid pLeo1 with P. leiognathilux CDABE genes under control of the lac promoter, that provide constitutive expression of thesegenes.
The effects of (−)-limonene, (+)-α-pinene, and CCCP on refolding kinetics of the heat-inactivated P. leiognathi luciferase in the wild-type strain (E. coli BW25113 (pLeo1)) and the ΔibpB mutant strain (E. coli JW3663 ibpB::kan (pLeo1)) are shown in Figure 9.
Protonophore CCCP (3-chlorophenylhydrazone) was used as a positive control. The presence of protonophore CCCP (50 µM) in the medium leads to a decrease in the intracellular concentration of ATP to almost a minimum during the first few minutes and at the same time completely inhibits DnaKJE-dependent refolding of heat-inactivated luciferase both in the wild-type strain (Figure 9A) and the ΔibpBmutant (Figure 9B). The action of (−)-limonene (10 µM) causes the same patterns (Figure 9). (+)-α-Pinene (20 µM), like ketones, only partially inhibits refolding in the ΔibpB mutant strain (Figure 9B). However, (−)-limonene (10 µM) shows a significantly higher ability to compete with the small chaperone IbpB for binding to hydrophobic sites in the denatured macromolecule and the DnaKJE chaperone since it completely inhibits refolding not only in the ΔibpB mutant strain but also in the wild strain BW25113 (Figure 9A).
3.3. Effect of (−)-Limonene and (+)-α-Pineneon Firefly Luciferase Activity
(−)-Limonene inhibits DnaKJE-dependent refolding, regardless of the presence of small chaperone IbpB in the cell, as entirely as the protonophore CCCP. Therefore it could be assumed that (−)-limonene, like the protonophore CCCP, decreases the intracellular concentration of ATP (ATP is a necessary factor for refolding with the ATP-dependent chaperones). The firefly L. mingrelica luciferase, whose enzymatic activity is utterly dependent on the presence of ATP, was used to test this hypothesis: ATP + O2+ D-luciferin → AMP + PPi+ CO2 + oxyluciferin + light (λmax = 566 nm). The data on the action of different concentrations of (−)-limonene and (+)-α-pinene on the firefly luciferase luminescence intensity in E. coli MG1655 (pLR) strain and cell lysate are represented in Figure 10. The plasmid pLR contains luc gene encoding the L. mingrelica luciferase under the Pr promoter of A. fischeri lux operon. The luminescence intensity gradually increased at all concentrations of (−)-limonene: the response amplitude (RA) at a concentration of 10 μM increases up to 5 and ~30 times after 10 and 30 min of incubation, respectively (Figure 10A). In comparison, the presence of protonophore CCCP (50 µM; used as a positive control) in the medium leads to a sharp decrease in luminescence intensity in E. coli MG1655 (pLR) (up to ~50–100 times from the initial level), which directly indicates a reduction in the intracellular concentration of ATP (data not shown). Therefore, the data show the absence of influence of (−)-limonene on the intracellular content of ATP in the early stages of action (in a period from 0 to 60 min of incubation).
The increase in the luminescence intensity of cells after adding (−)-limonene and (+)-α-pinene requires explaining this phenomenon. We assume that (−)-limonene and (+)-α-pinene, acting on cell membranes, increase their permeability for D-luciferin—a substrate of the luciferase, which very weakly penetrates bacterial membranes at pH 7.5 [40,41,42]. Confirmation of this assumption was obtained by adding (−)-limonene and (+)-α-pinene to firefly luciferase in anin vitroexperiment with E. coli MG1655 (pLR) living and lysed cells (Figure 10).
(−)-Limonene induces a gradual bioluminescence increase in E. coli MG1655 (pLR) cells without reducing the luminescence intensity of the cell lysate already at a concentration of 1 μM. However, at the concentration of 10 μM and especially at 100 μM, significant inhibition of luciferase enzymatic activity in the cell lysate is observed (Figure 10A). Different concentrations of (+)-α-pinene as well as (−)-limonene increase luminescence intensity in E. coli MG1655 (pLR) and, at the same time, decrease the luminescence intensity of cell lysates (Figure 10B). However, the activity of (+)-α-pinene is significantly lower than that of (−)-limonene.
3.4. Effect of (−)-Limonene and (+)-α-Pinene on the Growth of Bacteria
Antibacterial activity of (−)-limonene and (+)-α-pinene was tested using the agar diffusion method [13]. DIZs (diameters of inhibition zone) of different E. coli strains for (−)-limonene (in an amount of 3.36 mg (4 µL per sterile filter paper disc))are presented in Table 2. In this experiment, E. coli strains JW3914-1 ΔkatG729::kan and JW3933-3 ΔoxyR749::kan were used to determine the involvement of antioxidant enzymes in cell resistance to the action of VOCs. The obtained data (Table 2) demonstrate that the inhibitory effect of (−)-limonene is significantly enhanced in the absence of catalase and peroxidase enzymes in the bacterial cell in strain JW3914-1 ΔkatG729::kan, and especially in JW3933-3 ΔoxyR749::kan, which indicates the role of hydrogen peroxide in this process. (+)-α-Pinene in an amount of ≤5 mg (≤6 µL) almost does not inhibit the growth of E. coli strains (data not shown).
4. Discussion
It was shown that cyclic terpene limonene, when present in the growth medium during incubation for several hours, destroys the cell membrane in yeast cells (Z. rouxii) and bacterial cells (E. coli and S. aureus), which is accompanied by leakage of nucleic acids and proteins [8,13]. Moreover, the stereochemistry of limonene and α-pinene influences antimicrobial activity: in general, (−)-limonene and (+)-α-pinene are more active, and their antimicrobial activity is pathogen-specific [10,11,12]. Highly sensitive specific lux-biosensors were used in the present work to study the effects of (−)-limonene and (+)-α-pinene and determine the mechanism of action of the above-mentioned VOCs on bacterial cells. We found that starting from a concentration of 1 µM, (−)-limonene induces synthesis of considerable amounts of H2O2 in the bacteria within the first 20–30 min of incubationand induces the synthesis of superoxide anion radicals after 30 min. Moreover, this VOC causes damage to DNA and proteins after 40–50 min of action. We assume that the induction of oxidative stress, observed at the first stage (minutes) of the interaction of (−)-limonene with bacterial cells and associated with the synthesis of reactive oxygen species (ROS)—hydrogen peroxide and superoxide anion radical—is of considerable interest to researchers who use terpene derivatives. Previous studies have determined the reaction of cells to limonene at the late stage (hours) and recorded the damage to membranes. However, when using limonene as an inhibitor of yeast and bacteria in preparations of nutritious juices and other food products, it is necessary to know that limonene not only inhibits bacteria and yeast (i.e., it acts as an antibiotic on a specific target) but also induces the formation of ROS that can be dangerous to the human body. A similar situation arose in the study of the action of antibiotics when J. J. Collins et al. (USA) [43,44,45] found that antibiotics, aside from acting on the primary target (membranes, DNA-gyrase, etc.), induce the formation of a significant amount of ROS in the cell. This caused a great interest in the problem since ROS, penetrating human cells, cause DNA damage, mutations, and cancer degenerations.
As reported in [46], intracellular targets of 2,4-diacetylphloroglucinol action were assessed using bacterial biosensors with inducible bioluminescence corresponding to DNA and protein damage. However, unlike our study, it was not possible to register a positive response from any biosensor. As a result, the bactericidal effect of 2,4-DAPG is believed to be related to the destruction of bacterial barrier structures [46].
The data obtained in this work show that the observed effects (oxidative stress, SOS response, etc.) are unique since the investigated range of concentrations of (−)-limonene (<10 μM) weakly inhibits the enzyme-reporter activity of luciferase. Inhibition of the luciferase enzymatic activity by (−)-limonene observed at concentrations ≥10 μM (Figure 8) is reversible. It can be explained by the competition of VOC with the luciferase substrate, long-chain aldehyde, and/or FMNH2. This effect should be taken into account when working with luxbiosensors. In vitro, the reversible inhibition of the bacterial luciferase was determined for several compounds competitive with FMNH2 and long-chain aldehyde [47,48,49].
Summarizing the obtained data on the induction of oxidative stress by studied VOCs, it can be concluded that only (−)-limonene in a bacterial cell contributes to the synthesis of a significant amount of hydrogen peroxide and superoxide anion radical. On the other hand, the mechanism of the formation of reactive oxygen species (ROS) remains unclear. Apparently, for (+)-α-pinene, the mechanism of action on bacterial strains does not relate to forming ROS.
The method of using bacteria strains containing eukaryotic luciferase Pyrophorus plagiophtalamus (click beetle) activated by D-luciferin to measure membrane permeability was first applied in the analysis of the action of low-molecular membrane-lytic agents—cationic peptides—that form pores in biological membranes [50]. It was shown that melittin produced by Apis mellifera bees and the antibiotics polymyxin B and gramicidin S, at concentrations in the range of 10–100 µg/mL, completely inhibit the growth of E. coli MC1061 bacteria when seeding on Petri dishes. Furthermore, a significant increase in the luminescence of cells containing firefly luciferase was observed at the same concentrations, indicating an increase in the membrane permeability for D-luciferin [50]. However, unlike (−)-limonene, which gradually induces bioluminescence increase in cells within 30–50 min of incubation (Figure 10A), cationic peptides cause an increase in the luminescence of the cell suspension within a few seconds after being added to the cells, the passage of luminescence through a maximum within 20–30 s, and a relatively rapid decrease in luminescence over the next 2 min. Therefore, in the case of (−)-limonene, a much more moderate, prolonged effect of the toxic agent on the bacterial membrane structure is observed. Besides, the action of (−)-limonene does not lead to a decrease in the intracellular content of ATP compared to the mechanisms of cationic peptides and CCCP action.
