
Moderating Effects of BDNF Genetic Variants and Smoking on Cognition in PTSD Veterans

 , , , , ,
, , , , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
Participants
3. Results
3.1. The BDNF rs6265 and BDNF rs56164415 Frequency in Veterans with PTSD and Control Subjects
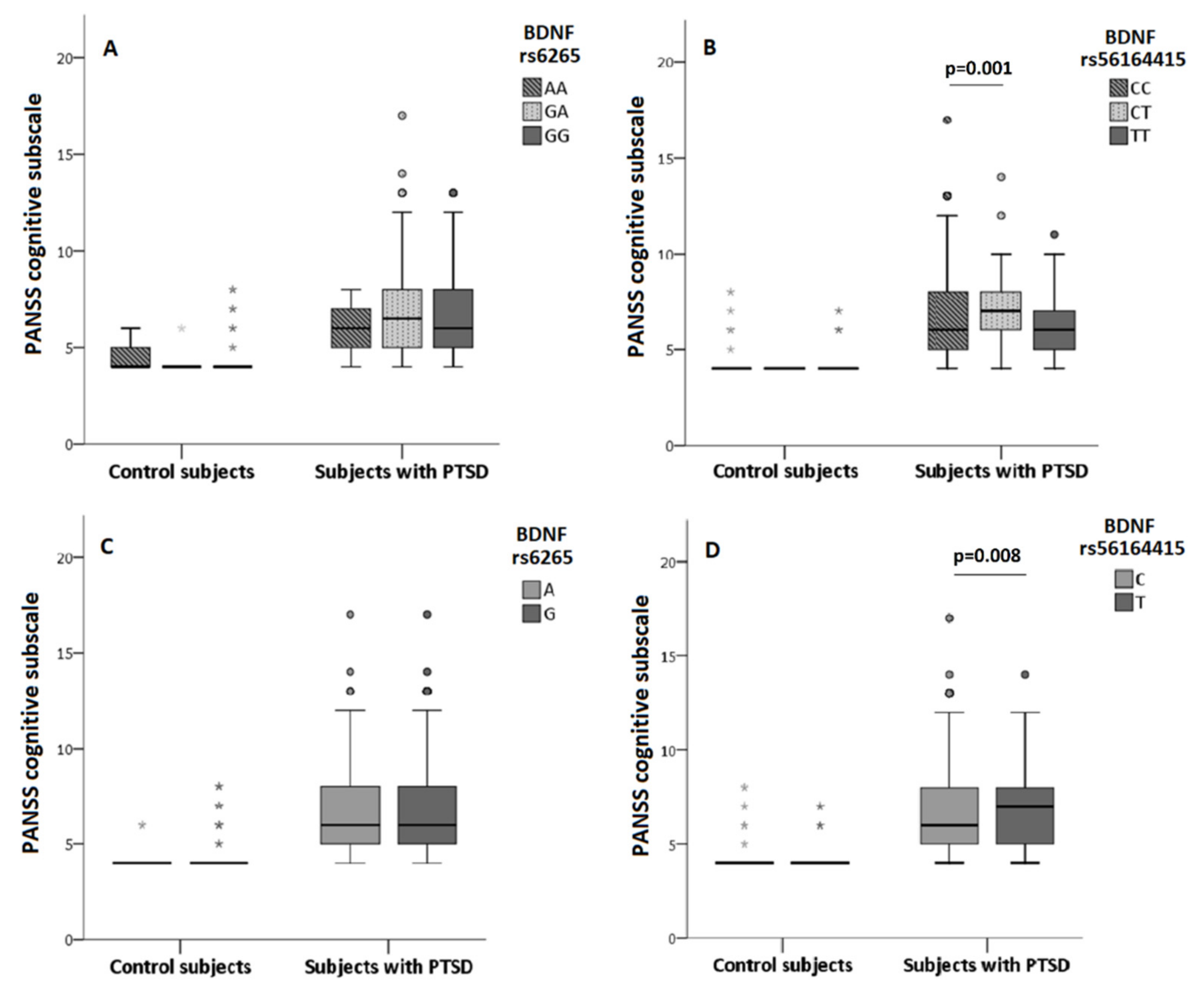
3.2. The Association of BDNF rs6265 and BDNF rs56164415 Polymorphism with Cognition
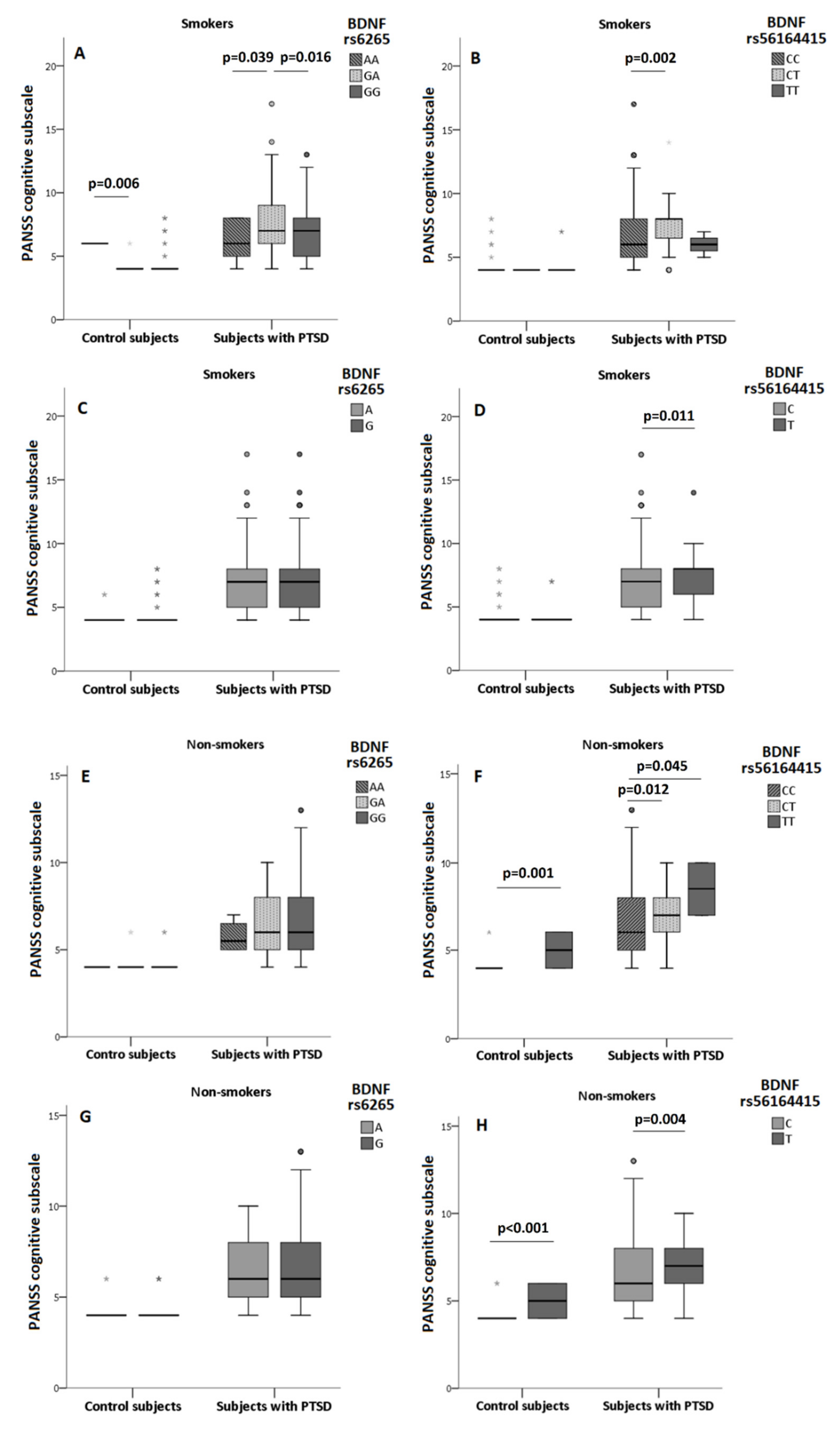
3.3. The Association of BDNF rs6265 and BDNF rs56164415 Polymorphism and Smoking on Cognition
4. Discussion
Limitations and Strengths
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shalev, A.; Liberzon, I.; Marmar, C. Post-traumatic stress disorder. N. Engl. J. Med. 2017, 376, 2459–2469. [Google Scholar] [CrossRef] [PubMed]
- Tortella-Feliu, M.; Fullana, M.A.; Pérez-Vigil, A.; Torres, X.; Chamorro, J.; Littarelli, S.A.; Solanes, A.; Ramella-Cravaro, V.; Vilar, A.; González-Parra, J.A.; et al. Risk factors for posttraumatic stress disorder: An umbrella review of systematic reviews and meta-analyses. Neurosci. Biobehav. Rev. 2019, 107, 154–165. [Google Scholar] [CrossRef] [PubMed]
- De Berardis, D.; Vellante, F.; Fornaro, M.; Anastasia, A.; Olivieri, L.; Rapini, G.; Serroni, N.; Orsolini, L.; Valchera, A.; Carano, A.; et al. Alexithymia, suicide ideation, affective temperaments and homocysteine levels in drug naïve patients with post-traumatic stress disorder: An exploratory study in the everyday ‘real world’ clinical practice. Int. J. Psychiatry Clin. Pract. 2020, 24, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Sherin, J.E.; Nemeroff, C.B. Post-traumatic stress disorder: The neurobiological impact of psychological trauma. Dialogues Clin. Neurosci. 2011, 13, 263–278. [Google Scholar] [CrossRef]
- De Berardis, D.; Marini, S.; Serroni, N.; Iasevoli, F.; Tomasetti, C.; de Bartolomeis, A.; Mazza, M.; Tempesta, D.; Valchera, A.; Fornaro, M.; et al. Targeting the Noradrenergic System in Posttraumatic Stress Disorder: A Systematic Review and Meta-Analysis of Prazosin Trials. Curr. Drug Targets 2015, 16, 1094–1106. [Google Scholar] [CrossRef]
- Aupperle, R.L.; Melrose, A.J.; Stein, M.B.; Paulus, M.P. Executive function and PTSD: Disengaging from trauma. Neuropharmacology 2012, 62, 686–694. [Google Scholar] [CrossRef] [Green Version]
- Scott, J.C.; Matt, G.E.; Wrocklage, K.M.; Crnich, C.; Jordan, J.; Southwick, S.M.; Krystal, J.H.; Schweinsburg, B.C. A quantitative meta-analysis of neurocognitive functioning in posttraumatic stress disorder. Psychol. Bull. 2015, 141, 105–140. [Google Scholar] [CrossRef]
- Karl, A.; Schaefer, M.; Malta, L.S.; Dörfel, D.; Rohleder, N.; Werner, A. A meta-analysis of structural brain abnormalities in PTSD. Neurosci. Biobehav. Rev. 2006, 30, 1004–1031. [Google Scholar] [CrossRef]
- Parslow, R.A.; Jorm, A.F. Pretrauma and posttrauma neurocognitive functioning and PTSD symptoms in a community sample of young adults. Am. J. Psychiatry 2007, 164, 509–515. [Google Scholar] [CrossRef]
- Couette, M.; Mouchabac, S.; Bourla, A.; Nuss, P.; Ferreri, F. Social cognition in post-traumatic stress disorder: A systematic review. Br. J. Clin. Psychol. 2019, 59, 117–138. [Google Scholar] [CrossRef]
- Havelka Mestrovic, A.; Tudor, L.; Nedic Erjavec, G.; Nikolac Perkovic, M.; Svob Strac, D.; Kovacic Petrovic, Z.; Pivac, N. The impact of BDNF Val66Met on cognitive skills in veterans with posttraumatic stress disorder. Neurosci. Lett. 2020, 735, 135235. [Google Scholar] [CrossRef]
- Pivac, N.; Kozaric-Kovacic, D. Pharmacotherapy of treatment-resistant combat related posttraumatic stress disorder with psychotic features. Croat. Med. J. 2006, 47, 440–451. [Google Scholar]
- Braakman, M.H.; Kortmann, F.A.M.; van den Brink, W. Validity of post-traumatic stress disorder with secondary psychotic features: A review of the evidence. Acta Psychiatr. Scand. 2009, 119, 15–24. [Google Scholar] [CrossRef]