The response of the luxbiosensor with the PibpA promoter, which fixes the heat shock, to the action of (−)-limonene and (+)-α-pinene is of particular interest and is shownfor the first time. As can be seen from the obtained data presented in Figure 7, (+)-α-pinene induces the opening of the heat shock promoter PibpA only during the longer incubation time compared to the action of (−)-limonene. However, (+)-α-pinene does not induce ROS formation in the cell (Figure 2 and Figure 3). (+)-α-Pinene, like (−)-limonene, inhibits refolding carried out by the DnaKJE chaperone, but only partially and only in the ΔibpB mutant strain lacking the small chaperone IbpB (Figure 9). These results are consistent with the data on the effects of ketones 2-heptanone, 2-nonanone, and 2-undecanone that were reported by us earlier [36]. It can be assumed that (−)-limonene and (+)-α-pinene, as hydrophobic compounds, are capable of complexing with the corresponding regions of the DnaKJE, hindering the chaperone protein, and inhibiting its ability to form a complex not only with denatured proteins but also with σ32 (the subunit σ32 is responsible for the synthesis of “heat shock” promoters [51,52,53,54]).
The effect of complete inhibition of the ATP-dependent chaperone (DnaKJE) activity by (−)-limonene should also be of considerable interest. Currently, some laboratories are searching for organic molecules that can inhibit ATP-dependent chaperones and exhibit an anticancer effect [55,56,57]. Several thousands of synthesized organic compounds have been tested, and only a few of them can hinderchaperones to some extent. However, as a rule, such compounds are toxic to the human body. In this work, we found that a natural product contained in citrus fruits completely and at very low concentrationsinhibits the DnaK-dependent refolding, which can be of practical use.
5. Conclusions
The initial stage of the action of (−)-limonene on the cell is the induction of oxidative stress, namely the formation of a significant amount of ROS (hydrogen peroxide and superoxide anion radical), which damages DNA (SOS response). At the same time, bacterial membranes are damaged, which leads to an increase in their permeability. Moreover, heat shock is induced by direct contact of (−)-limonene and (+)-α-pinene with the DnaKJE–σ32 complex.
The action of (−)-limonene at high concentrations and prolonged incubation time makes degrading processes in cells irreversible. This leads to the destruction of the membrane and the release of nucleic acids and proteins outside, which results in the lysis of bacteria.
Author Contributions
Conceptualization, G.B.Z.; methodology, O.E.M. and G.B.Z.; formal analysis, O.E.M., V.A.P. and I.A.K.; investigation, O.E.M.; writing—original draft preparation, O.E.M. and G.B.Z.; writing—review and editing, O.E.M., V.A.P., I.A.K. and G.B.Z.; visualization, V.A.P. and O.E.M.; funding acquisition, G.B.Z.; resources, G.B.Z.; supervision, I.A.K. and G.B.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported in part by the Russian Foundation for Basic Research (grant No. 19-04-00495), the Ministry of Science and Higher Education of the Russian Federation (Grant No. 075-15-2019-1658), and funds within the state assignment of NRC “Kurchatov Institute”—IMG for 2020–2021 (No. 121030200227-6).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The author elects not to share data.
Acknowledgments
The work was carried out using the strains of the Bioresource Center—All-Russian Collection of Industrial Microorganisms.
Conflicts of Interest
Authors declare no conflicts of interest.
References
- Böhme, K.; Barros-Velázquez, J.; Calo-Mata, P.; Aubourg, S.P. Antibacterial, Antiviral and Antifungal Activity of Essential Oils: Mechanisms and Applications. In Antimicrobial Compounds; Springer: Berlin/Heidelberg, Germany, 2014; pp. 51–81. [Google Scholar] [CrossRef]
- Burt, S. Essential Oils: Their Antibacterial Properties and Potential Applications in Foods—A Review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef]
- Mercier, B.; Prost, J.; Prost, M. The Essential Oil of Turpentine and Its Major Volatile Fraction (α- and β-Pinenes): A Review. Int. J. Occup. Med. Environ. Health 2009, 22, 331–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bevilacqua, A.; Corbo, M.R.; Sinigaglia, M. In Vitro Evaluation of the Antimicrobial Activity of Eugenol, Limonene, and Citrus Extract against Bacteria and Yeasts, Representative of the Spoiling Microflora of Fruit Juices. J. Food Prot. 2010, 73, 888–894. [Google Scholar] [CrossRef]
- Avalos, M.; van Wezel, G.P.; Raaijmakers, J.M.; Garbeva, P. Healthy Scents: Microbial Volatiles as New Frontier in Antibiotic Research? Curr. Opin. Microbiol. 2018, 45, 84–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giorgio, A.; de Stradis, A.; Lo Cantore, P.; Iacobellis, N.S. Biocide Effects of Volatile Organic Compounds Produced by Potential BiocontrolRhizobacteria on Sclerotinia sclerotiorum. Front. Microbiol. 2015, 6, 1056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyc, O.; Song, C.; Dickschat, J.S.; Vos, M.; Garbeva, P. The Ecological Role of Volatile and Soluble Secondary Metabolites Produced by Soil Bacteria. Trends Microbiol. 2017, 25, 280–292. [Google Scholar] [CrossRef]
- Cai, R.; Hu, M.; Zhang, Y.; Niu, C.; Yue, T.; Yuan, Y.; Wang, Z. Antifungal Activity and Mechanism of Citral, Limonene and Eugenol against Zygosaccharomyces rouxii. LWT 2019, 106, 50–56. [Google Scholar] [CrossRef]
- Lo Cantore, P.; Shanmugaiah, V.; Iacobellis, N.S. Antibacterial Activity of Essential Oil Components and Their Potential Use in Seed Disinfection. J. Agric. Food Chem. 2009, 57, 9454–9461. [Google Scholar] [CrossRef]
- Silva, A.C.R.D.; Lopes, P.M.; Azevedo, M.M.B.D.; Costa, D.C.M.; Alviano, C.S.; Alviano, D.S. Biological Activities of a-Pinene and β-Pinene Enantiomers. Molecules 2012, 17, 6305. [Google Scholar] [CrossRef] [Green Version]
- Van Vuuren, S.F.; Viljoen, A.M. Antimicrobial Activity of Limonene Enantiomers and 1,8-Cineole Alone and in Combination. Flavour Fragr. J. 2007, 22, 540–544. [Google Scholar] [CrossRef]
- Salehi, B.; Upadhyay, S.; ErdoganOrhan, I.; Kumar Jugran, A.; Jayaweera, S.L.D.; Dias, D.A.; Sharopov, F.; Taheri, Y.; Martins, N.; Baghalpour, N.; et al. Therapeutic Potential of α- and β-Pinene: A Miracle Gift of Nature. Biomolecules 2019, 9, 738. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.-H.; Cai, M.; Liu, Y.-S.; Sun, P.-L.; Luo, S.-L. Antibacterial Activity and Mechanisms of Essential Oil from Citrus Medica L. Var. Sarcodactylis. Molecules 2019, 24, 1577. [Google Scholar] [CrossRef] [Green Version]
- Vespermann, K.A.C.; Paulino, B.N.; Barcelos, M.C.S.; Pessôa, M.G.; Pastore, G.M.; Molina, G. Biotransformation of α- and β-Pinene into Flavor Compounds. Appl. Microbiol. Biotechnol. 2017, 101, 1805–1817. [Google Scholar] [CrossRef]
- Van Dyk, T.K.; Majarian, W.R.; Konstantinov, K.B.; Young, R.M.; Dhurjati, P.S.; LaRossa, R.A. Rapid and Sensitive Pollutant Detection by Induction of Heat Shock Gene-Bioluminescence Gene Fusions. Appl. Environ. Microbiol. 1994, 60, 1414–1420. [Google Scholar] [CrossRef] [Green Version]
- Ben-Israel, O.; Ben-Israel, H.; Ulitzur, S. Identification and Quantification of Toxic Chemicals by Use of Escherichia coli Carryinglux Genes Fused to Stress Promoters. Appl. Environ. Microbiol. 1998, 64, 4346–4352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bechor, O.; Smulski, D.R.; van Dyk, T.K.; LaRossa, R.A.; Belkin, S. Recombinant Microorganisms as Environmental Biosensors: Pollutants Detection by Escherichia coli Bearing FabA′::Lux Fusions. J. Biotechnol. 2002, 94, 125–132. [Google Scholar] [CrossRef]
- Vollmer, C.A.; van Dyk, T.K. Stress Responsive Bacteria: Biosensors as Environmental Monitors. Adv. Microb. Physiol. 2004, 49, 131–174. [Google Scholar] [CrossRef]
- Zavilgelsky, G.B.; Kotova, V.Y.; Manukhov, I.V. Action of 1,1-Dimethylhydrazine on Bacterial Cells Is Determined by Hydrogen Peroxide. Mutat. Res. Toxicol. Environ. Mutagen. 2007, 634, 172–176. [Google Scholar] [CrossRef]
- Kotova, V.Y.; Manukhov, I.V.; Zavilgelskii, G.B. Lux-Biosensors for Detection of SOS-Response, Heat Shock, and Oxidative Stress. Appl. Biochem. Microbiol. 2010, 46, 781–788. [Google Scholar] [CrossRef]
- Woutersen, M.; Belkin, S.; Brouwer, B.; van Wezel, A.P.; Heringa, M.B. Are Luminescent Bacteria Suitable for Online Detection and Monitoring of Toxic Compounds in Drinking Water and Its Sources? Anal. Bioanal. Chem. 2011, 400, 915–929. [Google Scholar] [CrossRef] [Green Version]
- Guyer, M.S.; Reed, R.R.; Steitz, J.A.; Low, K.B. Identification of a Sex-Factor-Affinity Site in E. coli as Γδ. Cold Spring Harb. Symp. Quant. Biol. 1981, 45, 135–140. [Google Scholar] [CrossRef]
- Datsenko, K.A.; Wanner, B.L. One-Step Inactivation of Chromosomal Genes in Escherichia coli K-12 Using PCR Products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baba, T.; Ara, T.; Hasegawa, M.; Takai, Y.; Okumura, Y.; Baba, M.; Datsenko, K.A.; Tomita, M.; Wanner, B.L.; Mori, H. Construction of Escherichia coli K-12 in-Frame, Single-Gene Knockout Mutants: The Keio Collection. Mol. Syst. Biol. 2006, 2, 2006.0008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlioz, A.; Touati, D. Isolation of Superoxide Dismutase Mutants in Escherichia Coli: Is Superoxide Dismutase Necessary for Aerobic Life? EMBO J. 1986, 5, 623–630. [Google Scholar] [CrossRef]
- Riether, K.; Dollard, M.-A.; Billard, P. Assessment of Heavy Metal Bioavailability Using Escherichia coli ZntAp::Lux and CopAp::Lux-Based Biosensors. Appl. Microbiol. Biotechnol. 2001, 57, 712–716. [Google Scholar] [CrossRef] [PubMed]
- Zavilgelsky, G.B.; Kotova, V.Y.; Manukhov, I.V. Sensor bioluminescent systems with lux operons for detection of toxical compounds. Russ. J. Phys. Chem. 2012, 31, 15–20. [Google Scholar]
- Van Dyk, T.K.; Rosson, R.A. Photorhabdus Luminescens LuxCDABE Promoter Probe Vectors. In Bioluminescence Methods and Protocols; Humana Press: Totowa, NJ, USA, 1998; Volume 102, pp. 85–96. [Google Scholar] [CrossRef]
- Zavilgelsky, G.B.; Zarubina, A.P.; Manukhov, I.V. Sequencing and Comparative Analysis of the Lux Operon of Photorhabdusluminescens Strain ZM1: ERIC Elements as Putative Recombination Spots. Mol. Biol. 2002, 36, 792–804. [Google Scholar] [CrossRef]
- Zavilgelsky, G.B.; Kotova, V.Y.; Manukhov, I.V. The Effects of the Regulatory Proteins RcsA and RcsB on the Expression of the Vibrio fischeri Lux Operon in Escherichia coli. Mol. Biol. 2003, 37, 704–711. [Google Scholar] [CrossRef]
- Lundovskich, I.A.; Leontieva, O.V.; Dementieva, E.I.; Ugarova, N.N. Bioluminescence and Chemiluminescence: Perspectives for 21st Century; Roda, A., Pazzagli, M., Kricka, L.J., Stanley, P.E., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 1999; pp. 420–424. [Google Scholar]
- Sambrook, J.; Russel, D.W. Molecular Cloning, 3-Volume Set: A Laboratory Manual; Cold Spring Harboc Laboratory Press: New York, NY, USA, 2001; p. 999. [Google Scholar]
- Lundin, A.; Thore, A. Comparison of Methods for Extraction of Bacterial Adenine Nucleotides Determined by Firefly Assay. Appl. Microbiol. 1975, 30, 713–721. [Google Scholar] [CrossRef]
- Nunoshiba, T.; Nishioka, H. ‘Rec-Lac Test’ for Detecting SOS-Inducing Activity of Environmental Genotoxic Substances. Mutat. Res. Repair 1991, 254, 71–77. [Google Scholar] [CrossRef]
- Goñi, P.; López, P.; Sánchez, C.; Gómez-Lus, R.; Becerril, R.; Nerín, C. Antimicrobial Activity in the Vapour Phase of a Combination of Cinnamon and Clove Essential Oils. Food Chem. 2009, 116, 982–989. [Google Scholar] [CrossRef]
- Melkina, O.E.; Khmel, I.A.; Plyuta, V.A.; Koksharova, O.A.; Zavilgelsky, G.B. Ketones 2-Heptanone, 2-Nonanone, and 2-Undecanone Inhibit DnaK-Dependent Refolding of Heat-Inactivated Bacterial Luciferases in Escherichia coli Cells Lacking Small Chaperon IbpB. Appl. Microbiol. Biotechnol. 2017, 101, 5765–5771. [Google Scholar] [CrossRef] [PubMed]
- Matuszewska, M.; Kuczyńska-Wiśnik, D.; Laskowska, E.; Liberek, K. The Small Heat Shock Protein IbpA of Escherichia coli Cooperates with IbpB in Stabilization of Thermally Aggregated Proteins in a Disaggregation Competent State. J. Biol. Chem. 2005, 280, 12292–12298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mogk, A.; Deuerling, E.; Vorderwülbecke, S.; Vierling, E.; Bukau, B. Small Heat Shock Proteins, ClpB and the DnaK System Form a Functional Triade in Reversing Protein Aggregation. Mol. Microbiol. 2003, 50, 585–595. [Google Scholar] [CrossRef]
- Mogk, A.; Schlieker, C.; Friedrich, K.L.; Schönfeld, H.-J.; Vierling, E.; Bukau, B. Refolding of Substrates Bound to Small Hsps Relies on a Disaggregation Reaction Mediated Most Efficiently by ClpB/DnaK. J. Biol. Chem. 2003, 278, 31033–31042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palomares, A.J.; DeLuca, M.A.; Helinski, D.R. Firefly Luciferase as a Reporter Enzyme for Measuring Gene Expression in Vegetative and Symbiotic Rhizobium meliloti and Other Gram-Negative Bacteria. Gene 1989, 81, 55–64. [Google Scholar] [CrossRef]
- Subramani, S.; DeLuca, M. Applications of the Firefly Luciferase as a Reporter Gene. In Genetic Engineering; Springer US: Boston, MA, USA, 1988; pp. 75–89. [Google Scholar] [CrossRef]
- Wood, K.V.; DeLuca, M. Photographic Detection of Luminescence in Escherichia coli Containing the Gene for Firefly Luciferase. Anal. Biochem. 1987, 161, 501–507. [Google Scholar] [CrossRef]
- Dwyer, D.J.; Kohanski, M.A.; Hayete, B.; Collins, J.J. Gyrase Inhibitors Induce an Oxidative Damage Cellular Death Pathway in Escherichia coli. Mol. Syst. Biol. 2007, 3, 91. [Google Scholar] [CrossRef]
- Dwyer, D.J.; Belenky, P.A.; Yang, J.H.; MacDonald, I.C.; Martell, J.D.; Takahashi, N.; Chan, C.T.Y.; Lobritz, M.A.; Braff, D.; Schwarz, E.G.; et al. Antibiotics Induce Redox-Related Physiological Alterations as Part of Their Lethality. Proc. Natl. Acad. Sci. USA 2014, 111, E2100–E2109. [Google Scholar] [CrossRef] [Green Version]
- Kohanski, M.A.; Dwyer, D.J.; Collins, J.J. How Antibiotics Kill Bacteria: From Targets to Networks. Nat. Rev. Microbiol. 2010, 8, 423–435. [Google Scholar] [CrossRef] [Green Version]
- Julian, W.T.; Vasilchenko, A.V.; Shpindyuk, D.D.; Poshvina, D.V.; Vasilchenko, A.S. Bacterial-Derived Plant Protection Metabolite 2,4-Diacetylphloroglucinol: Effects on Bacterial Cells at Inhibitory and Subinhibitory Concentrations. Biomolecules 2020, 11, 13. [Google Scholar] [CrossRef] [PubMed]
- Makemson, J.C.; Hastings, J.W. Poising of the Arginine Pool and Control of Bioluminescence in BeneckeaHarveyi. J. Bacteriol. 1979, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holzman, T.F.; Baldwin, T.O. Binding of 2,2-Diphenylpropylamine at the Aldehyde Site of Bacterial Luciferase Increases the Affinity of the Reduced Riboflavin 5′-Phosphate Site. Biochemistry 1981, 20, 5524–5528. [Google Scholar] [CrossRef]
- Holzman, T.; Baldwin, T. Corrections—Reversible Inhibition of the Bacterial Luciferase Catalyzed Bioluminescence Reaction by Aldehyde Substrate: Kinetic Mechanism and Ligand Effects. Biochemistry 1983, 22, 5464. [Google Scholar] [CrossRef]
- Virta, M.; Åkerman, K.E.O.; Saviranta, P.; Oker-Blom, C.; Karp, M.T. Real-Time Measurement of Cell Permeabilization with Low-Molecular-Weight Membranolytic Agents. J. Antimicrob. Chemother. 1995, 36, 303–315. [Google Scholar] [CrossRef]
- Gamer, J.; Bujard, H.; Bukau, B. Physical Interaction between Heat Shock Proteins DnaK, DnaJ, and GrpE and the Bacterial Heat Shock Transcription Factor Σ32. Cell 1992, 69, 833–842. [Google Scholar] [CrossRef]
- Liberek, K.; Galitski, T.P.; Zylicz, M.; Georgopoulos, C. The DnaK Chaperone Modulates the Heat Shock Response of Escherichia coli by Binding to the Sigma 32 Transcription Factor. Proc. Natl. Acad. Sci. USA 1992, 89, 3516–3520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomoyasu, T.; Gamer, J.; Bukau, B.; Kanemori, M.; Mori, H.; Rutman, A.J.; Oppenheim, A.B.; Yura, T.; Yamanaka, K.; Niki, H. Escherichia coli FtsH Is a Membrane-Bound, ATP-Dependent Protease Which Degrades the Heat-Shock Transcription Factor Sigma 32. EMBO J. 1995, 14, 2551–2560. [Google Scholar] [CrossRef]
- Arsène, F.; Tomoyasu, T.; Bukau, B. The Heat Shock Response of Escherichia coli. Int. J. Food Microbiol. 2000, 55, 3–9. [Google Scholar] [CrossRef]
- Cassel, J.A.; Ilyin, S.; McDonnell, M.E.; Reitz, A.B. Novel Inhibitors of Heat Shock Protein Hsp70-Mediated Luciferase Refolding That Bind to DnaJ. Bioorg. Med. Chem. 2012, 20, 3609–3614. [Google Scholar] [CrossRef]
- Moses, M.A.; Zuehlke, A.D.; Neckers, L. Molecular Chaperone Inhibitors. In Heat Shock Proteins in the Immune System; Springer International Publishing: Cham, Switzerland, 2018; pp. 21–40. [Google Scholar] [CrossRef]
- Lazarev, V.F.; Sverchinsky, D.V.; Mikhaylova, E.R.; Semenyuk, P.I.; Komarova, E.Y.; Niskanen, S.A.; Nikotina, A.D.; Burakov, A.V.; Kartsev, V.G.; Guzhova, I.V.; et al. Sensitizing Tumor Cells to Conventional Drugs: HSP70 Chaperone Inhibitors, Their Selection and Application in Cancer Models. Cell Death Dis. 2018, 9, 41. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The chemical structure of (−)-limonene (A) and (+)-α-pinene (B).