- Compean, E.; Hamner, M. Posttraumatic stress disorder with secondary psychotic features (PTSD-SP): Diagnostic and treatment challenges. Prog. Neuropsychopharmacol. Biol. Psychiatry 2019, 88, 265–275. [Google Scholar] [CrossRef]
- Svob Strac, D.; Kovacic Petrovic, Z.; Nikolac Perkovic, M.; Umolac, D.; Nedic Erjavec, G.; Pivac, N. Platelet monoamine oxidase type B, MAOB intron 13 and MAOA-uVNTR polymorphism and symptoms of post-traumatic stress disorder. Stress 2016, 19, 362–373. [Google Scholar] [CrossRef]
- Acierno, R.; Kilpatrick, D.G.; Resnick, H.; Saunders, B.; De Arellano, M.; Best, C. Assault, PTSD, family substance use, and depression as risk factors for cigarette use in youth: Findings from the National Survey of Adolescents. J. Trauma Stress 2000, 13, 381–396. [Google Scholar] [CrossRef]
- Buckley, T.C.; Mozley, S.L.; Bedard, M.A.; Dewulf, A.C.; Greif, J. Preventive health behaviors, health-risk behaviors, physical morbidity, and health-related role functioning impairment in veterans with post-traumatic stress disorder. Mil. Med. 2004, 169, 536–540. [Google Scholar] [CrossRef] [Green Version]
- Hapke, U.; Schumann, A.; Rumpf, H.J.; John, U.; Konerding, U.; Meyer, C. Association of smoking and nicotine dependence with trauma and posttraumatic stress disorder in a general population sample. J. Nerv. Ment. Dis. 2005, 193, 843–846. [Google Scholar] [CrossRef]
- Khantzian, E.J. The self-medication hypothesis of substance use disorders: A reconsideration and recent applications. Harv. Rev. Psychiatry 1997, 4, 231–244. [Google Scholar] [CrossRef]
- Froeliger, B.; Crowell Beckham, J.; Feldman Dennis, M.; Victoria Kozink, R.; Joseph McClernon, F. Effects of Nicotine on Emotional Reactivity in PTSD and Non-PTSD Smokers: Results of a Pilot fMRI Study. Adv. Pharmacol. Sci. 2012, 2012, 265724. [Google Scholar] [CrossRef]
- Kearns, N.T.; Carl, E.; Stein, A.T.; Vujanovic, A.A.; Zvolensky, M.J.; Smits, J.A.J.; Powers, M.B. Posttraumatic stress disorder and cigarette smoking: A systematic review. Depress. Anxiety 2018, 35, 1056–1072. [Google Scholar] [CrossRef] [PubMed]
- Heishman, S.J.; Kleykamp, B.A.; Singleton, E.G. Meta-analysis of the acute effects of nicotine and smoking on human performance. Psychopharmacology 2010, 210, 453–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, H.; Du, X.; Yin, G.; Zhang, Y.; Li, X.; Cai, J.; Huang, X.; Ning, Y.; Soares, J.C.; Wu, F.; et al. Effects of smoking on cognition and BDNF levels in a male Chinese population: Relationship with BDNF Val66Met polymorphism. Sci. Rep. 2019, 9, 217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos, M.W.; Serebrisky, D.; Castaldelli-Maia, J.M. Smoking and Cognition. Curr. Drug Abuse Rev. 2016, 9, 76–79. [Google Scholar] [CrossRef]
- Corley, J.; Gow, A.J.; Starr, J.M.; Deary, I.J. Smoking, childhood IQ, and cognitive function in old age. J. Psychosom. Res. 2012, 73, 132–138. [Google Scholar] [CrossRef] [Green Version]
- Huang, E.J.; Reichardt, L.F. Neurotrophins: Roles in neuronal development and function. Annu. Rev. Neurosci. 2001, 24, 677–736. [Google Scholar] [CrossRef] [Green Version]
- Andreska, T.; Aufmkolk, S.; Sauer, M.; Blum, R. High abundance of BDNF within glutamatergic presynapses of cultured hippocampal neurons. Front. Cell Neurosci. 2014, 8, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Aliaga, E.; Mendoza, I.; Tapia-Arancibia, L. Distinct subcellular localization of BDNF transcripts in cultured hypothalamic neurons and modification by neuronal activation. J. Neural Transm. 2009, 116, 23–32. [Google Scholar] [CrossRef]
- Parkhurst, C.N.; Yang, G.; Ninan, I.; Savas, J.N.; Yates, J.R., 3rd; Lafaille, J.J.; Hempstead, B.L.; Littman, D.R.; Gan, W.B. Microglia promote learning-dependent synapse formation through brain-derived neurotrophic factor. Cell 2013, 155, 1596–1609. [Google Scholar] [CrossRef] [Green Version]