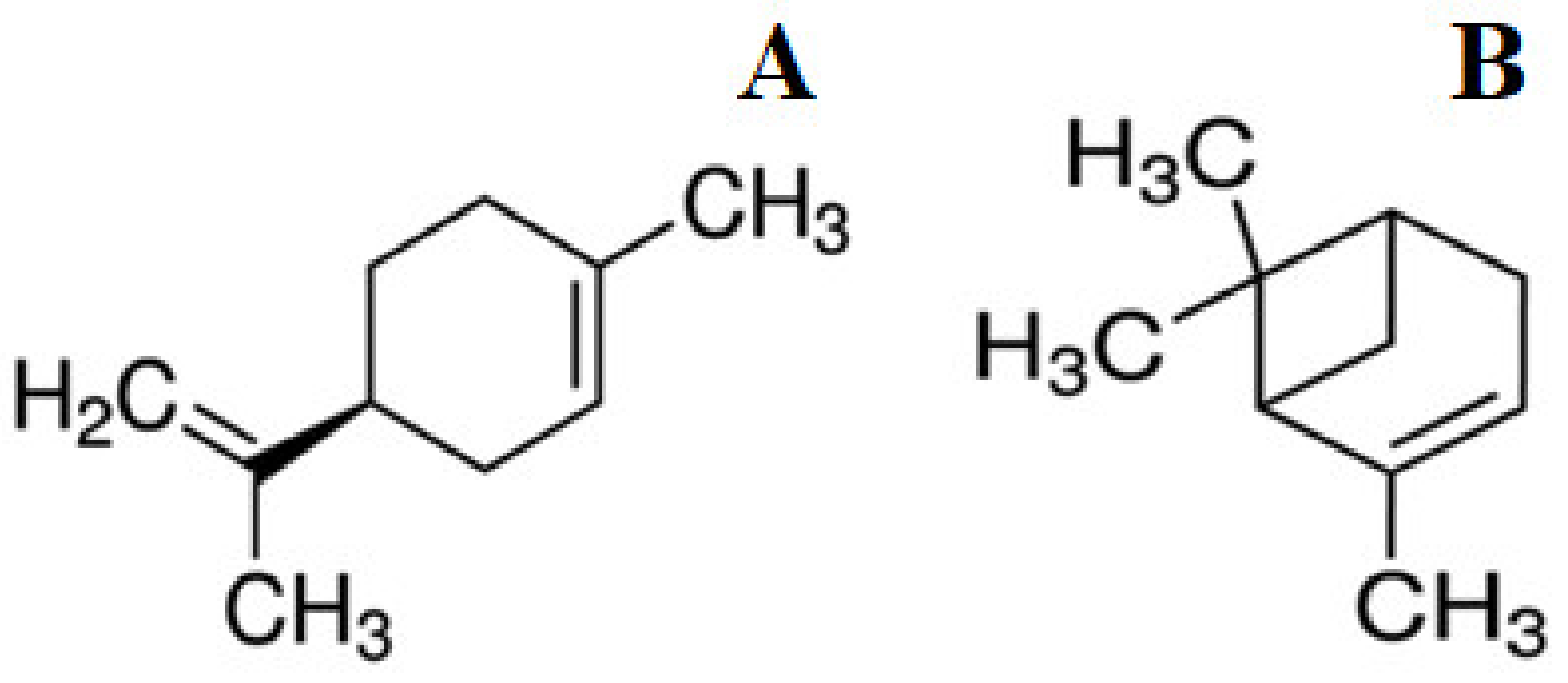
Figure 2.
Luminescent response of strain E. coli MG1655 (pKatG’::lux) to hydrogen peroxide (A), (−)-limonene (B), and (+)-α-pinene (C). Time course of mean values of light emission (in RLU) in the presence of different chemical concentrations (A,B): (◆) 0 μM (black diamond—Control), (◼) 1 μM (gray square), (▲) 5 μM (dark triangle), (×) 25 μM (black cross), (🞶) 50 μM (gray asterisk), and (⬤) 100 μM (light circle); (C): (◆) 0 μM (black diamond—control), (◼) 5 μM (light square), (▲) 50 μM (gray triangle), (⬤) 100 μM (black circle).
Figure 2.
Luminescent response of strain E. coli MG1655 (pKatG’::lux) to hydrogen peroxide (A), (−)-limonene (B), and (+)-α-pinene (C). Time course of mean values of light emission (in RLU) in the presence of different chemical concentrations (A,B): (◆) 0 μM (black diamond—Control), (◼) 1 μM (gray square), (▲) 5 μM (dark triangle), (×) 25 μM (black cross), (🞶) 50 μM (gray asterisk), and (⬤) 100 μM (light circle); (C): (◆) 0 μM (black diamond—control), (◼) 5 μM (light square), (▲) 50 μM (gray triangle), (⬤) 100 μM (black circle).

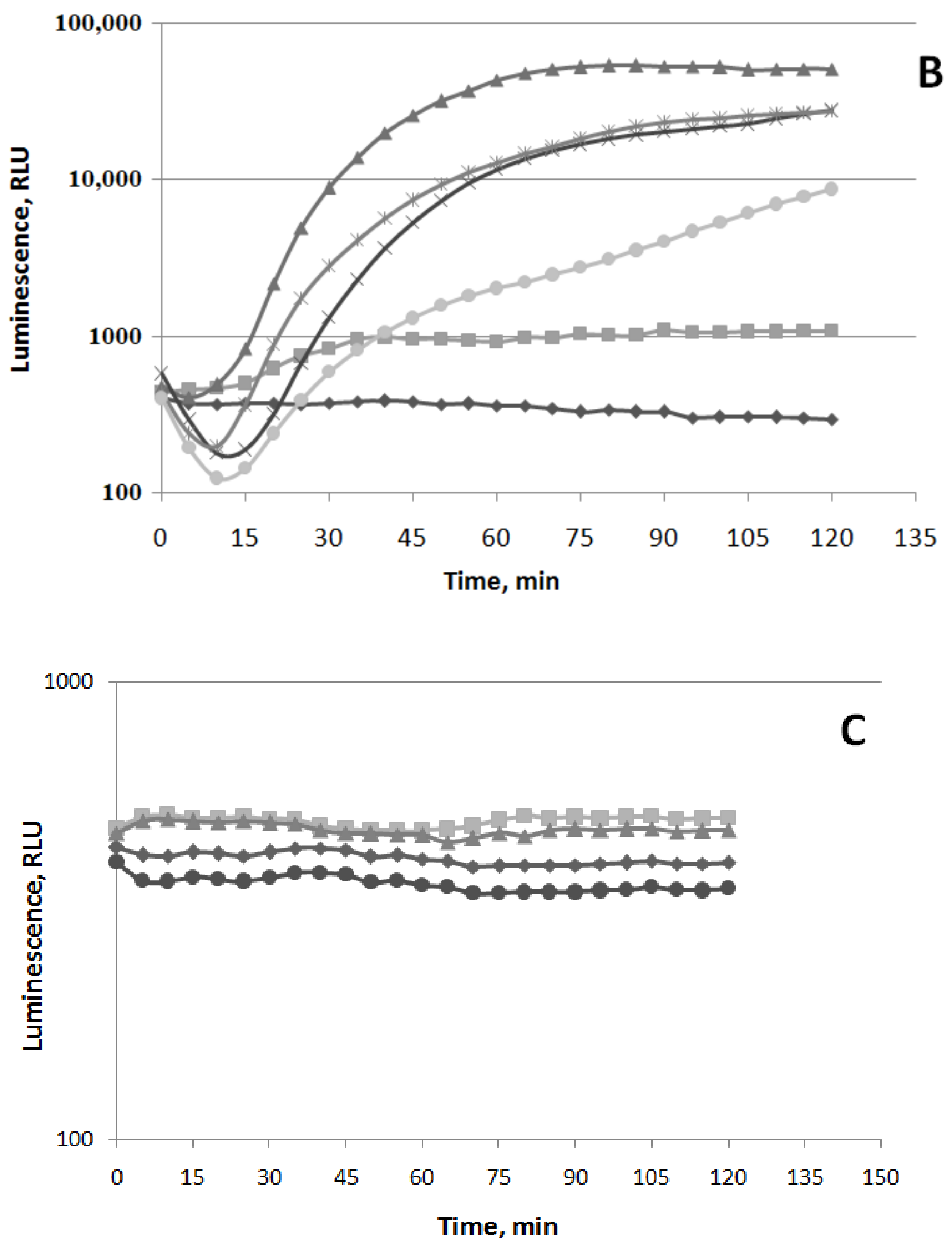
Figure 3.