- Pang, P.T.; Nagappan, G.; Guo, W.; Lu, B. Extracellular and intracellular cleavages of proBDNF required at two distinct stages of late-phase LTP. NPJ Sci. Learn. 2016, 1, 16003. [Google Scholar] [CrossRef] [Green Version]
- Bergami, M.; Rimondini, R.; Santi, S.; Blum, R.; Gotz, M.; Canossa, M. Deletion of TrkB in adult progenitors alters newborn neuron integration into hippocampal circuits and increases anxiety-like behavior. Proc. Natl. Acad. Sci. USA 2008, 105, 15570–15575. [Google Scholar] [CrossRef] [Green Version]
- Kim, G.S.; Cho, S.; Nelson, J.W.; Zipfel, G.J.; Han, B.H. TrkB agonist antibody pretreatment enhances neuronal survival and long-term sensory motor function following hypoxic ischemic injury in neonatal rats. PLoS ONE 2014, 9, e88962. [Google Scholar] [CrossRef] [Green Version]
- Colino-Oliveira, M.; Rombo, D.M.; Dias, R.B.; Ribeiro, J.A.; Sebastião, A.M. BDNF-induced presynaptic facilitation of GABAergic transmission in the hippocampus of young adults is dependent of TrkB and adenosine A(2A) receptors. Purinergic Signal. 2016, 12, 283–294. [Google Scholar] [CrossRef] [Green Version]
- Lu, B.; Nagappan, G.; Guan, X.; Nathan, P.J.; Wren, P. BDNF-based synaptic repair as a disease-modifying strategy for neurodegenerative diseases. Nat. Rev. Neurosci. 2013, 14, 401–416. [Google Scholar] [CrossRef]
- Costa, C.; Sgobio, C.; Siliquini, S.; Tozzi, A.; Tantucci, M.; Ghiglieri, V.; Di Filippo, M.; Pendolino, V.; de Iure, A.; Marti, M.; et al. Mechanisms underlying the impairment of hippocampal long-term potentiation and memory in experimental Parkinson’s disease. Brain 2012, 135, 1884–1899. [Google Scholar] [CrossRef] [Green Version]
- Komulainen, P.; Pedersen, M.; Hänninen, T.; Bruunsgaard, H.; Lakka, T.A.; Kivipelto, M.; Hassinen, M.; Rauramaa, T.H.; Pedersen, B.K.; Rauramaa, R. BDNF is a novel marker of cognitive function in ageing women: The DR’s EXTRA Study. Neurobiol. Learn. Mem. 2008, 90, 596–603. [Google Scholar] [CrossRef]
- Yu, H.; Zhang, Z.; Shi, Y.; Bai, F.; Xie, C.; Qian, Y.; Yuan, Y.; Deng, L. Association study of the decreased serum BDNF concentrations in amnestic mild cognitive impairment and the Val66Met polymorphism in Chinese Han. J. Clin. Psychiatry 2008, 69, 1104–1111. [Google Scholar] [CrossRef]
- Gunstad, J.; Benitez, A.; Smith, J.; Glickman, E.; Spitznagel, M.B.; Alexander, T.; Juvancic-Heltzel, J.; Murray, L. Serum brain-derived neurotrophic factor is associated with cognitive function in healthy older adults. J. Geriatr. Psychiatry Neurol. 2008, 21, 166–170. [Google Scholar] [CrossRef]
- Ciammola, A.; Sassone, J.; Cannella, M.; Calza, S.; Poletti, B.; Frati, L.; Squitieri, F.; Silani, V. Low brain-derived neurotrophic factor (BDNF) levels in serum of Huntington’s disease patients. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2007, 144B, 574–577. [Google Scholar] [CrossRef]
- Kenny, P.J.; File, S.E.; Rattray, M. Acute nicotine decreases, and chronic nicotine increases the expression of brain-derived neurotrophic factor mRNA in rat hippocampus. Brain Res. Mol. Brain Res. 2000, 85, 234–238. [Google Scholar] [CrossRef]
- Angelucci, F.; Ricci, V.; Gelfo, F.; Martinotti, G.; Brunetti, M.; Sepede, G.; Signorelli, M.; Aguglia, E.; Pettorruso, M.; Vellante, F.; et al. BDNF serum levels in subjects developing or not post-traumatic stress disorder after trauma exposure. Brain Cogn. 2014, 84, 118–122. [Google Scholar] [CrossRef]
- Mojtabavi, H.; Saghazadeh, A.; van den Heuvel, L.; Bucker, J.; Rezaei, N. Peripheral blood levels of brain-derived neurotrophic factor in patients with post-traumatic stress disorder (PTSD): A systematic review and meta-analysis. PLoS ONE 2020, 15, e0241928. [Google Scholar] [CrossRef]
- Rosas-Vidal, L.E.; Do-Monte, F.H.; Sotres-Bayon, F.; Quirk, G.J. Hippocampal--prefrontal BDNF and memory for fear extinction. Neuropsychopharmacology 2014, 39, 2161–2169. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.K.; Wiener, J.M. PTSD recovery, spatial processing, and the val66met polymorphism. Front. Hum. Neurosci. 2014, 8, 100. [Google Scholar] [CrossRef] [Green Version]
- Miranda, M.; Morici, J.F.; Zanoni, M.B.; Bekinschtein, P. Brain-Derived Neurotrophic Factor: A Key Molecule for Memory in the Healthy and the Pathological Brain. Front. Cell Neurosci. 2019, 13, 363. [Google Scholar] [CrossRef]
- Egan, M.F.; Kojima, M.; Callicott, J.H.; Goldberg, T.E.; Kolachana, B.S.; Bertolino, A.; Zaitsev, E.; Gold, B.; Goldman, D.; Dean, M.; et al. The BDNF val66met polymorphism affects activity-dependent secretion of BDNF and human memory and hippocampal function. Cell 2003, 112, 257–269. [Google Scholar] [CrossRef] [Green Version]
- Hariri, A.R.; Goldberg, T.E.; Mattay, V.S.; Kolachana, B.S.; Callicott, J.H.; Egan, M.F.; Weinberger, D.R. Brain-derived neurotrophic factor val66met polymorphism affects human memory-related hippocampal activity and predicts memory performance. J. Neurosci. 2003, 23, 6690–6694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Benedek, D.M.; Fullerton, C.S.; Forsten, R.D.; Naifeh, J.A.; Li, X.X.; Hu, X.Z.; Li, H.; Jia, M.; Xing, G.Q.; et al. PTSD risk is associated with BDNF Val66Met and BDNF overexpression. Mol. Psychiatry 2014, 19, 8–10. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Kaminga, A.C.; Wu, X.; Wen, S.W.; Tan, H.; Yan, J.; Deng, J.; Lai, Z.; Liu, A. Brain-Derived Neurotropic Factor Val66Met Polymorphism and Posttraumatic Stress Disorder among Survivors of the 1998 Dongting Lake Flood in China. Biomed. Res. Int. 2017, 2017, 4569698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pivac, N.; Kozaric-Kovacic, D.; Grubisic-Ilic, M.; Nedic, G.; Rakos, I.; Nikolac, M.; Blazev, M.; Muck-Seler, D. The association between brain-derived neurotrophic factor Val66Met variants and psychotic symptoms in posttraumatic stress disorder. World J. Biol. Psychiatry 2012, 13, 306–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Ozbay, F.; Lappalainen, J.; Kranzler, H.R.; van Dyck, C.H.; Charney, D.S.; Price, L.H.; Southwick, S.; Yang, B.Z.; Rasmussen, A.; et al. Brain derived neurotrophic factor (BDNF) gene variants and Alzheimer’s disease, affective disorders, posttraumatic stress disorder, schizophrenia, and substance dependence. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2006, 141B, 387–393. [Google Scholar] [CrossRef] [Green Version]
- Toh, Y.L.; Ng, T.; Tan, M.; Tan, A.; Chan, A. Impact of brain-derived neurotrophic factor genetic polymorphism on cognition: A systematic review. Brain Behav. 2018, 8, e01009. [Google Scholar] [CrossRef]
- Pitts, B.L.; Whealin, J.M.; Harpaz-Rotem, I.; Duman, R.S.; Krystal, J.H.; Southwick, S.M.; Pietrzak, R.H. BDNF Val66Met polymorphism and posttraumatic stress symptoms in U.S. military veterans: Protective effect of physical exercise. Psychoneuroendocrinology 2019, 100, 198–202. [Google Scholar] [CrossRef]
- Miller, J.K.; McDougall, S.; Thomas, S.; Wiener, J. The Impact of the Brain-Derived Neurotrophic Factor Gene on Trauma and Spatial Processing. J. Clin. Med. 2017, 6, 108. [Google Scholar] [CrossRef] [Green Version]
- Hori, H.; Itoh, M.; Yoshida, F.; Lin, M.; Niwa, M.; Hakamata, Y.; Ino, K.; Imai, R.; Ogawa, S.; Matsui, M.; et al. The BDNF Val66Met polymorphism affects negative memory bias in civilian women with PTSD. Sci. Rep. 2020, 10, 3151. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Li, X.X.; Hu, X.Z. Post-traumatic stress disorder risk and brain-derived neurotrophic factor Val66Met. World J. Psychiatry 2016, 6, 1–6. [Google Scholar] [CrossRef]
- Guo, J.C.; Li, X.; Guo, M.; Gao, Y.S.; Fu, L.Q.; Jiang, X.L.; Fu, L.M.; Huang, T. Association of BDNF gene polymorphism with endophenotypes in posttraumatic stress disorder. Rev. Assoc. Med. Bras. 2020, 66, 615–622. [Google Scholar] [CrossRef]
- Guo, X.J.; Hang, W.H.; Guo, L.Y. A family based association study of BDNF C270T polymorphism with schizophrenia. J. Nerv. Ment. Dis. 2011, 37, 568–570. [Google Scholar]