Effect of ΔkatG mutation on the induction of the PkatG promoter by (−)-limonene. Luminescent response of wild-type strain (E. coli MG1655 (pKatG’::lux)—dark symbols) and ΔkatG mutant strain (E. coli JW3914-1 ΔkatG729::kan (pKatG’::lux)—light symbols) to (−)-limonene. Kinetics of changes in mean values of light emission (in RLU) at various concentrations of (−)-limonene: (◆, ◇) 0 μM (control), (◼, ▢) 1 μM, (▲, △) 10 μM.
Figure 3.
Effect of ΔkatG mutation on the induction of the PkatG promoter by (−)-limonene. Luminescent response of wild-type strain (E. coli MG1655 (pKatG’::lux)—dark symbols) and ΔkatG mutant strain (E. coli JW3914-1 ΔkatG729::kan (pKatG’::lux)—light symbols) to (−)-limonene. Kinetics of changes in mean values of light emission (in RLU) at various concentrations of (−)-limonene: (◆, ◇) 0 μM (control), (◼, ▢) 1 μM, (▲, △) 10 μM.
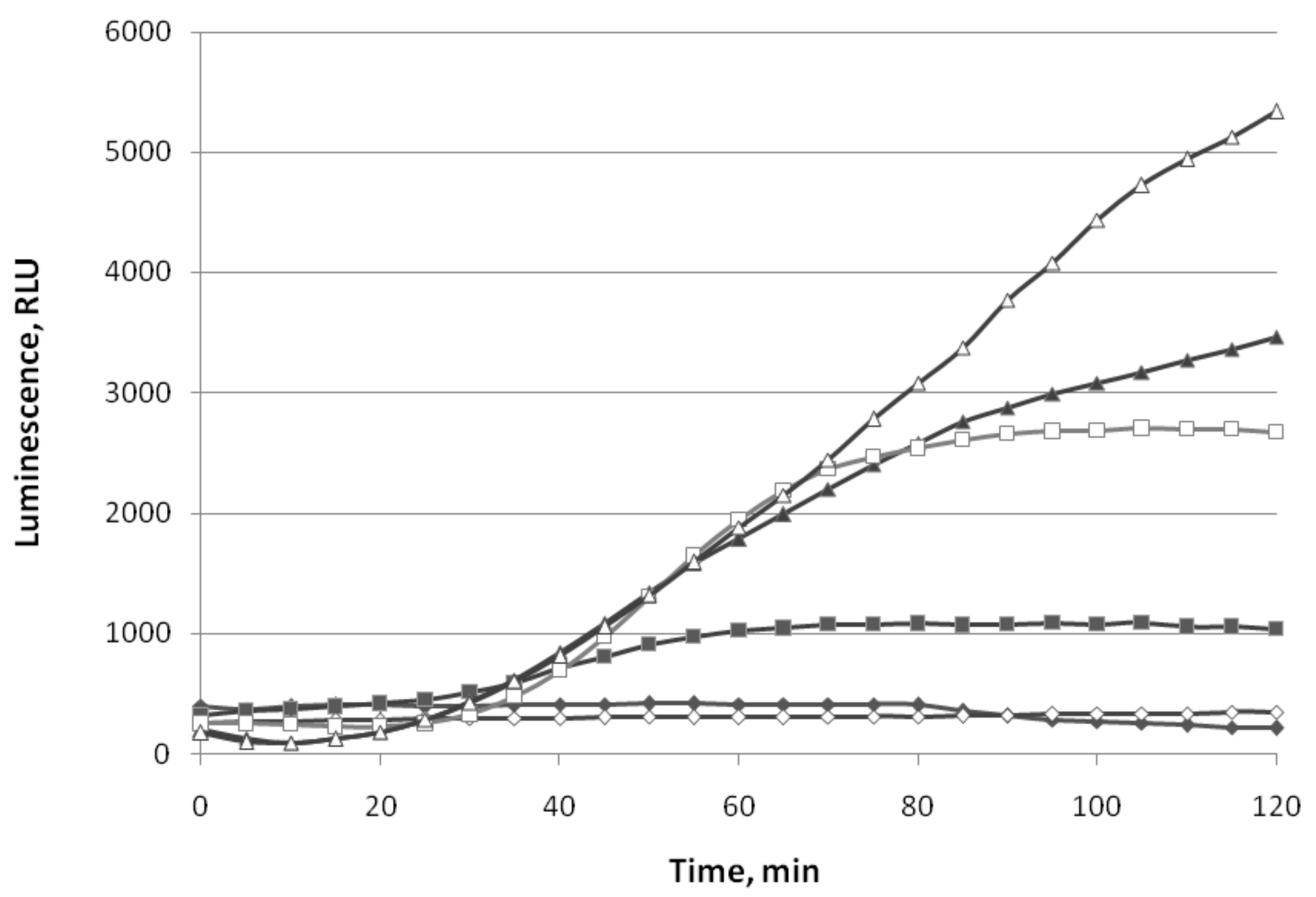
Figure 4.
Luminescence response of strain E. coli MG1655 (pSoxS’::lux) to paraquat (methyl viologen) (A), (−)-limonene (B), and (+)-α-pinene (C). Kinetics of changes in mean values of light emission (in RLU) at various chemical concentrations: (A): (◆) 0 μM (control), (◼) 5 μM, (▲)—50 μM, (⬤) 100 μM; (B): (◆) 0 μM (control), (◼) 1 μM, (▲) 5 μM, (×) 25 μM, (🞶) 50 μM, (⬤) 100 μM; (C): (◆) 0 μM (control), (▲) 5 μM, (🞶) 50 μM, (⬤) 100 μM.
Figure 4.
Luminescence response of strain E. coli MG1655 (pSoxS’::lux) to paraquat (methyl viologen) (A), (−)-limonene (B), and (+)-α-pinene (C). Kinetics of changes in mean values of light emission (in RLU) at various chemical concentrations: (A): (◆) 0 μM (control), (◼) 5 μM, (▲)—50 μM, (⬤) 100 μM; (B): (◆) 0 μM (control), (◼) 1 μM, (▲) 5 μM, (×) 25 μM, (🞶) 50 μM, (⬤) 100 μM; (C): (◆) 0 μM (control), (▲) 5 μM, (🞶) 50 μM, (⬤) 100 μM.

Figure 5.
Luminescence response of strain E. coli MG1655 (pColD’::lux) to mitomycin C, (−)-limonene, and (+)-α-pinene. Kinetics of changes in mean values of light emission (in RLU) at various chemical concentrations: (◆) 0 μM (control); (◼) mitomycin C 10 μM; (⬤) (+)-α-pinene 100 μM; (−)-limonene (▲) 5 μM, (×) 25 μM, and (🞶) 100 μM.
Figure 5.
Luminescence response of strain E. coli MG1655 (pColD’::lux) to mitomycin C, (−)-limonene, and (+)-α-pinene. Kinetics of changes in mean values of light emission (in RLU) at various chemical concentrations: (◆) 0 μM (control); (◼) mitomycin C 10 μM; (⬤) (+)-α-pinene 100 μM; (−)-limonene (▲) 5 μM, (×) 25 μM, and (🞶) 100 μM.