- Guo, J.C.; Yang, Y.J.; Guo, M.; Wang, X.D.; Juan, Y.; Gao, Y.S.; Fu, L.Q.; Jiang, X.L.; Fu, L.M.; Huang, T. Correlations of Four Genetic Single Nucleotide Polymorphisms in Brain-Derived Neurotrophic Factor with Posttraumatic Stress Disorder. Psychiatry Investig. 2018, 15, 407–412. [Google Scholar] [CrossRef]
- Weinstock-Guttman, B.; Benedict, R.H.; Tamaño-Blanco, M.; Ramasamy, D.P.; Stosic, M.; Polito, J.; Zivadinov, R.; Ramanathan, M. The rs2030324 SNP of brain-derived neurotrophic factor (BDNF) is associated with visual cognitive processing in multiple sclerosis. Pathophysiology 2011, 18, 43–52. [Google Scholar] [CrossRef]
- Shovit, R.; Praveen, K.S. Association of Brain-Derived Neurotrophic Factor (BDNF) Gene SNPs G196A and C270T with Alzheimer’s Disease: A Meta-Analysis. J. Alzheimers Dis. Parkinsonism 2017, 7, 323. [Google Scholar] [CrossRef] [Green Version]
- Nagata, T.; Shinagawa, S.; Nukariya, K.; Ochiai, Y.; Kawamura, S.; Agawa-Ohta, M.; Kasahara, H.; Nakayama, K.; Yamada, H. Association between brain-derived neurotrophic factor (BDNF) gene polymorphisms and executive function in Japanese patients with Alzheimer’s disease. Psychogeriatrics 2011, 11, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Desai, P.; Nebes, R.; DeKosky, S.T.; Kamboh, M.I. Investigation of the effect of brain-derived neurotrophic factor (BDNF) polymorphisms on the risk of late-onset Alzheimer’s disease (AD) and quantitative measures of AD progression. Neurosci. Lett. 2005, 379, 229–234. [Google Scholar] [CrossRef]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 4th ed.; American Psychiatric Association: Washington, DC, USA, 2000. [Google Scholar]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Association: Washington, DC, USA, 2013. [Google Scholar]
- Weathers, F.W.; Keane, T.M.; Davidson, J.R. Clinician-administered PTSD scale: A review of the first ten years of research. Depress. Anxiety 2001, 13, 132–156. [Google Scholar] [CrossRef]
- Lindstrom, E.; Tuninger, E.; Hodgins, S.; Levander, S. PECC—Factor structure and findings in three longitudinal materials of patients with schizophrenia. Nord. J. Psychiatry 2012, 66, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Tudor, L.; Konjevod, M.; Nikolac Perkovic, M.; Svob Strac, D.; Nedic Erjavec, G.; Uzun, S.; Kozumplik, O.; Sagud, M.; Kovacic Petrovic, Z.; Pivac, N. Genetic Variants of the Brain-Derived Neurotrophic Factor and Metabolic Indices in Veterans with Posttraumatic Stress Disorder. Front. Psychiatry 2018, 9, 637. [Google Scholar] [CrossRef] [Green Version]
- Miller, S.A.; Dykes, D.D.; Polesky, H.F. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res. 1988, 16, 1215. [Google Scholar] [CrossRef] [Green Version]
- The 1000 Genomes Project Consortium. A global reference for human genetic variation. Nature 2015, 526, 68–74. [Google Scholar] [CrossRef] [Green Version]
- Faul, F.; Erdfelder, E.; Lang, A.G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef]
- Havelka Mestrovic, A.; Tudor, L.; Nikolac Perkovic, M.; Nedic Erjavec, G.; Kovacic Petrovic, Z.; Svob Strac, D.; Konjevod, M.; Pivac, N. Significant association between catechol-O-methyltransferase (COMT) Val158/108Met polymorphism and cognitive function in veterans with PTSD. Neurosci. Lett. 2018, 666, 38–43. [Google Scholar] [CrossRef]
- Woon, F.L.; Farrer, T.J.; Braman, C.R.; Mabey, J.K.; Hedges, D.W. A meta-analysis of the relationship between symptom severity of Posttraumatic Stress Disorder and executive function. Cogn. Neuropsychiatry 2017, 22, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Sumner, J.A.; Hagan, K.; Grodstein, F.; Roberts, A.L.; Harel, B.; Koenen, K.C. Posttraumatic stress disorder symptoms and cognitive function in a large cohort of middle-aged women. Depress. Anxiety 2017, 34, 356–366. [Google Scholar] [CrossRef]