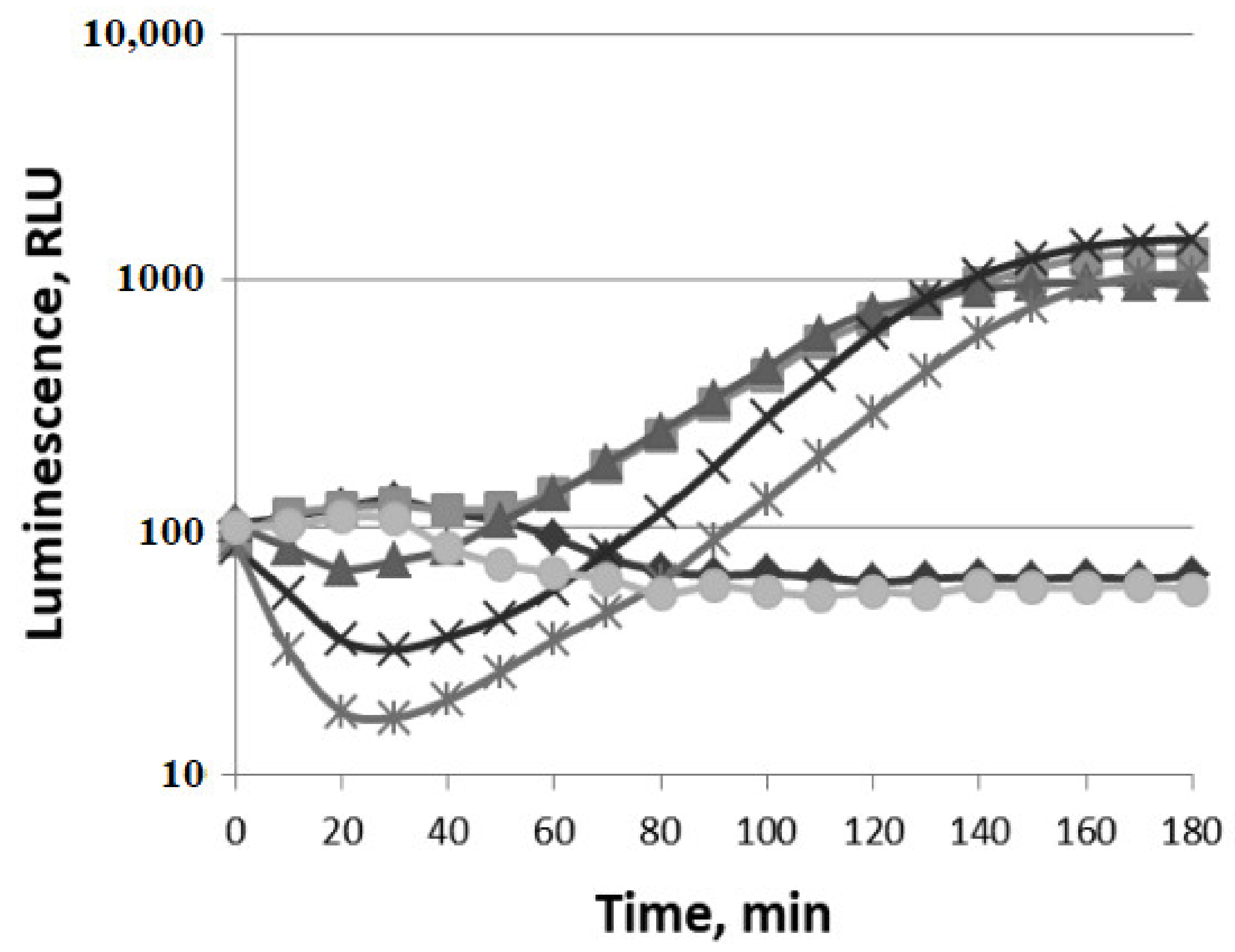
Figure 6.
Luminescent responses of strains E. coli MG1655 (pFabA’::lux) to Triton X-100 and (−)-limonene (A) and E. coli MG1655 (pMerR’::lux) to HgCl2 and (−)-limonene (B). Kinetics of changes in mean values of light emission (in RLU) at various chemical concentrations: (A): (◆) Control (0 μM); TritonX-100 (◼) 0.5 μg/mL, (▲) 2 μg/mL, and (×) 4 μg/mL; (−)-limonene (🞶) 2 μM, (+) 4 μM, and (⬤) 10 μM. (B): (◆) Control (0 μM); HgCl2(▲) 0.001 μM and (◼) 0.3 μM; (−)-limonene (×) 2 μM, (🞶) 4 μM, and (⬤) 10 μM.
Figure 6.
Luminescent responses of strains E. coli MG1655 (pFabA’::lux) to Triton X-100 and (−)-limonene (A) and E. coli MG1655 (pMerR’::lux) to HgCl2 and (−)-limonene (B). Kinetics of changes in mean values of light emission (in RLU) at various chemical concentrations: (A): (◆) Control (0 μM); TritonX-100 (◼) 0.5 μg/mL, (▲) 2 μg/mL, and (×) 4 μg/mL; (−)-limonene (🞶) 2 μM, (+) 4 μM, and (⬤) 10 μM. (B): (◆) Control (0 μM); HgCl2(▲) 0.001 μM and (◼) 0.3 μM; (−)-limonene (×) 2 μM, (🞶) 4 μM, and (⬤) 10 μM.
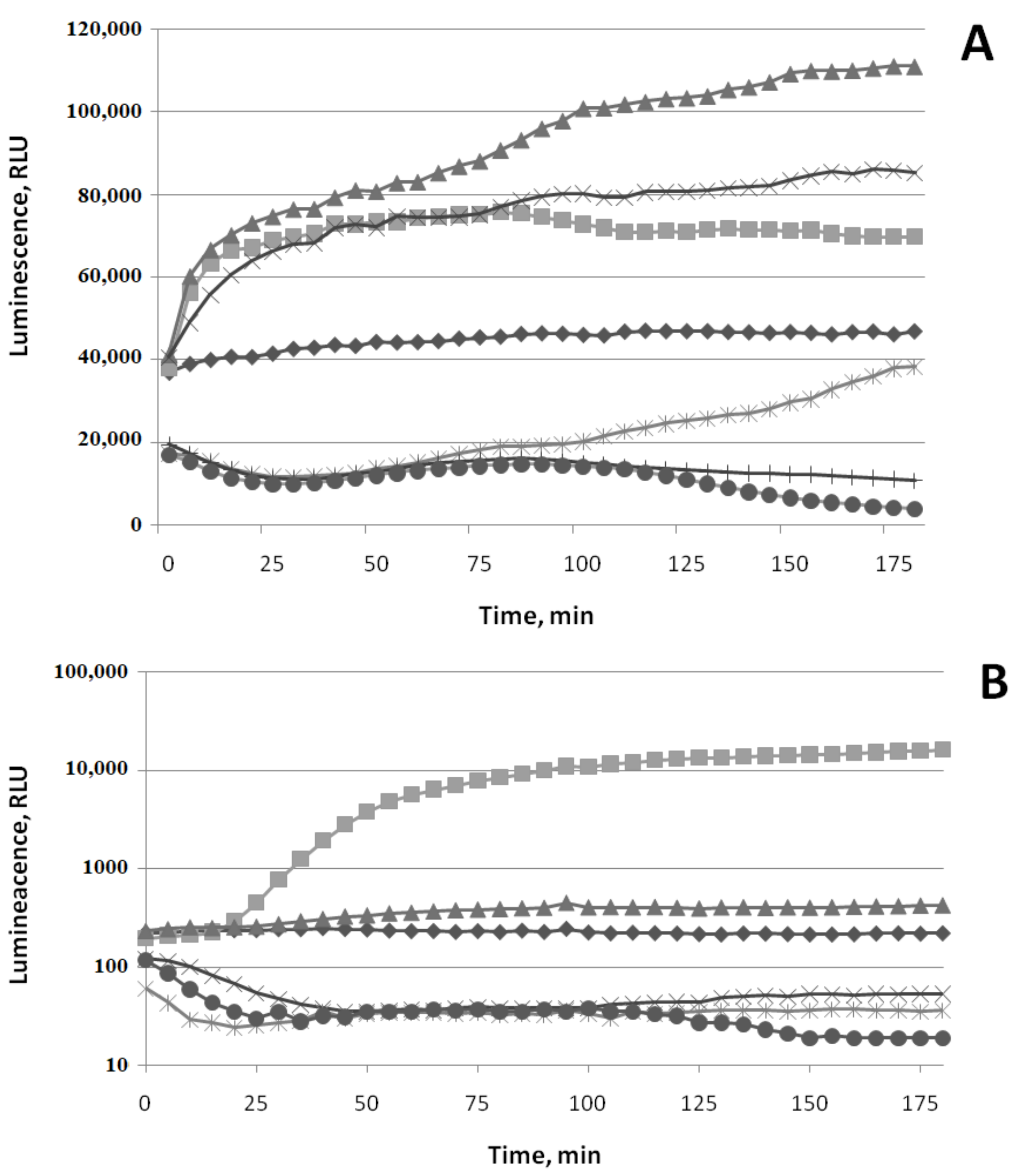
Figure 7.
Luminescence response of E. coli MG1655 (pIbpA’::lux) (“heatshock”) to(−)-limonene (A) and (+)-α-pinene (B). Kinetics of changes in mean values of light emission (in RLU) at various chemical concentrations: (A,B): (◆) 0 μM (control); (◼) ethanol (4%); (▲) 5 μM, (×) 10 μM, (🞶) 15 μM, and (⬤) 20 μM of (−)-limonene (A) and (+)-α-pinene (B).
Figure 7.