- Bell, C.J.; Frampton, C.M.; Colhoun, H.C.; Douglas, K.M.; McIntosh, V.V.; Carter, F.A.; Jordan, J.; Carter, J.D.; Smith, R.A.; Marie, L.M.; et al. Earthquake brain: Impairment of spatial memory following long-term earthquake-related stress. Aust. N. Z. J. Psychiatry 2019, 53, 37–47. [Google Scholar] [CrossRef]
- Kozaric Kovacic, D.; Grubisic Ilic, M.; Romic, Z.; Vidovic, A.; Jendricko, T.; Pivac, N. Body mass index in male Caucasian veterans with or without posttraumatic stress disorder. Progr. Neuro-Psychopharmacol. Biol. Psychiatry 2009, 33, 1447–1450. [Google Scholar] [CrossRef]
- Sagud, M.; Petrovic, B.; Vilibic, M.; Mihaljevic-Peles, A.; Vuksan-Cusa, B.; Rados, I.; Grea, A.; Trkulja, V. The relationship among psychopathology, religiosity, and nicotine dependence in Croatian war veterans with posttraumatic stress disorder. Croat. Med. J. 2018, 59, 165–177. [Google Scholar] [CrossRef] [Green Version]
- Bountress, K.E.; Bacanu, S.A.; Tomko, R.L.; Korte, K.J.; Hicks, T.; Sheerin, C.; Lind, M.J.; Marraccini, M.; Nugent, N.; Amstadter, A.B. The Effects of a BDNF Val66Met Polymorphism on Posttraumatic Stress Disorder: A Meta-Analysis. Neuropsychobiology 2017, 76, 136–142. [Google Scholar] [CrossRef]
- Felmingham, K.L.; Zuj, D.V.; Hsu, K.C.M.; Nicholson, E.; Palmer, M.A.; Stuart, K.; Vickers, J.C.; Malhi, G.S.; Bryant, R.A. The BDNF Val66Met polymorphism moderates the relationship between Posttraumatic Stress Disorder and fear extinction learning. Psychoneuroendocrinology 2018, 91, 142–148. [Google Scholar] [CrossRef] [Green Version]
- Young, D.A.; Neylan, T.C.; O’Donovan, A.; Metzler, T.; Richards, A.; Ross, J.A.; Inslicht, S.S. The interaction of BDNF Val66Met, PTSD, and child abuse on psychophysiological reactivity and HPA axis function in a sample of Gulf War Veterans. J. Affect. Disord 2018, 235, 52–60. [Google Scholar] [CrossRef]
- Zhang, L.; Hu, X.Z.; Benedek, D.M.; Fullerton, C.S.; Forsten, R.D.; Naifeh, J.A.; Li, X.; Biomarker Study Group. Genetic predictor of current suicidal ideation in US service members deployed to Iraq and Afghanistan. J. Psychiatr. Res. 2019, 113, 65–71. [Google Scholar] [CrossRef]
- Rabl, U.; Meyer, B.M.; Diers, K.; Bartova, L.; Berger, A.; Mandorfer, D.; Popovic, A.; Scharinger, C.; Huemer, J.; Kalcher, K.; et al. Additive gene-environment effects on hippocampal structure in healthy humans. J. Neurosci. 2014, 34, 9917–9926. [Google Scholar] [CrossRef] [Green Version]
- Tsai, S.J. Critical Issues in BDNF Val66Met Genetic Studies of Neuropsychiatric Disorders. Front. Mol. Neurosci. 2018, 11, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pivac, N.; Kim, B.; Nedic, G.; Joo, Y.H.; Kozaric-Kovacic, D.; Hong, J.P.; Muck-Seler, S. Ethnic differences in the brain-derived neurotrophic factor Val66Met polymorphism in Croatian and Korean healthy subjects. Croat. Med. J. 2009, 50, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Pregelj, P.; Nedic, G.; Videtic Paska, A.; Zupanc, T.; Nikolac, M.; Balazic, J.; Tomori, M.; Komel, R.; Muck Seler, D.; Pivac, N. The association between brain-derived neurotrophic factor polymorphism (BDNF Val66Met) and suicide. J. Affect. Disord. 2011, 128, 287–290. [Google Scholar] [CrossRef] [PubMed]
- Zhai, J.; Zhao, J.; Chen, M.; Li, J.; Su, Z. Relationship between brain-derived neurotrophic factor gene C270T polymorphisms and the psychotic symptoms and cognitive functioning of patients with schizophrenia. Shanghai Arch. Psychiatry 2012, 24, 328–334. [Google Scholar] [CrossRef]
- Kunugi, H.; Ueki, A.; Otsuka, M.; Isse, K.; Hirasawa, H.; Kato, N.; Nabika, T.; Kobayashi, S.; Nanko, S. A novel polymorphism of the brain-derived neurotrophic factor (BDNF) gene associated with late-onset Alzheimer’s disease. Mol. Psychiatry 2001, 6, 83–86. [Google Scholar] [CrossRef]
- Riemenschneider, M.; Schwarz, S.; Wagenpfeil, S.; Diehl, J.; Müller, U.; Förstl, H.; Kurz, A. A polymorphism of the brain-derived neurotrophic factor (BDNF) is associated with Alzheimer’s disease in patients lacking the Apolipoprotein E epsilon4 allele. Mol. Psychiatry 2002, 7, 782–785. [Google Scholar] [CrossRef] [Green Version]