Luminescence response of E. coli MG1655 (pIbpA’::lux) (“heatshock”) to(−)-limonene (A) and (+)-α-pinene (B). Kinetics of changes in mean values of light emission (in RLU) at various chemical concentrations: (A,B): (◆) 0 μM (control); (◼) ethanol (4%); (▲) 5 μM, (×) 10 μM, (🞶) 15 μM, and (⬤) 20 μM of (−)-limonene (A) and (+)-α-pinene (B).
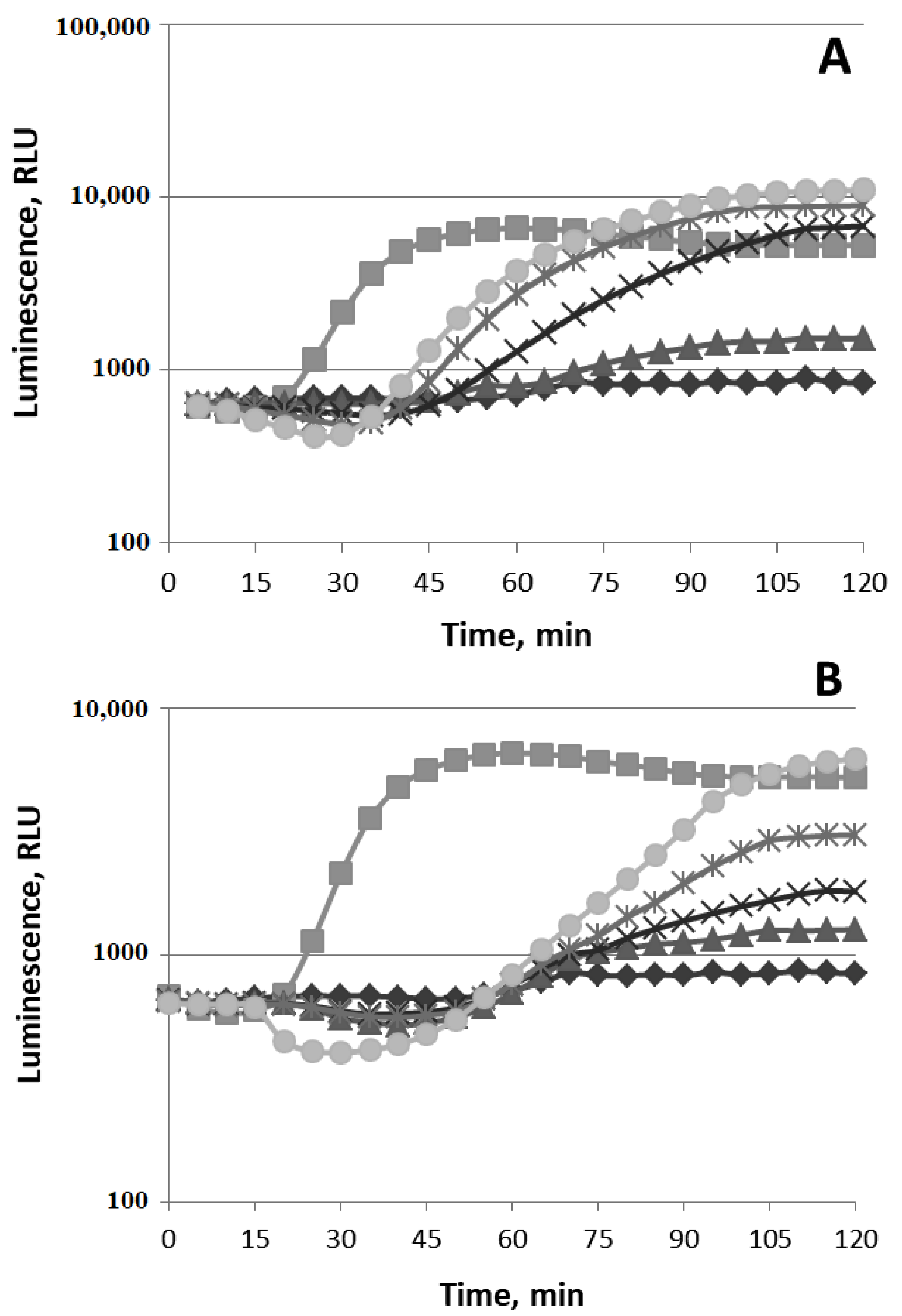
Figure 8.
Effect of (−)-limonene and (+)-α-pinene on luciferaseenzymatic activity of P. luminescens in the E. coli MG1655 (pXen7) strain. Kinetics of changes in mean values of light emission (in RLU) at various chemical concentrations: (◆) 0 μM (control); (+)-α-pinene (◼) 1 μM, (▲) 10 μM, and (+) 100 μM; (−)-limonene (🞶) 1 μM, (⬤) 10 μM, and (×) 100 μM. The ordinate axis shows the enzymatic activity of luciferase. The abscissa axis shows the time of incubation at 22 °C.
Figure 8.
Effect of (−)-limonene and (+)-α-pinene on luciferaseenzymatic activity of P. luminescens in the E. coli MG1655 (pXen7) strain. Kinetics of changes in mean values of light emission (in RLU) at various chemical concentrations: (◆) 0 μM (control); (+)-α-pinene (◼) 1 μM, (▲) 10 μM, and (+) 100 μM; (−)-limonene (🞶) 1 μM, (⬤) 10 μM, and (×) 100 μM. The ordinate axis shows the enzymatic activity of luciferase. The abscissa axis shows the time of incubation at 22 °C.
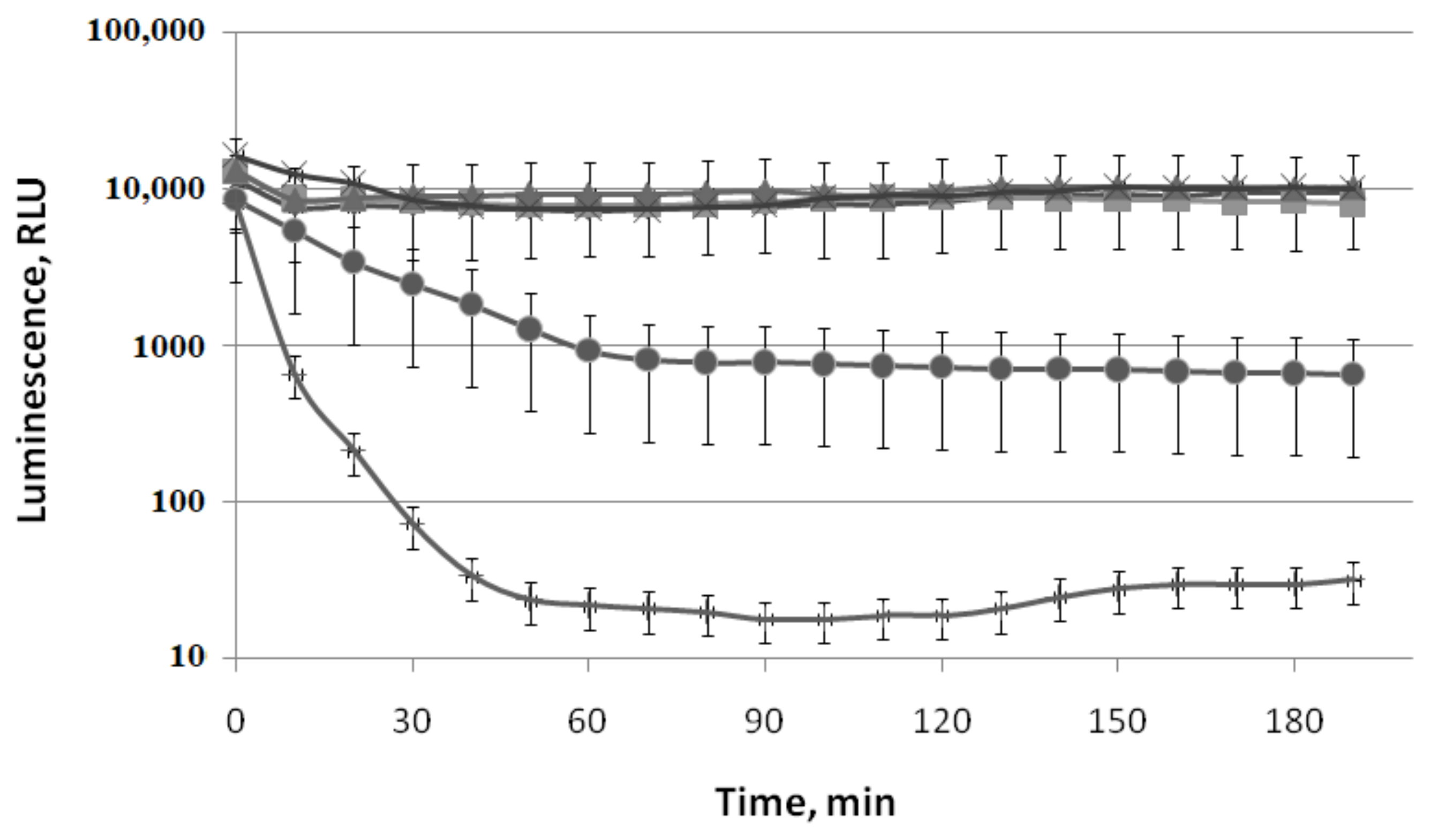
Figure 9.
Effect of (−)-limonene, (+)-α-pinene, and CCCP on the kinetics of refolding of heat-inactivated P. leiognathi luciferase in the wild-type strain (E. coli BW25113 (pLeo1)) (A) and ΔibpB mutant strain(E. coli JW3663 ibpB::kan (pLeo1)) (B). (◆) Control (0 μM), (×) 20 μM of (+)-α-pinene, (◼) 10 μM of (−)-limonene, (▲) 50 μM of CCCP. The ordinate axis shows the enzymatic activity of luciferase in the percentage of the initial level. The abscissa axis shows the time of incubation at 22 °C.
Figure 9.
Effect of (−)-limonene, (+)-α-pinene, and CCCP on the kinetics of refolding of heat-inactivated P. leiognathi luciferase in the wild-type strain (E. coli BW25113 (pLeo1)) (A) and ΔibpB mutant strain(E. coli JW3663 ibpB::kan (pLeo1)) (B). (◆) Control (0 μM), (×) 20 μM of (+)-α-pinene, (◼) 10 μM of (−)-limonene, (▲) 50 μM of CCCP. The ordinate axis shows the enzymatic activity of luciferase in the percentage of the initial level. The abscissa axis shows the time of incubation at 22 °C.


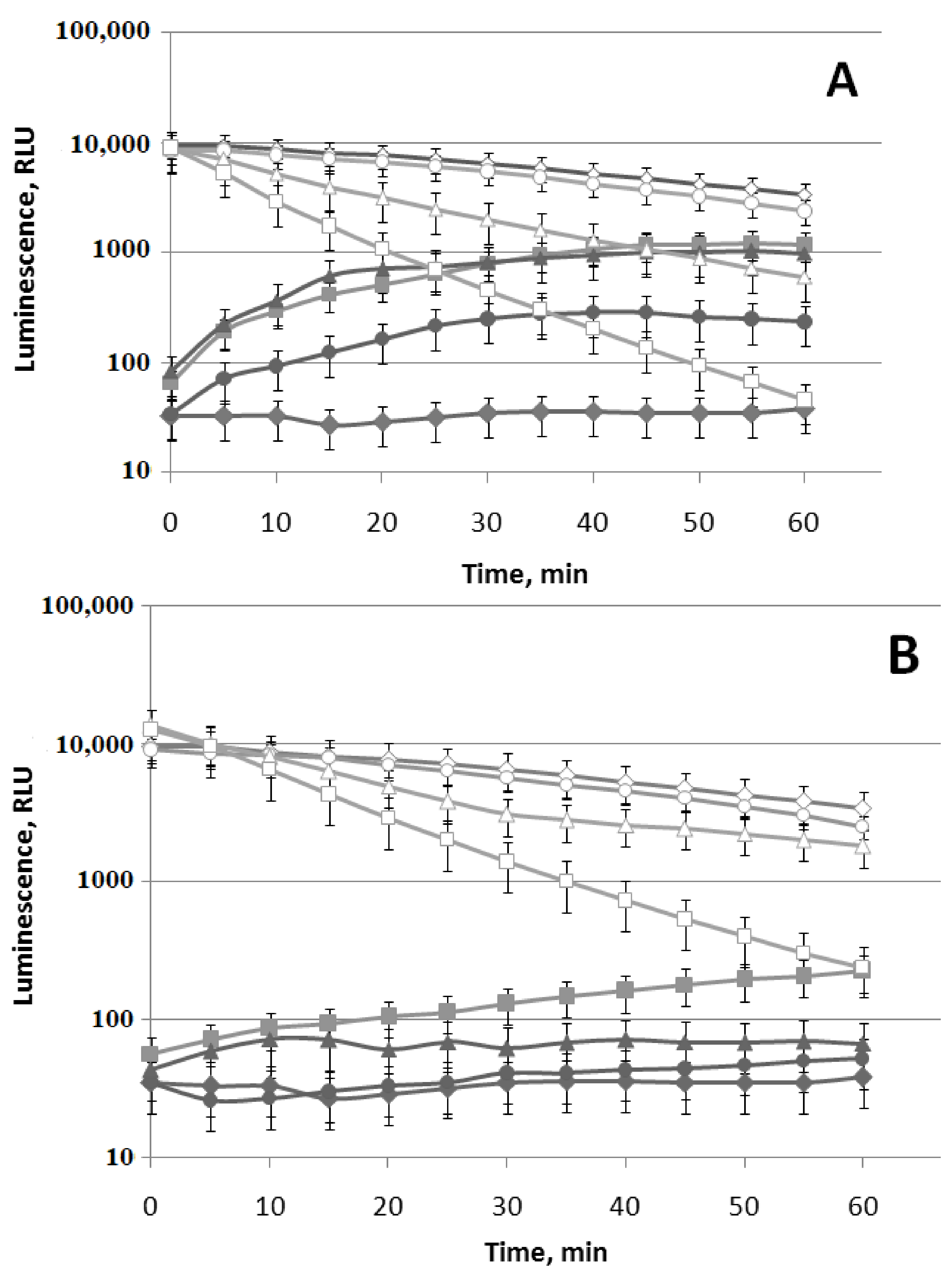
Figure 10.
Effect of (−)-limonene (A) and (+)-α-pinene (B) on the activity of firefly Luciola mingrelica luciferase in E. coli MG1655 (pLR)living and lysed cells. Kinetics of changes in mean values of light emission (in RLU) at various chemical concentrations. (A,B): E. coli MG1655 (pLR) (dark symbol) and bacterial cell lysate (light symbol); (◆, ◇) 0 μM (control), (⬤,🞅) 1 μM, (▲,△) 10 μM, (◼,□) 100 μM. The ordinate axis shows the enzymatic activity of luciferase. The abscissa axis shows the time of incubation at 20 °C.
Figure 10.
Effect of (−)-limonene (A) and (+)-α-pinene (B) on the activity of firefly Luciola mingrelica luciferase in E. coli MG1655 (pLR)living and lysed cells. Kinetics of changes in mean values of light emission (in RLU) at various chemical concentrations. (A,B): E. coli MG1655 (pLR) (dark symbol) and bacterial cell lysate (light symbol); (◆, ◇) 0 μM (control), (⬤,🞅) 1 μM, (▲,△) 10 μM, (◼,□) 100 μM. The ordinate axis shows the enzymatic activity of luciferase. The abscissa axis shows the time of incubation at 20 °C.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Escherichia coli strains used in this study.
| Strain | CGSC ** ID | Genotype | Reference | Comments |
|---|---|---|---|---|
| MG1655 | 6300 | F-ilvG rfb-50 rph-1 | [22] | Prototroph |
| BW25113 | 7636 | F−, Δ(araD-araB)567, ΔlacZ4787(::rrnB-3), л−, rph-1, Δ(rhaD-rhaB)568, hsdR514 | [23] | (This is) The parent strain for the Keio Collection of single-gene knockouts. |
| JW3914-1 | 10827 | ΔkatG729::kan | [24] | Keio Collection strain |
| JW3663 | 10689 | ibpB::kan | [24] | Keio Collection strain |
| JW3933-3 | 12039 | ΔoxyR749::kan | [24] | Keio Collection strain |
| QC868 | no | F−leu6 thrA1 proA2 thi-1 lacY1 tonA1 rpsL31 supE44 hsdR−sodA+sodB+Smr | [25] | The parent strain for QC871 |
| QC871 | no | F−leu6 thrA1 proA2 thi-1 lacY1 tonA1 rpsL31 supE44 hsdR−sodA25 sodB2CmrKanrSmr | [25] | The double mutant (sodAB−) without the manganese- and iron-cofactored superoxide dismutases |
** CGSC—The Coli Genetic StockCenter.
Table 2.
Diameters of inhibition zone (DIZs) of different E. coli strains afterincubation with (−)-limonene (4 µL (3.36 mg)).
Table 2.
Diameters of inhibition zone (DIZs) of different E. coli strains afterincubation with (−)-limonene (4 µL (3.36 mg)).
| E. coli Strain | DIZ, mm * |
|---|---|
| MG1655 (wt) | 7 ± 0.5 ** |
| QC871 sodAB- | 8 ± 0.5 |
| JW3933-3 ΔoxyR749::kan | 16 ± 0.5 |
| JW3914-1 ΔkatG729::kan | 10 ± 0.5 |
* All the assays were carried out in triplicate. The data were recorded as mean ± standard error The same results were obtained for BW25113 and QC868.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Melkina, O.E.; Plyuta, V.A.; Khmel, I.A.; Zavilgelsky, G.B. The Mode of Action of Cyclic Monoterpenes (−)-Limonene and (+)-α-Pinene on Bacterial Cells. Biomolecules 2021, 11, 806. https://doi.org/10.3390/biom11060806
AMA Style
Melkina OE, Plyuta VA, Khmel IA, Zavilgelsky GB. The Mode of Action of Cyclic Monoterpenes (−)-Limonene and (+)-α-Pinene on Bacterial Cells. Biomolecules. 2021; 11(6):806. https://doi.org/10.3390/biom11060806
Chicago/Turabian StyleMelkina, Olga E., Vladimir A. Plyuta, Inessa A. Khmel, and Gennadii B. Zavilgelsky. 2021. "The Mode of Action of Cyclic Monoterpenes (−)-Limonene and (+)-α-Pinene on Bacterial Cells" Biomolecules 11, no. 6: 806. https://doi.org/10.3390/biom11060806
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.