- Matsushita, S.; Arai, H.; Matsui, T.; Yuzuriha, T.; Urakami, K.; Masaki, T.; Higuchi, S. Brain-derived neurotrophic factor gene polymorphisms and Alzheimer’s disease. J. Neural Transm. 2005, 112, 703–711. [Google Scholar] [CrossRef]
- Olin, D.; MacMurray, J.; Comings, D.E. Risk of late-onset Alzheimer’s disease associated with BDNF C270T polymorphism. Neurosci. Lett. 2005, 381, 275–278. [Google Scholar] [CrossRef]
- Durmaz, A.; Kumral, E.; Durmaz, B.; Onay, H.; Aslan, G.I.; Ozkinay, F.; Pehlivan, S.; Orman, M.; Cogulu, O. Genetic factors associated with the predisposition to late onset Alzheimer’s disease. Gene 2019, 707, 212–215. [Google Scholar] [CrossRef]
- Ji, H.; Dai, D.; Wang, Y.; Jiang, D.; Zhou, X.; Lin, P.; Ji, X.; Li, J.; Zhang, Y.; Yin, H.; et al. Association of BDNF and BCHE with Alzheimer’s disease: Meta-analysis based on 56 genetic case-control studies of 12,563 cases and 12,622 controls. Exp. Ther. Med. 2015, 9, 1831–1840. [Google Scholar] [CrossRef] [Green Version]
- Tsai, S.J.; Hong, C.J.; Liu, H.C.; Liu, T.Y.; Liou, Y.J. The brain-derived neurotrophic factor gene as a possible susceptibility candidate for Alzheimer’s disease in a chinese population. Dement. Geriatr. Cogn. Disord. 2006, 21, 139–143. [Google Scholar] [CrossRef]
- Zdanys, K.F.; Kleiman, T.G.; Zhang, H.; Ozbay, F.; MacAvoy, M.G.; Gelernter, J.; van Dyck, C.H. BDNF variants, premorbid educational attainment, and disease characteristics in Alzheimer’s disease: An exploratory study. J. Alzheimers Dis. 2009, 17, 887–898. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Tian, D.; Li, J.; Chen, L.; Tang, L.; Fan, D. The Analysis of Two BDNF Polymorphisms G196A/C270T in Chinese Sporadic Amyotrophic Lateral Sclerosis. Front. Aging Neurosci. 2017, 9, 135. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Wang, W.; Jiang, Z.; Zhu, Z.; Liu, D.; Pan, F. Long-Term Effect of Post-traumatic Stress in Adolescence on Dendrite Development and H3K9me2/BDNF Expression in Male Rat Hippocampus and Prefrontal Cortex. Front. Cell Dev. Biol. 2020, 8, 682. [Google Scholar] [CrossRef]
- Kim, T.Y.; Kim, S.J.; Chung, H.G.; Choi, J.H.; Kim, S.H.; Kang, J.I. Epigenetic alterations of the BDNF gene in combat-related post-traumatic stress disorder. Acta Psychiatr. Scand. 2017, 135, 170–179. [Google Scholar] [CrossRef]
- Mentis, A.A.; Dardiotis, E.; Efthymiou, V.; Chrousos, G.P. Non-genetic risk and protective factors and biomarkers for neurological disorders: A meta-umbrella systematic review of umbrella reviews. BMC Med. 2021, 19, 6. [Google Scholar] [CrossRef]
- Smith, P.J.; Merwin, R.M. The Role of Exercise in Management of Mental Health Disorders: An Integrative Review. Annu. Rev. Med. 2021, 72, 45–62. [Google Scholar] [CrossRef]
- Ferrer, A.; Labad, J.; Salvat-Pujol, N.; Barrachina, M.; Costas, J.; Urretavizcaya, M.; de Arriba-Arnau, A.; Crespo, J.M.; Soriano-Mas, C.; Carracedo, Á.; et al. BDNF genetic variants and methylation: Effects on cognition in major depressive disorder. Transl. Psychiatry 2019, 9, 265. [Google Scholar] [CrossRef] [Green Version]
- Avgan, N.; Sutherland, H.G.; Spriggens, L.K.; Yu, C.; Ibrahim, O.; Bellis, C.; Haupt, L.M.; Shum, D.H.K.; Griffiths, L.R. BDNF Variants May Modulate Long-Term Visual Memory Performance in a Healthy Cohort. Int. J. Mol. Sci. 2017, 18, 655. [Google Scholar] [CrossRef] [Green Version]
- Pietrzak, R.H.; Goldstein, R.B.; Southwick, S.M.; Grant, B.F. Prevalence and Axis I comorbidity of full and partial posttraumatic stress disorder in the United States: Results from Wave 2 of the National Epidemiologic Survey on Alcohol and Related Conditions. J. Anxiety Disord. 2011, 25, 456–465. [Google Scholar] [CrossRef] [Green Version]
- Ryan, H.; Trosclair, A.; Gfroerer, J. Adult current smoking: Differences in definitions and prevalence estimates-NHIS and NSDUH, 2008. J. Environ. Public Health 2012, 918368. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Control Subjects | Subjects with PTSD | Statistics | ||
|---|---|---|---|---|
| Age/Median (25th; 75th) | 59 (52; 67) | 50 (41; 56) | U = 21,941.0; p < 0.001 | |
| BMI/Median (25th; 75th) | 29.1 (26.4; 30.5) | 27.8 (25.7; 30.4) | U = 16,725.0; p = 0.053 | |
| CAPS/Median (25th; 75th) | - | 81 (72; 90) | - | |
| CAPS severity/N (%) | Mild | - | 94 (14.9) | |
| Moderate | - | 443 (68.9) | ||
| Severe | - | 106 (16.5) | ||
| PANSS scores | 32 (30; 33) | 54 (48; 61) | U = 1720.5; p < 0.001 | |
| ROCF copy | 20 (20; 20) | 20 (20; 20) | U = 11,905.0; p = 0.128 | |
| ROCF immediate recall | 19 (18; 20) | 12 (9; 15) | U = 1684.0; p < 0.001 | |
| ROCF delayed recall | 17 (16; 18) | 7 (5; 9) | U = 1598.0; p < 0.001 | |
| Smoking/N (%) | Yes | 67 (55.8) | 344 (67.2) | χ2 = 4.636; df = 1; p = 0.031 |
| No | 53 (44.2) | 175 (34.2) | ||
| Number of cigarettes/Median (25th; 75th) | 10 (10; 16) | 18 (15; 22) | U = 3723.0; p < 0.001 | |
| SNP | Genotype/Allele | Control Subjects | Subjects with PTSD | Total | Statistics |
|---|---|---|---|---|---|
| BDNF rs6265 | AA | 4 (3.3) | 18 (2.8) | 22 (2.9) | χ2 = 1.379; df = 2; p = 0.502 |
| GA | 48 (39.7) | 222 (34.5) | 270 (35.3) | ||
| GG | 69 (57.0) | 403 (62.7) | 472 (61.8) | ||
| A carriers | 52 (43.0) | 240 (37.3) | 292 (38.2) | χ2 = 1.377; df = 1; p = 0.241 | |
| GG | 69 (57.0) | 403 (62.7) | 472 (61.8) | ||
| A | 56 (23.1) | 258 (20.1) | 314 (20.5) | χ2 = 1.182; df = 1; p = 0.277 | |
| G | 186 (76.9) | 1028 (79.9) | 1214 (79.5) | ||
| BDNF rs56164415 | CC | 105 (86.8) | 473 (73.6) | 578 (75.6) | χ2 = 58.335; df = 2; p < 0.001 |
| CT | 2 (1.7) | 159 (24.7) | 161 (21.1) | ||
| TT | 14 (11.6) | 11 (1.7) | 25 (3.3) | ||
| T carriers | 16 (13.3) | 170 (26.4) | 186 (24.4) | χ2 = 9.653; df = 1; p = 0.002 | |
| CC | 105 (86.8) | 473 (73.6) | 578 (75.6) | ||
| C | 212 (87.6) | 1105 (85.9) | 1317 (86.2) | χ2 = 0.482; df = 1; p = 0.488 | |
| T | 30 (12.4) | 181 (14.1) | 211 (13.8) |
| Control Subjects | Subjects with PTSD | |||
|---|---|---|---|---|
| Smokers | Non-Smokers | Smokers | Non-Smokers | |
| PANSS total scores | 32 (30; 34) | 32 (30; 33) | 57 (50; 62) | 54 (48; 61) |
| U = 1546.5; p = 0.210 | U = 25,862.0; p = 0.009 | |||
| PANSS cognitive subscale scores | 4 (4; 4) | 4 (4; 4) | 7 (5; 8) | 6 (5; 8) |
| U = 1691.5; p = 0.394 | U = 26,315.0; p = 0.018 | |||
| ROCF Copy scores | 20 (20; 20) | 20 (20; 20) | 20 (20; 20) | 20 (20; 20) |
| U = 308.0; p = 0.149 | U = 1.019.0; p = 0.844 | |||
| ROCF Immediate Recall scores | 19 (18; 19) | 19 (18; 20) | 12 (9; 15) | 13 (11; 15) |
| U = 305.5; p = 0.529 | U = 956.0; p = 0.548 | |||
| ROCF Delayed Recall scores | 17 (17; 17) | 17 (16; 18) | 7 (5; 10) | 6 (5; 9) |
| U = 282.0; p = 0.290 | U = 939.5; p = 0.464 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nedic Erjavec, G.; Nikolac Perkovic, M.; Tudor, L.; Uzun, S.; Kovacic Petrovic, Z.; Konjevod, M.; Sagud, M.; Kozumplik, O.; Svob Strac, D.; Peraica, T.; et al. Moderating Effects of BDNF Genetic Variants and Smoking on Cognition in PTSD Veterans. Biomolecules 2021, 11, 641. https://doi.org/10.3390/biom11050641
Nedic Erjavec G, Nikolac Perkovic M, Tudor L, Uzun S, Kovacic Petrovic Z, Konjevod M, Sagud M, Kozumplik O, Svob Strac D, Peraica T, et al. Moderating Effects of BDNF Genetic Variants and Smoking on Cognition in PTSD Veterans. Biomolecules. 2021; 11(5):641. https://doi.org/10.3390/biom11050641
Chicago/Turabian StyleNedic Erjavec, Gordana, Matea Nikolac Perkovic, Lucija Tudor, Suzana Uzun, Zrnka Kovacic Petrovic, Marcela Konjevod, Marina Sagud, Oliver Kozumplik, Dubravka Svob Strac, Tina Peraica, and et al. 2021. "Moderating Effects of BDNF Genetic Variants and Smoking on Cognition in PTSD Veterans" Biomolecules 11, no. 5: 641. https://doi.org/10.3390/biom11050641